


-
规律间隔成簇短回文重复序列及其相关蛋白(clustered regularly interspaced short palindromic repeats/CRISPR associated proteins,CRISPR/Cas)系统是古细菌和大多数细菌在生物进化过程中抵御病毒入侵而形成的一种适应性免疫防御系统. 1987年Nakata等[1]首次报道了它的特殊结构,crRNA(CRISPR RNA)包含间隔序列与重复序列[2],间隔序列来源于噬菌体和共轭质粒等外源基因,用于识别目标核酸,重复序列则用于结合Cas蛋白并指引其发挥作用. 随后科学家们在超过40%的细菌和90%的古生菌中发现了类似结构的序列[3],命名为CRISPR/Cas[4]. 对Cas蛋白的研究发现Cas12a和Cas13a不仅可以特异性顺式切割目标双链DNA(dsDNA)、单链DNA/RNA(ssDNA/RNA),还具有非特异性切割溶液中的游离核酸的“反式切割”[5 − 7]. 由于CRISPR/Cas系统具有特异性识别目标核酸、非特异性切割游离核酸及可编程性等优点,无需专业设备即可提供价格低廉、快速、准确的即时检测,被用来开发快速、高效、低成本和高灵敏度的分子传感器,例如SHERLOCK[8]和DETECTR[9]等,在新型冠状病毒检测中发挥了重要作用[10 − 12].
近年来,除重金属离子外,抗生素、内分泌干扰物、持久性难降解有机物等新型污染物越来越受人们关注,已成为环境分析的重要靶物. 传统上,这些非核酸靶标的检测依靠高效液相色谱(HPLC)[13]、气相色谱/质谱(GC/MS)[14]或酶联免疫吸附测定(ELISA)[15]等方法. 然而,这些方法通常耗时长、成本高,或需要昂贵的仪器和专业操作人员,不利于现场即时检测或快速筛查. 鉴于CRISPR/Cas系统的成熟可靠和可扩展性,研究人员尝试将其应用于环境/食品安全领域的非核酸靶标检测中,开发出了一系列高灵敏度、高特异性的便携式检测设备. 然而,相对于核酸检测应用,CRISPR/Cas系统在非核酸靶标的检测研究相对滞后. 这主要因为Cas的活化依赖于核酸底物,而构筑高灵敏度、高特异性和通用性的元件将非核酸靶标信息转换为核酸信息仍然面临挑战. 为此,本文综述了基于CRISPR/Cas系统的生物传感器在环境监测领域中非核酸靶标检测的应用进展,包括了重金属离子、阴离子、新污染物、农药、真菌毒素以及有害细菌等目标物,重点讨论了将非核酸靶标的信息转换为核酸信息的策略,并对不同的信号输出方式和检测能力进行分析比较(信息汇总于表1),最后对该领域研究存在的问题和可能的发展方向进行探讨.
-
重金属离子对环境的污染可直接或间接对人体造成严重危害. 传统的重金属检测依赖于大型检测仪器,成本高且耗时长,因此建立一种方便、高效、灵敏的检测方法很有必要. 借助于可特异性识别重金属离子的RNA-cleaving DNAzyme(RCD)、适配体以及变构转录因子等作为信息转换元件,CRISPR系统已被用来构建快速检测重金属离子的生物传感器. 这些方法具有很高的灵敏度、操作简便、成本低以及可扩展等优点,具有应用前景. 本节重点介绍利用以上信息转换元件和CRISPR系统构筑Pb2+[16 − 20]、Cd2+[21]、Zn2+[22]等几种重金属离子的生物传感器以及应用潜力,并对存在的问题进行探讨.
RCD是通过体外筛选技术(in vitro selection)得到的具有催化功能的DNA序列,可以在特定辅因子的作用下切割对应RNA底物. 目前,已有不少针对特定金属离子的RCD被公开报道[44 − 45]. RCD切割底物释放的片段可被CRISPR/Cas系统识别,从而使CRISPR/Cas的核酸切割活性被激活,该过程可将对金属离子的识别转换为核酸切割信号. 该策略仅需在RCD的底物一端设计可被crRNA特异性识别的DNA序列,在特定重金属离子作用下RCD将底物切割为两段;将可被crRNA识别的片段转移到CRISPR/Cas系统中,从而激活Cas蛋白切割报告分子. 上述CRISPR/Cas系统还可与第二个RCD组合,通过切割报告分子产生的片段再激活此RCD,形成多酶串联的信号放大系统,可以方便地通过荧光、比色、电化学等方式进行输出. 基于上述原理,科学家构建了基于RCD-CRISPR/Cas系统的检测Pb2+的生物传感器[16]. 首先在底物的5’端设计可被crRNA特异性识别的目标链,并在底物3’端标记生物素使其被固定在链霉亲和素磁珠上. 当Pb2+存在时,RCD将底物切割为两段,磁吸移除带有生物素的一段产物,然后将含目标链的产物转移到CISPR/Cas系统,与crRNA识别激活Cas蛋白. 最后,Cas蛋白切割标记有荧光和猝灭基团报告分子,从而产生信号. 经过优化研究,发现底物长度增加到20-nt时,切割效率最佳,检出限(LOD)达到0.053 nmol·L−1,远低于饮用水中铅的浓度限值(72 nmol·L−1). 然而,由于RCD的底物常含有rA碱基,在检测实际样品如血清样品时,存在被核酸酶降解产生较高背景信号的问题. 有研究报道在高盐条件下,寡核苷酸在纳米颗粒上密集排列成三维球形,具有很强的稳定性和抗降解能力[46 − 47]. 因此,采取将RCD的底物固定在金纳米颗粒(AuNP)形成球形核酸的方法,有助于提高其稳定性,降低背景信号. 该策略组合CRISPR/Cas12a来检测Pb2+(图1a)[17],最低检测限可以达到fmol·L−1级别,并且在实际样本如血清、空气颗粒和土壤中也有很好的检测性能. 该系统还可与芯片检测设备整合,用智能手机显示检测结果,提高现场检测的实用性.
利用荧光报告分子可以快速、准确地显示结果,但荧光修饰的报告分子价格昂贵,而且依赖荧光检测设备. 为降低成本,开发了基于CRISPR/Cas12a-RCD的比色传感器[18]. 在该平台中,将报告分子设计为MnO2NRs(氧化锰纳米棒)-DNA-MB(磁珠)复合材料,当存在目标分析物时,报告分子断裂,携带MnO2NRs的产物片段氧化TMB产生肉眼可见的颜色变化. 该系统被用于酒和食用油等样本中Pb2+的检测,回收率达到97%—104%,LOD达到0.54 nmol·L−1. 虽然其灵敏度相比荧光法有所下降,但可视化的信号输出方式使其在实验室外的检测更为便捷.
环境中重金属离子的浓度通常较低,对传感器的灵敏度的要求高. 基于CRISPR/Cas的传感器检测核酸分析物时,可将目标核酸进行预扩增处理,如滚环扩增(Rolling Circle Amplification, RCA)、链置换扩增技术(Strand Displacement Amplification, SDA)、催化发夹自组装(Catalytic Hairpin Assembly, CHA)及杂交链式反应(Hybridization Chain Reaction, HCR)等,以提升灵敏度. 这些放大策略同样也可和CRISPR/Cas整合来检测Pb2+[19]. 首先目标分析物激活RCD切割底物,随后掉落的产物底物片段启动SDA扩增,产生可被crRNA识别的DNA,进而引导Cas蛋白切割报告分子. 该平台的信号输出采用电化学设备,将报告分子ssDNA-Fc(二茂铁)在ZrO2/CeO/PAH的帮助下固定在玻璃碳电极上,根据报告分子断裂引起电流的变化来判断是否存在目标物. 该方法对Pb2+的检测限达到0.02 pmol·L−1,可检测酒、油、大米等样本中的Pb2+.
适配体(aptamer)和变构转录因子(allosteric transcription factor, aTF)也被用作CRISPR/Cas系统的信息转换元件. 研究人员在CRISPR/Cas12a中引入了Cd2+的适配体,构建了可检测Cd2+的生物传感器(图1b)[21]. 在SDA的引物P设计了Cd2+的适配体序列,当样品中没有Cd2+时,引物与模板结合后在DNA聚合酶的作用下产生dsDNA. 随后,Nt.BbvCI切口酶在引物延伸区域生成聚合酶的识别位点,启动二次延伸,同时产生大量的ssDNA被crRNA特异性识别并引导Cas蛋白切割荧光报告分子. 当存在Cd2+时,Cd2+与适配体结合,阻止引物P与模板T的结合,使SDA的效率降低,荧光信号变弱. 该方法检测Cd2+的LOD可达到60 pmol·L−1,远低于美国环保署饮用水中Cd2+的限量标准(44.5 nmol·L−1),回收率为97.59%—104.94%,具有很好的实际应用潜力. 研究人员还利用响应Zn2+的aTF作为信息转换元件构建了检测Zn2+的CRISPR/Ca13a生物传感器[22]. 目标分析物的存在使aTF与DNA的亲和力减弱,从而转录出可被crRNA特异性识别的激活剂,引导Cas蛋白酶对底物进行反式切割,产生可被检测的荧光信号. 在市政水样中,Zn2+浓度为1 μmol·L−1时可以利用手持式荧光检测设备观察到荧光信号.
总的来说,在重金属离子检测中,RCD仍然是CRISPR/Cas系统的主要信息转换元件. 目前,依赖Cu2+、Ag+、Cr3+、Uo2+以及镧系金属离子Lu3+等具有生态环境风险的重金属离子发挥作用的RCD已经被报道[48 − 50],这些进展为CRISPR/Cas系统在环境分析中的应用提供了新机遇. 然而,不少RCD对金属离子的专一性较差[44],因此仍有必要优化筛选条件,获得更具选择性的识别工具. 另外,高特异性区分Fe2+和Fe3+的RCD已经被筛选出来[51],启示如果能够筛选依赖不同价态(如Cr3+和Cr6+)或不同形态(如有机汞和无机汞)的RCD可能具有实际应用价值. 目前已报道的RCD的催化转化速率大多低于蛋白质酶(目前公认活性最高的RCD为10-23 DNAzyme,kcat ≈ 10 min−1)[44],而且实际应用中RCD的催化活性易受如pH、温度、有机溶剂等条件的影响[52]. 因此,开发能够耐受多种环境条件、催化活性更高的RCD,将有助于RCD和CRISPR/Cas系统整合的生物传感器的实际应用. 除RCD以外,能够专一性识别某些重金属离子如Zn2+、Cd2+、Pb2+、Co2+、Hg2+和Cu2+的适配体也被陆续报道[53 − 58],这为基于CRISPR的金属离子传感器的信息转换元件提供了更多选择.
-
水体中富含诸多无机阴离子,如F−、Cl−、NO2−、Br−、NO3−、PO43−、SO32−、SO42−等,当其含量超过一定阈值,可能具有潜在的生态风险. 响应氟离子(F−)的核糖开关作为转导元件与CRISPR/Cas系统整合,在大肠杆菌内构建了F−传感器(图1c)[22]. 在该系统中,RNA聚合酶的识别位点被F−核糖开关占据. 当F−与核糖开关结合时,RNA聚合酶可执行转录产生RNA,然后被crRNA特异性识别,引导Cas蛋白酶反式切割报告分子,产生荧光信号. 该系统对F−的LOD可达1 μmol·L−1. 然而,由于大肠杆菌的RNA对氟化物极为敏感,F−浓度超过100 μmol·L−1时抑制Cas的蛋白活性,从而限制了其线性范围. 为了准确的定量氟化物的浓度,研究人员将细胞内的转录过程移到体外,构建了无细胞的F−传感器,获得了更宽的线性响应范围[23]. 在该系统中,DNA模板包含启动子、间隔序列、核糖开关以及靶向crRNA的序列. 当F−存在时,F−核糖开关直接触发转录产生RNA,被crRNA特异性识别激活Cas蛋白切割报告分子,该方法使F−的检测上限提高到800 μmol·L−1,并且LOD为1.7 μmol·L−1,可检测瓶装水、自来水和湖水中的F−. 然而,除F−外,对于Br−、NO3−、SO42−等众多可溶于水的阴离子,仍缺乏特异性强、亲和力高的信息转换元件,限制了CRISPR/Cas系统在阴离子检测中的应用.
-
新污染物如持久性有机污染物、内分泌干扰物、药品和个人护理品等对于生态环境和人体健康构成潜在威胁,已经成为国际上高度关注的环境问题[59]. 目前,新污染物的检测主要依赖于可精确分析多种污染物分子的大型仪器,对于污染物类型的环境调查或非靶向监测具有重要作用. 然而,对于具有重要风险的污染物的靶向监测,可能更需要能在现场应用的快速检测工具,而这些大型仪器则难以适应.
核酸适配体是指通过指数富集的配体系统进化技术(SELEX)筛选得到的一段寡核苷酸序列. 核酸适体可折叠形成特定三维结构,以高亲和力和特异性结合靶标,其靶标范围涵盖金属离子、小分子、蛋白质到细胞等多种类型[60]. 相较于抗体,适配体易合成、易修饰、稳定性强、靶分子范围广,因此在环境分析领域也有诸多应用[61 − 62]. 理论上,适配体与靶分子结合引起的构象变化[63 − 65],可产生被CRISPR/Cas系统识别的核酸激活剂. 得益于筛选技术的快速发展,研究人员已经获得了不少对新污染物类分子具有高亲和力和选择性的适配体,使得适配体作为CRISPR/Cas系统检测这些化合物的信息转换元件具有应用潜力[60,66]. 此外,小分子的抗体也可作为CRISPR/Cas系统的信息转换元件,研究人员通常将抗体与可被crRNA识别的DNA以生物素和链霉亲和素连接,进而通过抗体-抗原的特异性识别将非核酸靶转换为核酸信号.
-
内分泌干扰物也被称为环境激素,是一大类具有内分泌干扰毒性的物质. 常见的内分泌干扰物有在塑料中大量使用的双酚A(BPA)、邻苯二甲酸酯类化合物以及天然和人工合成的类固醇激素等化合物. 这些物质在自然环境和人类生活中无处不在,对动物和人类的生殖、发育等产生持久性危害,导致多种与内分泌相关的疾病,严重威胁生态系统和人类健康. 雌二醇(E2)和BPA是广受关注的内分泌干扰物,二者的适配体作为信息转换元件被整合进CRISPR/Cas系统,构建了生物传感器[24]. 作者首先通过适配体链将DNA激活剂锁住,当目标分析物存在时,与适配体结合引起其构象变化,释放激活剂DNA片段,从而被crRNA识别激活Cas蛋白. 该方法对E2和BPA检测限分别为0.08 nmol·L−1和0.06 nmol·L−1,并且在污水和尿液中仍具有很好的分辨能力. 为了拓宽E2检测的应用场景,可视化的信号输出方式被引入适配体-CRISPR/Cas12a的生物传感器[25]. 该策略通过适配体将非核酸靶标转化为核酸信号,并通过CHA来放大信号,扩增产物被crRNA识别并引导Cas蛋白将DNA水凝胶包被的金属有机框架(MOF)裂解,催化TMB与 H2O2反应产生颜色变化(图2a). 该系统采用上转换材料替代荧光团,有效地提高了信噪比和荧光稳定性,并通过CHA将信号进一步放大,提高了检测的灵敏度. 并通过牛奶、猪肉等样品验证了其实际应用能力,与荧光法相比,该方法的灵敏度略低(LOD = 0.015 ng·mL−1).
与传统的信号输出方式相比,侧向层析测定(Lateral flow assay, LFA)因其操作简便、分析时间短、价格低廉备受关注. Li团队[26]将LFA用于信号输出,开发了一种检测E2的CRISPR/Cas-适配体生物传感器. 该平台主要由三部分组成(图2b):报告分子为Biotin-ssDNA-抗地高辛抗体复合物,LFA的测试区涂抹链霉亲和素,控制区涂抹抗小鼠IgG抗体,以及用于信号显示的地高辛抗体-Au@BDT@Au复合物. E2与适配体结合可使Cas蛋白失去活性,不切割报告分子;测试区的链霉亲和素捕获Biotin标记的抗地高辛抗体以及用于信号输出的地高辛抗体-Au@BDT@Au,产生信号,剩余样品流至控制区. 相反,不存在E2时,适配体序列被crRNA特异性识别,引导Cas蛋白切割报告分子,导致地高辛抗体-Au@BDT@Au不能被捕获在测试区,因而测试区域信号降低或没有信号. 该方法结合了CRISPR/Cas12a的信号放大能力和间隙增强拉曼标签(Au@BDT@Au)的高灵敏度,较传统的LFA灵敏度有很大提升(LOD = 180 fmol·L−1),可用于实际样品包括血清、尿液、牛奶和自来水中的E2检测.
-
抗生素广泛用于人类医学和畜牧业,然而抗生素的滥用及其在水环境和土壤中的累积间接地危害人类健康,在临床中常用的抗生素包括磺胺、喹诺酮、β内酰胺、四环素、大环内酯、氨基糖苷等六大类. 目前,已报道了基于CRISPR系统开发的卡那霉素[27 − 29]、妥布霉素[30]、四环素[31]、氨苄青霉素[32 − 33]等的生物传感器,在抗生素的快速现场检测中具有应用前景. 基于适配体-CRISPR/Cas检测非核酸靶的策略大多通过一条适配体链将激活链锁住,这种方法可能会产生较高的背景值. 为降低背景信号,研究人员使用两段卡那霉素的适配体序列将激活剂锁定[27]. 该系统的报告分子铥(Tm)-Rep-DNA-Bio(生物素)被裂解后,携带生物素的部分被链霉亲和素磁珠分离,带有Tm-Rep的一段可以通过同位素法监测. 该方法可以在30 min内检测卡那霉素,LOD低至4.06 pmol·L−1. 该方法使用镧系元素Tm,虽然在生物和环境中的干扰极低,但应用成本高,对仪器设备要求高. 为此,研究人员开发了成本更低、灵敏度更高的适配体-CRISPR/Cas12a系统[28]. 该系统仍采用适配体与DNA链互锁的方式,通过加入靶分子诱导适配体构象变化,使被其锁住的DNA链释放,启动HCR,生成被crRNA特异性识别的dsDNA,引导Cas蛋白切割报告分子. 报告分子ssDNA两端分别标记磁珠和葡萄糖转化酶,被切断后,分离带有转化酶的片段,催化蔗糖水解成葡萄糖,最后用便携式血糖仪定量葡萄糖来评估卡那霉素的浓度(图2c). 该方法对卡那霉素的LOD达到了1 pmol·L−1. 此外,适配体-CRISPR/Cas系统的信号也可被LFA输出[29],检测卡那霉素的LOD达到了14.8 nmol·L−1,该系统用于牛奶中卡那霉素检测的效果与市售试纸条相当.
Li等[30]在CRISPR/Cas系统中引入适配体构建了两种用于妥布霉素检测的生物传感器sensor-ss和sensor-ds. 在sensor-ss中,采用ssDNA作为激活剂并经过SDA扩增以提高传感器的灵敏度. 当目标分析物存在时,适配体结构发生变化使得激活剂被释放并启动SDA,扩增产物被crRNA识别引导Cas蛋白切割荧光报告分子(图2d). 而sensor-ds的激活剂则是使用含有原间隔序列邻近基序位点(PAM)的dsDNA,未采用核酸扩增技术放大信号. 结果显示sensor-ss(LOD = 1.542 pmol·L−1)比sensor-ds(LOD = 3.719 pmol·L−1)具有更高的灵敏度,但由于经过扩增导致反应过快达到检测平台其检测线性范围较窄(前者5—30 pmol·L−1,后者10—300 pmol·L−1).
-
持久性有机污染物的种类很多,具有环境持久性、生物累积性、远距离迁移性和高毒性等特点. 此类污染物分子苯环多,如多环芳烃、多氯联苯、多溴联苯醚等,分子结构相似且水溶性很差,筛选核酸适体或制备抗体的难度大,限制了CRISPR系统的应用. 目前开发的基于CRISPR/Cas的工具仅用于少数分子的检测. 研究人员利用抗体-抗原的特异性识别实现信息转换,与CRISPR/Cas系统整合(称为“iPOCT”)用于苯并[a]芘(Benzo (a) pyrene, BaP)的检测[34]. 该检测体系包括:固定在微孔板由小分子偶联牛血清白蛋白(BSA)构建的抗原,生物素化的抗体,链霉亲和素标记的dsDNA—用来结合crRNA并激活Cas蛋白,标记荧光和淬灭基团的ssDNA报告分子(图2e). 当待测样品中没有目标分子时,固定化的抗原与生物素化的抗体结合,然后捕获链霉亲和素标记的dsDNA;洗去未结合的dsDNA,再向微孔板内加入CRISPR/Cas系统,保留在孔内的dsDNA结合crRNA,激活Cas蛋白切割报告分子产生荧光增强的信号. 待测样品中含有目标分子,则游离的待测分子与生物素化的抗体结合,导致抗体与固定化的抗原的结合受到抑制,从而使微孔捕获的dsDNA含量减少,进而减少了CRISPR/Cas切割底物的荧光增强信号. 除了BaP,该策略也成功用于其他小分子化合物的检测,说明该策略具有一定的通用性,而且还可通过酶标仪进行高通量检测多个目标化合物. 优化后的传感器对目标分子的LOD达到4.971 fg·mL−1,在湖水等实际样品中也有很好的检测能力. 然而,该策略检测目标分子时输出的信号低于非目标物对照的信号,属于信号减小的响应模式. 此外,基于CRISPR生物传感器也被用来检测与持久性有机污染物类似的对羟基苯甲酸分子 [35].
虽然基于CRISPR系统的生物传感器在新污染物检测中展示了应用前景,但受限于信息转换元件的可及性,此类生物传感器尚未得到普遍应用. 以抗体作为靶分子识别与信息转换元件的CRISPR系统在新污染物检测中的应用较少,主要在于新污染物大多为分子量小于1000Da的小分子,没有免疫原性,很难制备相应的抗体. 适配体因其广泛的分子识别能力,作为CRISPR的信息转换元件更有优势. 目前已有一些新污染物分子,如全氟辛酸(PFOA)[67]、多氯联苯(PCB77)[68]、邻苯二甲酸酯(PAEs)[69]、四环素类、氨基糖苷类、β-内酰胺类、大环内酯类、氯霉素类、喹诺酮类等抗生素[70]的适配体被筛选出,为基于CRISPR的检测技术开发奠定了基础. 然而,新污染物的种类很多,很多分子具有相似的结构且水溶性差,缺少活性位点,使得通过靶分子固定的方法筛选适配体挑战性很大,所得适配体对结构非常相似的分子的选择性不高. 以上因素制约了适配体作为CRISPR的信息转换元件在新污染物检测中的实际应用. 发展新的筛选技术,获得亲和力更高、选择性更好、能够适应实际检测环境的适配体,将有助于推动CRISPR系统在新污染物检测中的实际应用.
-
虽然RCD是基于CRISPR/Cas系统的生物传感器的理想信号转换元件,但目前尚未有专门响应某一农药分子的RCD被开发出来,限制了该元件在农药分子检测的直接使用. 为此,研究人员设计了乙酰胆碱酯酶-RCD-CRISPR/Cas三酶级联的策略来检测有机磷农药(图3a)[36]. 首先,有机磷农药的加入使乙酰胆碱酯酶水解乙酰硫代胆碱产生硫代胆碱,硫代胆碱将MnO2纳米片还原成Mn2+,激活依赖Mn2+的RCD将底物切断释放激活剂,最后激活剂被crRNA特异性识别并引导Cas蛋白切割报告分子,产生荧光信号. 该策略成功检测了实际样品柑橘类水果、卷心菜中的对氧磷、敌敌畏和内吸磷的3种农药,LOD分别达到了270、406、218 pg·mL−1,远低于农药使用标准中的最低残留量. 然而,有机磷农药对乙酰胆碱酯酶的活性影响没有特异性,应用该方法不能区分具体的农药分子,在实际应用中仅可用于该类农药的初筛. 为实现特异性检测某一农药成分,研究人员将适配体引入CRISPR/Cas系统构建了检测啶虫脒的传感器[37]. 首先将啶虫脒适配体组装成球形核酸,将对啶虫脒的识别信号转换为核酸信号,进而激活Cas蛋白的活性,可在生菜中检测该农药残留检测(LOD = 2.7 pmol·L−1),为农药残留监测提供了更加精准、便捷的方法.
-
真菌毒素是真菌在农作物、食品中产生的次级代谢产物,可在人体中累积对人体健康造成危害. 常见的真菌毒素有黄曲毒素类、赭曲毒素A及镰孢菌毒素类,此类毒素具有很强的毒性以及致癌性,是环境监测真菌毒素中的重点关注对象. 利用抗体作为CRISPR/Cas系统的信号转换元件的策略被用来开发脱氧雪腐镰刀菌烯醇(deoxynivalenol, DON)的传感器[38]. 作者首先制作了DON抗体用以捕获目标分析物,其次通过ssDNA将PS微球与磁性纳米颗粒(MNPs)连接构建信号探针,最后是将DON通过牛血清白蛋白(BSA)与激活crRNA的DNA链偶联. DON的存在会使抗体捕获DON-激活剂,从而被crRNA特异性识别引导Cas蛋白切割报告分子PS-ssDNA-MNP,然后通过磁吸将PS微球分离出去,通过粒子计数器测量PS微球的数量用于DON的定量分析. 该方法对DON的LOD 达到了0.061 ng·mL−1,与传统的ELISA和HPLC相比表现出更宽的线性范围和更高的灵敏度. 此策略也用于黄曲霉毒素B1(AFB1)检测[34]. 然而,这些毒素的分子量小于
1000 kD,没有免疫原性,通常需要将它们制成半抗原偶联到蛋白质分子如BSA上,再进行免疫反应制备抗体,不仅流程繁琐而且难以保证选择性;另外抗体稳定性较差,保存困难. 适配体被称为化学抗体,针对小分子的核酸适体可通过SELEX技术筛选得到. 目前科学家已经分离了不少特异性结合AFB1的适配体,其结构也得到了详细表征[71],为开发基于适配体的AFB1检测工具打下坚实基础. 研究人员将AFB1的适配体与CRISPR/Cas系统联用构建检测AFBI的平台(图3b)[39],利用crRNA锁住适配体,当靶标存在时引起适配体构象变化,抑制其与crRNA的结合,降低Cas蛋白的切割能力. 该方法在酒、牛奶样品中实现了AFB1的高灵敏度检测(LOD = 0.8 ng·mL−1). 该系统的关键在于crRNA要与适配体-靶标结合的区域互补配对,这样靶标的加入才会对crRNA和适配体杂交产生很大影响,为此作者设计了28种crRNA用来优化与适配体杂交的位点. 该研究提示如果适配体与靶分子相互作用的机制明确,将对设计适配体-CRRISPR/Cas的分子传感器很有帮助.鉴于单一信号输出方式常常限制了检测范围或LOD,因此提出了基于CRISPR/Cas-适配体的双信号生物传感器[40]. 首先将报告分子ssDNA-Fc固定在金电极表面,ssDNA-Fc通过碱基互补将适配体固定,而适配体通过与激活剂1的结合互相锁定. 加入胶霉毒素时,诱导适配体构象变化,释放激活剂1,使其与H4ETTC-ssDNA结合产生聚集诱导发光;H4ETTC-ssDNA锁定的激活剂2则被释放,与crRNA识别,引导Cas切割ssDNA-Fc产生电化学信号(图3c). 传感器通过电化学、荧光双信号输出,具有很高的灵敏度(LOD = 2.4 fmol·L−1),作者应用该系统对苹果、玉米等食物样品中胶霉毒素残留进行检测. 借鉴上转换材料相较于荧光探针的优势,有研究人员利用其构建了CRISPR生物传感器用于赭曲霉毒素的监测[41].
农药和真菌毒素的残留监测对于环境/食品安全具有重要意义,目前科学家已开发出了多个针对农药和真菌毒素的适配体[72 − 73],这为基于CRISPR/Cas检测农药、真菌毒素研究提供了强有力的支持. 然而,利用适配体作为信息转换元件常常需要靶分子诱导其发生构象变化,因此需要研究人员更系统地研究适配体与靶分子的相互作用机制,以提高分子设计的成功率.
-
有害细菌的快速检测也是环境监测和食品安全检测的重要任务. 与细菌培养、抗体分析和核酸分析相比,CRISPR/Cas系统为病原菌的灵敏检测提供了一种简单便捷的方法. 研究人员将适配体引入CRISPR/Cas系统,并结合HCR设计了一种检测鼠伤寒沙门氏菌的生物传感器[42](图4a). 当靶标存在时,竞争结合适配体以启动HCR,HCR的产物可以被crRNA特异性识别并引导Cas蛋白切割报告分子. 通过HCR扩增,显著增强了反式切割活性,从而实现了灵敏检测(LOD = 20 CFU·mL−1),并可用于牛奶样品中的鼠伤寒沙门氏菌检测,提高了在临床诊断的应用潜力.
目前基于CRISPR/Cas的检测方法都使用线性crRNA(LcrRNA)作为引导RNA. 然而,在自然界中存在一种共价闭合环状RNA,可以通过化学或酶连接的方法制备,目前很少被用于CRISPR生物传感系统. 鉴于此,Wu等[43]将crRNA设计成环状引导RNA的形式(CcrRNA),证实了环化会使crRNA失去引导作用,无法激活正常的Cas蛋白的切割反应. 随后,研究团队使用大肠杆菌依赖性DNAzyme和肺炎克雷伯菌依赖的RCD作为信息转换元件,用于识别大肠杆菌和肺炎球菌. 当目标分析物存在时,DNAzyme活性被激活,切割CcrRNA形成LcrRNA(如图4b). 这样就使得crRNA获得引导作用,引导Cas蛋白切割报告分子. 经过优化,使得大肠杆菌和肺炎球菌的LOD达到了102 CFU·mL−1,并使用该系统对尿路感染的患者进行诊断,灵敏度和特异性分别达到100%、90%. 该方法将环化RNA设计进CRISPR-Cas系统,使CRISPR Cas系统摆脱线性crRNA的束缚,极大降低了背景值的同时,还灵活应用了细菌特异性的RCD(也称为适配体酶)切割RNA的特性. 此外利用RCD切割环化crRNA可以在同一管内实现信号转换和Cas蛋白的激活,避免了繁琐的实验步骤. 这也是首次通过控制crRNA的结构变化来调节CRISPR反应活性,为后续开发RCD耦联CRISPR/Cas系统用于非核酸靶标检测提供了新思路.
-
基于CRISPR/Cas的生物传感系统具有高特异性、可编程性及高催化活性,在快速、灵敏、特异的核酸检测中极具应用前景. 然而,在非核酸靶标的研究方面,该系统尚处于起步阶段. 由于Cas蛋白的活化依赖于核酸底物,因此将非核酸识别信息转换为核酸信息是其应用的关键. 目前的信息转换策略中,以DNAzyme和适配体作为信息转换元件较为普遍. 这些策略在非核酸靶标的检测上取得了良好的开端,但其进一步应用仍面临着诸多挑战.
基于CRISPR/Cas系统的非核酸靶标检测依赖于信号转导元件. 虽然已有研究通过逻辑门的方法实现了多目标分析物的检测,然而受信息转换元件的限制,其应用范围有限,尚未见类似SHERLOCK[8]这样通用的平台被开发出来. 近期科学家开发了一种将小型化和自组装的微阵列系统与基于CRISPR的特异性核酸检测相结合的方法[74],建立了一种可扩展的多重检测平台,这为基于CRISPR/Cas系统的非核酸靶标的高通量检测提供了思路.
相对于环境监测的目标物而言,已开发并实际应用的转换元件仍然很少. 而且,现有的转换元件,如针对金属离子的DNAzyme,针对小分子的适配体,还存在特异性不够强的问题. 因此筛选更多、特异性更好的分子识别工具,是未来对环境分析样品实现即时现场快速检测的重要内容. 再者,适配体用于分子识别一般不能直接输出信号,大多经过复杂的分子设计实现构象转换等来释放信号,除了设计非常复杂,还可能导致亲和力下降. 这些因素制约了适配体与CRISPR/Cas系统的整合应用. Li课题组[75 − 76]以及Liu课题组[77]筛选得到的特异性识别有害细菌的DNAzyme,在进行分子识别的同时输出信号,是典型的智能分子. 这种系统可以方便地与CRISPR/Cas整合,这对推动CRISPR/Cas系统在环境分析中的应用有重要价值.
在环境分析中,目标分析物通常以较低浓度存在,这对传感器的灵敏度提出了挑战. 基于电化学法的信号输出具有很高的灵敏度,然而其抗干扰能力可能较差,应用于复杂环境中的分析物监测存在挑战. 开发抗污的电化学分析材料或设备,对于应用该系统进行环境分析有重要价值.
CRISPR/Cas系统的大多数识别或信息转换元件都涉及核酸,这些分子多在水溶液中开发. 然而,实际的环境分析对象,如新污染物,大多难溶于水,需要有机溶剂提取和溶解,因此开发适用于含有机溶剂的体系的分子识别/信息转换工具箱具有应用价值. 我们团队首先在含35% DMSO的缓冲溶液体系中筛选鉴定了一种高度依赖该条件的DNAzyme[78],并构建了一种能在35% DMSO体系中对靶分子进行识别并产生核酸切割信号的适配体酶系统,证明了可以在含高浓度有机溶剂的环境中筛选出高活性的DNAzyme. 为开发难溶于水且需要有机溶剂溶解的有害物质的快速检测工具提供了新思路,也提示有必要在真实的环境样品或应用场景中筛选出具有专一性强的DNAzyme、适配体或具有定向进化适应能力强的CRISPR/Cas系统,以提升生物传感器在环境分析的实际应用价值.
基于CRISPR/Cas系统的生物传感器在环境分析中的应用进展
Advances in CRISPR/Cas-based biosensors for environmental analysis applications
-
摘要: CRISPR/Cas可改造为靶核酸刺激-响应的传感系统,可视为集“分子识别与信号输出”一体的生物传感器,目前在核酸即时诊断中已得到广泛应用. 鉴于该系统具有成熟、可靠和可扩展的优点,研究人员尝试将其用于环境分析领域的非核酸分析物检测,已开发了一系列具有应用前景的便携式设备. 然而,如何将非核酸靶标信息转换为核酸信息仍存在挑战,这限制了CRISPR/Cas系统在环境分析中的应用推广. 为此,本文综述了CRISPR/Cas系统在重金属离子和阴离子、新污染物、农药与真菌毒素以及有害细菌等非核酸靶标检测中的应用进展,重点介绍了构筑非核酸靶标信息转换元件的策略,以期为推进该系统在环境分析中的应用提供支持.
-
关键词:
- CRISPR/Cas /
- 新污染物 /
- 核酸适体 /
- RNA-cleaving DNAzyme /
- 生物传感器.
Abstract: The CRISPR/Cas can be engineered as a stimuli-responsive sensing system for the target nucleic acids, which enables it to serve as a “two-in-one” biosensor with both molecular recognition element and signal output element. Up to date, this type of biosensing systems have been widely employed in the Point of Care Testing (POCT) for nucleic acids. Taking advantage of being mature, reliable and scalable, the CRISPR/Cas system has been developed as portable devices for the identification of non-nucleic acid analytes in the field of environmental analytical chemistry. However, challenges remain to be resolved in converting the information from non-nucleic acid targets to nucleic acids, and thus limiting its utilization in environmental analysis. Here, we review the applications of the CRISPR/Cas system in the detection of non-nucleic acid targets, including heavy metal ions and anions, emerging contaminants, pesticides, mycotoxins, and harmful bacteria. Moreover, we focus on the strategies of constructing non-nucleic acid target transducers in order to promote the utilization of this system in environmental analysis.-
Key words:
- CRISPR/Cas /
- emerging contaminants /
- aptamer /
- RNA-cleaving DNAzyme /
- biosensor.
-
镉(Cd)是水产品中常见的重金属污染物,可以在水环境以及水生动物之间进行迁移转化,并沿着食物链在生物体内蓄积[1 − 2]. 前人研究发现,某些甲壳类水生动物对镉具有明显的蓄积特异性. 其中,三疣梭子蟹(Portunus trituberculatus)的镉蓄积问题尤为突出,引起了人们广泛关注.
三疣梭子蟹是一种生活在水深10—30 m砂质泥或砂质海底的杂食性螃蟹,广泛分布于太平洋的西海岸,北起日本的北海道,南至东南亚的越南、泰国等地[3]. 三疣梭子蟹肉质鲜美,风味独特,是一种深受中国人喜爱的具有较高营养和经济价值的海产品. 《2023中国渔业统计年鉴》[4]指出,2022年,我国海捕梭子蟹的产量就达到45.82万吨,占海捕蟹的70.75%. 然而,膳食摄入是人类暴露镉的主要途径[5],食物中的镉进入人体后会对肝脏、肾脏、骨骼、大脑等部位造成损伤,引起人体慢性中毒,进而产生生殖系统损伤、死亡率升高、预期寿命减少等负面影响[6]. 因此,三疣梭子蟹的食用安全风险问题同样受到广泛关注.
此外,相关的毒理学研究指出,不同形态的镉可能具有不同的生物学毒性[7]. 因此,在进行三疣梭子蟹的食用健康风险评价时,需考虑不同镉形态的毒性影响. 三疣梭子蟹中镉的赋存形态可以分为无机镉形态和有机镉形态[8]. 无机离子态镉可以通过不同的途径对生物体的某一器官和组织产生多方面的危害,如与体内的生物分子,包括酶和核酸等生物大分子相互作用[9]. 有机镉的具体形态目前尚未明确,并且关于有机镉的毒性机理研究也少有报道. 林建云等[10]实验指出有机结合态镉对水产动物的毒理效应和代谢作用与离子态镉(Cd2+)存在着明显的差异. 目前,国内外主要以总镉含量来评价三疣梭子蟹中镉的污染水平及食用健康风险,鲜少有针对镉形态的深入研究. 这使得三疣梭子蟹中镉的安全性评价缺乏不同镉形态的可靠数据与科学依据.
迄今为止,大部分研究主要围绕三疣梭子蟹中的镉蓄积水平、地区差异等问题进行,也有部分科学家开展了三疣梭子蟹中镉的蓄积机制及赋存形态问题的初步研究,但尚未有关于此问题的清晰明确的定论. 基于此,围绕三疣梭子蟹对镉的特异性蓄积问题,本文对三疣梭子蟹中镉的蓄积含量、分布特征及污染现状进行了综述,同时对三疣梭子蟹不同可食用组织中镉的赋存形态和健康风险评估的研究进展进行了总结分析,期望能为后续开展有关三疣梭子蟹中镉蓄积机制及赋存形态的研究提供科学参考与基础指导.
1. 三疣梭子蟹中镉的来源、污染现状及国内外限量标准(Cadmium in Portunus trituberculatus: sources, pollution situation ,domestic and international limit standards)
1.1 三疣梭子蟹中镉来源的研究进展
据报道[11 − 12],水生动物中的重金属来源主要分为两种,一种是表皮细胞或鳃等从周围水环境中直接吸收重金属,然后积累在外皮硬组织中;另一种来源则是含有重金属的颗粒状物质(饵料和沉积物颗粒)被水生动物摄食,通过食物链传递,积累在内脏软组织中. 对于甲壳类水生动物,研究人员认为重金属主要通过4个途径进入其体内. 第一,在甲壳类进行鳃呼吸时,重金属通过呼吸作用进入动物体内. 第二,某些甲壳类饵料中的金属离子通过摄食途径进入动物体内;第三,部分甲壳类动物的体表可通过与水体的渗透交换作用富集重金属;第四,水生动植物中富集的重金属通过食物链进入甲壳类体内[13].
三疣梭子蟹作为典型的甲壳类动物,目前关于其体内的镉来源的研究较少,并且对于三疣梭子蟹各可食组织中蓄积的镉具体来源也未见明确报道. 根据其他蟹类的相关研究[14 − 18],可以推测三疣梭子蟹中镉可能来源于两种途径:①环境途径,三疣梭子蟹的蟹壳和蟹腮与含镉介质(如水体、沉积物)直接接触,通过自由/被动扩散或Ca2+通道截留环境介质中的镉. ②摄食途径,三疣梭子蟹通过食物链从饵料或浮游动植物中富集镉. 作为研究三疣梭子蟹镉蓄积机制的重要组成部分,镉来源的研究还需要结合相关毒理学和生物学方法开展深入实验,以查明镉的不同来源对三疣梭子蟹体内蓄积机制的影响,同时有利于后续开展镉在三疣梭子蟹体内迁移、转运和归趋的相关研究.
1.2 三疣梭子蟹中镉的污染现状
三疣梭子蟹体内可检测出多种金属元素,镉是其中重要的有毒污染物. 尤炬炬等[19]对浙江沿海68个梭子蟹样品进行了铅、镉、总汞和无机砷的检测,发现镉含量远远高于其他重金属. 吴烨飞等[20]检测了梭子蟹中铅、镉、总汞和无机砷,只有镉存在污染问题. 可见,三疣梭子蟹中存在着明显的镉污染状况.
根据《食品安全国家标准 食品中污染物限量(GB2762—2012/GB2762—2017)》,甲壳类水产品中的镉含量不应超过0.5 mg·kg−1. 但在2012—2022年有关水产品重金属检测的报道中,全国各地市售的三疣梭子蟹普遍存在着镉超标问题(表1). 如高志杰等[21]采集了宁波市售的具有代表性的海产品进行重金属检测,发现其中镉含量最高的是梭子蟹6.808 mg·kg−1,超标率达74.5%,存在严重的镉污染现象. 樊伟等[22]研究了2007—2014年绍兴地区7类992份水产品的重金属污染情况,发现梭子蟹的镉污染最为严重,超标率为38.38%. 庞雨樵等[23]对新疆地区市售的6类水产品进行抽检,发现超标样品均为梭子蟹,最大镉浓度为2.1 mg·kg−1. 然而,即使三疣梭子蟹中镉超标问题严重,仍不能仅凭限量判定镉对三疣梭子蟹食用者的毒性大小和风险高低,还需要结合更科学的方法去研究和评价三疣梭子蟹中镉对人类的危害.
表 1 相关文献报道的三疣梭子蟹中镉含量水平及超标率Table 1. The cadmium concentrations and exceedance rates in Portunus trituberculatus reported in relevant literature采样地点Sampling region 采样年份Sampling year 报道年份Year of reporting 平均含量/(mg·kg−1)Average content 超标率/%Exceedance rate 参考文献Reference 宁波Ningbo 2012 2014 6. 808 74.5 [21] 绍兴Shaoxing 2007—2014 2016 0.90±1.35 38.38 [22] 廊坊Langfang — 2016 1.06 73.1 [24] 青岛Qingdao — 2017 2.79 100 [8] 渤海Bohai Sea 2014 2018 1.60 — [25] 温州Wenzhou 2013-2017 2020 1.99 88.4 [26] 山东Shandong — 2020 2.26 — [27] 舟山Zhoushan 2016 2021 1.44 75.6 [28] 莱州Laizhou — 2021 0.36 18.6 [29] 上海Shanghai 2021 2022 2.635 ±1.784 — [30] 注:“—”代表引用文献中未提供该信息. Note:‘—’represents in formation is not provided in the cited references. | Show Table DownLoad:
CSV
DownLoad:
CSV
1.3 国内外对三疣梭子蟹的限量
三疣梭子蟹对镉具有显著的蓄积特异性[31],其体内的镉含量往往远高于其他甲壳类水产品[32 − 33]. 可能是因为口虾姑[34]和三疣梭子蟹等海水蟹[35]具有不同于其他甲壳类水产品对镉的强蓄积特异性,在我国新实行的国标GB2762—2022《食品安全国家标准 食品中污染物限量》中,就将海蟹和虾蛄中的镉限量指标单独设立,把镉的限量值由原来的0.5 mg·kg−1调整为3 mg·kg−1. 除此之外,对限量值的改动,还可能是因为镉在不同生物体内的不同的赋存形态具有不同的生物利用率和生物学毒性. 依据我国相关检测标准,对水产品进行检测得到的镉总量是以毒性最高的无机离子态镉为限量值的评价标准. 但已有研究指出,三疣梭子蟹和虾蛄中的镉大部分以毒性较低的有机形态存在[8]. 因此,仅以0.5 mg·kg−1作为镉的限量标准,可能会高估三疣梭子蟹等海蟹和虾蛄中镉的毒性. 在港澳台地区,三疣梭子蟹中镉的限量标准有所不同. 澳门规定了去除内脏后的甲壳类镉的最高限量为2 mg·kg−1. 香港则是明确要求去除蟹壳和鳃后的整体(包括性腺、肝及其他消化器官)中镉的限量为2 mg·kg−1. 台湾地区仅规定甲壳类可食用组织限量值不超过0.5 mg·kg−1.
对于三疣梭子蟹中镉的限量值,国际上并没有统一的限量标准. 查阅相关法案文件(表2),可以发现不同国家相关的镉限量值有所不同,最低可至0.5 mg·kg−1,最高可至5.0 mg·kg−1,相差可至10倍. 不同国家的甲壳类水产品中镉含量限定对象的范围也有所不同. 比如,韩国、欧盟等国家会针对甲壳类不同组织进行限量值的规定. 对于各国镉限量差异问题,原因在于各国在划定限量值时没有统一的科学参考依据. 因此,有必要完善甲壳类中镉限量的判断指标. 不仅要根据甲壳类自身特征与生长环境进行评估,还要继续研究不同镉形态的毒理学效应,根据甲壳类中无机镉与总镉的占比情况,更细致地划分限量对象与限量值,更好地保障公众的食品安全.
表 2 部分组织或国家有关三疣梭子蟹中镉的限量值比较Table 2. The comparison of limit values for cadmium in Portunus trituberculatus in some organizations or countries组织或国家Organizations or countries 执行对象Implementation aims 限量值/(mg·kg−1)Limited value 执行标准/法规Implementing standards/regulations 中国China 甲壳类(海蟹、虾蛄除外)Crustaceans(except sea crabs and mantis shrimps) 0.5 GB2762—2022[36] 海蟹、虾蛄Sea crabs and mantis shrimps 3.0 中国澳门Macau, China 甲壳类(去除内脏)Crustaceans(Removal of internal organs) 2.0 第23/2018号行政法规食品中重金属污染物最高限量[37]Administrative Regulation No. 23/2018Maximum limits for heavy metal contaminants in food[37] 中国香港Hong Kong, China 蟹(去除壳和鳃后的整体)Crab (whole after shell and gills removed) 2.0 2018年食物搀杂(金属杂质含量)(修订)规例[38]Food adulteration (Metallic Contamination) (Amendment) Regulation 2018[38] 中国台湾Taiwan, China 甲壳类可食用组织Edible tissues of crustaceans 0.5 食品中污染物质及毒素卫生标准[39]Hygienic Standards for Contaminants and Toxins in Food [39]. 欧盟European Union 蟹类附肢的肌肉Muscles of crab appendages 0.5 European commission (EU) 2023/915 Regulation on the maximum levels of certain contaminants in food[40] 俄罗斯Russia 甲壳类Crustaceans 2.0 Customs Union TR CU 021/2011[41] 韩国South Korea 甲壳类Crustaceans 1.0 Food Code 2021[42] 带内脏的梭子蟹Swimming crab with entrails 5.0 美国American 甲壳类Crustaceans 3.0 Food and Drug Administration[43] 土耳其Turkey 甲壳类(蟹类的棕肉除外)Crustaceans(excluding brown meat of crabs) 0.5 Food Code 2011[44] 孟加拉Bangladesh 甲壳类Crustaceans 0.5 Bangladesh Bulletin 2014[41] | Show TableDownLoad:
CSV
目前,我国对于食品中污染物的限量仍以总的可食用部分计算,并没有针对甲壳类特定组织的浓度限制. 在进行水产品质量安全检测时,若仅将各个可食组织混合检测,并不能全面准确地评价重金属对甲壳类动物的污染程度,也无法准确评估不同可食组织的重金属潜在健康风险. 因此,对甲壳类的不同食用组织进行分类限量,将有助于科学评价水产品质量.
2. 三疣梭子蟹的可食用组织中总镉的分布特征(Distribution characteristics of total cadmium in edible tissues of Portunus trituberculatus)
环境中的镉可以通过各种途径进入三疣梭子蟹体内,并以不同的速率在各种器官中积累[45]. 前人研究发现,镉在三疣梭子蟹体内不同组织的蓄积量存在差异[46]. 本文总结了三疣梭子蟹不同可食组织中总镉含量的研究现状,以了解镉在各可食组织中的分布特征,期待为后续镉蓄积相关研究提供参考.
2.1 肝胰腺
作为三疣梭子蟹体内重要的消化代谢器官,肝胰腺兼具肝脏和胰腺的功能,能够直接参与金属的摄取、存储和排泄[47]. 同时,肝胰腺作为镉的靶标器官,还会从生物体内的其他组织积累镉[48]. 此外,肝胰腺中存在的金属硫蛋白(metallothionein,MT)还被认为与三疣梭子蟹对重金属的解毒机制有关[49]. 有报道指出,三疣梭子蟹肝胰腺对镉的响应较敏感,能够在感应到镉的胁迫后迅速提高MT的表达[50],产生的MT结合游离的镉,形成低毒状态的金属硫蛋白-镉(cadmium-metallothionein,Cd-MT),从而降低血淋巴中的镉浓度,保证其他维持生命的器官和组织功能不受影响[51 − 53]. 因此,有学者认为三疣梭子蟹的肝胰腺可能是镉的主要蓄积组织[54].
由于肝胰腺与性腺位置相连,难以剥离,研究人员常检测的是肝胰腺和性腺的混合组织(棕肉),而不是单独检测肝胰腺组织. 因此,当前单独针对三疣梭子蟹肝胰腺组织进行镉检测的报道较少. 在仅有的报道中,有学者指出三疣梭子蟹的肝胰腺中镉含量显著高于其他可食组织[33]. 如牛红鑫等[55]对上海市售的梭子蟹中镉的残留量进行了调查,发现梭子蟹各部位中镉的含量为肝胰腺(18.44±10.50) mg·kg−1>性腺(0.49±0.37) mg·kg−1>肌肉(0.22±0.15) mg·kg−1. 然而,仅少量文献不足以说明肝胰腺对镉的蓄积特异性,往后还需要更多的研究支撑.
目前,关于肝胰腺中镉的来源、迁移与归趋的机理问题尚未有科学解释,也未有更深入的分子生物学上的研究. 并且,除了金属硫蛋白之外,肝胰腺是否存在其他蓄积镉的途径也未有明确定论. 综上可知,有关三疣梭子蟹肝胰腺的镉蓄积问题还需要不断探索.
2.2 性腺
三疣梭子蟹的性腺是消费者极为喜爱食用的组织,尤其是发育成熟的性腺[27]. 然而,专门检测三疣梭子蟹性腺中镉含量的报道非常少,绝大多数检测的是性腺与肝胰腺混合物(棕肉)中镉的含量水平. 因为三疣梭子蟹的性腺与肝胰腺位置极为相近,以往有观点认为性腺也可能是三疣梭子蟹中重要的镉蓄积组织. 有学者认为肝胰腺与性腺分布相近,两者在重金属富集分布上具有高度的相似性[33]. 此外,对于肝胰腺中某些脂类及蛋白质会在蟹类性成熟时期通过血淋巴转运至生殖腺,有学者推测某些脂溶性的镉形态可能会随之转移至性腺中[56]. 但棕肉中的镉含量并不能说明镉在性腺中蓄积的情况. 因为棕肉中的成分会因性腺发育时期的不同而产生差异. 例如,有的文献[24]检测的棕肉样品中性腺尚未发育,此时棕肉的成分几乎全是肝胰腺. 另有文献[30]对性成熟的梭子蟹样品中的棕肉进行检测,此时棕肉成分中除了肝胰腺以外,性腺也占有不小的比例. 因此,不能仅依靠棕肉的镉含量水平去单独判断肝胰腺或性腺对镉的蓄积能力.
目前,有极少量报道[32]单独检测了性腺中的镉含量,指出肝胰腺中的镉含量要高于性腺. 但是如此少量的报道,并不足以说明镉在三疣梭子蟹性腺中的蓄积情况,仍需要后续对性腺进行单独检测和研究. 性腺是三疣梭子蟹繁殖的重要器官,探究性腺中镉的蓄积情况,对研究镉对三疣梭子蟹可能存在的生殖毒性[57],以及对消费者食用三疣梭子蟹的安全风险评估研究有着重要意义. 值得注意的是,目前对于三疣梭子蟹性腺不同发育时期的镉含量检测未见报道. 性腺不同发育时期中镉含量的变化规律也是研究三疣梭子蟹中镉蓄积机制的重要部分. 因此,对于三疣梭子蟹性腺中镉的蓄积问题后续需要针对性腺不同发育时期进行更为详细和深入的实验与研究.
2.3 肌肉
在绝大多数报道中,三疣梭子蟹的肌肉(包括胸肌、腿肌、钳肌)较其他可食组织的镉含量水平最低. 如Mei等[28]对东海舟山渔场捕捞的三疣梭子蟹进行了检测,发现内脏组织的Cd水平大约是肌肉组织的9.5倍. 有研究指出,蟹类肌肉组织中的重金属含量与多种复杂因素有关,包括环境参数、种间的生理变异、以及重金属的理化性质、生物活性和累积特性等[58]. 蟹类肌肉中的镉主要来源于血淋巴的输送[59]. 研究指出,环境中的镉进入三疣梭子蟹体内后,会先经过肝胰腺的解毒,若是肝胰腺中重金属过度积累,会加快镉向肌肉组织迁移的速度[49]. 研究指出,镉与生物体中内源性物质亲和力的差异导致其在组织器官蓄积的异质性[60]. 因此,可能是肌肉对重金属的低亲和力导致了镉在肌肉中的蓄积量远低于其他组织[61].
图1展示了部分文献报道的三疣梭子蟹各个可食用组织中镉含量的堆积百分比. 可见,在多数报道中,三疣梭子蟹的可食组织被分为棕肉和白肉(肌肉)组织进行检测,且棕肉中镉含量占比显著高于白肉;对于肝胰腺和性腺分开检测的报道较少,仅有的报道显示肝胰腺中镉含量占比远高于性腺和肌肉组织. 综合而言,关于镉在三疣梭子蟹各可食组织中的分布特征,目前较为明确的是肌肉组织的镉含量水平明显低于其他可食组织,尤其是低于棕肉组织和肝胰腺组织;对于肝胰腺组织,较多学者猜测其为三疣梭子蟹的镉主要蓄积组织,但对其单独进行镉含量检测的报道较少,需要后续进行更多的实验和研究;对于性腺组织,有观点认为性腺也可能是三疣梭子蟹中重要的镉蓄积组织,但相关报道太少并不足以支撑此观点,因此后续仍需要学者们对性腺展开相关研究,尤其要针对性腺不同发育时期进行研究.

3. 镉在三疣梭子蟹可食组织中的赋存形态研究进展(The progress of chemical speciation of cadmium in the edible tissues of Portunustri tuberculatus)
重金属在环境中的迁移转化规律及其对生物的毒理学效应,并非取决于重金属的总浓度,而是由其赋存形态的性质决定[60]. 镉在三疣梭子蟹体内的赋存形态可分类为两种,一种是镉与有机物结合形成的有机镉形态,第二种则是镉以游离的无机离子态形式存在[65]. 多数毒理学研究指出,镉的毒性与其存在形态有关,无机游离态镉的毒性较高,有机结合态镉毒性较低[66]. 目前,镉在生物体内具体的赋存形态尚未清晰,针对具有镉蓄积特异性的三疣梭子蟹开展不同镉形态的分析,不仅有助于揭示三疣梭子蟹中镉的蓄积机制,同时也能为不同形态镉在其他生物体内的研究提供探索思路与数据支持.
3.1 不同镉形态的提取方法的研究进展
研究镉的形态,首先需要将不同形态的镉从生物体内提取和分离出来. 因此,选择适当的提取剂是准确测定镉形态的前提条件. 并且提取形态所用的提取剂应能将生物体内不同形态的镉完整而充分地提取且不破坏其原有形态结构. 水生生物中镉形态的研究起步较晚,关于土壤[67 − 69]、陆生植物[70 − 74]、藻类[75 − 76]中的镉形态提取分离模式和方法研究较多,且大多数提取方法是利用连续化学浸提法,即利用不同的提取剂进行分级提取后测定提取剂中的镉. 至于这些提取剂是否会在连续提取的过程中影响和破坏镉的具体化学形态,鲜有报道.
目前,有学者对水生生物中镉形态的提取开展了相关的研究. 先是有学者尝试用碱性酶缓冲液对海豚肝脏进行提取,发现提取到的可溶态镉只占镉总量的45%左右,且这部分镉全部都与蛋白质(非金属硫蛋白)相结合[77]. 除此之外,还有学者尝试用柠檬酸和琥珀酸两种有机酸溶剂提取牡蛎匀浆液中的镉,提取率可达到90.2%—91.8%,但未研究此种提取法是否会对镉形态产生影响[78]. 针对不同的浸提方法,李敏[65]分析了渔用饲料的镉形态,发现有机溶剂环己烷和乙醇无法浸提出饲料中的有机态镉,同时发现较低浓度的盐酸就能使有机态镉发生解离. 也有研究人员对比了不同提取剂的逐级提取效果,田姣姣等[79]对比了Tris-HCl缓冲溶液、HCl(pH值3.5)和碱性蛋白酶3种不同的提取剂对三疣梭子蟹可食用组织中镉形态逐级提取的效果,结果表明Tris-HCl具备较好的提取效果,对肝胰腺、性腺等组织的镉提取率高达90%,但对肌肉组织中的提取率仅为60%,仍有37%的非游离态镉存在,而经碱性蛋白酶处理后可有效溶出.
综上所述,镉形态的提取常用到酸性溶液、有机溶剂以及碱性蛋白酶缓冲液,但这些提取溶液的使用都有可能会引起有机镉形态的破坏,从而影响形态检测的结果. 目前,有研究人员开始使用与生物体生理条件相似的中性Tris-HCI缓冲溶液,以最大程度保证镉的形态在提取过程中不发生变化. 对于镉形态的研究,提取剂的选择固然重要,但目前大部分的镉形态研究仍停留在依赖不同的提取剂将镉形态进行分类后检测其中无机镉含量,未能实现直接检测镉的具体化学形态. 因此,往后对于提取方法的研究应当针对镉的具体化学形态,这对于镉具体化学形态的研究也具有重要意义.
3.2 三疣梭子蟹中无机镉形态的研究现状
国内有关水产品中无机镉的分析研究起步较晚,且所用方法大多为高效液相色谱在线联用电感耦合等离子体质谱法(high performance liquid chromatography-inductively coupled plasma mass spectrometer,HPLC-ICP-MS). 章红等[80]最早建立了水产品中无机镉的测定方法. 他们使用石墨炉原子吸收法(graphite furnace atomic absorption spectrometry, GFAAS),利用盐酸作为浸提液,测定了7种水产品中无机镉的含量. 但此方法并未分析使用盐酸浸提对样品中无机镉离子的提取效果,也没有考虑到是否存在有机镉形态被盐酸破坏转化为无机镉. 随后,赵艳芳等[81]利用Tris-HCl缓冲溶液作为浸提液,借助CG5A阳离子保护柱和CS5A阳离子分析柱对样品中Cd2+分离,建立了HPLC-ICP-MS法测定海水贝中离子态镉的含量. 自此之后,大部分有关于水产品中无机镉离子的检测方法都在此研究基础上做出优化. 如姜芳等[82]同样采用HPLC-ICP-MS法测定了海水贝中无机镉离子含量,通过进一步优化前处理过程及流动相浓度,完善了海水贝中无机镉离子的定性和定量分析方法.
国内有关三疣梭子蟹中无机镉离子的形态研究少之又少. 仅田姣姣[83]和冯瑞雪等[84]对三疣梭子蟹不同组织中的无机镉进行了探究. 在赵艳芳[81]的研究基础上,田姣姣[83]探讨了流动相浓度、pH、流速对HPLC-ICP-MS检测Cd2+的影响规律,建立了三疣梭子蟹各部位离子态镉检测的前处理方法,并且测得的三疣梭子蟹中只有蟹膏部位含有离子态镉,约占蟹膏部位总镉的9.4%. 这说明三疣梭子蟹不同部位镉的赋存形态存在差异. 而在此基础上,冯瑞雪等[84]进一步优化了HPLC-ICP-MS检测离子态镉的方法,探究了经过蒸煮和模拟消化后三疣梭子蟹各部位镉的变化. 发现经过蒸煮加工及模拟消化后,肌肉中Cd2+含量上升,蟹膏、蟹黄中Cd2+含量下降. 他们认为在蒸煮过程中,可能因汁液流失、浓缩效应或在不同部位间存在的迁移转化使样本中Cd2+含量发生变化. 综合来看,针对三疣梭子蟹体内无机镉形态的研究尚处于起步阶段,仍需要后续不断优化和完善提取、检测、定量方法,以便深入地探索三疣梭子蟹体内镉赋存形态的分布特征及蓄积机制,同时也为后续更加科学合理地评估三疣梭子蟹的食用安全风险提供研究数据.
3.3 三疣梭子蟹中有机镉形态的研究现状
研究发现,三疣梭子蟹体内毒性较高的无机镉离子含量占总镉的比例较小,间接地表明了低毒性的有机镉形态在总镉中的占比较大[85]. 目前,分析生物体中有机镉形态最常用的方法是体积排阻色谱(size exclusion chromatography,SEC)与高效液相色谱(HPLC)串联电感耦合等离子体质谱(ICP-MS)联用技术[73, 76, 86 − 87]. 然而,有机镉形态的检测存在着标准物质缺乏的问题,难以对生物体内的有机镉形态进行定性. 为此,有学者尝试将无机镉的标准溶液与其他溶液进行络合,根据镉络合物的保留时间来初步判断镉可能在生物体内存在的形态[73].
在前人的研究基础上,赵艳芳等[8]运用了SEC-HPLC-ICP-MS首次对三疣梭子蟹中的有机镉形态进行分析. 发现三疣梭子蟹肝胰腺组织中的镉主要以金属硫蛋白-镉(metallothionein-Cadmium,MT-Cd)和半胱氨酸-镉(cysteine-Cadmium,Cys-Cd)两种形态存在. 并且,根据总镉量的不同,肝胰腺中有机镉的主要存在形态也有所不同. 而对于肌肉组织,镉主要以Cys-Cd形态存在. 此后,田姣姣等[79]进一步对三疣梭子蟹体内镉赋存形态开展了研究. 他们利用Tris-HCl缓冲液进行提取,结合超滤结合透析的方式,对提取液中不同分子质量镉复合物进行分离测定. 结果显示,分子质量>10 kDa的镉复合物占比最高,可能是MT以外的其它蛋白或多糖、脂类等与镉形成大分子复合物. 这说明三疣梭子蟹在MT所介导的途径之外,可能存在其它的蓄积代谢机制;同时<500 Da的镉组分约占4%—25%,可能是存在离子态镉、氨基酸-镉等小分子镉组分.
目前,三疣梭子蟹体内有机镉形态尚未完全明确,并且有机镉的标准物质缺乏,定性与定量存在困难. 后续需要进一步确定三疣梭子蟹体内有机镉的具体形态,同时确定不同结合态的有机镉的毒性效应和转化机制,对于三疣梭子蟹中无机镉与有机镉的占比问题也值得进行更深入的探索与验证.
无论有机镉还是无机镉,国内外针对三疣梭子蟹中镉的赋存形态分析的研究非常少. 并且,研究镉形态的检测手段、测定方法并不完善,仪器设备的发展水平和镉形态的提取与分离都限制了镉形态分析. 此外,由于缺少镉形态的标准品,无法明确三疣梭子蟹等水产品中镉存在的不同形态及其相关毒理效应[31]. 因此,后续需要对镉形态的定性和定量分析开展进一步的研究,以便更科学地评估不同形态镉的毒性效应,并将研究结论应用于完善相关限量标准.
4. 三疣梭子蟹中镉的人类健康风险评估(Human health risk assessment of cadmium in Portunus trituberculatus)
镉作为一种高危有毒的环境污染物,仅关注其在三疣梭子蟹中的蓄积量不足以评估其对生物体健康的危害风险. 因此,对三疣梭子蟹中的镉进行健康风险评估可以将三疣梭子蟹中镉的膳食摄入量与机体负荷有机联系起来,同时监测镉对人体健康的潜在影响. 人体健康风险评估不单取决于食品中重金属的摄入量,还需要借助各种指标和参数[88]. 目标危害系数(Target hazard quotient, THQ)和靶标癌症风险(Target cancer risk, TCR)是人类健康风险评估最常关注的指标. THQ可以评估通过水产品途径的接触单一重金属的非致癌健康风险[89]. TCR是一种衡量个体在食用水产品期间长期暴露于潜在致癌物而患癌症风险的指标[90]. 除此之外,生物可及性(Bioaccessibility)也常用于评估食品中有毒物质的危害性. 它是指在胃肠消化过程中,营养素或污染物从食物基质中释放溶出并可以被人体吸收的百分比[91]. 考虑到一定比例的重金属不具有生物可及性[92],将生物可及性因素纳入风险计算有助于更全面地进行评估.
目前,有关三疣梭子蟹中镉的人类健康风险评估的研究大多着眼于两个方面. 一方面,研究主要针对不同食用人群进行健康风险评估[93 − 94],年龄、性别和体重是重要的影响因素. 据报道,对于同性别不同年龄段人群,三疣梭子蟹中引起的健康风险趋势表现为年龄越小,健康风险越大;对于同年龄段不同性别人群,三疣梭子蟹中镉引起的健康风险趋势表现为女性大于男性. 有学者认为,因为计算THQ时体重作为分母,低龄人群和女性群体较轻的体重导致了THQ结果较大,因此,低龄儿童和女性群体食用三疣梭子蟹的风险相对较大,并且在食用三疣梭子蟹时需要避免食用过量. 另一方面,针对三疣梭子蟹不同可食组织进行健康风险评估. 对于肌肉组织,有研究指出[49],无论镉暴露浓度的高低(1—50 μg·L−1),肌肉中的THQ值均小于限值1,这说明食用三疣梭子蟹肌肉组织造成的健康风险属于低关注级别. 然而,对于肝胰腺或棕肉组织,常常显示较高的食用风险[27, 30]. 如Yang等[95]分别评估了食用三疣梭子蟹的白肉和棕肉样品的人类健康风险,发现棕肉的非致癌风险(THQ)是白肉的16倍,致癌风险(TCR)是白肉的100倍. 即使经过烹饪后棕肉中镉的含量下降,但THQ仍接近风险限值. Zhao等[27]利用镉的生物可及性浓度计算了三疣梭子蟹组织允许每周消耗量,并由此评估了山东沿海地区三疣梭子蟹的食用安全性,指出在山东的成年人每周食用0.13 kg棕色肉和1.56 kg白肉,对机体不会产生明显的危害,并且不建议食用棕肉,更推荐食用白肉. 此外,有研究指出[95],煮熟后的三疣梭子蟹中的镉浓度有所下降,建议消费者最好只食用熟蟹黄或熟蟹肉,避免食用烹饪螃蟹的汁水或油,以减少镉对人体健康的危害. 综上,对于三疣梭子蟹不同的可食组织,消费者最好少食用或不食用肝胰腺或棕肉,更推荐食用安全性更高的肌肉组织.
关于三疣梭子蟹的人类健康风险评估,后续研究除了可以针对如孕妇、幼童等易受镉污染影响的敏感人群进行研究,还可以针对三疣梭子蟹食用频次和数量较高的沿海居民进行健康风险观察. 另外,野生捕捞和规模养殖的三疣梭子蟹中镉的健康风险尚未见报道. 总而言之,食用三疣梭子蟹作为一种可预见的、可判断的人类膳食摄入镉的途径,研究三疣梭子蟹中镉的人类健康风险有助于相关限量的制定以及对食用量和膳食人群做出推荐.
5. 结论与展望(Conclusion and prospect)
针对三疣梭子蟹中镉的特异性蓄积问题,相关研究大多着眼于三疣梭子蟹不同组织中总镉的含量水平,而有关镉在三疣梭子蟹中的蓄积机制及赋存形态问题的研究仍处于探索阶段. 虽然已经有研究初步检测了三疣梭子蟹不同组织中的镉形态,但受限于镉形态的提取、分离、定性和定量方法的不完善,后续仍需要建立相应的技术方法,以便更好地研究三疣梭子蟹中不同形态镉的具体类型以及各组织中不同形态镉的占比问题,为深入研究三疣梭子蟹中镉的蓄积机制和赋存形态提供技术基础,同时为食用三疣梭子蟹的镉安全性问题提供新的风险评价思路.
基于三疣梭子蟹中镉的蓄积机制及赋存形态尚未明晰的现状,今后可从以下几方面着手进行更深入的研究:①镉在三疣梭子蟹不同组织中的迁移、转化、蓄积以及释放的全过程未有研究,可以开展相关的检测实验,寻找镉在三疣梭子蟹中的变化规律,进而探索镉在三疣梭子蟹体内的蓄积机制,还可为后续标准限量的修订提供科学参考依据. ②为更深入探索镉在三疣梭子蟹中的蓄积机制,也为更加全面评估不同毒性的镉形态对人类健康的影响,三疣梭子蟹各组织中赋存的镉形态研究必不可少. 镉形态标准物缺乏、分离分析方法不完善、仪器设备不够精密等问题,有待后续的学者们持续研究和解决. 建立科学的方法,才能够更清晰地了解三疣梭子蟹中镉的赋存形态及分布规律,最终摸索出三疣梭子蟹对镉的蓄积机制. ③研究三疣梭子蟹中镉的特异性蓄积问题,终是为了能更加准确地评估人们食用三疣梭子蟹的膳食风险. 虽然前人已经进行了相关研究,但考虑到人群和时空的差异,不能仅依靠过往的评估结果进行食用三疣梭子蟹的安全性判断,需要持续对食用三疣梭子蟹进行镉的安全性评估. 在进行评估时,孕妇、幼童等易受镉污染影响的敏感人群以及食用三疣梭子蟹频次和数量较高的沿海居民都是重点研究对象. 此外,还可以进行不同养殖方式、不同食用组织、不同镉赋存形态等方面的健康风险评估,全面分析三疣梭子蟹中镉的膳食风险,保障人们在食用三疣梭子蟹时不受重金属镉的威胁.
-

图 1 基于CRISPR/Cas系统的离子传感器:(a)RCD-AuNP球形核酸与CRISPR/Cas整合的Pb2+传感器[17];(b)适配体-CRISPR/Cas系统检测Cd2+[21];(c)核糖开关-CRISPR/Cas系统检测F−[22]
Figure 1. CRISPR/Cas-based biosensor for the detection of heavy metal ions and anion (F−): (a) RCD-AuNP spherical nucleic acids and CRISPR/Cas system for the sensing of Pb2+ [17]; (b) An aptamer-CRISPR/Cas system as a Cd2+ sensor [21]; (c) A riboswitch-CRISPR/Cas system for the detection of F− [22]
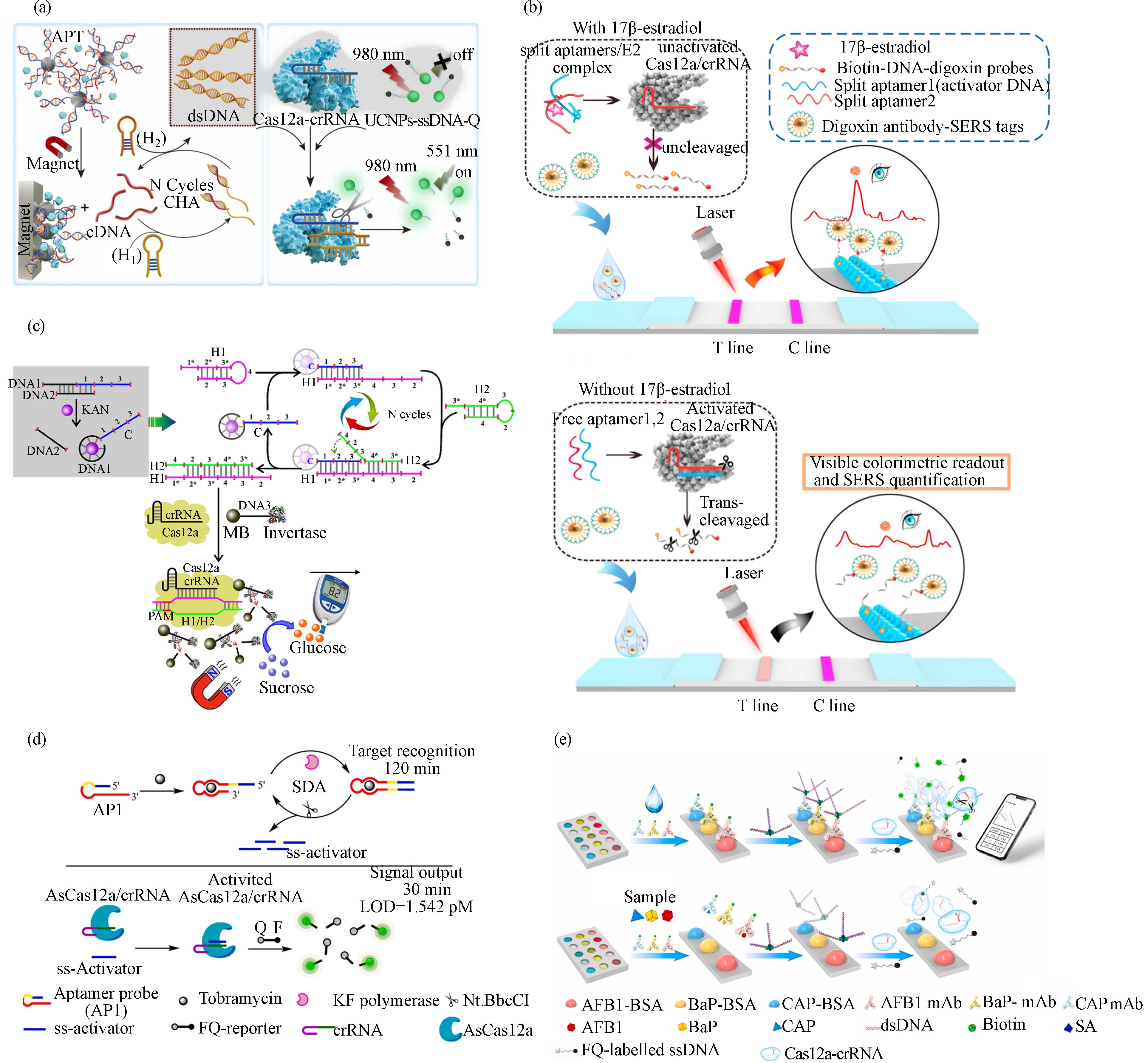
图 2 基于CRISPR/Cas系统的内分泌干扰物、抗生素和持久性有机污染物的生物传感器:(a)适配体-CRISPR/Cas系统检测E2[25];(b)整合ELISA与CRISPR/Ca系统检测E2[26];(c)基于适配体-HCR-CRISPR/Cas的卡那霉素传感器[28];(d)基于适配体-SDA-CRISPR/Cas的妥布霉素传感器[30];(e)检测BaP、AFB1和CAP的“iPOCT”系统[34]
Figure 2. CRISPR/Cas-based biosensors for the detection of endocrine disrupting chemicals, antibiotics, and persistent organic pollutants: (a) Aptamer-CRISPR/Cas system for the sensing of E2[25]; (b) Combination of ELISA and CRISPR/Cas system for the detection of E2[26]; (c) Aptamers, HCR, and CRISPR/Cas-based biosensors for kanamycin[28]; (d) Aptamer-SDA-CRISPR/Cas system for Tobramycin detection[30]; (e) An “iPOCT” system for the detection of BaP, AFB1, and CAP[34]

图 3 基于CRISPR/Cas系统检测农药和真菌毒素的生物传感器:(a)检测农药的三酶级联系统 [36];(b)适配体-CRISPR/Cas系统检测AFB1[39];(c)适配体-CRISPR/Cas系统检测胶霉毒素[40]
Figure 3. CRISPR/Cas -based biosensors for the detection of pesticides and mycotoxins: (a) A three-enzymes-cascade system for the detection of pesticides[36]; (b), and (c) Aptamer-CRISPR/Cas-based biosensor for AFB1 (b) [39]; and gliotoxin (c) [40]

图 4 基于CRISPR/Cas检测细菌的生物传感器. (a)适配体-CRISPR/Cas系统检测鼠伤寒沙门菌[42];(b)基于RNA-cleaving DNAzyme切割ccrRNA激活CRISPR/Cas系统的策略检测大肠杆菌和肺炎克雷伯菌[43]
Figure 4. CRISPR/Cas-based biosensors for the detection of bacteria: (a) Aptamer-CRISPR/Cas-based biosensor for Salmonella typhimurium[42]; (b) A CRISPR/Cas-based biosensor that is activated by a ccrRNA-cleaving DNAzyme for the detection of Escherichia coli and Klebsiella pneumoniae[43]
表 1 基于CRISPR/Cas系统的传感器在环境分析中的应用
Table 1. Application of CRISPR/Cas-based biosensors in the environmental analysis field
靶标Targets 信息转换元件Information converting element 效应蛋白The type of Cas 检出限Limit of detection 检测时间Detection time 信号输出Signal output 样品Samples 检测设备Testing equipment 文献Ref. 重金属离子 Pb2+ DNAzyme(GR-5) Cas12a 0.053 nmol·L−1 15 min 荧光法 — — [16] Pb2+ DNAzyme(8-17E) Cas12a 86 fmol·L−1 — 荧光法 血清、空气颗粒、土壤 智能手机 [17] Pb2+ DNAzyme(GR-5) Cas12a 0.54 nmol·L−1 — 比色法 食用油、白酒 肉眼观察 [18] Pb2+ DNAzyme(GR-5) Cas12a 0.02 pmol·L−1 — 电化学 酒、花生、大米、食用油 电化学分析仪 [19] Pb2+ DNAzyme(GR-5) Cas12a 0.48 nmol·L−1 — 荧光法 矿泉水 便携式3D打印设备 [20] Cd2+ 适配体 Cas12a 60 pmol·L−1 120 min 荧光法 湖水、大米 — [21] Zn2+ 变构转录因子(SmtB) Cas13a 1 μmol·L−1 20 min 荧光法 市政水样 便携式3D打印设备 [22] 阴离子 F− 核糖开关(crcB) Cas13a 1 μmol·L−1 20 min 荧光法 — 便携式3D打印设备 [22] F− 核糖开关 Cas13a 1.7 μmol·L−1 30 min 荧光法 瓶装水、自来水、湖水 荧光计 [23] 内分泌干扰物 双酚A、雌二醇 适配体 Cas12a 0.06、0.08 nmol·L−1 20 min 荧光法 尿液、污水 — [24] 雌二醇 适配体 Cas12a 0.015 ng·mL−1 — 比色法 牛奶、蛋类、猪肉 肉眼观察 [25] 内分泌干扰物 雌二醇 适配体 Cas12a 180 fmol·L−1 — 比色法 牛奶、自来水、血清、尿液 LFA [26] 抗生素 卡那霉素 适配体 Cas12a 4.06 pmol·L−1 30 min 同位素法 野生鱼血清、肌肉、肝脏 电感耦合等离子体-质谱法 [27] 卡那霉素 适配体 Cas12a 1 pmol·L−1 — 酶催化蔗糖转化为葡萄糖 河水、牛奶 血糖仪 [28] 卡那霉素 适配体 Cas12a 14.8 nmol·L−1 26min 比色法 牛奶 侧向层析测定 [29] 妥布霉素 适配体 Cas12a 1.542 pmol·L−1 — 荧光法 湖水、牛奶 紫外灯 [30] 四环素 变构转录因子(TetR) Cas12a 2 μmol·L−1 — 荧光法 环境水样 手持可视化荧光仪 [31] 氨苄青霉素 适配体 Cas12a 0.01 nmol·L−1 30 min 荧光法 鲜奶、生蛋清、生蜂蜜 荧光仪 [32] 氨苄青霉素 适配体 Cas14a 2.06 nmol·L−1 45 min 同位素法 江河水样 ICP-MS [33] 持久性有机污染物 苯并[a]芘 抗体 Cas12a 4.971 fg·mL−1 — 荧光法 湖水、大豆油 智能手机 [34] 对羟基苯甲酸 变构转录因子 Cas12a 1.8 nmol·L−1 — 荧光法 — 荧光仪 [35] 农药 对氧磷、敌敌畏、内吸磷 DNAzyme(8-17E变体) Cas12a 270、406、218 pg·mL−1 — 荧光法 柑橘类水果、卷心菜 — [36] 啶虫脒 适配体 Cas12a 2.7 pmol·L−1 — 电化学发光 生菜 肉眼观察 [37] 真菌毒素 脱氧雪腐镰刀菌烯醇 抗体 Cas12a 0.061 ng·mL−1 30 min 粒子计数 玉米、水 粒子计数器 [38] 黄曲霉毒素 抗体 Cas12a 0.00257 fg·mL−1— 荧光法 花生、面粉 智能手机 [34] 黄曲霉毒素 适配体 Cas12a 0.8 ng·mL−1 20 min 比色法 红酒、啤酒、牛奶 荧光板读取仪 [39] 胶霉毒素 适配体 Cas12a 2.4 fmol·L−1 55 min 电化学 苹果、胡萝卜、红薯、马铃薯、玉米 手持式电化学分析仪 [40] 赭曲霉毒素 适配体 Cas12a 0.83 ng/mL 60 min 荧光法 玉米粉 荧光仪 [41] 细菌 沙门氏菌 适配体 Cas12a 20 CFU·mL−1 — 电化学 牛奶 电化学工作站 [42] 大肠杆菌 DNAzyme(EC1) Cas12a 102 CFU·mL−1 107 min 荧光法 尿液 荧光仪 [43] 肺炎球菌 DNAzyme(KP6) Cas12a 102 CFU·mL−1 — 荧光法 — 荧光仪 [43] “—”,文献缺乏相应信息. “—”, Lack of information.
下载: 导出CSV
-
[1] ISHINO Y, SHINAGAWA H, MAKINO K, et al. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product[J]. Journal of Bacteriology, 1987, 169(12): 5429-5433. doi: 10.1128/jb.169.12.5429-5433.1987 [2] MOJICA F J, DIEZ-VILLASENOR C, GARCIA-MARTINEZ J, et al. Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements[J]. Journal Of Molecular Evolution, 2005, 60(2): 174-182. doi: 10.1007/s00239-004-0046-3 [3] MOJICA F J, DIEZ-VILLASENOR C, SORIA E, et al. Biological significance of a family of regularly spaced repeats in the genomes of Archaea, Bacteria and mitochondria[J]. Molecular Microbiology, 2000, 36(1): 244-246. doi: 10.1046/j.1365-2958.2000.01838.x [4] JANSEN R, EMBDEN J D, GAASTRA W, et al. Identification of genes that are associated with DNA repeats in prokaryotes[J]. Molecular Microbiology, 2002, 43(6): 1565-1575. doi: 10.1046/j.1365-2958.2002.02839.x [5] QIN J J, WANG W, GAO L Q, et al. Emerging biosensing and transducing techniques for potential applications in point-of-care diagnostics[J]. Chemical Science, 2022, 13(10): 2857-2876. doi: 10.1039/D1SC06269G [6] KIM S, JI S, KOH H R. CRISPR as a Diagnostic Tool[J]. Biomolecules, 2021, 11(8): 1162. doi: 10.3390/biom11081162 [7] YUE H H, HUANG M Q, TIAN T, et al. Advances in clustered, regularly interspaced short palindromic repeats (CRISPR)-based diagnostic assays assisted by micro/nanotechnologies[J]. ACS Nano, 2021, 15(5): 7848-7859. doi: 10.1021/acsnano.1c02372 [8] GOOTENBERG J S, ABUDAYYEH O O, LEE J W, et al. Nucleic acid detection with CRISPR-Cas13a/C2c2[J]. Science, 2017, 356(6336): 438-442. doi: 10.1126/science.aam9321 [9] CHEN J S, MA E, HARRINGTON L B, et al. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity[J]. Science, 2018, 360(6387): 436-439. doi: 10.1126/science.aar6245 [10] BROUGHTON J P, DENG X D, YU G X, et al. CRISPR–Cas12-based detection of SARS-CoV-2[J]. Nature Biotechnology, 2020, 38(7): 870-874. doi: 10.1038/s41587-020-0513-4 [11] SELVAM K, NAJIB M A, KHALID M F, et al. RT-LAMP CRISPR-Cas12/13-Based SARS-CoV-2 Detection Methods[J]. Diagnostics, 2021, 11(9): 1646. doi: 10.3390/diagnostics11091646 [12] LIANG Y H, LIN H Q, ZOU L R, et al. CRISPR-Cas12a-Based Detection for the Major SARS-CoV-2 Variants of Concern[J]. Microbiology Spectrum, 2021, 9(3): e0101721. doi: 10.1128/Spectrum.01017-21 [13] RANJBARIAN F, SHARMA S, FALAPPA G, et al. Isocratic HPLC analysis for the simultaneous determination of dNTPs, rNTPs and ADP in biological samples[J]. Nucleic Acids Research, 2022, 50(3): e18. doi: 10.1093/nar/gkab1117 [14] ZHANG S M, WANG H B, ZHU M J. A sensitive GC/MS detection method for analyzing microbial metabolites short chain fatty acids in fecal and serum samples[J]. Talanta, 2019, 196: 249-254. doi: 10.1016/j.talanta.2018.12.049 [15] ALBERTI D, VAN'T ERVE M, STEFANIA R, et al. A quantitative relaxometric version of the ELISA test for the measurement of cell surface biomarkers[J]. Angewandte Chemie International Edition, 2014, 53(13): 3488-3491. doi: 10.1002/anie.201310959 [16] LI J J, YANG S S, ZUO C, et al. Applying CRISPR-Cas12a as a Signal Amplifier to Construct Biosensors for Non-DNA Targets in Ultralow Concentrations[J]. ACS Sensors, 2020, 5(4): 970-977. doi: 10.1021/acssensors.9b02305 [17] LI Y Y, LI H D, FANG W K, et al. Amplification of the Fluorescence Signal with Clustered Regularly Interspaced Short Palindromic Repeats-Cas12a Based on Au Nanoparticle-DNAzyme Probe and On-Site Detection of Pb2+ Via the Photonic Crystal Chip[J]. ACS Sensors, 2022, 7(5): 1572-1580. doi: 10.1021/acssensors.2c00516 [18] XU S Q, WANG S T, GUO L, et al. Nanozyme-catalysed CRISPR-Cas12a system for the preamplification-free colorimetric detection of lead ion[J]. Analytica Chimica Acta, 2023, 1243: 340827. doi: 10.1016/j.aca.2023.340827 [19] YUE Y Y, WANG S T, JIN Q, et al. A triple amplification strategy using GR-5 DNAzyme as a signal medium for ultrasensitive detection of trace Pb2+ based on CRISPR/Cas12a empowered electrochemical biosensor[J]. Analytica Chimica Acta, 2023, 1263: 341241. doi: 10.1016/j.aca.2023.341241 [20] CHEN Y J, WU H, QIAN S W J, et al. Applying CRISPR/Cas system as a signal enhancer for DNAzyme-based lead ion detection[J]. Analytica Chimica Acta, 2022, 1192: 339356. doi: 10.1016/j.aca.2021.339356 [21] MA X C, SUO T Y, ZHAO F R, et al. Integrating CRISPR/Cas12a with strand displacement amplification for the ultrasensitive aptasensing of cadmium(II)[J]. Analytical and Bioanalytical Chemistry, 2023, 415(12): 2281-2289. doi: 10.1007/s00216-023-04650-6 [22] IWASAKI R S, BATEY R T. SPRINT: a Cas13a-based platform for detection of small molecules[J]. Nucleic Acids Research, 2020, 48(17): e101. doi: 10.1093/nar/gkaa673 [23] MA Y, MOU Q B, YAN P, et al. A highly sensitive and selective fluoride sensor based on a riboswitch-regulated transcription coupled with CRISPR-Cas13a tandem reaction[J]. Chemical Science, 2021, 12(35): 11740-11747. doi: 10.1039/D1SC03508H [24] ZHAO Y Q, ZHU L, DING Y X, et al. Simple and cheap CRISPR/Cas12a biosensor based on plug-and-play of DNA aptamers for the detection of endocrine-disrupting compounds[J]. Talanta, 2023, 263: 124761. doi: 10.1016/j.talanta.2023.124761 [25] WANG Y, PENG Y, LI S, et al. The development of a fluorescence/colorimetric biosensor based on the cleavage activity of CRISPR-Cas12a for the detection of non-nucleic acid targets[J]. Journal of Hazardous Materials, 2023, 449: 131044. doi: 10.1016/j.jhazmat.2023.131044 [26] LI Q, LI X B, ZHOU P X, et al. Split aptamer regulated CRISPR/Cas12a biosensor for 17beta-estradiol through a gap-enhanced Raman tags based lateral flow strategy[J]. Biosensors and Bioelectrons, 2022, 215: 114548. doi: 10.1016/j.bios.2022.114548 [27] HU J Y, SONG H J, ZHOU J, et al. Metal-Tagged CRISPR/Cas12a Bioassay Enables Ultrasensitive and Highly Selective Evaluation of Kanamycin Bioaccumulation in Fish Samples[J]. Analytical Chemistry, 2021, 93(42): 14214-14222. doi: 10.1021/acs.analchem.1c03094 [28] CHEN J H, SHI G, YAN C. Portable biosensor for on-site detection of kanamycin in water samples based on CRISPR-Cas12a and an off-the-shelf glucometer[J]. Science of The Total Environment, 2023, 872: 162279. doi: 10.1016/j.scitotenv.2023.162279 [29] LI X P, CHEN X J, MAO M X, et al. Accelerated CRISPR/Cas12a-based small molecule detection using bivalent aptamer[J]. Biosensors and Bioelectronics, 2022, 217: 114725. doi: 10.1016/j.bios.2022.114725 [30] LI D W, LING S, WU H S, et al. CRISPR/Cas12a-based biosensors for ultrasensitive tobramycin detection with single- and double-stranded DNA activators[J]. Sensors and Actuators B: Chemical, 2022, 355: 131329. doi: 10.1016/j.snb.2021.131329 [31] MAHAS A, WANG Q C, MARSIC T, et al. Development of Cas12a-Based Cell-Free Small-Molecule Biosensors via Allosteric Regulation of CRISPR Array Expression[J]. Analytical Chemistry, 2022, 94(11): 4617-4626. doi: 10.1021/acs.analchem.1c04332 [32] YEE B J, SHAFIQAH N F, MOHD-NAIM N F, et al. A CRISPR/Cas12a-based fluorescence aptasensor for the rapid and sensitive detection of ampicillin[J]. International Journal of Biological Macromolecules, 2023, 242(Pt 4): 125211. [33] HU J Y, ZHOU J, LIU R, et al. Element probe based CRISPR/Cas14 bioassay for non-nucleic-acid targets[J]. Chemical Communications, 2021, 57(80): 10423-10426. doi: 10.1039/D1CC03992J [34] ZHAO Y, WU W Q, TANG X Q, et al. A universal CRISPR/Cas12a-powered intelligent point-of-care testing platform for multiple small molecules in the healthcare, environment, and food[J]. Biosensors and Bioelectronics, 2023, 225: 115102. doi: 10.1016/j.bios.2023.115102 [35] LIANG M D, LI Z L, WANG W S, et al. A CRISPR-Cas12a-derived biosensing platform for the highly sensitive detection of diverse small molecules[J]. Nature Communications, 2019, 10(1): 3672. doi: 10.1038/s41467-019-11648-1 [36] FU R J, WANG Y W, LIU Y L, et al. CRISPR-Cas12a based fluorescence assay for organophosphorus pesticides in agricultural products[J]. Food Chemistry, 2022, 387: 132919. doi: 10.1016/j.foodchem.2022.132919 [37] LI Y, YANG F, YUAN R, et al. Electrochemiluminescence covalent organic framework coupling with CRISPR/Cas12a-mediated biosensor for pesticide residue detection[J]. Food Chemistry, 2022, 389: 133049. doi: 10.1016/j.foodchem.2022.133049 [38] LI L T, HONG F, PAN S X, et al. "Lollipop" particle counting immunoassay based on antigen-powered CRISPR-Cas12a dual signal amplification for the sensitive detection of deoxynivalenol in the environment and food samples[J]. Journal of Hazardous Materials, 2023, 455: 131573. doi: 10.1016/j.jhazmat.2023.131573 [39] NIU C Q, XING X H, ZHANG C. A novel strategy for analyzing aptamer dominated sites and detecting AFB1 based on CRISPR–Cas12a[J]. Sensors & Diagnostics, 2023, 2(1): 155-162. [40] MA X, ZHANG Y, QIAO X J, et al. Target-Induced AIE Effect Coupled with CRISPR/Cas12a System Dual-Signal Biosensing for the Ultrasensitive Detection of Gliotoxin[J]. Analytical Chemistry, 2023, 95(31): 11723-11731. doi: 10.1021/acs.analchem.3c01760 [41] MAO Z F, WANG X J, CHEN R P, et al. Upconversion-mediated CRISPR-Cas12a biosensing for sensitive detection of ochratoxin A[J]. Talanta, 2022, 242: 123232. doi: 10.1016/j.talanta.2022.123232 [42] LIU X, BU S J, FENG J Q, et al. Electrochemical biosensor for detecting pathogenic bacteria based on a hybridization chain reaction and CRISPR-Cas12a[J]. Analytical and Bioanalytical Chemistry, 2022, 414(2): 1073-1080. doi: 10.1007/s00216-021-03733-6 [43] WU Y P, CHANG D R, CHANG Y Y, et al. Nucleic Acid Enzyme-Activated CRISPR-Cas12a With Circular CRISPR RNA for Biosensing[J]. Small, 2023, 19(41): e2303007. doi: 10.1002/smll.202303007 [44] SANTORO S W, JOYCE G F. A general purpose RNA-cleaving DNA enzyme[J]. Proceedings of the National Academy of Sciences of the United States of America, 1997, 94(9): 4262-4266. [45] LI J, ZHENG W, KWON A H, et al. In vitro selection and characterization of a highly efficient Zn(II)-dependent RNA-cleaving deoxyribozyme[J]. Nucleic Acids Research, 2000, 28(2): 481-488. doi: 10.1093/nar/28.2.481 [46] GAO P, LIU B, PAN W, et al. A Spherical Nucleic Acid Probe Based on the Au–Se Bond[J]. Analytical Chemistry, 2020, 92(12): 8459-8463. doi: 10.1021/acs.analchem.0c01204 [47] CUTLER J I, AUYEUNG E, MIRKIN C A. Spherical Nucleic Acids[J]. Journal of the American Chemical Society, 2012, 134(3): 1376-1391. doi: 10.1021/ja209351u [48] CHEN Y, CHEN L, OU Y, et al. DNAzyme-based biosensor for Cu2+ ion by combining hybridization chain reaction with fluorescence resonance energy transfer technique[J]. Talanta, 2016, 155: 245-249. doi: 10.1016/j.talanta.2016.04.057 [49] SARAN R, LIU J W. A Silver DNAzyme[J]. Analytical Chemistry, 2016, 88(7): 4014-4020. doi: 10.1021/acs.analchem.6b00327 [50] ZHOU W H, VAZIN M, YU T M, et al. In vitro selection of chromium-dependent DNAzymes for sensing chromium(III) and chromium(VI)[J]. Chemistry, 2016, 22(28): 9835-9840. doi: 10.1002/chem.201601426 [51] Wu Y T, Torabi S F, Lake R J, et al. Simultaneous Fe2+/Fe3+ imaging shows Fe3+ over Fe2+ enrichment in Alzheimer's disease mouse brain[J]. Science Advances, 2023, 9(16): eade7622. doi: 10.1126/sciadv.ade7622 [52] 郑星, 马明, 崔力, 等. 应用于环境分析的切割RNA的脱氧核酶研究进展[J]. 化学通报, 2022, 85(7): 770-780. ZHENG X, MA M, CUI L, et al. Advances in RNA-Cleaving DNAzymes for environmental analysis[J]. Chemistry, 2022, 85(7): 770-780 (in Chinese).
[53] RAJENDRAN M, ELLINGTON A D. Selection of fluorescent aptamer beacons that light up in the presence of zinc[J]. Analytical and Bioanalytical Chemistry, 2008, 390(4): 1067-1075. doi: 10.1007/s00216-007-1735-8 [54] WANG H Y, CHENG H, WANG J N, et al. Selection and characterization of DNA aptamers for the development of light-up biosensor to detect Cd(II)[J]. Talanta, 2016, 154: 498-503. doi: 10.1016/j.talanta.2016.04.005 [55] WU Y G, ZHAN S S, WANG L M, et al. Selection of a DNA aptamer for cadmium detection based on cationic polymer mediated aggregation of gold nanoparticles[J]. Analyst, 2014, 139(6): 1550-1561. doi: 10.1039/C3AN02117C [56] CHEN Y, LI H H, GAO T, et al. Selection of DNA aptamers for the development of light-up biosensor to detect Pb(II)[J]. Sensors and Actuators B: Chemical, 2018, 254: 214-221. doi: 10.1016/j.snb.2017.07.068 [57] WRZESINSKI J, CIESIOLKA J. Characterization of structure and metal ions specificity of Co2+-binding RNA aptamers[J]. Biochemistry, 2005, 44(16): 6257-6268. doi: 10.1021/bi047397u [58] QU H, CSORDAS A T, WANG J P, et al. Rapid and Label-Free Strategy to Isolate Aptamers for Metal Ions[J]. ACS Nano, 2016, 10(8): 7558-7565. doi: 10.1021/acsnano.6b02558 [59] 王斌, 邓述波, 黄俊, 等. 我国新兴污染物环境风险评价与控制研究进展[J]. 环境化学, 2013, 32(7): 1129-1136. doi: 10.7524/j.issn.0254-6108.2013.07.003 WANG B, DENG S B, HUANG J, et al. Environmental risk assessment and control of emerging contaminants in china[J]. Environmental Chemistry, 2013, 32(7): 1129-1136 (in Chinese). doi: 10.7524/j.issn.0254-6108.2013.07.003
[60] ZHUO Z J, YU Y Y, WANG M L, et al. Recent Advances in SELEX technology and aptamer applications in biomedicine[J]. International Journal of Molecular Sciences, 2017, 18(10): 2142. doi: 10.3390/ijms18102142 [61] 陈慧甜, 孙清, 时国庆, 等. 核酸适配体在环境分析中的应用[J]. 环境化学, 2015, 34(1): 89-96. doi: 10.7524/j.issn.0254-6108.2015.01.2014101401 CHEN H T, SUN Q, SHI G Q, et al. Application of aptamers to environmental analysis[J]. Environmental Chemistry, 2015, 34(1): 89-96 (in Chinese). doi: 10.7524/j.issn.0254-6108.2015.01.2014101401
[62] 孟雪洁, 张瑜, 刘京华, 等. 纳米金-适配体电化学传感器用于环境水样中双酚A检测[J]. 环境化学, 2023, 42(2): 379-387. doi: 10.7524/j.issn.0254-6108.2021102403 MENG X J, ZHANG Y, LIU J H, et al. Gold nanoparticles-aptamer electrochemical sensor for detection of bisphenol A in environmental waters[J]. Environmental Chemistry, 2023, 42(2): 379-387 (in Chinese). doi: 10.7524/j.issn.0254-6108.2021102403
[63] LIU H M, LU A X, FU H L, et al. Affinity capture of aflatoxin B(1) and B(2) by aptamer-functionalized magnetic agarose microspheres prior to their determination by HPLC[J]. Mikrochimica Acta, 2018, 185(7): 326. doi: 10.1007/s00604-018-2849-8 [64] CAI S D, YAN J H, XIONG H J, et al. Investigations on the interface of nucleic acid aptamers and binding targets[J]. Analyst, 2018, 143(22): 5317-5338. doi: 10.1039/C8AN01467A [65] LEI Z, LEI P, GUO J F, et al. Recent advances in nanomaterials-based optical and electrochemical aptasensors for detection of cyanotoxins[J]. Talanta, 2022, 248: 123607. doi: 10.1016/j.talanta.2022.123607 [66] MONDAL B, RAMLAL S, LAVU P S R, et al. A combinatorial systematic evolution of ligands by exponential enrichment method for selection of aptamer against protein targets[J]. Applied Microbiology and Biotechnology, 2015, 99(22): 9791-9803. doi: 10.1007/s00253-015-6858-9 [67] PARK J Y, YANG K A, CHOI Y J, et al. Novel ssDNA aptamer-based fluorescence sensor for perfluorooctanoic acid detection in water[J]. Environment International, 2022, 158: 107000. doi: 10.1016/j.envint.2021.107000 [68] CHEN J H, SHI G, YAN C. Visual test paper for on-site polychlorinated biphenyls detection and its logic gate applications[J]. Analytical Chemistry, 2021, 93(46): 15438-15444. doi: 10.1021/acs.analchem.1c03309 [69] CHEN Y Q, WANG Z M, LIU S Y, et al. A highly sensitive and group-targeting aptasensor for total phthalate determination in the environment[J]. Journal of Hazardous Materials, 2021, 412: 125174. doi: 10.1016/j.jhazmat.2021.125174 [70] 高羽菲, 甄建辉, 赵杰, 等. 比色法适配体传感器在抗生素检测中的研究进展 [J]. 分析实验室, 2023, 1-18. GAO Y F, ZHEN J H, ZHAO J, et al. Advances in antibiotics detection based on colorimetric aptasensors[J]. Chinese Journal of Analysis Laboratory, 2023, 1-18 (in Chinese).
[71] XU G H, WANG C, YU H, et al. Structural basis for high-affinity recognition of aflatoxin B1 by a DNA aptamer[J]. Nucleic Acids Research, 2023, 51(14): 7666-7674. doi: 10.1093/nar/gkad541 [72] MCCONNELL E M, NGUYEN J, LI Y F. Aptamer-based biosensors for environmental monitoring[J]. Frontiers in Chemistry, 2020, 8: 434. doi: 10.3389/fchem.2020.00434 [73] CHANG T J, HE S S, AMINI R, et al. Functional nucleic acids under unusual conditions[J]. ChemBioChem, 2021, 22(14): 2368-2383. doi: 10.1002/cbic.202100087 [74] ACKERMAN C M, MYHRVOLD C, THAKKU S G, et al. Massively multiplexed nucleic acid detection with Cas13[J]. Nature, 2020, 582(7811): 277-282. doi: 10.1038/s41586-020-2279-8 [75] ALI M M, WOLFE M, TRAM K, et al. A DNAzyme‐Based colorimetric paper sensor for helicobacter pylori[J]. Angewandte Chemie International Edition, 2019, 58(29): 9907-9911. doi: 10.1002/anie.201901873 [76] ROTHENBROKER M, MCCONNELL E M, GU J, et al. Selection and Characterization of an RNA-Cleaving DNAzyme Activated by Legionella pneumophila[J]. Angewandte Chemie International Edition, 2021, 60(9): 4782-4788. doi: 10.1002/anie.202012444 [77] ZHOU Q B, ZHANG G X, WU Y P, et al. In Vitro selection of M2+-independent, fast-responding acidic deoxyribozymes for bacterial detection[J]. Journal of the American Chemical Society, 2023, 145(39): 21370-21377. doi: 10.1021/jacs.3c06155 [78] CHANG T J, LI G P, CHANG D R, et al. An RNA‐Cleaving DNAzyme that requires an organic solvent to function[J]. Angewandte Chemie International Edition, 2023, 62(42): e202310941. doi: 10.1002/anie.202310941 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 2081
- HTML全文浏览数: 2081
- PDF下载数: 45
- 施引文献: 0
