


-
我国是世界上畜禽养殖第一大国,畜禽养殖业排放化学需氧量(chemical oxygen demand, COD)、氨氮和总氮的量占农业污染排放总量的比例分别达到了95.2%、76.8%和62.4%[1]。畜禽养殖过程产生大量高氨氮有机废水,已成为水体污染的最主要来源,对这部分废水的有效处理成为畜禽养殖业污染控制的关键[2]。对于畜禽养殖废水的处理通常采用除碳+脱氮的耦合工艺,其中除碳过程通过厌氧消化将有机物转化为甲烷回收,而脱氮过程往往采用生物脱氮方法。传统的生物脱氮技术需要结合自养菌的好氧硝化作用和异养菌的缺氧反硝化作用,但是,当硝化和反硝化在2个独立单元中进行时(如A/O工艺),需要进行混合液回流,通常具有占地面积大和建设投资成本高等缺点。虽然当硝化和反硝化在同一个反应器内(SBR工艺)进行时,能够节省占地和成本,但需要分时段控制曝气和投加碳源,增加了操作复杂性。在处理畜禽养殖废水的厌氧消化液时,一方面,高氨氮质量浓度易对自养硝化产生抑制;另一方面,厌氧消化处理后出水中仍会残留很多有机物,这将进一步抑制自养硝化效果。因此,十分有必要开发更适宜的高氨氮有机废水处理技术。
异养硝化-好氧反硝化(heterotrophic nitrification-aerobic denitrification, HN-AD)菌能够适应高质量浓度的氨氮和有机物,通过异养硝化与好氧反硝化作用的耦合,在好氧条件下能够将废水中的氨氮转化为氮气,实现高效脱氮,整个代谢过程几乎没有亚硝态氮/硝态氮的积累,同时有机物也得到了降解和去除。1984年,ROBERTSON等[3]将首株HN-AD菌Thiosphaera pantotropha从硫氧化脱硝废水处理装置中分离出来。近年来,越来越多的HN-AD菌被发现,它们大多具有世代时间短和耐受性强等优势,对高质量浓度的氨氮和有机物有较好的耐受和处理效果[4]。HN-AD菌Comamonas WXZ-17可耐受817 mg·L−1的氨氮质量浓度,Acinetobacter sp. TN-14能在氨氮质量浓度高达1 200 mg·L−1的环境下生长[5]。HN-AD菌耐受高氨氮的同时能实现对其转化脱除,如Thauera sp. SND5的平均氮去除速率约为2.85 mg·(L·h)−1[6];Bacillus methylotrophicus L7在初始氨氮质量浓度为1 121.2 mg·L−1的条件下,总氮去除速率可达3.8 mg·(L·h)−1[7]。因此,HN-AD技术可以适应高氨氮有机废水的脱氮处理,反应速率高、处理时间短,同时能够在同一个处理单元中在好氧条件下实现有机物和氨氮、总氮的同步去除,降低工艺复杂度,有望为高氨氮有机废水提供一种具有更高效率和更低成本的新技术。目前相关的研究以纯菌HN-AD系统较多,而实际工程中很难做到纯菌环境,那么具有HN-AD功能的污泥驯化就非常重要,是实现技术应用的关键,然而相关的研究仍然较为缺乏。SONG等[8]针对高盐榨菜废水的处理,经过105 d驯化建立了HN-AD混菌系统,COD和总氮去除率分别达到了93.2%和82.4%。但是,上述方法的驯化时间长,系统启动较慢,迫切需要一种在实际应用场景中低成本、快速、有效驯化具有HN-AD功能活性污泥的方法以及有机碳源对系统运行的影响及其优化相关的研究。
因此,本研究考察了在固定C/N比条件下驯化HN-AD活性污泥的方法,并进行了HN-AD效果验证,选择了3种碳源以探究碳源种类和C/N比对系统运行效果的影响,分析了系统中有机物降解和脱氮的动力学特征,揭示了系统中的优势功能菌,以期为实际工程中畜禽养殖废水厌氧消化液等高氨氮有机废水的高效处理提供技术支撑。
-
本研究采用间歇式反应器,材质为有机玻璃,内径为150 mm,主体高170 mm,有效容积为3.0 L。在反应器底部放置曝气盘,外接空气曝气泵,同时,在反应器上方设搅拌桨充分混合活性污泥和废水,以确保溶解氧(DO)分布均匀。反应器中插有pH和DO电极,用于在线监测pH和DO。
实验用水选用模拟配水,以NH4Cl(100~600 mg·L−1, 以氮浓度计)作为氮源,添加KH2PO4(20 mg·L−1, 以磷浓度计)补充磷源,添加微量元素(1 mL·L−1),添加乙酸钠、柠檬酸钠、丁二酸钠的混合碳源(溶液中三者的COD比=1:1:1)作为生物可利用有机碳源(2 000~12 000 mg·L−1,以COD计)。通过调整进水碳源和氮源质量浓度以控制配水的碳氮比。
-
反应器采用序批式运行模式,每个周期分为5个阶段:进水(10 min)、曝气加搅拌运行、沉降(20 min)、出水(10 min)、空闲(1 h)。反应在室温下运行,通过实时监测DO来控制曝气阶段的运行时长,采用蠕动泵进水和排水,出水阶段排水比为0.25。实验装置共运行175 d,设置了系统启动、进水负荷提升和C/N比优化3个阶段。
第Ⅰ阶段:启动HN-AD系统,将北京某污水厂二沉池的活性污泥投入反应器,通过定期排泥(污泥龄12~15 d)排出部分死菌和细胞分泌物,以维持微生物的新陈代谢活性。初始进水COD和氨氮质量浓度分别为2 000 mg·L−1 和100 mg·L−1。根据自养氨氧化过程,每氧化1 g氨氮需要消耗7.14 g碱度(以碳酸钙计),在进水中投加NaHCO3补充至所需碱度。保持C/N比为20,逐步提高进水氨氮质量浓度(每5个周期提升100 mg·L−1)来驯化HN-AD菌群。验证HN-AD系统,当系统启动成功后不再额外补充碱度。第Ⅱ阶段:HN-AD系统的进水氨氮质量浓度进一步提升到300 mg·L−1,待系统运行稳定后,每5个周期提升100 mg·L−1,逐步将进水氨氮提升至600 mg·L−1。第Ⅲ阶段:保持进水氨氮质量浓度为600 mg·L−1,探究不同C/N比(15、20、25)对HN-AD系统脱氮效率的影响,优化出最佳运行条件。
-
从反应器中取出50 mL活性污泥,移至250 mL锥形瓶,随后加入50 mL模拟配水进行实验。设置模拟配水中C/N=20,采用混合碳源(乙酸钠、丁二酸钠和柠檬酸钠的COD比=1:1:1)。模拟配水中进水氨氮质量浓度为100 mg·L−1。在实验组中添加自养硝化抑制剂3,4-二甲基吡唑磷酸盐(C5H8N2·H3O4P,DMPP),于25 ℃、160 r·min−1振荡培养36 h,每间隔一段时间取水样测试氨氮、硝态氮、亚硝态氮的质量浓度。
-
相比于糖类等大分子物质,乙酸钠、柠檬酸钠和丁二酸钠等小分子更容易被HN-AD菌所利用,能够直接参与三羧酸循环代谢过程。考虑到畜禽养殖废水厌氧消化液中常含有小分子挥发性脂肪酸的特点,因此,本研究选用乙酸钠、柠檬酸钠和丁二酸钠作为实验所用碳源,以研究碳源种类的影响。
从反应器中取出50 mL活性污泥,移至250 mL锥形瓶,随后加入50 mL模拟配水(含氨氮200 mg·L−1)进行实验。在模拟配水中,分别采用乙酸钠、丁二酸钠、柠檬酸钠及3种有机物作为混合碳源,以确保锥形瓶内在进水后的初始COD和氨氮质量浓度分别为2 000 mg·L−1和100 mg·L−1,于25 ℃、160 r·min−1振荡培养36 h,每间隔一段时间取水样测试氨氮、硝态氮、亚硝态氮的质量浓度。
-
HN-AD系统启动成功并稳定运行后,在初始有机物和氨氮质量浓度分别为3 000 mg·L−1和150 mg·L−1的单个周期,实时监测COD值、氮质量浓度变化。采用修正的Gompertz模型[9](式(1))对底物去除过程进行拟合,解析底物质量浓度与反应时间的关系。
式中:S为t时刻底物质量浓度,mg·L−1;S0为初始底物质量浓度,mg·L−1;Rm为最大去除速率,mg·(L·h)−1;t0为迟滞时间,h。
-
1)常规水质分析方法。COD值和各种氮化合物的质量浓度用标准方法测定。用纳氏分光光度法在425 nm处测定氨氮,用比色法在540 nm处测定亚硝态氮,用紫外分光光度法在220 nm和275 nm处测量硝态氮。用手持便携式分析仪测量pH和溶解氧(德国WTW Multi 3320)。铵(NH4+-N)由于水解反应会转化为分子态氨,也称为游离氨(free ammonia, FA),FA质量浓度由NH4+-N、pH和温度确定,根据式(2)[10]进行计算。
式中:ρFA为游离氨质量浓度,mg·L−1;
CNH+4-N 为氨氮质量浓度,mg·L−1;pH为溶液的酸碱度;T为温度, ℃。2)微生物群落结构测定。从反应器中取一定量的活性污泥,根据E.Z.N.A.® soil DNA kit(Omega Bio-tek, Norcross,美国)说明书进行微生物群落总DNA抽提,用NanoDrop2000(赛默飞世尔科技,美国)测定DNA的浓度和纯度。使用引物338F(5’-ACTCCTACGGGAGGCAGCAG-3’)和806R(5’-GGACTACHVGGGTWTCTAAT-3’)对16S rRNA基因V3~V4可变区进行PCR扩增,并用琼脂糖凝胶电泳检测PCR结果。使用Illumina MiSeq平台(美吉生物医药科技有限公司,上海)对纯化的聚合酶链反应产物进行测序。
-
在C/N比为20的条件下,通过进水氨氮质量浓度的梯度提升来驯化活性污泥,增加HN-AD菌的相对丰度,进而构建HN-AD脱氮系统,结果如图1所示。结果表明,系统启动初始,进水氨氮质量浓度为100 mg·L−1,逐步提升进水氨氮质量浓度,在1~10周期(进水氨氮≤200 mg·L−1),氨氮去除率(>95%)较高,出水中亚硝态氮质量浓度(<0.1 mg·L−1)较低,但积累了一定质量浓度的硝态氮(>20 mg·L−1),推测系统中发生了以自养硝化为主的硝化过程,此时总氮去除率只有约38%。由图2(a)可见,在系统启动初期(第7周期)的反应过程可以看出,随着氨氧化的进行,亚硝态氮质量浓度先升高后降低,硝态氮质量浓度逐步升高并出现积累。在该周期的反应过程中,游离氨质量浓度在6 h时升至20 mg·L−1,高于抑制亚硝酸盐氧化菌(nitrite oxidizing bacteria, NOB)的FA阈值(0.1~5.0 mg·L−1),亚硝态氮出现积累且质量浓度到达峰值(9 mg·L−1),但随着反应的进行,FA质量浓度迅速降低至5 mg·L−1以下,亚硝态氮积累消失且硝态氮质量浓度不断上升。
由图1可见,在第11周期,将进水氨氮质量浓度提升至300 mg·L−1,对应进水FA达到76.8 mg·L−1,已高于大多数研究报道的自养氨氧化菌(ammonia oxidizing bacteria, AOB)和NOB的抑制阈值。虽然该周期氨氮去除率降低至76%,但出水中不再有硝态氮累积(<2 mg·L−1),总氮去除率达到67.8%,从这一周期开始,系统不再额外补充碱度。当FA质量浓度为20~40 mg·L−1时,自养AOB的氨氧化性能会受到严重影响[11],而HN-AD菌属,如Acinetobacter YB、Bacillus WXZ-8和Zobellella DN-7等,在较高质量浓度FA时(>100 mg·L−1)仍能正常发挥氨氧化功能[12-13]。由此可知,异养硝化菌对FA的耐受能力远高于自养硝化菌,推断此时系统中较高质量浓度的FA抑制了自养AOB,硝化作用主要由HN-AD菌贡献。随着反应器运行,HN-AD系统脱氮性能不断提升。由图2(b)可见,第15周期的氨氮去除率和总氮去除率分别提升至92%和83%,有机物和氨氮实现了同步降解,并且未出现硝态氮和亚硝态氮的积累。在该周期FA质量浓度始终维持在40 mg·L−1以上,且在6 h时达到70.5 mg·L−1,从而较稳定地实现对自养AOB和NOB的抑制,有利于HN-AD菌的富集。当第16周期进水氨氮质量浓度进一步提升至400 mg·L−1,氨氮去除率和总氮去除率依然保持在91%和85%,至此认为HN-AD系统启动成功。
通过投加抑制剂DMPP抑制自养硝化,根据氨氮转化性能验证了系统中发生的主要硝化反应类型。由图3(a)可知,驯化前,投加DMPP实验组氨氮质量浓度基本没有下降,而空白组则正常进行氨氧化过程,这表明抑制剂显著抑制了氨氮的转化。由图3(b)可知,驯化后,投加DMPP实验组与空白组的氨氮质量浓度呈现相似下降趋势,表明氨氧化过程并未受到抑制。综上所述,系统启动成功后,体系中硝化反应的主要类型是异养硝化。
-
选择合适的碳源有利于实现最佳的菌体生长和脱氮性能,可以提高反应效率并缩短反应时间。本研究选用乙酸钠、柠檬酸钠和丁二酸钠作为实验所用碳源,在混菌系统中研究了有机碳源种类的影响。从图4可以看出,用乙酸钠、柠檬酸钠、丁二酸钠作唯一碳源时,48 h的氨氮去除率分别达到了79.5%、83.3%和87.9%;而将上述3种碳源混合使用时,48 h的氨氮去除率达到了96.1%。不同种类碳源的分子构成和氧化还原电位存在差异,因此,HN-AD菌对他们的利用程度各不相同[13]。有研究表明,以丁二酸钠为碳源时,Thauera sp. SND5菌株对氮的去除效果较好[6];利用柠檬酸钠为碳源时,Alcaligenes faecalis C16菌株的脱氮效率最高[14];而对于Paracoccus pantotrophus菌株,乙酸钠是发挥最佳脱氮性能的碳源[15]。因此,在本研究的HN-AD系统中,不同的单一碳源会表现出脱氮性能的差异,而混合碳源可以更好地满足混菌体系中不同种HN-AD菌属的需求,从而使系统具有更好的脱氮性能。由于畜禽养殖废水中存在大量乙酸等小分子有机酸,所以当用HN-AD系统处理该类废水时,有望能够保证较高的脱氮效率。
-
在HN-AD系统采用混合碳源的基础上,进一步研究了系统进水C/N比的影响。结果表明,C/N比对HN-AD菌的脱氮效率有显著影响。
如图5(a)所示,当C/N比为15时,系统的氨氮去除率为80.3%,同时出水中几乎没有硝态氮或亚硝态氮积累;将C/N比提升为20后,HN-AD系统脱氮效率明显上升,随着系统运行,HN-AD系统运行稳定,氨氮去除率提高到了95%;将C/N比提高到25后,脱氮性能有所下降,氨氮去除率降低为90%。总氮的去除率也随着C/N比的优化而发生变化,在C/N比为20时,总氮去除率达到了89.6%,之后继续提升C/N比并不能显著提升总氮去除率。不同HN-AD菌的最优C/N比不同,Aliidiomarina在C/N比为9时,氨氮去除率达到93.7%[16];Thauera sp.TN9在C/N比为22时氮去除效率最高,达到99.2%[17];Paracoccus versutus LYM在C/N比为20时,氮去除率达到97.09%[18]。一方面,C/N比过低会导致碳源不足,使得细胞生长受限以及缺乏电子供体,进而导致脱氮效率下降;另一方面,C/N比过高可在一定程度上抑制脱氮效果[19]。因此,合适的C/N比对于HN-AD系统的低耗高效运行至关重要,后续可将C/N比设为20~25进一步优化,以确定HN-AD系统的最佳C/N比。
如图5(b)所示,随着进水C/N比的提高,系统出水的COD值不断增加,由于出水中没有检测到进水所用碳源,因此推测出水中的COD来源于微生物的细胞分泌物。对出水进行三维荧光测试,结果显示较明显的荧光峰(Ex/Em=260~300 nm/300~370 nm)为色氨酸荧光蛋白峰,同时,另一个较明显的荧光峰(Ex/Em=330~370 nm/400~475 nm)为类腐殖酸的峰,表明蛋白质和腐殖酸是出水中有机物的组成部分。这可能是由于高C、N质量浓度环境刺激了微生物产生大量细胞代谢产物,形成高浓度的胞外聚合物(extracellular polymeric substances, EPS),EPS主要由多糖和蛋白质组成,可分为溶解态和结合态2种形态,其中大部分溶解态EPS可以随出水排出系统[20]。WANG等[21]研究表明,在微生物处于极端的生存环境时,EPS中色氨酸和芳香类蛋白荧光强度会显著增强。
本研究进一步对出水中的蛋白质和多糖进行了测定,结果如图6(b)所示,发现其含量随着C/N比提高而不断增加,换算成COD当量后发现,蛋白质和多糖分别占出水总COD的60%~70%。当对高碳质量浓度有机废水进行脱氮处理时,HN-AD菌的繁殖速度远远快于自养硝化菌,但其生长过程产生的大量溶解态EPS很可能导致出水有机物超标,后续可以通过增加膜组件构成膜-生物反应器对这些大分子物质截留,从而保证出水水质达到排放标准。
-
HN-AD系统具有同步除碳脱氮的特性,通过修正的Gompertz模型[9]来进一步解析底物去除过程。针对典型周期,分别对COD和氨氮质量浓度随时间的变化进行动力学拟合,结果如图7所示。COD的最大去除速率为174.1 mg·(L·h)−1(R2=0.992),氨氮的最大去除速率为8.66 mg·(L·h)−1 (R2=0.999)。已有研究[22]表明,大部分HN-AD纯菌的氨氮去除速率为3~8 mg·(L·h)−1,而本研究所构建HN-AD系统显示出比纯菌体系更高的氨氮去除速率。此外,对典型周期各时间点的COD和氨氮质量浓度进行相关性拟合。如图7(c)所示,反应过程中两者呈现出较好的相关性(R2=0.997),从而间接证明了HN-AD过程氨氮转化需要碳源,有助于实现碳氮协同降解。上述拟合结果有助于深入了解系统性能,并优化HN-AD系统在实际应用时的设计和运行参数,以实现高质量浓度含氮有机废水处理过程高效稳定的除碳脱氮。
-
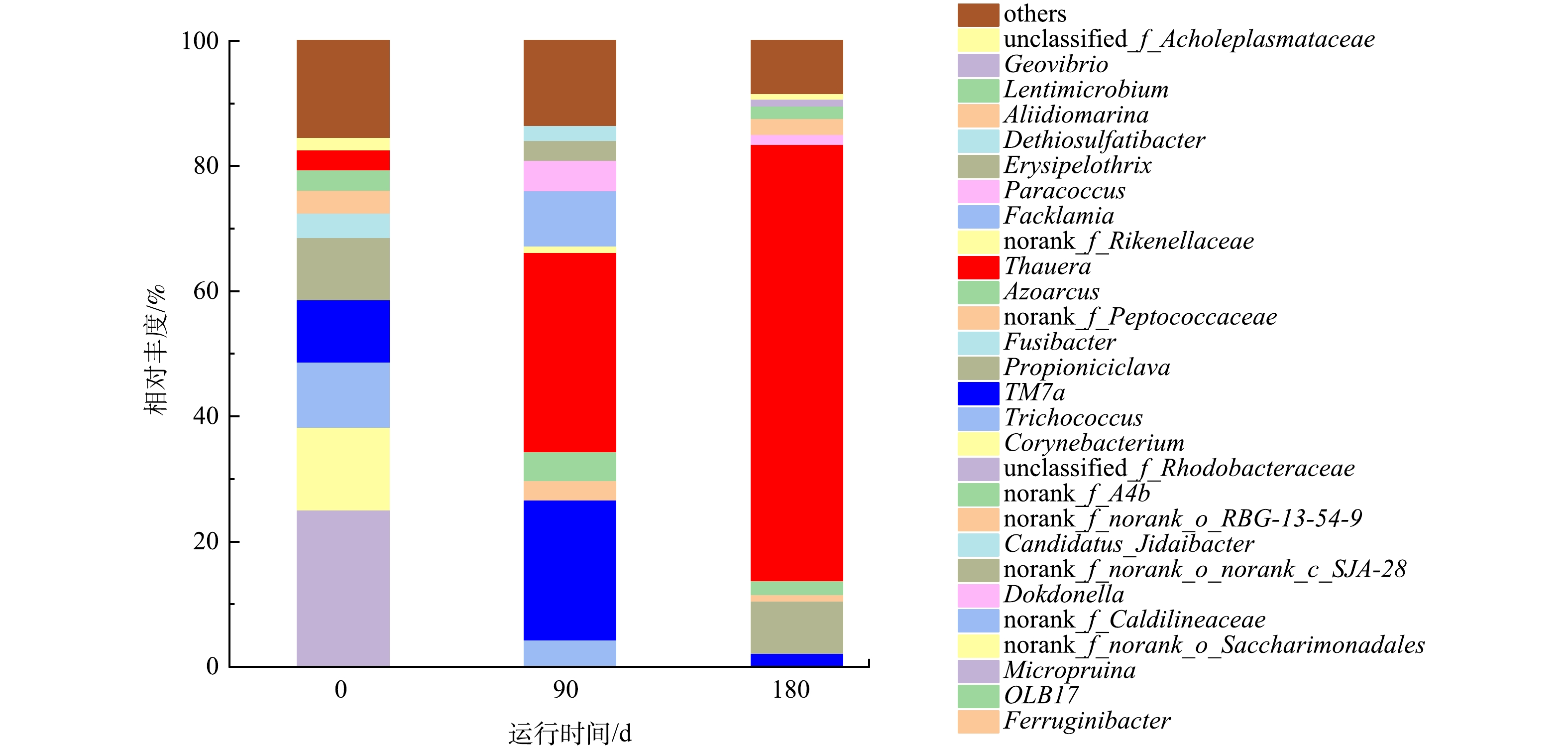
在属水平上的污泥物种组成分析结果如图8所示,HN-AD系统中的优势菌为Thauera(69.7%),而自养硝化菌的丰度较低(<1%)。已有研究[6,17]表明,Thauera细菌可以参与氮循环过程,具有异养硝化-好氧反硝化功能,通常存在于极端环境中,对高COD和高氨氮环境具有较强的适应能力,Thauera细菌具有降解有机物的能力,特别是一些具有环境污染潜力的有机污染物,如芳香烃类化合物和氯化有机化合物,其代谢能力可以用于生物修复和废水处理等环境应用。相对丰度第2高的Propioniciclava(8.3%),该菌属是一种潜在聚磷菌,能够在厌氧阶段完成聚磷代谢[23]。属水平微生物群落分析结果表明,在驯化过程中,Thauera相对丰度显著增加,成为主导性的菌种。以上结果对于深入了解HN-AD系统的微生物群落动态和功能特性具有重要意义。
-
1)通过逐渐提升进水COD和氨氮质量浓度的方法可以快速实现HN-AD污泥的驯化,并通过单个周期的COD和氨氮质量浓度监测证实了HN-AD功能的启动。
2)有机碳源对系统的脱氮效果有重要影响,柠檬酸钠、乙酸钠和丁二酸钠的混合碳源更有利于异养硝化脱氮,当C/N比为20时,HN-AD系统的脱氮效率最高。采用Gompertz动力学模型分别拟合了氨氮和COD底物的去除过程,模型相关系数R2均在0.9以上,证实了氨氮去除和有机物去除的相关性。
3)在优化条件下,HN-AD系统实现了95%的氨氮去除率、89.6%的总氮去除效率和92%的COD去除率,这表明该工艺可同步实现高效脱氮和有机物的去除。高通量分析结果表明,Thauera相对丰度显著增加,并成为了HN-AD系统中的优势菌种。
有机碳源对异养硝化-好氧反硝化生物脱氮的影响及其优化
Effect of organic carbon on the biological nitrogen removal through heterotrophic nitrification-aerobic denitrification and its optimization
-
摘要: 异养硝化-好氧反硝化(HN-AD)是一种新型的高氨氮有机废水生物脱氮技术,目前在纯菌系统中的研究较多而更适合真实应用场景的混菌系统相关研究则较为缺乏,限制了技术的推广应用。本研究首先开发了具有HN-AD功能的活性污泥驯化方法并进行了验证,然后研究了碳源种类和C/N比的影响并进行了优化。结果表明,HN-AD污泥被成功驯化,自养硝化菌的丰度占比较低(<1%),在最佳脱氮效果运行条件下,Thauera逐渐富集并成为HN-AD系统的优势功能菌,其丰度占比高达69.7%。有机碳源对HN-AD脱氮系统具有重要影响,采用混合碳源(柠檬酸钠、乙酸钠、丁二酸钠,溶液中三者的COD比=1:1:1)并在C/N=20时实现了95%的氨氮去除率和89.6%的总氮去除效率,同时COD去除率高达92%,可实现高效脱氮和有机物去除。
-
关键词:
- 异养硝化-好氧反硝化 /
- 碳氮比 /
- 氨转化率 /
- 微生物群落 /
- 反应动力学
Abstract: Heterotrophic nitrification-aerobic denitrification(HN-AD) is a novel biological nitrogen removal technology for high-ammonium organic wastewater. Currently, related studies mainly focus on the HN-AD in pure culture using single bacteria, while studies on mixed bacteria systems suitable for realistic application scenario are relatively scarce, limiting the application of this technology. This study developed and validated an activated sludge domestication method with HN-AD function. Subsequently, the impacts of carbon source types and C/N ratio were investigated and optimized. Results showed that the HN-AD sludge was successfully domesticated, with a low proportion of autotrophic nitrifying bacteria (<1%). Under the optimal condition of nitrogen removal, Thauera was gradually enriched and became the dominant functional bacterium in the HN-AD system, accounting for a high relative abundance of 69.7%. Organic carbon sources had a significant impact on the HN-AD nitrogen removal system. At a C/N ratio of 20 and the mixed carbon sources (sodium citrate, sodium acetate, sodium succinate, the related COD ratio of these carbon sources was 1:1:1) in the HN-AD system, the removal efficiency of ammonium and total nitrogen could reach 95% and 89.6%, respectively, and COD removal efficiency could reach 92%, which could achieve effective total nitrogen and organic matter removal. -
类金属砷是自然界(大气、土壤、水等环境介质)中普遍存在的元素,对人体有一定的毒性作用,易致癌[1]。自然条件下砷的释放很少达到对人体有害的程度,砷的释放、迁移和污染大多是由人类活动造成的,包括含砷矿物的开采、冶炼以及含砷物质的使用[2]。在我国,有超过15万km2的地区,近2 000万人生活在砷含量超过10 μg·L−1(WTO饮用水标准)水高砷污染区中;全世界约有2×109人受到高砷暴露的威胁[3]。因此,开发合理、高效的砷污染治理技术对保护人类健康、保障饮水安全具有重要意义。
砷在污染水体中主要形态为+3价(还原环境,亚砷酸根(AsO33−))和+5价(氧化环境,砷酸根(AsO43−)),其中,As(Ⅲ)毒性约为As(V)的25~60倍[4]。依据砷的不同形态及包括吸附、氧化-还原和混凝-沉淀等去除原理,目前治理砷污染水体的方法主要有离子交换和膜分离技术[5-6]、混凝沉淀法[7]、吸附法[8]和生物修复技术[9]。其中,离子交换法和膜分离技术是最有效的去除方法,但因使用价格昂贵,很难推广应用;混凝沉淀和吸附法,作为传统的水净化方法,需要消耗大量的化学制品,且因砷污染水体中含有复杂的有机物和竞争离子存在,使去除效果成为一项重大挑战;生物修复技术因其应用成本低和环境友好性等优势受到广泛关注,但因修复效率低,处理时间长和适宜生长环境控制较难的问题而受到很大限制。
近年来,基于铁碳微电解材料产生微原电池反应去除水体污染物的处理方法,受到越来越多研究者的关注[10]。该方法利用铁碳微电解材料兼具催化氧化、絮凝沉淀和氧化还原等污染物去除原理,具有效果稳定,应用范围广,操作方便等优点[11-12]。但传统铁碳微电解材料主要是将还原铁粉或废铁屑、工业碳粉和一定的粘结剂按照一定比例混匀由固定模具压制成型后通过高温烧结制备完成[13],需消耗大量碳粉和粘结剂等工业制成品,提升了制备成本。并且工业粘结剂在高温焙烧下易板结,不利于铁碳材料烧制过程中孔隙结构的产生,在砷污染水体的去除过程中,降低了Fe-C反应电极与含砷废水的接触面积,阻碍微原电池反应过程中的电子转移,去除效率不佳[14-15]。
鉴于此,在笔者所在团队前期研究基础上[16],提出了一种利用人工湿地芦苇制备得到的植物粉末取代工业碳粉,膨润土作为支撑剂和粘结剂,与还原铁粉(Fe0)按一定添加比例混匀,在一次性梯度升温条件下烧制成型的制备工艺。此外,有研究表明,植物基生物质材料在碳化后,具有较好稳定性和电化学性能,与还原性铁Fe0形成持续的Fe-C微原电池反应,而被当做良好的阴极材料[17]。但在一定的烧制温度下,碳基材料的性质如电子传递、稳定性和氧化还原能力,在很大程度上由其生物质前驱体的不同组分(半纤维素、纤维素和木质素)决定[18]。因此,本研究采用响应面法探究芦苇叶和芦苇秆植物基铁碳微电解材料最优制备条件,并且探讨了2种植物基中各主要生物质组分与制备条件相互作用对铁碳微电解材料除As(Ⅲ)效率与机理的影响,以期为铁碳微电解材料优化制备及其应用于含砷废水的处理提供参考。
1. 材料与方法
1.1 试剂与仪器
还原铁粉(Fe0)、亚砷酸钠(NaAsO2)、氢氧化钠(NaOH)、盐酸(HCl)等均为分析纯(国药集团化学试剂有限公司);纤维素、半纤维素和木质素(阿拉丁试剂(上海)有限公司);钙基膨润土(辽宁省锦州市锦州金泰膨润土有限公司);实验用水均为去离子水。
实验过程所使用的仪器:DFT-100A型小型粉碎机(浙江大德仪器有限公司);TSQ-280型恒温振荡器(上海精宏有限公司);SXW-1200 ℃型马弗炉(上海实研有限公司)。
分析测试方法:铁碳微电解材料中铁含量采用《直接还原铁-金属铁含量的测定-三氯化铁分解重铬酸钾滴定法》(GB/T 38812.2-2020)方法进行测定[19];植物基原材料芦苇秆与芦苇叶中碳元素含量采用元素分析仪(Elementar Vario EL III,德国Elementar公司)测定;溶液中砷浓度采用电感耦合等离子体原子发射光谱(ICP-OES,Optima,美国PerkinElmer公司)测定;铁碳微电解材料中碳化铁(Fe3C)采用X射线衍射仪(XRD,D8 Advance,布鲁克AXS公司)进行物相分析[20];铁碳微电解材料的形貌采用场发射扫描电子显微镜(蔡司Gemini360/560场发射扫描电镜,德国蔡司公司)进行分析;铁碳微电解材料及纤维素、半纤维素及木质素在氮气保护下采用热重-差热综合热分析仪(TGA,Diamond TG/DTA,美国PerkinElmer公司)进行热稳定性分析(TG-DTG);铁碳微电解材料比表面积及孔隙度采用比表面积和孔隙度分析仪(TriStar II Plus Series,美国麦克仪器公司)测试。
1.2 植物基铁碳微电解材料的制备
选用西南科技大学流域生态环境与水污染控制实训中心人工湿地的芦苇为生物质原料,将其按芦苇秆和芦苇叶进行分选,去除杂质并洗净,置于烘箱中在80 ℃条件下烘至恒重后利用破碎机进行破碎,过200目筛,收集于棕色瓶中保存待用。将还原铁粉、植物粉末样品、膨润土、水按一定配比混匀加入造粒机中制备成直径大小为1~1.5 cm的球形颗粒,然后移入马弗炉烧制。烧制步骤为:先由室温升至设定碳化温度并恒温一段时间,进行绝氧热解碳化处理,在碳化过程结束后,继续升温至设定焙烧温度并恒温一段时间,进行绝氧焙烧成型处理,最后自然冷却得到植物基铁碳微电解材料。
2. 结果与讨论
2.1 响应面法研究优化制备条件
制备植物基铁碳微电解材料的主要影响因子包括铁碳摩尔比(Fe/C)、碳化温度和焙烧温度。第一步根据单因素预备实验,确定各因子最佳水平的适当范围,后采用Box-Behnken模型进行实验设计。
1)单因素预备实验。取100 mL 10 mg·L−1的NaAsO2溶液于250 mL锥形瓶中,调节pH至3后,加入5 g植物基铁碳微电解材料,放入恒温摇床(25 ℃,100 r·min−1),避光振荡,计算反应48 h时间下的As(Ⅲ)去除率。铁碳微电解材料中砷去除率根据式(1)进行计算。
R=(A0−AiA0)×100% (1) 式中:R表示砷脱除率,%;A0表示铁碳微电解材料处理前溶液中砷的质量浓度,mg·L−1;Ai表示铁碳微电解材料处理后溶液中砷的质量浓度,mg·L−1。
铁碳摩尔比(Fe/C)最适条件范围。在铁碳微电解原材料配比中,铁与植物材料的添加量直接影响除砷过程中的“Fe-C原电池对”数量。根据芦苇秆与芦苇叶中的碳含量(表1),通过式(2)计算得到还原性铁添加量。
表 1 湿地植物基材料的理化性质(基于干基计算)Table 1. Physicochemical properties of plant - based materials in wetland样品名称 元素组成% 原子比 灰分% C H N O S H/C O/C (O+N/C) 芦苇叶 39.42 3.65 1.05 49.49 0.12 1.11 0.94 0.96 8.37 芦苇秆 44.64 4.42 1.84 44.93 0.09 1.19 0.75 0.79 7.76 注:O%=100%-(C%-N%+H%+S%+灰分%) | Show Table DownLoad:
CSV
DownLoad:
CSV
M1=56PM012 (2) 式中:M0为植物粉末质量,g;M1为还原铁粉(Fe0)质量,g;P为植物粉末中碳元素质量分数。
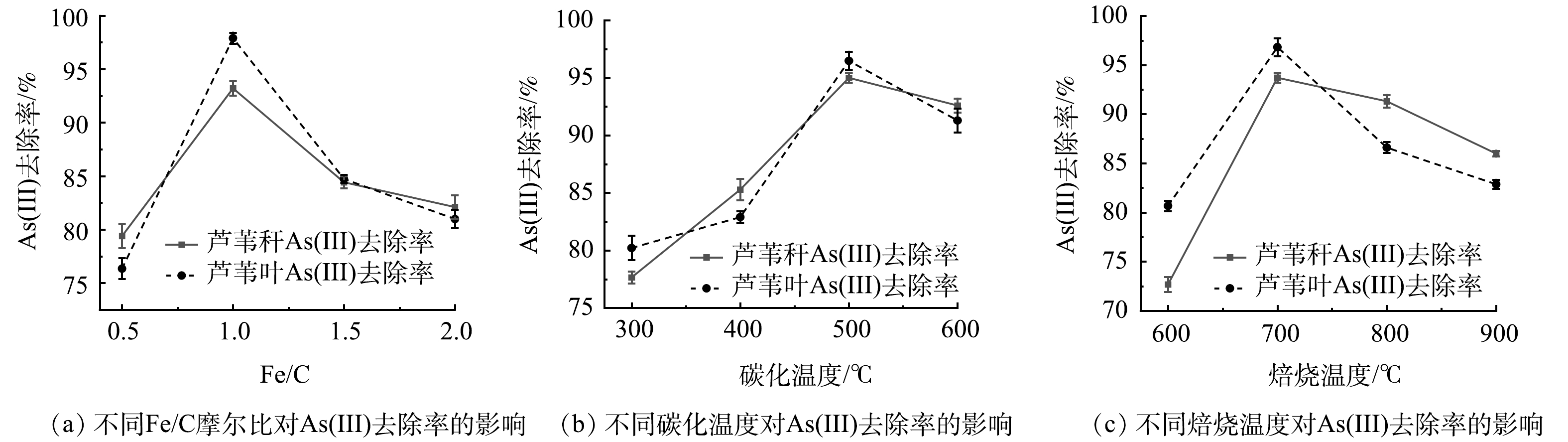
当碳化温度为500 ℃,恒温120 min,焙烧温度为700 ℃,恒温60 min时,不同Fe/C摩尔比条件下As(Ⅲ)的去除率如图1(a)所示。由图1(a)可知,As(Ⅲ)去除率随着Fe/C摩尔比的增加呈现先增加后下降的趋势,在Fe/C摩尔比为1时As(Ⅲ)去除效果最好。因此,选择Fe/C摩尔比为1进行后续实验。
当Fe/C摩尔比为1时,在碳化恒温120 min以及焙烧温度700 ℃、恒温60 min条件下,制得不同碳化温度条件下的铁碳微电解材料,As(Ⅲ)的去除率如图1(b)所示。由图1(b)可知,As(Ⅲ)去除率随着碳化温度的增加呈现逐渐增加再降低的趋势,在500 ℃去除效果最好。因此,选择碳化温度为500 ℃进行后实验。
当在Fe/C摩尔比为1,碳化温度为500 ℃时,在碳化恒温120 min和焙烧恒温60 min条件下,制得不同焙烧温度条件下的铁碳微电解材料,As(Ⅲ)的去除率如图1(c)所示。由图1(c)可知,以芦苇秆和芦苇叶为植物基材料制备的铁碳微电解材料对As(Ⅲ)去除率随着焙烧温度的升高,呈现先升后降的趋势,且在700 ℃时As(Ⅲ)的去除率最高。
2)响应面Box-Behnken设计。在前期单因素预备实验得到的各因子最佳水平的适当范围,运用Design Expert 11.1.0.1软件中的中心复合设计模块(Box-Behnken,BBD)[21]以3个因素作为自变量,砷去除率为响应值设计得到如表2所示的3因素3水平的响应曲面实验。
表 2 Box-Behnken 设计实验因素与水平Table 2. Factors and levels in Box-Behnken design水平 因素 A(铁碳摩尔比(Fe/C)) B(碳化温度/ ℃) C(焙烧温度/ ℃) −1(低水平) 0.8 450 600 0(中水平) 1.0 500 700 1(高水平) 1.2 550 800 | Show TableDownLoad:
CSV
在响应曲面交互作用实验中,实验影响因素(自变量)与实验指数(因变量)之间的回归关系可以通过响应面方法(RSM)[22]进行分析,这种回归既可能是曲线关系,也可能是曲面关系,如式(3)所示。
Y=b0+b1A+b2B+b3C+b12AB+b13AC+b23BC+b11A2+b22B2+33C2 (3) 式中:bi是线性效应;bii是第二次效应;bij是相互作用效应;A为铁炭摩尔比;B为碳化温度;C为焙烧温度。
2.2 响应面模型优化设计结果及模型检验
根据Box-Behnken实验设计制备得到的铁碳微电解材料对As(Ⅲ)去除效率如表3所示。
表 3 Box-Bennken响应面实验设计及结果Table 3. Box-Behnken response surface experiment design and results实验编号 因素 As(Ⅲ)去除率(芦苇秆)/% As(Ⅲ)去除率(芦苇叶)/% A B C 1 0.8 450 700 85.47 90.71 2 1.2 450 700 91.86 94.02 3 0.8 550 700 90.83 92.26 `4 1.2 550 700 95.36 96.26 5 0.8 500 600 70.36 74.71 6 1.2 500 600 76.39 78.44 7 0.8 500 800 96.49 87.78 8 1.2 500 800 99.26 93.26 9 1 450 600 70.18 72.43 10 1 550 600 76.68 80.36 11 1 450 800 96.14 93.25 12 1 550 800 97.48 87.58 13 1 500 700 95.53 98.33 14 1 500 700 96.05 98.36 15 1 500 700 95.81 99.12 16 1 500 700 96.42 98.92 17 1 500 700 96.63 98.56 | Show TableDownLoad:
CSV
将表3中的总砷去除率输入到Design Expert 11.1.0.1软件中进行数据分析和多元二次回归拟合,得到依照编码因数的二次响应曲面方程如式(4)和式(5)所示,实际因素的二次响应曲面方程如式(6)和式(7)所示。
Y(芦苇杆)=96.09+2.5A+2.09B+12.01C−0.47AB−0.74AC−1.29BC−2.31A2−2.89B2−8.07C2 (4) Y(芦苇叶)=96.66+2.06A+0.76B+6.99C+0.17AB+0.44AC−3.4BC−2.6A2−2.74B2−12.51C2 (5) Y(芦苇杆)=−903.68+177.36A+1.43B+1.42C−0.047AB−0.037AC−0.00026BC−57.85A2−0.00116B2−0.000807C2 (6) Y(芦苇叶)=−1134.61+116.46A+1.57B+2.14C+0.0173AB+0.0219AC−0.00068BC−65.04A2−0.0011B2−0.00125C2 (7) 为说明响应面模型及回归方程的有效性,对回归方程的可行性及相关性进行分析,得到分析结果如表4所示。R2均在95%置信区间内,同时校正系数与预测系数差值(Radj-RPred)均小于0.2,表明实测值与预测值之间偏差较小,拟合度好,可信度高;失拟影响因素项结果均不显著(P>0.05),代表失拟因素对于纯粹的误差可忽略不计,无失拟因数存在,3个自变量与响应值之间存在显著的相关性,误差小;精密度均大于4(一般认为较好拟合模型的精密度需大于4[23]),说明可实用性高,真实实验结果可用上述回归方程代替分析。同时从三因素间交互作用的相关系数可以看出:铁碳摩尔比(A)与焙烧温度(C)、碳化温度(B)与焙烧温度(C)之间的交互影响大于铁碳摩尔比(A)与碳化温度(B)间的交互作用,并表现为显著水平(P<0.05)。因此,本研究选择AC与BC因素之间的交互影响探究不同植物基生物质组分对铁碳微电解材料的除砷效率的影响。
表 4 回归方程可行性分析Table 4. Feasibility analysis of regression equation模型项 芦苇秆 芦苇叶 F值 P值 F值 P值 模型 582.18 <0.000 1 777.89 <0.000 1 A-Fe/C 163.52 <0.000 1 193.40 <0.000 1 B-碳化温度 113.79 0.001 7 25.94 0.001 4 C-焙烧温度 3 764.79 <0.000 1 2 216.75 <0.000 1 AB 2.82 0.136 8 0.674 8 0.438 5 AC 7.15 0.031 8 4.34 0.045 7 BC 21.73 0.002 3 262.14 <0.000 1 A2 73.59 <0.000 1 161.55 <0.000 1 B2 115.10 <0.000 1 179.73 <0.000 1 C2 895.90 <0.000 1 3 735.07 <0.000 1 失拟项 2.27 0.222 7 2.04 0.250 9 R2 0.998 7 0.999 0 Radj 0.997 0 0.997 7 RPred 0.985 8 0.989 7 精密度 70.043 1 81.958 7 注:P <0.05表示差异显著。 | Show TableDownLoad:
CSV
2.3 不同生物质组分对最适制备温度的影响
由响应面分析得到碳化温度和焙烧温度2个因素交互作用对总砷去除率的影响如图2和图3所示。在等高线图中,当中心图形呈现椭圆形时,交互作用强;而接近圆形时,交互作用弱。在响应曲面中,响应面坡度越大,自变量条件的改变对响应值的影响越大,反之影响越小。而当两者图形中的颜色越深代表响应值越大,效果越明显[24]。
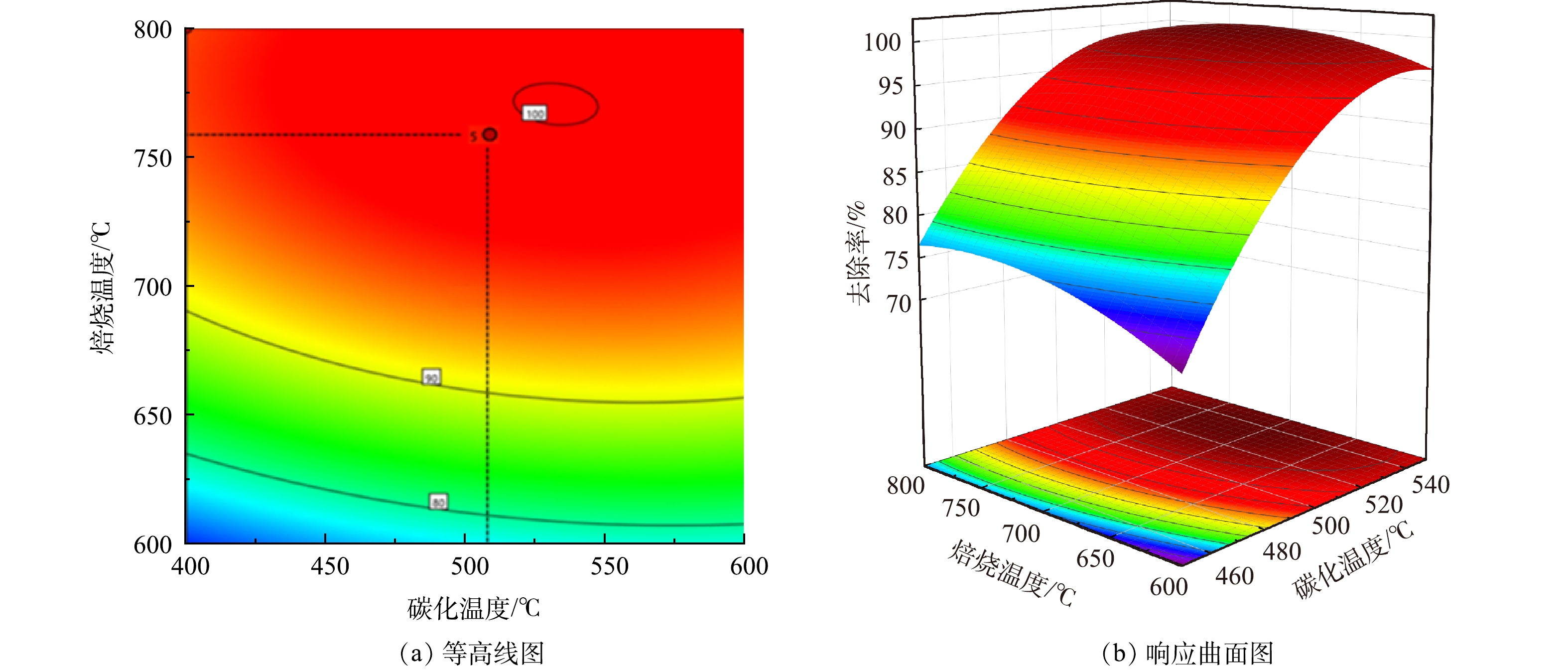 图 2 芦苇秆植物基碳化温度与焙烧温度交互作用的等高线与响应面图Figure 2. Contour line and response surface diagram of interaction between carbonization temperature and roasting temperature of reed rod
图 2 芦苇秆植物基碳化温度与焙烧温度交互作用的等高线与响应面图Figure 2. Contour line and response surface diagram of interaction between carbonization temperature and roasting temperature of reed rod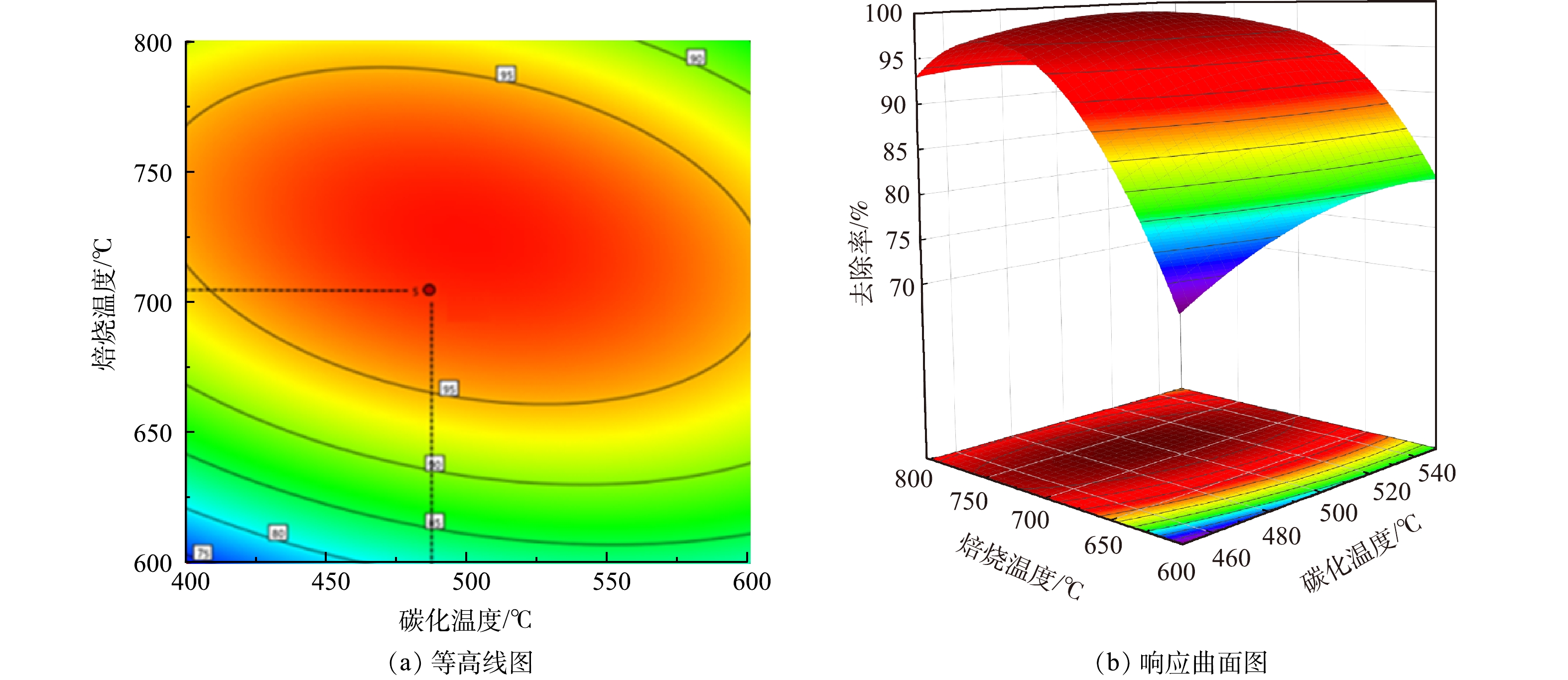 图 3 芦苇叶植物基碳化温度与焙烧温度交互作用的等高线与响应面图Figure 3. Contour line and response surface diagram of interaction between carbonization temperature and roasting temperature of reed leaves
图 3 芦苇叶植物基碳化温度与焙烧温度交互作用的等高线与响应面图Figure 3. Contour line and response surface diagram of interaction between carbonization temperature and roasting temperature of reed leaves由图2、图3中的等高线图看出,碳化温度与As(Ⅲ)的去除率呈正相关,焙烧温度与As(Ⅲ)去除率呈现先增后减的趋势,这与单因素实验结果一致。同时由等高线图也可以看出,沿C因素向峰值移动的等高线密度明显高于沿B因素向峰值移动的等高线密度,表明焙烧温度对响应值的贡献最大,这与方差分析得到的结果一致。响应面坡度陡峭,说明碳化温度与焙烧温度具有显著的交互作用,对As(Ⅲ)的去除影响较大。
作为植物基铁碳微电解阴极材料与Fe0在水溶液中形成微原电池,发生原电池反应,其生物质组分主要由木质素、纤维素和半纤维素三种主要有机物构成,通过非共价键相互交联在一起构成植物骨架。在隔氧条件下进行梯度升温,会生成生物炭、生物油以及不可冷凝气体。生物炭具有稳定的芳香化、脂肪族链状结构及丰富的孔隙分布,具有良好的稳定性和吸附性[25],能提高植物基铁碳微电解材料对As(Ⅲ)的去除。生物炭产量、表面官能团的种类和数量以及碳化方式与温度紧密相关。目前,主要碳化方式有慢速热解、高温气化和水热碳化法[26]。与其它热解方法相比,慢速热解因其固相收率较高,气相产物释放较慢且均匀,更适于植物基材料碳化。在隔氧梯度升温过程中,植物基碳化为生物炭中碳含量增多,氧、氢及灰分比例下降。温度越高,碳化程度越高,生物炭越稳定,但生物炭产量与烧制温度成非线性关系,主要是由于不同湿地植物生物质组分中纤维素、半纤维素和木质素含量和稳定性的差异性造成的[27-28]。通过热重分析仪测试分析得到纤维素、半纤维素和木质素热稳定性如图4所示。
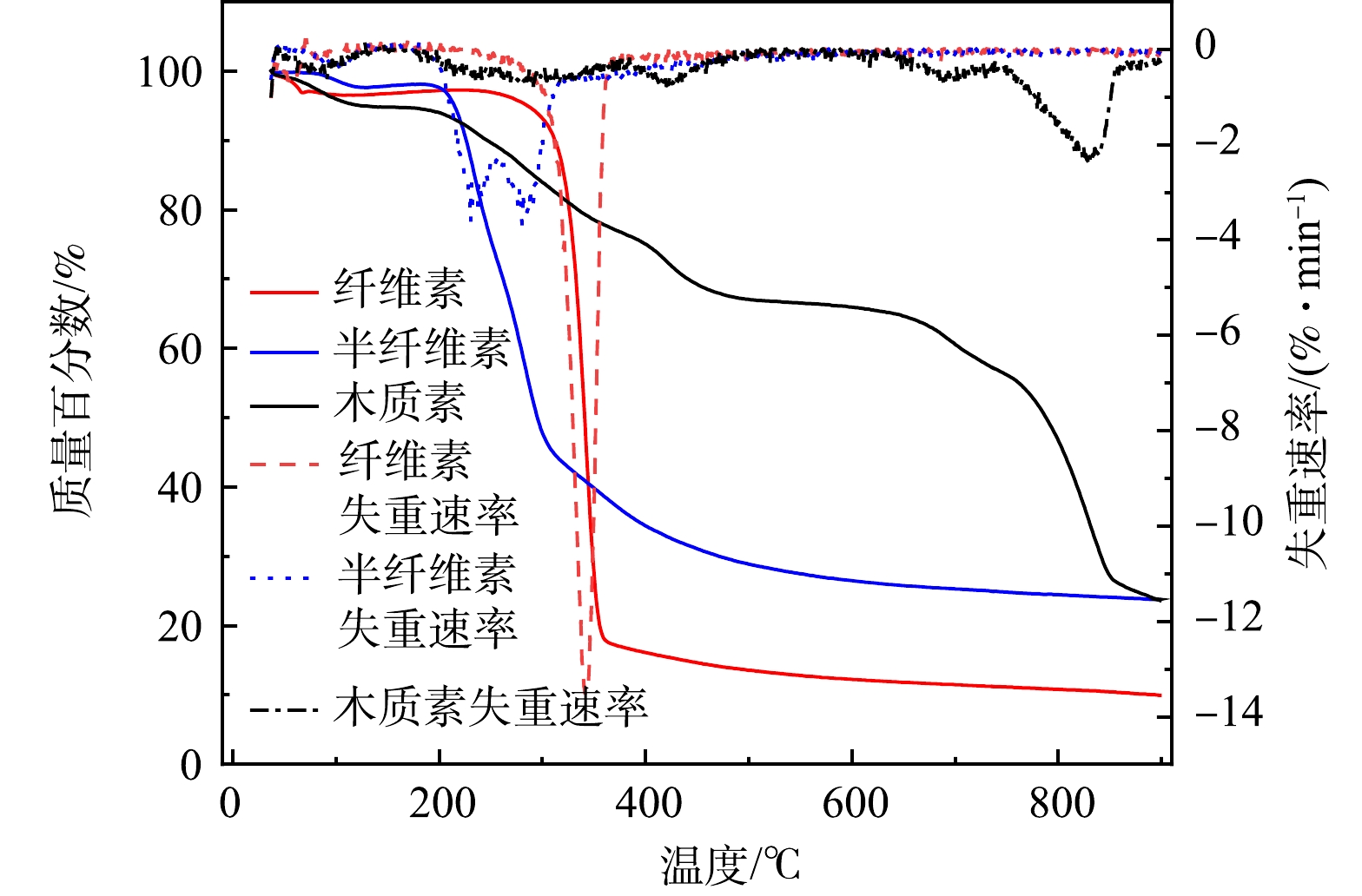 图 4 纤维素、半纤维素及木质素热解稳定性分析Figure 4. Pyrolysis stability of cellulose, hemicellulose and lignin
图 4 纤维素、半纤维素及木质素热解稳定性分析Figure 4. Pyrolysis stability of cellulose, hemicellulose and lignin由图4中可以看出,当碳化温度超过400 ℃时,大部分半纤维素和纤维素被热解。木质素因高稳定性而最难分解,分解温度范围跨度大(160~900 ℃),能在超过700 ℃热解条件下使更多的碳基结构被保留了下来,成为植物基碳基成分中最主要的碳元素来源[29]。
芦苇叶与芦苇秆植物基铁碳微电解材料的微商热重分析(DGT)与失重速率(TG)曲线如图5所示。由图5可以看出,在100 ℃左右时开始出现由铁碳微电解材料水分蒸发所导致的的失重峰。随着梯度升温至400 ℃左右时,植物基铁碳材料出现了不同程度的失重,其中芦苇叶植物基铁碳微电解材料的失重大于芦苇秆植物基铁碳微电解材料(△m1>△m3)。其原因可能是前者纤维素与半纤维素含量大于后者,在较低温度时更容易热解成生物炭,质量损失较快。但随着温度梯度的继续升高伴随着木质素的大量热解,含有更多木质素组分的芦苇秆植物基铁碳微电解材料迅速碳化,质量损失加快(△m2>△m4)。
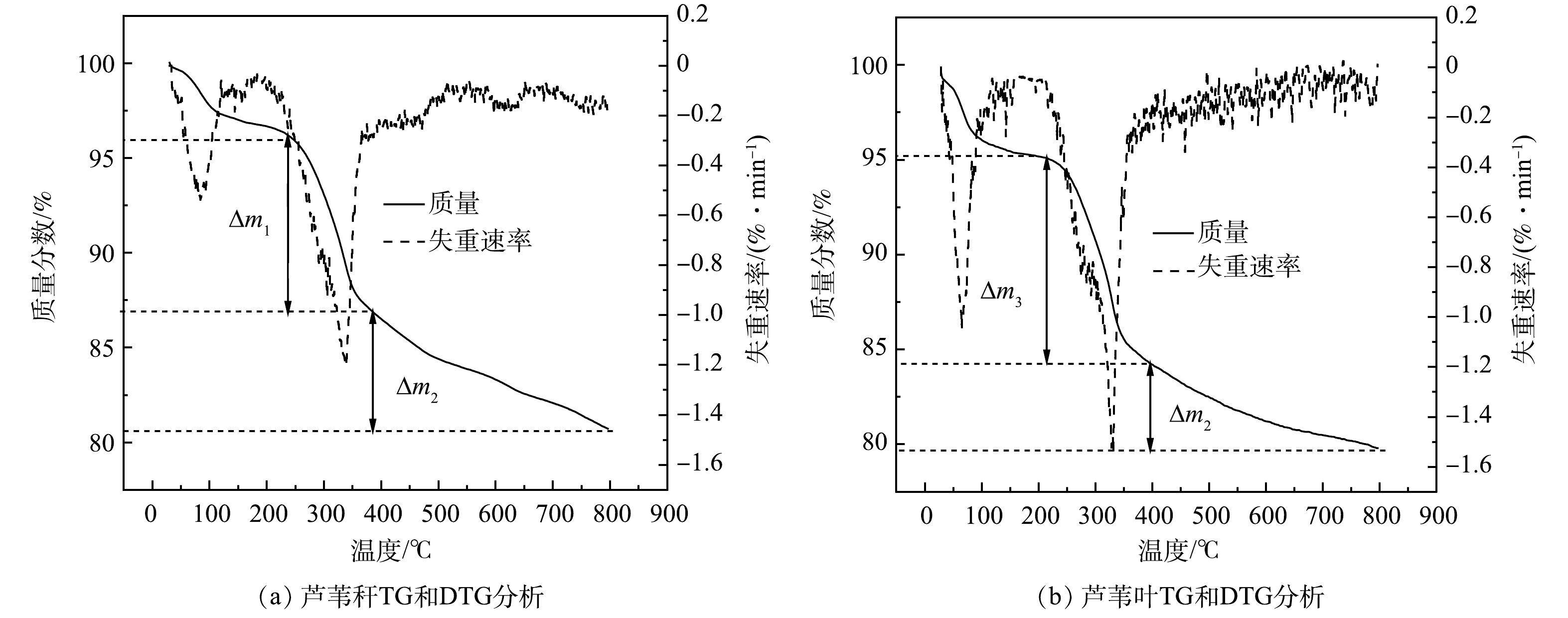 图 5 芦苇秆、芦苇叶植物基铁碳微电解材料热重(TG-DTG)分析Figure 5. Thermogravimetric (TG-DTG) analysis of reed rod and Reed leaf plant-based Fe-C microelectrolysis material
图 5 芦苇秆、芦苇叶植物基铁碳微电解材料热重(TG-DTG)分析Figure 5. Thermogravimetric (TG-DTG) analysis of reed rod and Reed leaf plant-based Fe-C microelectrolysis material因此,在相同Fe/C摩尔比和低于700 ℃焙烧温度下,芦苇叶植物基铁碳微电解材料对As(Ⅲ)去除率高于芦苇秆植物基铁碳微电解材料。分析认为,在低于700 ℃焙烧温度时,芦苇秆植物基有机成分中含有大量木质素,难以被完全碳化形成稳定的碳基材料,所以铁碳材料中有效碳元素比例较低;而芦苇叶植物基拥有大量能在较低温度下碳化的纤维素与半纤维素,碳化充分,有效碳元素含量多,能与Fe0形成大量的Fe0-C原电池参与反应,因此As(Ⅲ)去除率高。当焙烧温度超过700 ℃时,芦苇秆植物基中未被完全碳化的木质素被二次碳化,有效碳元素含量增加,Fe0-C原电池数量上升,As(Ⅲ)去除率提高。而芦苇叶植物基在第一次碳化过程中形成的碳基成分在二次碳化过程中灰分质量分数增加,有效碳元素降低,Fe0-C原电池数量下降,导致去除率下降。
2.4 不同生物质组成对还原铁粉(Fe0)投加量的影响
Fe/C摩尔比和焙烧温度2个因素交互作用对As(Ⅲ)去除率的影响如图6和图7所示。由响应面结果可以看出,并非碳含量比例越高,处理效果越明显。当Fe/C=0.8时,随着植物基生物质组分的不断碳化,碳含量线性增加,不仅导致Fe0的相对含量降低,微原电池数量的损失,而且过多的碳化成分在植物基铁碳微电解材料表面堆积,导致Fe0活性点位与As(Ⅲ)接触减少,导致去除率降低[30]。当Fe/C=1.2,即Fe0的含量过量时,随着焙烧温度上升,植物基的有效碳含量增加,微电解体系中阴极数量增多,配对原电池数量增加,电极反应加快。但过量Fe0和水中微电解产生的Fe2+使溶液体系保持为还原环境,抑制As(Ⅲ)向As(Ⅴ)的转化过程,降低铁盐对As(Ⅲ)的去除率。原因在于植物基铁碳材料中铁盐混凝作用去除水溶液中As(Ⅲ)主要是通过在微原电池反应中失去电子形成的Fe3+和Fe2+离子与As(Ⅲ)和As(Ⅴ)形成亚砷酸铁、砷酸铁等沉淀作用;以及Fe(OH)2及Fe(OH)3等沉淀对水相中的As(Ⅲ)和As(Ⅴ)的静电吸附作用[31]。主要的As(Ⅲ)化合物(AsO33−)具有中性电荷,As(V)化合物(H2AsO4−、HAsO42−)在4-10的pH范围内带负电荷。因此,对于铁盐的氢氧化物沉淀而言,对As(V)化合物静电引力更强;As(V)化合物更容易被扩散吸附到铁盐水解沉积物表面,从而被吸附去除。
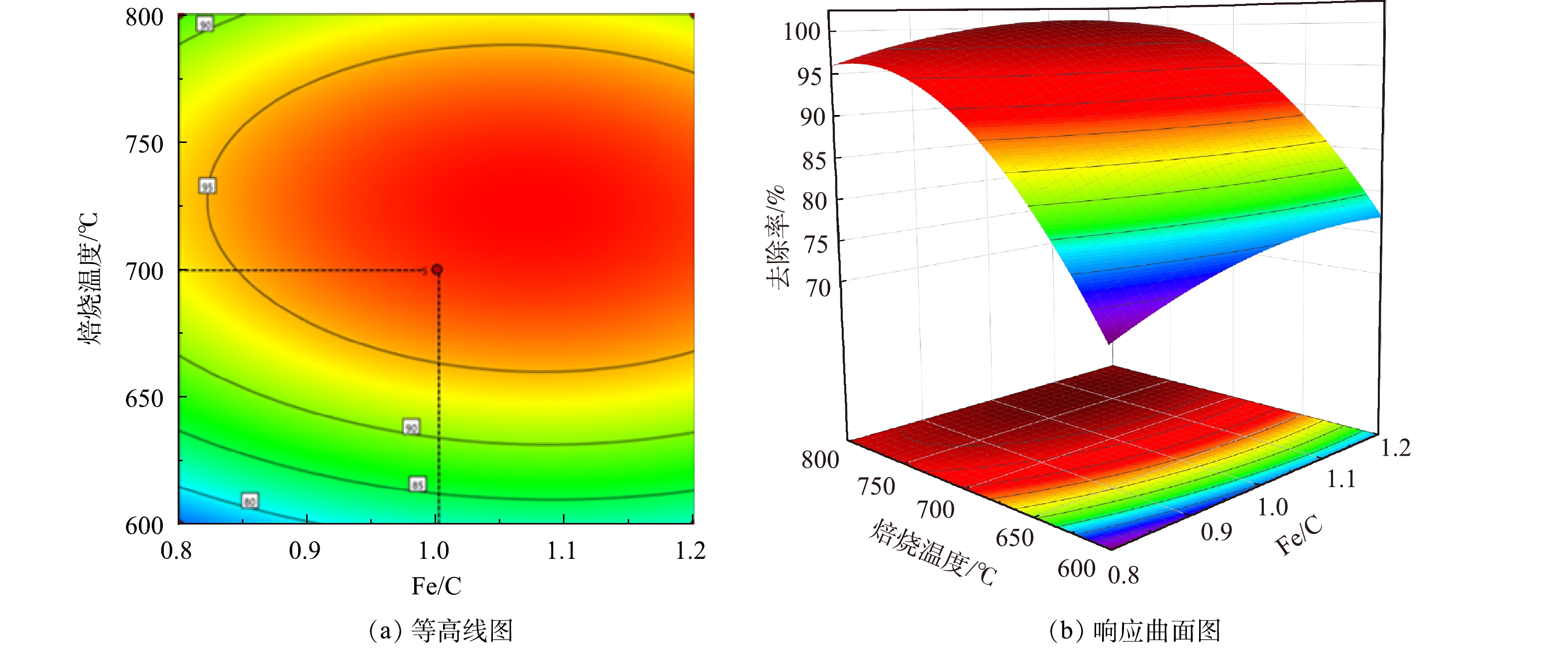 图 6 芦苇秆植物基Fe/C与焙烧温度交互影响总砷去除率的等高线与响应面Figure 6. Contour line and response surface of interaction of reed plant-based Fe/C and calcination temperature on total arsenic removal
图 6 芦苇秆植物基Fe/C与焙烧温度交互影响总砷去除率的等高线与响应面Figure 6. Contour line and response surface of interaction of reed plant-based Fe/C and calcination temperature on total arsenic removal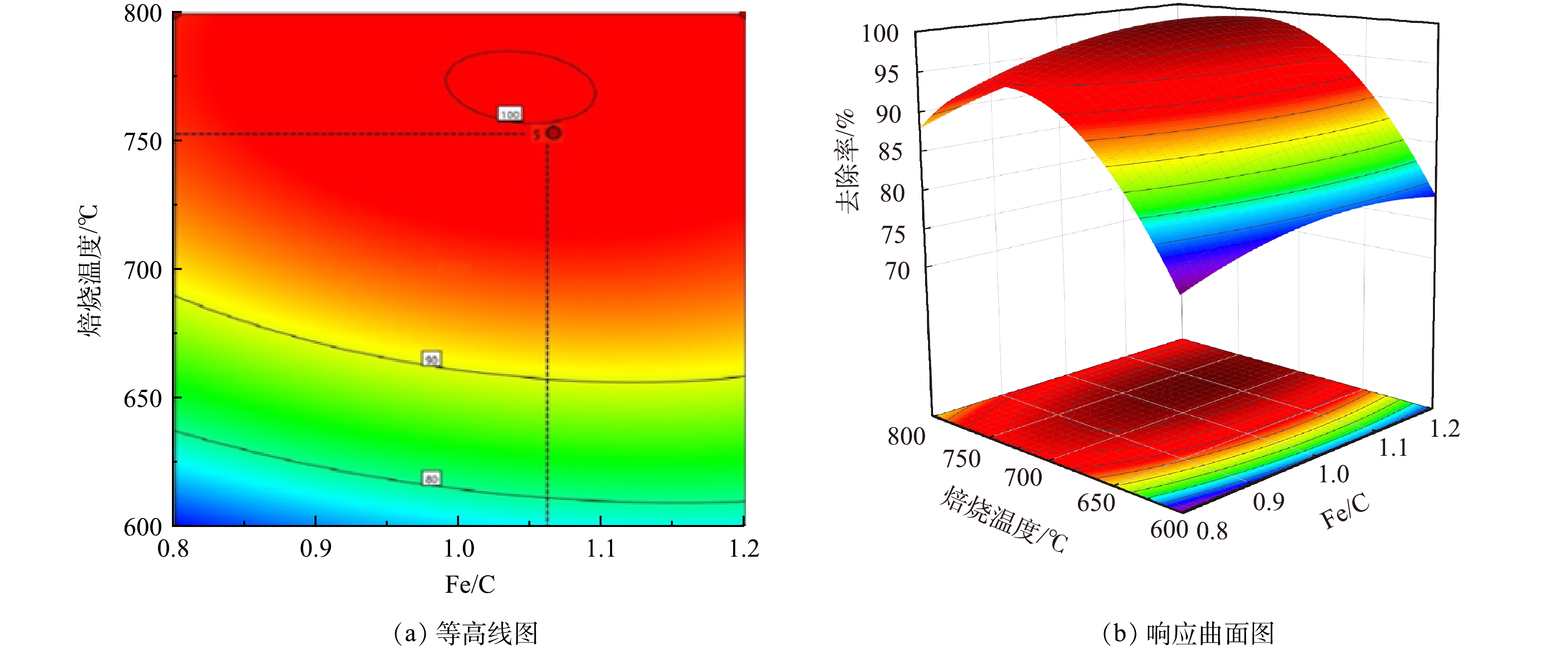 图 7 芦苇叶植物基Fe/C与焙烧温度交互影响总砷去除率的等高线与响应面Figure 7. Contour line and response surface of interaction of Phragmites australis leaves-based Fe/C and calcination temperature on total arsenic removal
图 7 芦苇叶植物基Fe/C与焙烧温度交互影响总砷去除率的等高线与响应面Figure 7. Contour line and response surface of interaction of Phragmites australis leaves-based Fe/C and calcination temperature on total arsenic removal芦苇秆植物基铁碳微电解材料在Fe/C=1.06时,即Fe0过量6%左右时取得最大响应值。一方面,在铁碳微电解材料制备过程中,材料表面Fe0存在被氧化的风险,且温度越高,氧化越明显。因此,在高温烧制芦苇秆植物基铁碳微电解材料时,需投加过量Fe0来保证Fe0-C微原电池数量,同时不会对溶液氧化还原环境造成破坏,使反应持续进行。另一方面,在超过700 ℃的焙烧温度下,Fe0与植物基中碳化成分进一步碳化形成Fe3C[32],减少铁碳微电解材料表面积碳,释放Fe0活性位点,使得阴阳两电极之间电子转移速率提高。并且Fe3C可作为微原电池反应的阴极,能与Fe0形成Fe3C-Fe0微原电池反应[33],有效填补高温焙烧下,富含木质素芦苇秆植物基碳化成分中作为阴极碳元素含量的减少,使得Fe3C-Fe0和Fe0-C 2种微原电池能共同作用,提高溶液中的原电池数量。对于木质素含量不足的芦苇叶植物基,随着焙烧温度的提高,在接近750 ℃时,Fe3C作为微原电池阴极,不能够有效填补高温焙烧下,植物基碳化成分中作为阴极碳元素含量的减少,导致阴极数量减少,从而降低As(Ⅲ)的去除率,因此焙烧温度不宜超过700 ℃,在Fe/C=1.01时取得最佳制备条件。铁碳微电解表面材料的SEM和XRD分析表征结果如图8所示。可见,芦苇秆植物基铁碳微电解材料750 ℃出现类似鱼鳞状Fe3C特征物象及Fe3C特征峰[34],证实有Fe3C的生成。
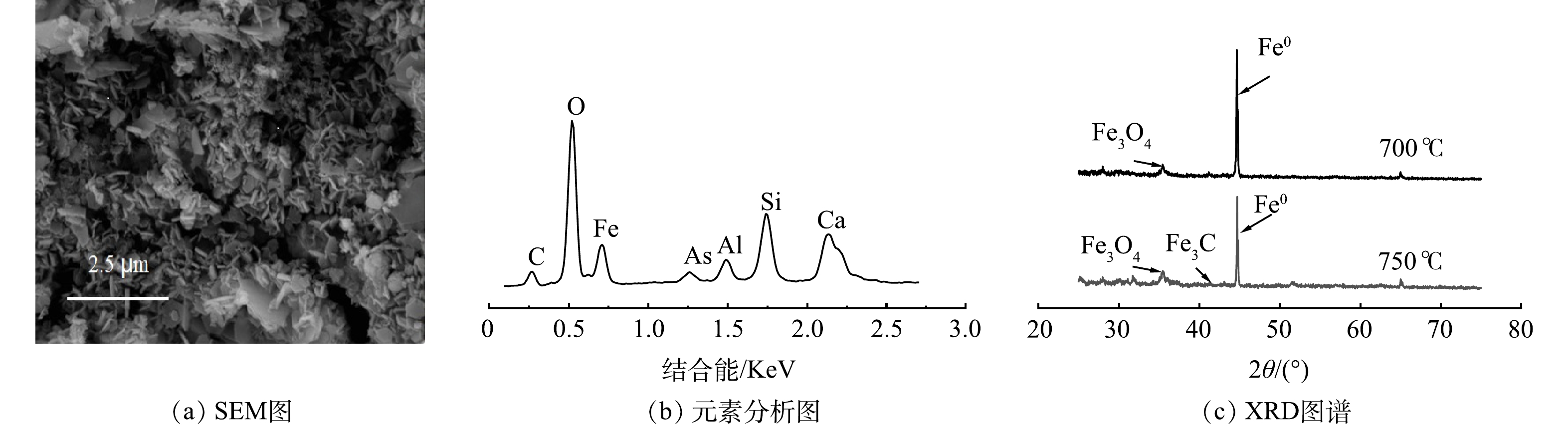 图 8 芦苇秆植物基铁碳微电解材料750 ℃表征图谱Figure 8. Characterization of reed rod plant-based Fe-C microelectrolysis materials at 750 ℃
图 8 芦苇秆植物基铁碳微电解材料750 ℃表征图谱Figure 8. Characterization of reed rod plant-based Fe-C microelectrolysis materials at 750 ℃2.5 不同生物质组成对电子转移及氧化As(Ⅲ)能力的影响
芦苇叶和芦苇秆在300~800 ℃下制备的碳基材料的元素组成以及H/C、O/C和O+N/C比如表5所示。由表5可知,随着梯度升温的进行,在碳化过程中碳元素和灰分相对含量增加,氧元素和氢元素含量降低。这是由于温度升高,C—H、C—O键断裂,氢和氧元素在此过程中发生脱附丢失所致[35-36]。对于碳基材料,石墨晶格中存在的层状组成分子(LCM)更稳定。因此,在碳化过程中,随着热解温度的升高更多的杂原子丢失(O、N、S、P),碳原子趋向重排为六元环系统的大分子网络,以试图获得这种更稳定的类似石墨状态[37]的芳香结构。其中,由H/C原子比可知碳基材料芳香性的大小,H/C越低,芳香化程度越高。在植物基铁碳微电解材料中,高芳香性的碳基结构,具有更高的反应活性[38],有利于促进Fe0-C微原电池的电子转移,加快反应的进行。木质素因碳含量高和热稳定性强的因素,在高温条件下,生物质中木质素占比越高,H/C越低,碳基成分芳香性越强。因此,在高温焙烧条件下,生物质中木质素含量越高的植物基铁碳微电解材料,越有利于Fe-C微原电池反应的进行。
表 5 不同温度烧制下植物基碳化成分的元素组成和原子比Table 5. Elemental composition and atomic ratio of plant-based carbonization components fired at different temperatures样品 元素组成/% 原子比 灰分/% C H N O S H/C O/C (O+N/C) 芦苇叶300 45.12 3.25 1.52 36.76 0.16 0.86 0.63 0.64 16.23 芦苇叶400 50.76 3.12 2.46 26.07 0.17 0.74 0.39 0.43 22.34 芦苇叶500 48.15 2.19 1.92 21.16 0.16 0.55 0.33 0.36 30.26 芦苇叶600 46.12 2.04 1.76 15.02 0.19 0.53 0.24 0.28 38.39 芦苇叶700 44.23 1.63 1.71 9.24 0.32 0.44 0.16 0.19 46.29 芦苇叶800 41.23 1.14 1.29 7.37 0.17 0.33 0.13 0.16 51.38 芦苇秆300 51.58 4.23 1.77 35.06 0.54 0.98 0.51 0.54 10.36 芦苇秆400 57.21 3.02 2.42 24.82 1.05 0.63 0.33 0.36 16.32 芦苇秆500 59.39 2.27 2.13 14.49 0.95 0.46 0.18 0.21 25.03 芦苇秆600 61.54 1.45 1.94 7.02 1.26 0.28 0.09 0.58 30.67 芦苇秆700 63.25 1.13 2.61 5.36 0.83 0.21 0.07 0.10 32.04 芦苇秆800 52.49 1.09 1.98 4.75 0.75 0.25 0.07 0.10 43.28 注:O%=100%-(C%-N%+H%+S%+灰分%) | Show TableDownLoad:
CSV
具有不同生物质组分的植物基材料其热解产物在氧化还原性质上也表现出较明显的差异[39]。热解产物氧化还原性质主要由有机官能团决定,如提供电子的羧基和酚羟基(还原剂)和接收电子的醌类和半醌类(氧化剂)等。随着热解的进行,有机成分中碳、氢和氧在热解过程中发生化学键的断裂,重组形成新的含氧和含碳官能团,从而表现出不同的氧化还原性质[25]。
不同烧制温度下芦苇秆与芦苇叶植物基的傅里叶红外光谱图(FTIR)如图9所示。在低温热解过程中(200~500 ℃),植物有机质热解产生大量含氢和氧官能团,此时碳基材料官能团以羧基和酚羟基的供电子官能团为主,包括C—H(1 384~1 402 cm−1)、—COOH(1 692 cm−1)、CH2(2 825 cm−1)、O—H(3 200~3 400 cm−1)等。芦苇叶植物基因富含易热解的半纤维素和纤维素组分,在低温热解过程官能团演化更完全[40],此时有芦苇叶植物基还原能力强于芦苇秆植物基。随着热解温度的提高,—COOH、—CH2、C—H和O—H的伸缩振动的强度逐渐降低直至消失,原因在于随着温度的升高,大量C—H、C—O键断裂,氢和氧以水分子形式丢失,官能团转以氧原子为中心的向以碳原子为中心转移。此时—O—(1 114 cm−l)、C=O、C=C(1 612~1 692 cm−1)和C—O(1 210 cm−1)伸缩振动存在,碳基材料以羰基和醌基的得电子氧化官能团为主。木质素因结构刚性强,难以分解,所以在高热解温度下官能团演变更稳定[41],因此,芦苇秆植物基在1 612~1 692 cm−1处C=O、C=C和1 210 cm−1处C—O的峰值更强,氧化能力强于芦苇叶植物基。并在1 556 cm−1处出现新的特征峰,可能是由于木质类生物质在高温下芳香化,形成具有类石墨结构的Π共轭的晶格体系[37]。
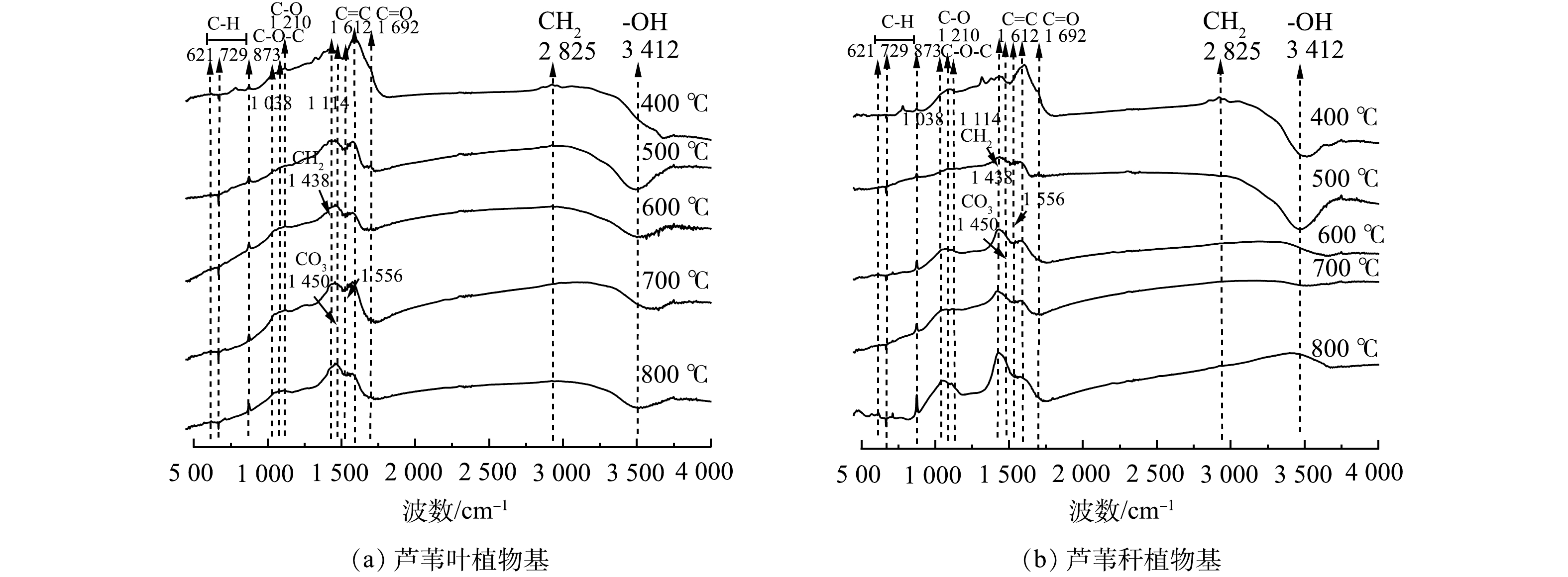 图 9 不同热解温度下不同植物基材料的傅里叶红外光谱图Figure 9. Fourier infrared spectra of different plant-based materials at different pyrolysis temperatures
图 9 不同热解温度下不同植物基材料的傅里叶红外光谱图Figure 9. Fourier infrared spectra of different plant-based materials at different pyrolysis temperatures2.6 植物基铁碳微电解材料对As(Ⅲ)去除机理
因植物基材料在热解过程中受热碳化并产生物油和不可冷凝气体,从而体积萎缩,使得植物基铁碳微电解材料形成连续贯通的多孔结构。本研究通过压汞法测得的植物基铁碳微电解材料的比表面积为1.1~1.2 m2·g−1、孔隙率为64.8%~65.32%,这有利于Fe-C微原电池反应的进行。植物基铁碳微电解材料对As(Ⅲ)去除机理可包括以下2条路径:一是在开始的反应过程中,As(Ⅲ)直接被植物基铁碳微电解材料吸附而去除;二是As(Ⅲ)被碳基材料中C=O、C=C和C—O等得电子官能团氧化为As(V),与在微原电池反应中失去电子形成的Fe3+和Fe2+共沉淀形成Fe2(AsO4)3、Fe3(AsO4)2,被吸附固定于铁碳微电解材料表面和多孔结构中,从而实现As(Ⅲ)的去除,去除机理如图10所示。
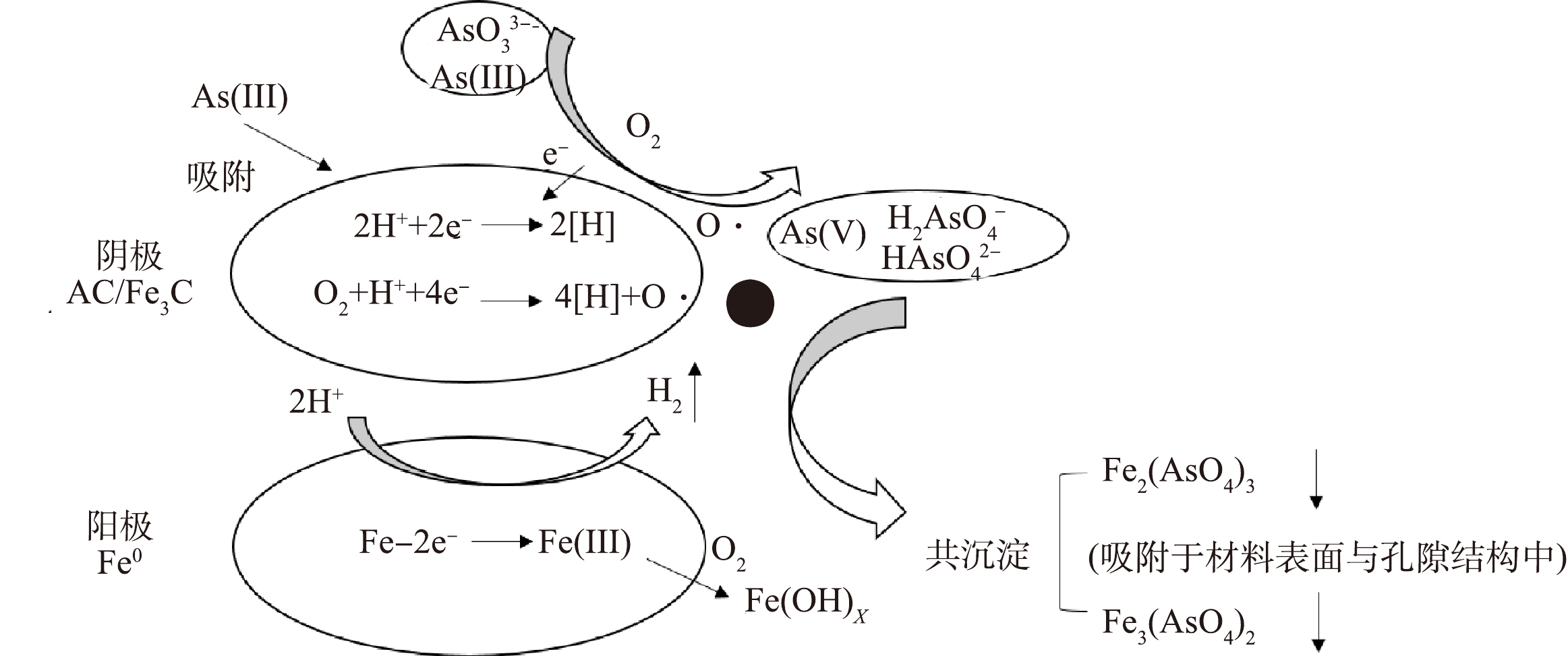 图 10 植物基铁碳微电解材料对As(III)去除的机理图Figure 10. Mechanism diagram of As(III) removal by plant-based Fe-C microelectrolytic materials
图 10 植物基铁碳微电解材料对As(III)去除的机理图Figure 10. Mechanism diagram of As(III) removal by plant-based Fe-C microelectrolytic materials3. 优化条件确定及模型
利用Design Expert 11.1.0.1软件对制备条件进行优化得到各植物基铁碳微电解材料最优预测模型,结果如表6所示。取100 mL 10 mg·L−1的NaAsO2溶液于250 mL锥形瓶中,调节pH至3后,加入根据最优制备条件分别制得的5 g植物基铁碳微电解材料,放入恒温摇床(25 ℃,100 r·min−1),避光振荡,计算得到反应48 h后芦苇杆和芦苇叶对As(Ⅲ)的去除率分别为98.12%和97.22%,与模型预测值的相对偏差分别为1.01%、1.46%,再次表明回归方程拟合度高,说明模型能够较好地适用于3个因数对芦苇叶和芦苇秆植物基铁碳微电解材料对砷去除率的影响。
表 6 各植物基铁碳微电解材料最优制备条件Table 6. The optimum preparation conditions of plant-based Fe-C microelectrolysis materials碳基材料 Fe/C摩尔比 碳化温度/℃ 焙烧温度/℃ As(Ⅲ)去除率/% 相对偏差/% 预测值 实验值 芦苇秆 1.06 507.87 751.92 99.19 98.12 1.07 芦苇叶 1.01 498.65 701.09 98.66 97.22 1.46 | Show TableDownLoad:
CSV
4. 结论
1)在Fe/C=1.06、碳化温度为507.87 ℃、焙烧温度为751.92 ℃下制备的芦苇秆植物基铁碳微电解材料对As(Ⅲ)去除率最高;在Fe/C=1.01、碳化温度为498.65 ℃、焙烧温度为701.09 ℃条件下制备得到的芦苇叶植物基铁碳微电解材料对As(Ⅲ)去除率最高。5 g芦苇杆和芦苇叶对 10 mg·L−1 NaAsO2去除率分别为98.12%和97.22%。
2)植物基生物质的不同组分中,木质素含量占比越大,制备铁碳微电解材料烧制温度及还原性铁投加量越高;同时在超过700 ℃焙烧温度下有Fe3C生成,形成Fe3C-Fe微原电池反应,从而提高对As(Ⅲ)的去除效率。
3)当植物基生物质的不同组分中纤维素和半纤维素含量较高,木质素含量较低时,较低的碳化和焙烧温度能降低材料中灰分含量,提高有效碳质量分数,增加微原电池数量,从而提升对As(Ⅲ)的去除率;当植物基生物质组分中木质素含量较高时,提升焙烧温度,植物基氧化官能团增多并形成具有类石墨结构的Π共轭的晶格体系,使得植物基铁碳材料氧化及电子传递能力提高,从而可提高As(Ⅲ)的去除效率。
-

图 1 HN-AD系统启动过程的脱氮效果
Figure 1. The efficiency of nitrogen removal at the start-up of the HN-AD process

图 2 HN-AD系统成功启动前后的单周期过程特征
Figure 2. Characteristics of a single-cycle before and after the successful start-up of the HN-AD system
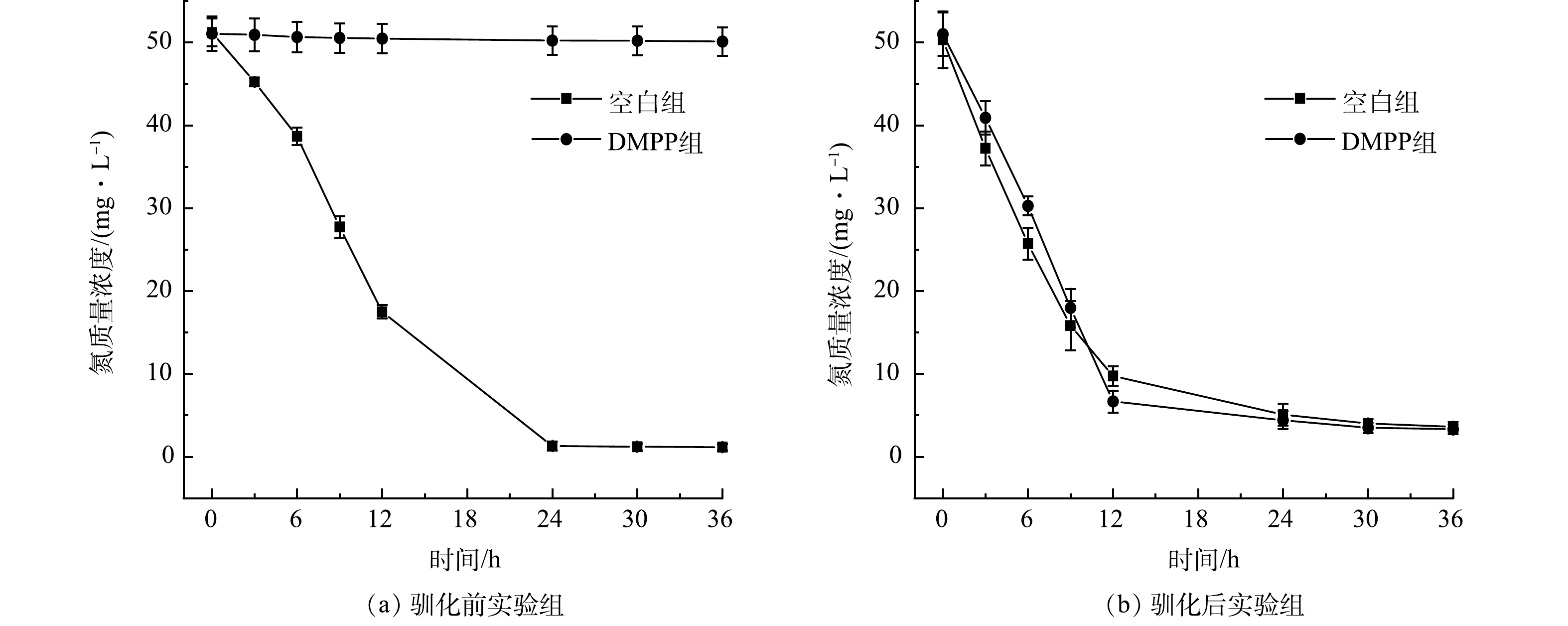
图 3 硝化抑制剂对异养硝化脱氮系统的影响
Figure 3. Effect of nitrification inhibitors on the heterotrophic nitrification and denitrification system
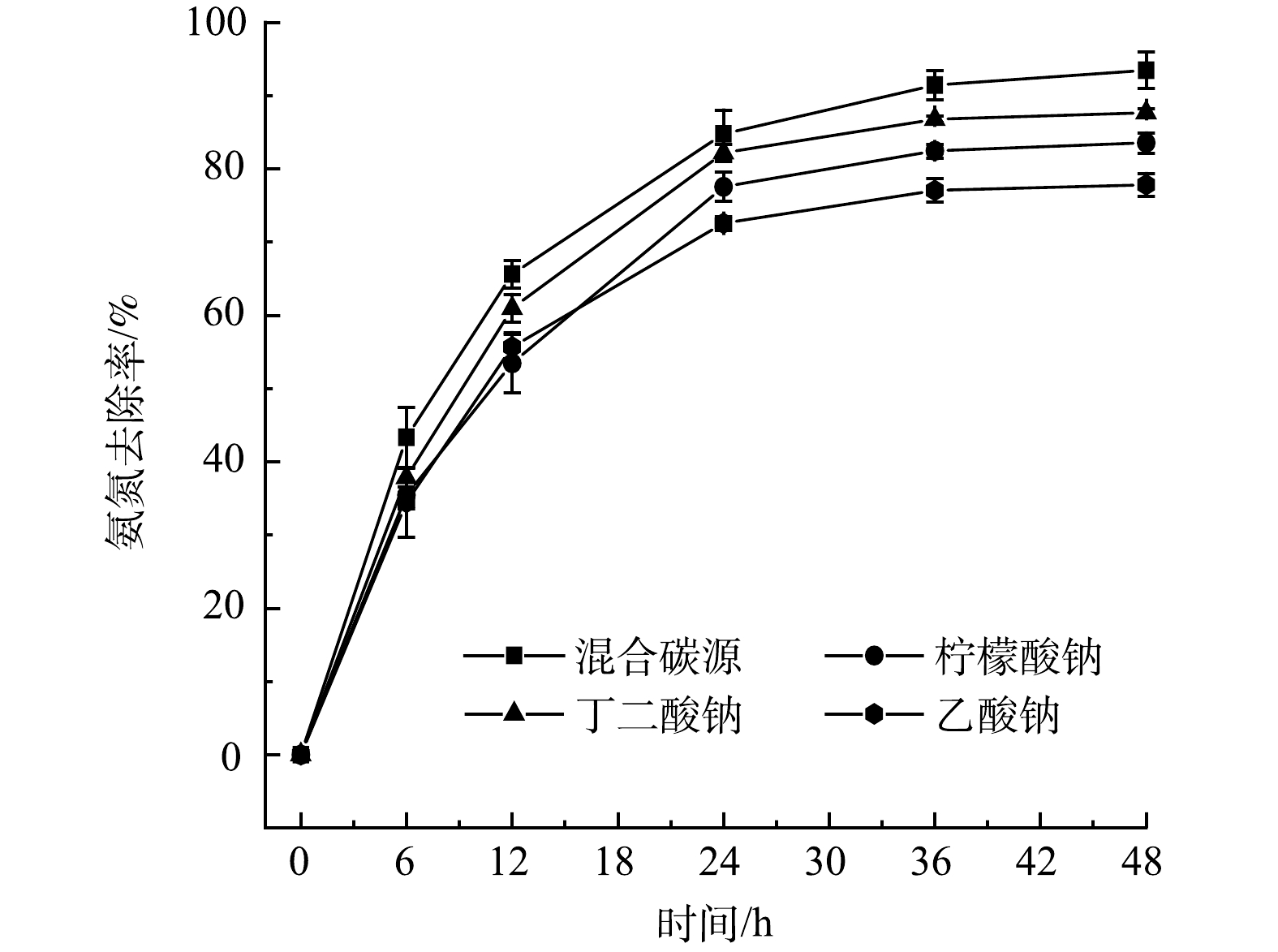
图 4 不同有机碳源下的氨氮去除效果
Figure 4. Removal efficiency of ammonium with different organic carbon sources
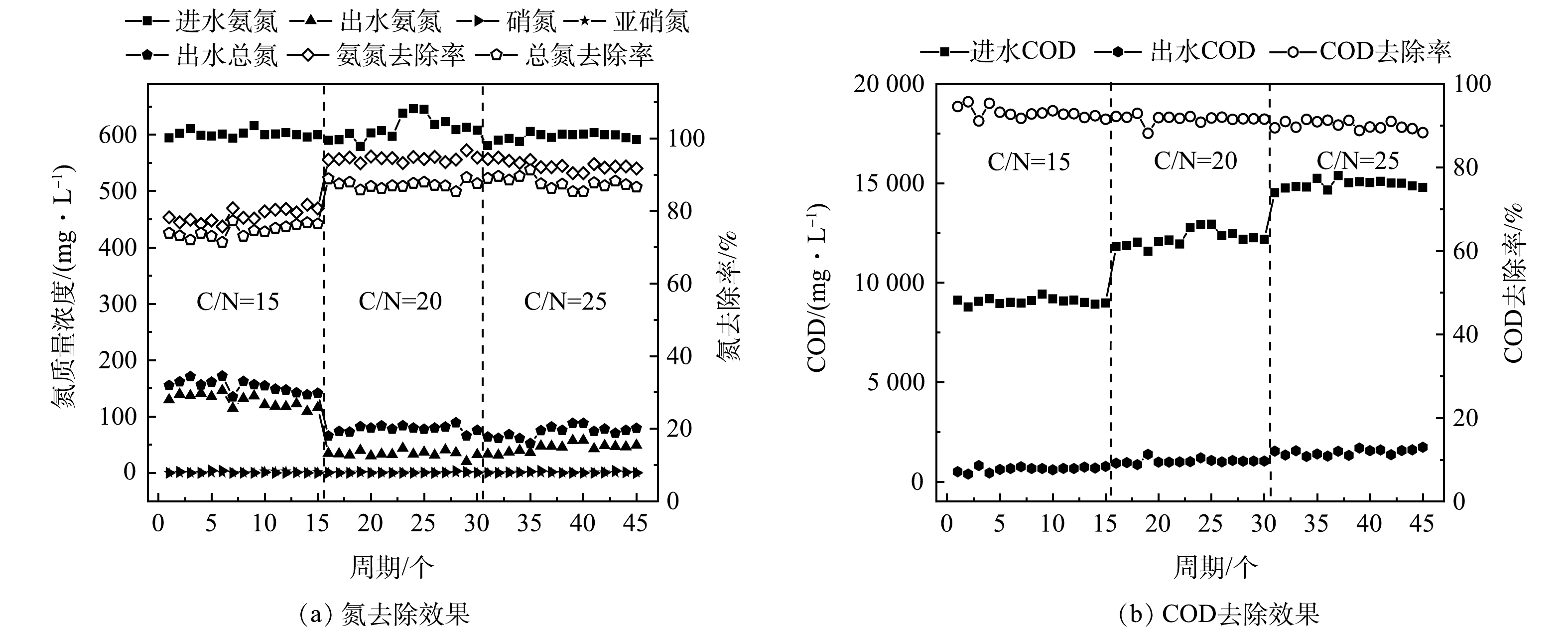
图 5 不同C/N比对模拟废水中NH4+-N、TN、COD的去除效果
Figure 5. Removal efficiencies of NH4+-N, TN and COD from the simulated wastewater at different C/N ratios
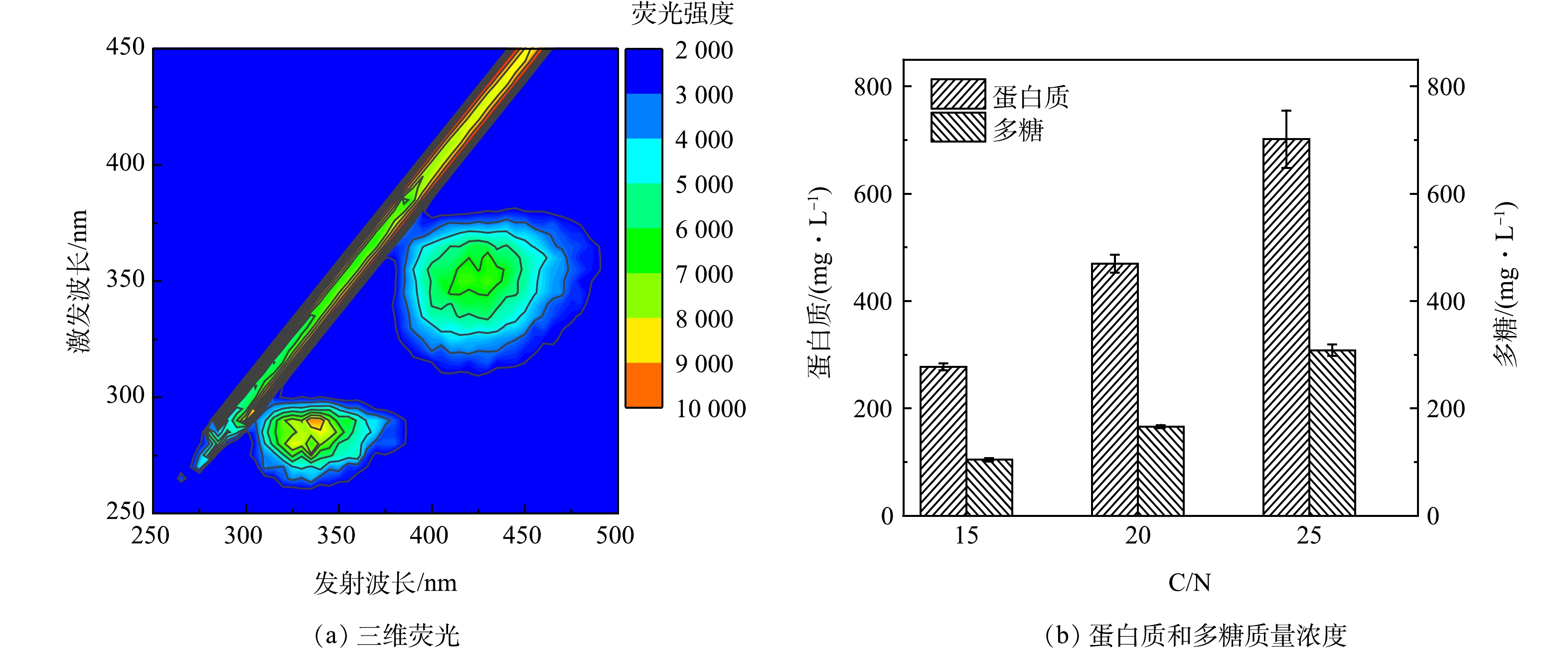
图 6 反应器出水三维荧光图及不同C/N比出水蛋白质和多糖质量浓度
Figure 6. Three-dimensional fluorescence pattern of reactor effluent and protein and polysaccharide concentrations of effluent at different C/N ratios

图 7 异养硝化系统COD和氨氮去除的动力学拟合曲线
Figure 7. Kinetic fitting curves of the removal of COD and ammonium in the heterotrophic nitrification and denitrification system
-
[1] SHIMOKAWA S. Sustainable meat consumption in China[J]. Journal of Integrative Agriculture, 2015, 14(6): 1023-1032. [2] YANG L, WANG X H, CUI S, et al. Simultaneous removal of nitrogen and phosphorous by heterotrophic nitrification-aerobic denitrification of a metal resistant bacterium Pseudomonas putida strain NP5[J]. Bioresource Technology, 2019, 285: 121360. [3] ROBERTSON L A, KUENEN J G. Aerobic denitrification: A controversy revived[J]. Archives of Microbiology, 1984, 139: 351-354. [4] CHEN J, GU S, HAO H, et al. Characteristics and metabolic pathway of Alcaligenes sp. TB for simultaneous heterotrophic nitrification-aerobic denitrification[J]. Applied Microbiology and Biotechnology, 2016, 100(22): 9787-9794. [5] 信欣, 姚力, 鲁磊, 等. 耐高氨氮异养硝化-好氧反硝化菌TN-14的鉴定及其脱氮性能[J]. 环境科学, 2014, 35(10): 3926-3932. [6] WANG Q, HE J. Complete nitrogen removal via simultaneous nitrification and denitrification by a novel phosphate accumulating Thauera sp. strain SND5[J]. Water Research, 2020, 185: 116300. [7] ZHANG T, SHAO M F, YE L. 454 Pyrosequencing reveals bacterial diversity of activated sludge from 14 sewage treatment plants[J]. The ISME Journal, 2012, 6(6): 1137-1147. [8] SONG K, GAO Y, YANG Y, et al. Performance of simultaneous carbon and nitrogen removal of high-salinity wastewater in heterotrophic nitrification-aerobic denitrification mode[J]. Journal of Environmental Chemical Engineering, 2023, 11(3): 109682. [9] CHEN J, ZHAO B, AN Q, et al. Kinetic characteristics and modelling of growth and substrate removal by Alcaligenes faecalis strain NR[J]. Bioprocess and Biosystems Engineering, 2016, 39: 593-601. [10] ANTHONISEN A, LOEHR R, PRAKASAM T, et al. Inhibition of nitrification by ammonia and nitrous acid[J]. Journal (Water Pollution Control Federation), 1976: 835-852. [11] REN S, WANG Z, JIANG H, et al. Stable nitritation of mature landfill leachate via in-situ selective inhibition by free nitrous acid[J]. Bioresource Technology, 2021, 340: 125647. [12] LEI Y, WANG Y, LIU H, et al. A novel heterotrophic nitrifying and aerobic denitrifying bacterium, Zobellella taiwanensis DN-7, can remove high-strength ammonium[J]. Applied Microbiology and Biotechnology, 2016, 100: 4219-4229. [13] HU B, LU J, QIN Y, et al. A critical review of heterotrophic nitrification and aerobic denitrification process: Influencing factors and mechanisms[J]. Journal of Water Process Engineering, 2023, 54: 103995. [14] LIU Y, WANG Y, LI Y, et al. Nitrogen removal characteristics of heterotrophic nitrification-aerobic denitrification by Alcaligenes faecalis C16[J]. Chinese Journal of Chemical Engineering, 2015, 23(5): 827-834. [15] OTANI Y, HASEGAWA K, HANAKI K. Comparison of aerobic denitrifying activity among three cultural species with various carbon sources[J]. Water Science and Technology, 2004, 50(8): 15-22. [16] JIN Y, WANG W, LI W, et al. Efficient denitrification of liquid digestate with its indigenous microflora[J]. Journal of Environmental Chemical Engineering, 2023, 11(5): 110636. [17] ZHAO Y, HUANG J, ZHAO H, et al. Microbial community and N removal of aerobic granular sludge at high COD and N loading rates[J]. Bioresource Technology, 2013, 143: 439-446. [18] ZHANG Y, SHI Z, CHEN M, et al. Evaluation of simultaneous nitrification and denitrification under controlled conditions by an aerobic denitrifier culture[J]. Bioresource Technology, 2015, 175: 602-605. [19] PAN Z, ZHOU J, LIN Z, et al. Effects of COD/TN ratio on nitrogen removal efficiency, microbial community for high saline wastewater treatment based on heterotrophic nitrification-aerobic denitrification process[J]. Bioresource Technology, 2020, 301: 122726. [20] 樊华, 方凡, 刘强, 等. 基于平行因子分析的藻菌共生膜污染机制研究[J]. 膜科学与技术, 2020, 40(4): 1007-8924. [21] WANG Z, GAO M, WANG S, et al. Effect of hexavalent chromium on extracellular polymeric substances of granular sludge from an aerobic granular sequencing batch reactor[J]. Chemical Engineering Journal, 2014, 251: 165-174. [22] HU J, YAN J, WU L, et al. Simultaneous nitrification and denitrification of hypersaline wastewater by a robust bacterium Halomonas salifodinae from a repeated-batch acclimation[J]. Bioresource Technology, 2021, 341: 125818. [23] ZHAO Y, ZHU Z, CHEN X, et al. Discovery of a novel potential polyphosphate accumulating organism without denitrifying phosphorus uptake function in an enhanced biological phosphorus removal process[J]. Science of the Total Environment, 2024, 912: 168952. 期刊类型引用(2)
1. 高航. 基于协同硫自养反硝化的工业酸洗电镀废水处理技术. 中国资源综合利用. 2025(03): 266-270 .  百度学术
百度学术
2. 张梦,张龙真,陶然,廖若梅,杨扬,邰义萍,傅木星. 不同填料曝气生物滤池对养猪沼液的处理效果及微生物群落响应. 环境工程学报. 2024(09): 2418-2427 . 本站查看
其他类型引用(1)
-

 点击查看大图
点击查看大图
计量
- 文章访问数: 2250
- HTML全文浏览数: 2250
- PDF下载数: 86
- 施引文献: 3

