









</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M15">$ \Delta{M}_{{\mathrm{N}\mathrm{O}}_{3}^{-}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M15.jpg"><img class="graphic" src="202307113-wanghengxin_M15.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M16">$ \Delta{M}_{{\mathrm{N}\mathrm{O}}_{2}^{-}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M16.jpg"><img class="graphic" src="202307113-wanghengxin_M16.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M17">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{1}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M17.jpg"><img class="graphic" src="202307113-wanghengxin_M17.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M18">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{2}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M18.jpg"><img class="graphic" src="202307113-wanghengxin_M18.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
</tr>
</thead>
<tbody>
<tr>
<td align="center" valign="middle">5</td>
<td align="center" valign="middle">335.16</td>
<td align="center" valign="middle">79.35</td>
<td align="center" valign="middle">74.87</td>
<td align="center" valign="middle">91.31</td>
<td align="center" valign="middle">243.84</td>
</tr>
<tr>
<td align="center" valign="middle">24</td>
<td align="center" valign="middle">394.68</td>
<td align="center" valign="middle">101.39</td>
<td align="center" valign="middle">88.34</td>
<td align="center" valign="middle">128.47</td>
<td align="center" valign="middle">266.2</td>
</tr>
<tr>
<td align="center" valign="middle">36</td>
<td align="center" valign="middle">468.91</td>
<td align="center" valign="middle">103.59</td>
<td align="center" valign="middle">94.3</td>
<td align="center" valign="middle">124.77</td>
<td align="center" valign="middle">344.14</td>
</tr>
<tr>
<td align="center" valign="middle">48</td>
<td align="center" valign="middle">407.25</td>
<td align="center" valign="middle">102.71</td>
<td align="center" valign="middle">93.03</td>
<td align="center" valign="middle">124.46</td>
<td align="center" valign="middle">282,79</td>
</tr>
<tr>
<td align="center" valign="middle" class="table_bottom_border">72</td>
<td align="center" valign="middle" class="table_bottom_border">376.14</td>
<td align="center" valign="middle" class="table_bottom_border">96.54</td>
<td align="center" valign="middle" class="table_bottom_border">84.47</td>
<td align="center" valign="middle" class="table_bottom_border">121.76</td>
<td align="center" valign="middle" class="table_bottom_border">254.38</td>
</tr>
</tbody>
</table></div></foreignObject></svg>"></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M15">$ \Delta{M}_{{\mathrm{N}\mathrm{O}}_{3}^{-}} $</tex-math><img class="inline-formula" style="display:none;" src="data:image/svg+xml,<svg xmlns='http://www.w3.org/2000/svg' width='350' height='600'><foreignObject width='2000' height='100%'><div xmlns='http://www.w3.org/1999/xhtml' style='font-size:16px;'><table>
<thead>
<tr>
<td align="center" valign="middle" class="table_top_border table_bottom_border2">饥饿时长/h</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M14">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{0}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M14.jpg"><img class="graphic" src="202307113-wanghengxin_M14.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M15">$ \Delta{M}_{{\mathrm{N}\mathrm{O}}_{3}^{-}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M15.jpg"><img class="graphic" src="202307113-wanghengxin_M15.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M16">$ \Delta{M}_{{\mathrm{N}\mathrm{O}}_{2}^{-}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M16.jpg"><img class="graphic" src="202307113-wanghengxin_M16.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M17">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{1}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M17.jpg"><img class="graphic" src="202307113-wanghengxin_M17.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M18">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{2}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M18.jpg"><img class="graphic" src="202307113-wanghengxin_M18.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
</tr>
</thead>
<tbody>
<tr>
<td align="center" valign="middle">5</td>
<td align="center" valign="middle">335.16</td>
<td align="center" valign="middle">79.35</td>
<td align="center" valign="middle">74.87</td>
<td align="center" valign="middle">91.31</td>
<td align="center" valign="middle">243.84</td>
</tr>
<tr>
<td align="center" valign="middle">24</td>
<td align="center" valign="middle">394.68</td>
<td align="center" valign="middle">101.39</td>
<td align="center" valign="middle">88.34</td>
<td align="center" valign="middle">128.47</td>
<td align="center" valign="middle">266.2</td>
</tr>
<tr>
<td align="center" valign="middle">36</td>
<td align="center" valign="middle">468.91</td>
<td align="center" valign="middle">103.59</td>
<td align="center" valign="middle">94.3</td>
<td align="center" valign="middle">124.77</td>
<td align="center" valign="middle">344.14</td>
</tr>
<tr>
<td align="center" valign="middle">48</td>
<td align="center" valign="middle">407.25</td>
<td align="center" valign="middle">102.71</td>
<td align="center" valign="middle">93.03</td>
<td align="center" valign="middle">124.46</td>
<td align="center" valign="middle">282,79</td>
</tr>
<tr>
<td align="center" valign="middle" class="table_bottom_border">72</td>
<td align="center" valign="middle" class="table_bottom_border">376.14</td>
<td align="center" valign="middle" class="table_bottom_border">96.54</td>
<td align="center" valign="middle" class="table_bottom_border">84.47</td>
<td align="center" valign="middle" class="table_bottom_border">121.76</td>
<td align="center" valign="middle" class="table_bottom_border">254.38</td>
</tr>
</tbody>
</table></div></foreignObject></svg>"></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M16">$ \Delta{M}_{{\mathrm{N}\mathrm{O}}_{2}^{-}} $</tex-math><img class="inline-formula" style="display:none;" src="data:image/svg+xml,<svg xmlns='http://www.w3.org/2000/svg' width='350' height='600'><foreignObject width='2000' height='100%'><div xmlns='http://www.w3.org/1999/xhtml' style='font-size:16px;'><table>
<thead>
<tr>
<td align="center" valign="middle" class="table_top_border table_bottom_border2">饥饿时长/h</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M14">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{0}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M14.jpg"><img class="graphic" src="202307113-wanghengxin_M14.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M15">$ \Delta{M}_{{\mathrm{N}\mathrm{O}}_{3}^{-}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M15.jpg"><img class="graphic" src="202307113-wanghengxin_M15.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M16">$ \Delta{M}_{{\mathrm{N}\mathrm{O}}_{2}^{-}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M16.jpg"><img class="graphic" src="202307113-wanghengxin_M16.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M17">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{1}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M17.jpg"><img class="graphic" src="202307113-wanghengxin_M17.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M18">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{2}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M18.jpg"><img class="graphic" src="202307113-wanghengxin_M18.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
</tr>
</thead>
<tbody>
<tr>
<td align="center" valign="middle">5</td>
<td align="center" valign="middle">335.16</td>
<td align="center" valign="middle">79.35</td>
<td align="center" valign="middle">74.87</td>
<td align="center" valign="middle">91.31</td>
<td align="center" valign="middle">243.84</td>
</tr>
<tr>
<td align="center" valign="middle">24</td>
<td align="center" valign="middle">394.68</td>
<td align="center" valign="middle">101.39</td>
<td align="center" valign="middle">88.34</td>
<td align="center" valign="middle">128.47</td>
<td align="center" valign="middle">266.2</td>
</tr>
<tr>
<td align="center" valign="middle">36</td>
<td align="center" valign="middle">468.91</td>
<td align="center" valign="middle">103.59</td>
<td align="center" valign="middle">94.3</td>
<td align="center" valign="middle">124.77</td>
<td align="center" valign="middle">344.14</td>
</tr>
<tr>
<td align="center" valign="middle">48</td>
<td align="center" valign="middle">407.25</td>
<td align="center" valign="middle">102.71</td>
<td align="center" valign="middle">93.03</td>
<td align="center" valign="middle">124.46</td>
<td align="center" valign="middle">282,79</td>
</tr>
<tr>
<td align="center" valign="middle" class="table_bottom_border">72</td>
<td align="center" valign="middle" class="table_bottom_border">376.14</td>
<td align="center" valign="middle" class="table_bottom_border">96.54</td>
<td align="center" valign="middle" class="table_bottom_border">84.47</td>
<td align="center" valign="middle" class="table_bottom_border">121.76</td>
<td align="center" valign="middle" class="table_bottom_border">254.38</td>
</tr>
</tbody>
</table></div></foreignObject></svg>"></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M17">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{1}} $</tex-math><img class="inline-formula" style="display:none;" src="data:image/svg+xml,<svg xmlns='http://www.w3.org/2000/svg' width='350' height='600'><foreignObject width='2000' height='100%'><div xmlns='http://www.w3.org/1999/xhtml' style='font-size:16px;'><table>
<thead>
<tr>
<td align="center" valign="middle" class="table_top_border table_bottom_border2">饥饿时长/h</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M14">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{0}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M14.jpg"><img class="graphic" src="202307113-wanghengxin_M14.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M15">$ \Delta{M}_{{\mathrm{N}\mathrm{O}}_{3}^{-}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M15.jpg"><img class="graphic" src="202307113-wanghengxin_M15.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M16">$ \Delta{M}_{{\mathrm{N}\mathrm{O}}_{2}^{-}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M16.jpg"><img class="graphic" src="202307113-wanghengxin_M16.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M17">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{1}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M17.jpg"><img class="graphic" src="202307113-wanghengxin_M17.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M18">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{2}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M18.jpg"><img class="graphic" src="202307113-wanghengxin_M18.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
</tr>
</thead>
<tbody>
<tr>
<td align="center" valign="middle">5</td>
<td align="center" valign="middle">335.16</td>
<td align="center" valign="middle">79.35</td>
<td align="center" valign="middle">74.87</td>
<td align="center" valign="middle">91.31</td>
<td align="center" valign="middle">243.84</td>
</tr>
<tr>
<td align="center" valign="middle">24</td>
<td align="center" valign="middle">394.68</td>
<td align="center" valign="middle">101.39</td>
<td align="center" valign="middle">88.34</td>
<td align="center" valign="middle">128.47</td>
<td align="center" valign="middle">266.2</td>
</tr>
<tr>
<td align="center" valign="middle">36</td>
<td align="center" valign="middle">468.91</td>
<td align="center" valign="middle">103.59</td>
<td align="center" valign="middle">94.3</td>
<td align="center" valign="middle">124.77</td>
<td align="center" valign="middle">344.14</td>
</tr>
<tr>
<td align="center" valign="middle">48</td>
<td align="center" valign="middle">407.25</td>
<td align="center" valign="middle">102.71</td>
<td align="center" valign="middle">93.03</td>
<td align="center" valign="middle">124.46</td>
<td align="center" valign="middle">282,79</td>
</tr>
<tr>
<td align="center" valign="middle" class="table_bottom_border">72</td>
<td align="center" valign="middle" class="table_bottom_border">376.14</td>
<td align="center" valign="middle" class="table_bottom_border">96.54</td>
<td align="center" valign="middle" class="table_bottom_border">84.47</td>
<td align="center" valign="middle" class="table_bottom_border">121.76</td>
<td align="center" valign="middle" class="table_bottom_border">254.38</td>
</tr>
</tbody>
</table></div></foreignObject></svg>"></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M18">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{2}} $</tex-math><img class="inline-formula" style="display:none;" src="data:image/svg+xml,<svg xmlns='http://www.w3.org/2000/svg' width='350' height='600'><foreignObject width='2000' height='100%'><div xmlns='http://www.w3.org/1999/xhtml' style='font-size:16px;'><table>
<thead>
<tr>
<td align="center" valign="middle" class="table_top_border table_bottom_border2">饥饿时长/h</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M14">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{0}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M14.jpg"><img class="graphic" src="202307113-wanghengxin_M14.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M15">$ \Delta{M}_{{\mathrm{N}\mathrm{O}}_{3}^{-}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M15.jpg"><img class="graphic" src="202307113-wanghengxin_M15.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M16">$ \Delta{M}_{{\mathrm{N}\mathrm{O}}_{2}^{-}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M16.jpg"><img class="graphic" src="202307113-wanghengxin_M16.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M17">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{1}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M17.jpg"><img class="graphic" src="202307113-wanghengxin_M17.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
<td align="center" valign="middle" class="table_top_border table_bottom_border2"><inline-formula><tex-math id="M18">$ \Delta{M}_{\mathrm{H}\mathrm{A}{\mathrm{c}}_{2}} $</tex-math><alternatives><img class="graphic" src="202307113-wanghengxin_M18.jpg"><img class="graphic" src="202307113-wanghengxin_M18.png"></alternatives></inline-formula>/(mg·L<sup>−1</sup>)</td>
</tr>
</thead>
<tbody>
<tr>
<td align="center" valign="middle">5</td>
<td align="center" valign="middle">335.16</td>
<td align="center" valign="middle">79.35</td>
<td align="center" valign="middle">74.87</td>
<td align="center" valign="middle">91.31</td>
<td align="center" valign="middle">243.84</td>
</tr>
<tr>
<td align="center" valign="middle">24</td>
<td align="center" valign="middle">394.68</td>
<td align="center" valign="middle">101.39</td>
<td align="center" valign="middle">88.34</td>
<td align="center" valign="middle">128.47</td>
<td align="center" valign="middle">266.2</td>
</tr>
<tr>
<td align="center" valign="middle">36</td>
<td align="center" valign="middle">468.91</td>
<td align="center" valign="middle">103.59</td>
<td align="center" valign="middle">94.3</td>
<td align="center" valign="middle">124.77</td>
<td align="center" valign="middle">344.14</td>
</tr>
<tr>
<td align="center" valign="middle">48</td>
<td align="center" valign="middle">407.25</td>
<td align="center" valign="middle">102.71</td>
<td align="center" valign="middle">93.03</td>
<td align="center" valign="middle">124.46</td>
<td align="center" valign="middle">282,79</td>
</tr>
<tr>
<td align="center" valign="middle" class="table_bottom_border">72</td>
<td align="center" valign="middle" class="table_bottom_border">376.14</td>
<td align="center" valign="middle" class="table_bottom_border">96.54</td>
<td align="center" valign="middle" class="table_bottom_border">84.47</td>
<td align="center" valign="middle" class="table_bottom_border">121.76</td>
<td align="center" valign="middle" class="table_bottom_border">254.38</td>
</tr>
</tbody>
</table></div></foreignObject></svg>"></inline-formula>/(mg·L<sup>−1</sup>)</td>
</tr>
</thead>
<tbody>
<tr>
<td align="center" valign="middle">5</td>
<td align="center" valign="middle">335.16</td>
<td align="center" valign="middle">79.35</td>
<td align="center" valign="middle">74.87</td>
<td align="center" valign="middle">91.31</td>
<td align="center" valign="middle">243.84</td>
</tr>
<tr>
<td align="center" valign="middle">24</td>
<td align="center" valign="middle">394.68</td>
<td align="center" valign="middle">101.39</td>
<td align="center" valign="middle">88.34</td>
<td align="center" valign="middle">128.47</td>
<td align="center" valign="middle">266.2</td>
</tr>
<tr>
<td align="center" valign="middle">36</td>
<td align="center" valign="middle">468.91</td>
<td align="center" valign="middle">103.59</td>
<td align="center" valign="middle">94.3</td>
<td align="center" valign="middle">124.77</td>
<td align="center" valign="middle">344.14</td>
</tr>
<tr>
<td align="center" valign="middle">48</td>
<td align="center" valign="middle">407.25</td>
<td align="center" valign="middle">102.71</td>
<td align="center" valign="middle">93.03</td>
<td align="center" valign="middle">124.46</td>
<td align="center" valign="middle">282,79</td>
</tr>
<tr>
<td align="center" valign="middle" class="table_bottom_border">72</td>
<td align="center" valign="middle" class="table_bottom_border">376.14</td>
<td align="center" valign="middle" class="table_bottom_border">96.54</td>
<td align="center" valign="middle" class="table_bottom_border">84.47</td>
<td align="center" valign="middle" class="table_bottom_border">121.76</td>
<td align="center" valign="middle" class="table_bottom_border">254.38</td>
</tr>
</tbody>
</table></div></foreignObject></svg>)
-
循环活性污泥法(cyclic activated sludge technology, CAST)作为一种基于序批式反应器(sequencing batch reactor, SBR)工艺改良的新技术,因其运行简便、结构紧凑及运行费用低等优点而被广泛应用于城镇污水处理厂中[1-2]。CAST反应池通常由选择池、缺氧池和好氧池3部分构成。在生物选择池内,将原水和混合液按体积比为5∶1的比例混合,使回流污泥处于高负荷条件下,这能有效抑制丝状菌的增殖,避免污泥膨胀[3-4]。由于污泥回流过程中会带有一定量的硝态氮(NO3−-N),反硝化菌在选择池和缺氧池中利用有机物将NO3−-N还原为氮气(N2)的同时,也将原水中的有机物储存为细胞内碳源[5],这为好氧池内的同步硝化-反硝化脱氮(simultaneous nitrification and denitrification, SND)创造了条件。BIAN等[6]同时研究CAST工艺的脱碳与脱氮性能,结果表明,TN主要通过以亚硝酸盐为间歇产物的同步硝化-反硝化机制去除,曝气期间同步硝化-反硝化的发生是由于反应器悬浮载体中存在微缺氧环境所致。WANG等[7]指出通过改变关键操作条件(温度,DO等)可以控制脱氮路径,控制同步硝化-反硝化成为主要脱氮路径,并在不同温度下通过调整有机负荷改善脱氮效率。WANG等[8]在对CAST系统进行脱氮性能的评估中,同样证实同步硝化-反硝化脱氮贡献与操作条件有关,通过优化操作条件可提高同步硝化-反硝化过程的脱氮性能,推测DO在1 mg·L−1时亚硝酸盐氧化菌的抑制作用会影响亚硝酸盐的积累。然而,目前关于CAST工艺的脱氮路径及脱氮特性的报道不多。基于此,对CAST工艺的脱氮性能进行评价十分必要。
为此,本研究以某污水处理厂CAST工艺为研究对象,通过对典型周期内氮组分、污泥浓度等进行测定,结合污泥活性、同步硝化-反硝化速率和饱食-饥饿(feast-famine)等批次实验,评价了该工艺的脱氮性能,以期为我国城市污水处理厂CAST工艺的设计、运行及水厂提质增效提供参考。
-
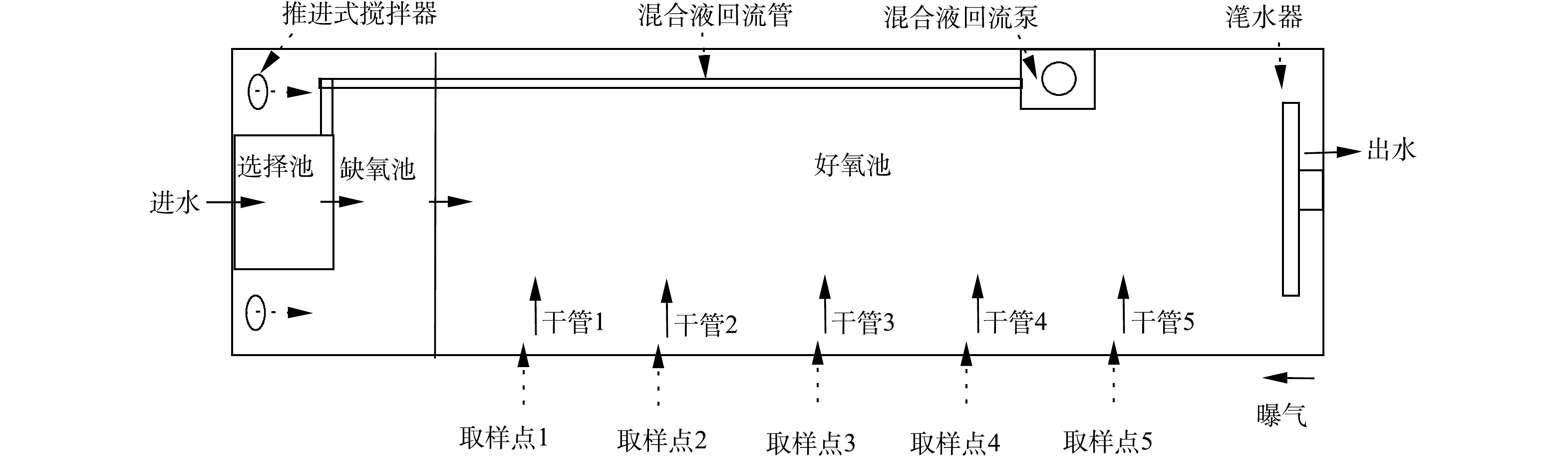
该污水处理厂处理规模为3.5×104 m3·d−1,其中一期工程2.0×104 m3·d−1,采用CAST工艺;二期工程1.5×104 m3·d−1,采用A2/O工艺。CAST工艺由4组反应池构成,每一组反应池的尺寸为76 m×11.6 m×6 m,选择池、缺氧池和好氧池体积比为1∶4∶25。该工艺水力停留时间(hydraulic retention time, HRT)为18.8 h,污泥龄(sludge retention time, SRT)为14.7 d,污泥回流比为20%。CAST工艺包括4个运行步骤:进水曝气、曝气、沉淀和出水,每步骤时长为1 h。缺氧池中安装潜水混合器,保证缺氧池内活性污泥处于悬浮状态;好氧池内安装微孔曝气器,以维持所需溶解氧(dissolved oxygen, DO)浓度及搅拌。在好氧池末端设有混合液回流泵,每运行周期前2 h进行回流,流量为80 m3·d−1。厌氧池和缺氧池设有氧化还原电位(oxidation-reduction potential, ORP)测定仪,在线显示氧化还原电位。好氧池中设有溶解氧仪和污泥浓度仪,在线检测DO浓度和污泥浓度,并反馈至中控室,随时调节鼓风机送风量和排泥量。
-
氨氮(NH4+-N)、亚硝氮(NO2−-N)和硝氮(NO3−-N)等的测定参照标准分析方法[9]。NH4+-N的测定采用纳氏试剂分光光度法,NO2−-N的测定采用N-(1-萘基)-乙二胺分光光度法,NO3−-N的测定采用紫外分光光度法,MLSS和MLVSS的测定采用重量法。DO由便携式溶氧仪(SevenGo pro SG6)测定。本研究中以乙酸钠作为批次实验的碳源,其测定方法为气象色谱法,测定仪器为SP-3420A型气象色谱仪。
-
为研究CAST工艺好氧池同步硝化-反硝化脱氮特性,沿池长设定5个取样点(图1),在进水曝气和曝气段每间隔15 min取样且测定氮组分浓度和记录DO浓度。
CAST反应池的脱氮主要由3部分贡献:缺氧池反硝化脱氮(DEN)、同步硝化-反硝化脱氮(SND)、沉淀阶段的内源反硝化脱氮(ED)。总脱氮量根据式(1)计算,同步硝化-反硝化脱氮量根据式(2)计算,内源反硝化脱氮量根据式(3)计算,选择池及缺氧池脱氮量根据式(4)计算,每周期开始时池内总氮量根据式(5)计算,每周期进水总氮量根据式(6)计算,每周期出水总氮量根据式(7)计算,曝气结束时池内总氮量根据式(8)计算。
式中:M 为每周期总脱氮量,mg(以N计);Minf为每周期进水总氮量,mg;Meff为每周期出水总氮量,mg;Msnd为每周期同步硝化-反硝化脱氮量,mg;Mst为每周期起始时CAST池内总氮量,mg;Mhd为每周期缺氧池脱氮量,mg;Med为每周期内源反硝化脱氮量,mg;Mend为每周期曝气结束时CAST池总氮量,mg; V1为混合液回流量,80 m3·h−1;T为回流时间,2 h;C1为回流混合液平均NO3−-N浓度,mg·L−1;V2为每周期开始时池内混合液体积,m3;C2为每周期开始时池内混合液总氮浓度,mg·L−1;V3为进水体积,m3;C3为进水总氮浓度,mg·L−1;C4为出水总氮浓度,mg·L−1;C5为曝气结束时池内混合液总氮浓度,mg·L−1。
-
为研究CAST反应池内不同DO浓度下的同步硝化-反硝化脱氮特性,本研究在CAST工艺进水结束时取1号采样点的泥水混合物于5 L试验装置内,在DO浓度分别为1.0、1.5、2.0和2.5 mg·L−1下进行同步硝化-反硝化率测定。试验装置内设有搅拌器,在反应器顶端设有取样口。测定过程中,每隔15 min取样并测定NH4+-N和NO3−-N浓度,随后根据浓度变化拟合得出NH4+-N消耗速率和NO3−-N生成速率,最终根据两速率计算得出同步硝化-反硝化率(式(9))。
式中:
R 为同步硝化反硝化率,%;RNH+4-N 为NH4+-N消耗速率,mg·(g·h)−1(mg以氮计、g以VSS计);RNH+4-N 为NO3−-N生成速率,mg·(g·h)−1。 -
为确定CAST工艺中饥饿时长对微生物吸收乙酸的影响,本研究将活性污泥等分为5份,置于1 L血清瓶中,活性污泥(SS)浓度为3 608 mg·L−1,VSS·SS−1为0.55。向每一份污泥加入足量的NO3−-N使其处于饥饿状态,饥饿时长依次为5、24、36、48和72 h。饥饿结束后加入足量的乙酸、NO3−-N和NH4+-N,间隔15 min进行取样,测定基质浓度变化,根据式(10)~式(12)计算最大乙酸吸收量。
式中:
ΔMHAc1 为将NO3−-N还原为NO2−-N及N2所消耗的乙酸量,mg·L−1;ΔMNO−3 为反应过程中浓度减少量,mg·L−1;ΔMNO−2 为反应过程中浓度增加量,mg·L−1;ΔMHAc0 为反应过程中消耗的乙酸量,mg·L−1;ΔMHAc2 为反应过程中吸收的乙酸量,mg·L−1;CVSS 为污泥浓度,g·L−1;ΔMHAc 为单位质量微生物吸收的乙酸量,g·g−1(以VSS计)。 -
活性污泥的内源反硝化活性采用基质消耗速率法测定,具体操作如下:CAST工艺每周期曝气结束时,在好氧池末端取一定量的活性污泥,用水淘洗去除残余基质,将淘洗过的活性污泥加入密闭的反应器内,向其中加入硝酸钠,控制NO3−-N为20 mg·L−1,每隔30 min取泥水混合物进行过滤,之后测定滤液中的NO3−-N、NO2−-N和NH4+-N的浓度,活性通过基质浓度变化线性拟合得出。
-
CAST工艺自2013年投入运行以来,各项水质指标均满足《城镇污水处理厂污染物排放标准》(GB 18918-2002)一级B标准。2021年10月—2022年5月,CAST工艺的主要进出水水质见表1。可以看出,CAST工艺出水COD、TN和PO43--P浓度分别小于30、15和0.3 mg·L−1,均达到A标准的水质要求。而对于出水NH4+-N,尽管略高于A标准所规定的1.5 mg·L−1,但在二期A2/O工艺出水稀释下,污水厂总出水可达标排放。
-
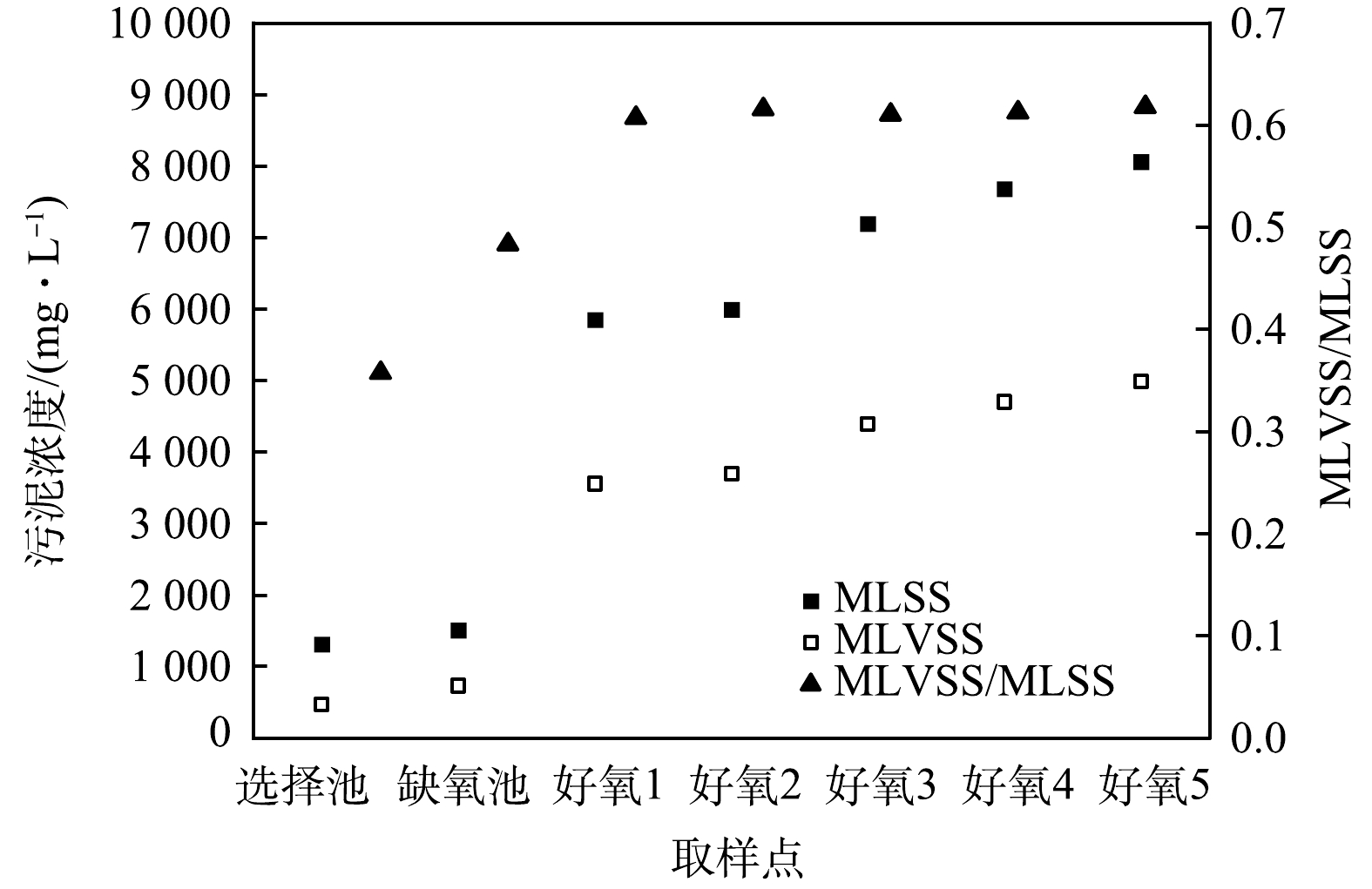
CAST工艺属于SBR工艺的一种演变,其运行方式也与SBR工艺相同,但CAST的推流式进水方式使得好氧池的反应并非完全混合式。考虑到CAST工艺的脱氮主要在好氧池。因此,首先对好氧池流态进行分析,结合选择池、缺氧池和好氧池的污泥浓度测定,判断各反应区的流体力学特性。由测定结果可知(图2),各反应区的污泥浓度分布并不均匀,在选择池和缺氧池内污泥浓度基本不变,说明这2个反应区的流态呈完全混合式;而好氧池进水端污泥浓度最低,约6 000 mg·L−1;出水端最高,约8 000 mg·L−1,说明回流混合液和原水混合进入好氧池后,以推流式方式前进,由于混合液回流比较低,使得污泥浓度沿程增加。
CAST工艺序批式的运行方式及推流式进水方式使其具有独特的脱氮特性。在周期开始时,回流混合液和原水在选择池内混合,此时混合液中的NO3−-N被反硝化菌利用原水中的有机物还原成N2(反硝化脱氮)。值得注意的是,CAST工艺的回流比仅为20%,使得回流至缺氧池的NO3−-N含量有限,继而选择池和缺氧池对TN脱除的贡献率较低;其次,回流混合液中的活性污泥在选择池和缺氧池中将原水中的有机物以胞内聚合物的形式储存,之后进入好氧池内,进而作为同步硝化-反硝化脱氮的碳源;最后,当曝气结束后,好氧池内活性污泥开始沉淀,池中剩余DO首先被消耗殆尽,此时好氧池部分区域进入缺氧状态,反硝化菌开始利用胞内聚合物及内碳源进行内源反硝化脱氮。综上所述,CAST工艺脱氮途径包括选择池及缺氧池反硝化脱氮、好氧池同步硝化-反硝化脱氮及内源反硝化脱氮。
为明确3种脱氮途径对TN脱除的贡献率,结合典型周期水质指标,利用式(1)~(8)对其进行计算,结果见表2。可以看出,选择池和缺氧池中反硝化脱氮贡献率最低,仅为(1.64±0.05)%,这一结果主要和混合液回流比较低(20%)有关,其可通过增大回流比提升,但需对回流泵进行更换;沉淀出水阶段的内源反硝化脱氮贡献最大,可达(62.86±4.13)%,类似结果在二沉池中也有观察到[10-11],但受限于沉淀时间短等因素,其脱氮贡献率难以进一步提升;除去反硝化和内源反硝化脱氮,TN的脱除全由好氧池同步硝化-反硝化途径完成,贡献率可达(35.50±4.15)%,对其进行优化,或成为进一步提升CAST工艺脱氮性能的突破口。基于此,本研究从如何优化同步硝化-反硝化过程入手,通过探究DO浓度和饥饿时长对同步硝化-反硝化脱氮性能的影响机制,从而确定适宜CAST工艺的运行策略。
-
为分析DO浓度对同步硝化-反硝化过程的影响特性,测定了不同DO浓度下好氧池曝气阶段DO、NH4+-N及NO3−-N浓度随时间的变化情况,结果如图3所示。图3(a)~(b)为低DO浓度(0.5~1 mg·L−1)下同步硝化-反硝化的脱氮特性。由图3(a)可知,反应过程中NH4+-N仅很少一部分被硝化,曝气结束时仍有约12 mg·L−1NH4+-N,而NO3−-N浓度几乎为零,不能为同步硝化-反硝化过程提供足够的电子受体,进而导致脱氮效率极低。
图3(c)~(d)为中等DO浓度(1~4 mg·L−1)下同步硝化-反硝化脱氮特性。由图3(c)可知,取样点1、3和5的NH4+-N和NO3−-N浓度随反应的进行呈现先升后降的趋势。在进水曝气阶段,取样点1 NH4+-N浓度由3.80 mg·L−1升至10.72 mg·L−1,而NO3−-N浓度仅由1.85 mg·L−1升至2.94 mg·L−1,这一结果由原水和硝化反应联合贡献。原水会带入一定量的NH4+-N,但此时硝化能力有限,无法及时将NH4+-N氧化为NO3−-N,于是NH4+-N浓度不断升高。对NO3−-N而言,原水会对其进行稀释,但伴随着硝化反应的进行,NO3−-N会不断积累,因而NO3−-N浓度略有增加,但会明显低于NH4+-N浓度的增加;在曝气阶段,若仅存在硝化反应,NH4+-N的下降将同NO3−-N的升高相匹配。但实际情况是,NH4+-N和 NO3−-N浓度同时下降,在曝气结束时分别降至4.09 mg·L−1和5.29 mg·L−1,此结果证实了同步硝化-反硝化脱氮的存在。由于取样点3和5距进水口有一定距离,推流式进水方式使得其起始NH4+-N和COD较低,进而DO浓度快速升高。而当进水逐渐推移到这两点时(取样点3为20~40 min;取样点5为40~60 min),由于COD降低和硝化反应的进行,DO浓度开始下降,之后随着NH4+-N和COD的降低,DO浓度开始缓慢回升。依据计算可以得出,同步硝化-反硝化、内源反硝化和反硝化途径对TN的脱除贡献率分别为65.7%、32.5%和1.80%。依据计算可得,同步硝化-反硝化、内源反硝化和反硝化途径对TN的脱除贡献分别为65.7%、32.5%和1.80%(进水和出水TN分别为38.80 mg·L−1和6.01 mg·L−1, TN去除率为84.51%)。
图3(e)~(f)为高DO浓度(1~6 mg·L−1)下同步硝化-反硝化脱氮特性。由图3(e)和图3(f)可知,3个取样点的 DO 浓度随时间变化规律与中 DO 水平下的变化规律基本一致,但其浓度均高于后者,取样点1、3、5的DO平均浓度分别为3.41、4.48和6.07 mg·L−1。在高DO浓度条件下,氧扩散能力增强,活性污泥絮体内部难以维持缺氧环境,同步硝化-反硝化脱氮过程所需环境遭到破坏,致使NO3−-N不断积累,曝气结束时3个取样点的NH4+-N和NO3−-N浓度依次达到1.41、1.27、0.58 mg·L−1和13.92、12.48、9.88 mg·L−1。依据计算可得,同步硝化-反硝化、内源反硝化和反硝化途径对TN的脱除贡献分别为39.2%、58.6%和2.20%(进水和出水TN分别为40.01和10.20 mg·L−1,TN去除率为74.51%)。
综上所述,中DO浓度下可观察到明显的同步硝化-反硝化过程,同步硝化-反硝化脱氮效果最理想,而高DO和低DO浓度下无法观察到明显的同步硝化-反硝化过程,脱氮效果较差。因此,CAST工艺好氧池DO应控制为中等浓度,这对同步硝化-反硝化过程进行更有利,从而提高TN去除率。
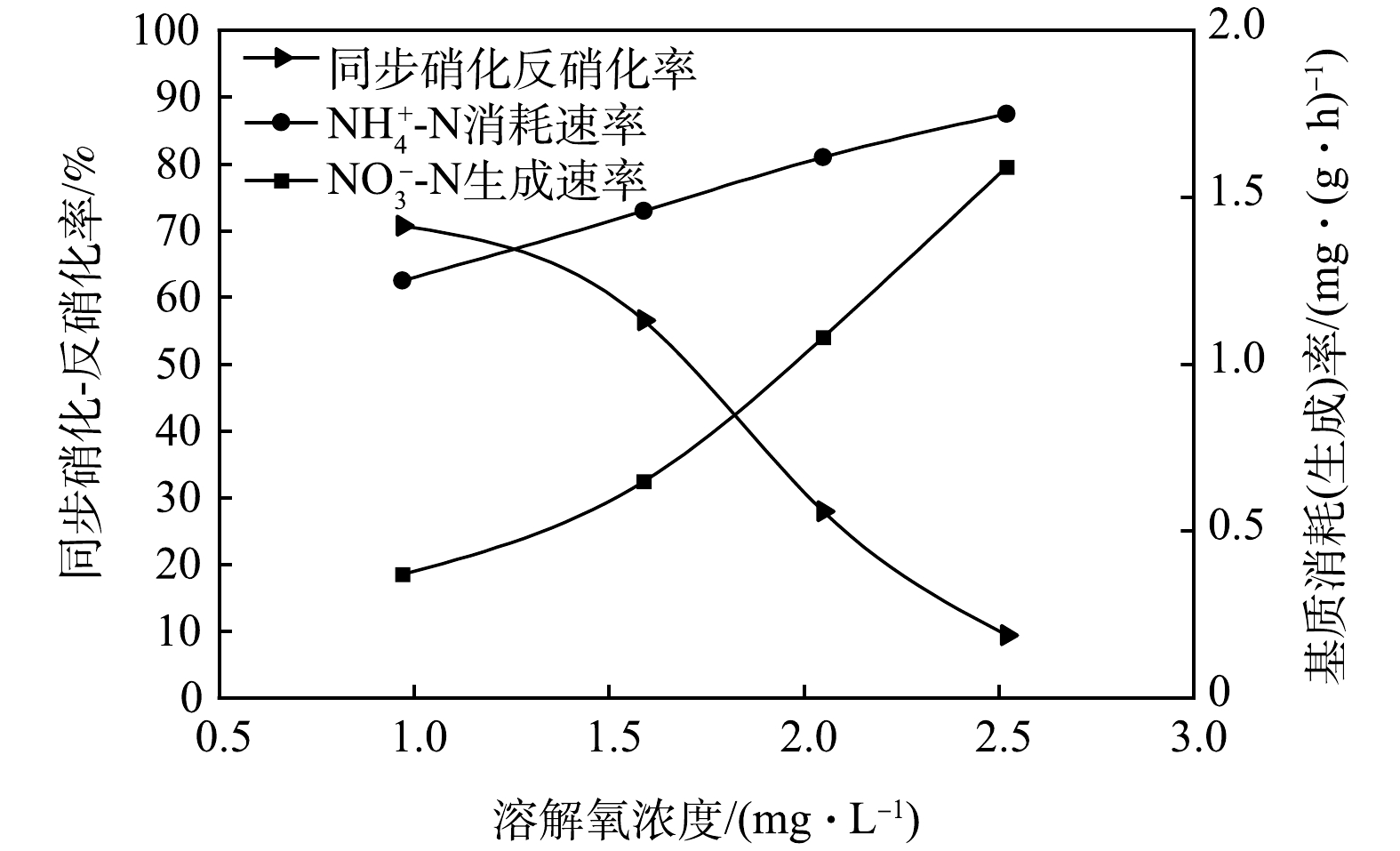
基于不同DO浓度下典型周期内同步硝化-反硝化对TN脱除的贡献变化,特进行同步硝化-反硝化率测定,以确定同步硝化-反硝化脱氮的最适宜DO浓度。在CAST反应池进水结束后取取样点1的混合液,利用序批式反应器对不同DO浓度(1.0~2.5 mg·L−1)下的同步硝化-反硝化率进行测定,结果如图4所示。由图4可知,随着DO浓度升高,同步硝化-反硝化率逐渐下降。当DO浓度控制在 1 mg·L−1 时,同步硝化-反硝化率达到最高值(70.66%),NH4+-N消耗速率和NO3−-N生成速率分别为1.25 mg·(g·h)−1和0.37 mg·(g·h)−1。而当DO浓度控制在2.52 mg·L−1时,同步硝化-反硝化率降为9.34%,NH4+-N消耗速率和NO3−-N生成速率分别为1.75 mg·(g·h)−1和1.59 mg·(g·h)−1。相比较而言,随着DO浓度的升高,NH4+-N消耗速率增幅并不明显,仅为40%,但NO3−-N生成速率增幅明显,达到329.73%。这表明在DO浓度为2.52 mg·L−1时同步硝化-反硝化过程已严重受限。因此,CAST工艺DO浓度应控制在1.0~1.5 mg·L−1,此时同步硝化-反硝化性能最强,这一结果与实际运行过程内中等DO浓度下所得结果一致,同样符合徐凤等[12]认为的最佳溶解氧浓度(0~2mg·L−1)的观点。
-
CAST工艺中20%的混合液回流比使得回流至CAST反应池前端的污泥有限,且污泥在好氧池内以推流式方式前进,污泥从好氧池末端回流至前端理论需要3~5 d,于是微生物将长期处于饥饿状态,即CAST工艺运行具有饱食-饥饿的特性,即非平衡增长[13-14]。饱食-饥饿理论[15]认为污泥所处饥饿时长会影响胞内聚合物的储存量,而同步硝化-反硝化脱氮能力又取决于胞内聚合物的量。因此,确定合适的饥饿时长,进而明确合适的污泥回流比对提高CAST工艺脱氮效率尤为重要。
为探究饥饿时长对微生物吸收有机物的影响,进行批次实验,结果见表3。由表3可知,随着饥饿时长增加,活性污泥对HAc的吸收量呈现出先上升后下降的趋势。在饥饿时长为5 h时
ΔMHAc 最小,仅为0.123 g·g−1;随着饥饿时长的增加,活性污泥内原有细胞聚合物被逐渐消耗,使得活性污泥对HAc的吸收能力和储存胞内聚合物的空间得以提升,当饥饿时长为36 h时ΔMHAc 达到最大,为0.173 g·g−1;随着饥饿时长的进一步增加,微生物开始利用细胞体进行反硝化,导致参与吸收的微生物数量有所减少(可检测到NH4+-N浓度升高),进而吸收量减小,当饥饿时长为72 h时ΔMHAc 仅有0.173 g·g−1。综上所述,最佳饥饿时长应控制为36 h,此时的HAc吸收量最大,更有利于同步硝化-反硝化脱氮。本研究中CAST工艺好氧池有效容积为4 254 m3,若活性污泥停留时间为36 h,则每天由好氧池末端回流至选择池的污泥量为3 016 m3,CAST工艺每天运行6个周期,每周期回流量为502 m3,基于每周期进水量为1 125 m3,CAST工艺的最佳回流比应控制为45%,这对好氧池的同步硝化-反硝化和内源反硝化脱氮过程更有利。 -
为了解CAST工艺好氧池内源反硝化脱氮特性,在曝气结束时取1、3和5取样点的泥水混合物,对其内源反硝化活性进行了测定,结果见图5。由图5可知,取样点1泥水混合物内源反硝化活性最大,达到1.48 mg·(g·h)−1,明显高于取样点3(1.08 mg·(g·h)−1)和取样点5 (1.05 mg·(g·h)−1)。造成这一结果的原因是取样点1靠近进水端,活性污泥所吸附的有机物及微生物储存的胞内聚合物较多,所以可用于内源反硝化反应的电子供体较多。而对于取样点3和取样点5的活性污泥,较长的污泥龄导致外源有机物和胞内聚合物已被大量消耗,内源反硝化反应只能以细胞作为碳源进行,致使内源反硝化速率较低。内源反硝化主要是利用内碳源(细胞体)进行反硝化,细胞体的化学结构通式为C5H7NO2,化学计量反应如式(13)所示。
由式(13)可知,每去除1 g的NO3−-N理论上会产生0.25 g的NH4+-N。而在CAST工艺中,取样点1、3和5每去除1 g的NO3−-N依次产生0.09、0.13和0.20 g NH4+-N,均低于利用细胞体进行反硝化的理论值,但沿着推流方向逐渐接近理论值。此结果证实内源反硝化过程不单以细胞体作为碳源,还包括污泥所吸附的有机物及微生物储存的胞内聚合为碳源。
-
表4列举了CAST、A2/O和氧化沟工艺的脱氮指标。由表4可知,3种工艺出水COD、NH4+-N和TN浓度相当,均满足相关排放要求。而对于出水TN,CAST、A2/O和氧化沟工艺的去除量分别为29.1、36.06和39.03 mg,每去除1 mg TN所需COD依次为5.77、6.41和15.94 mg,此结果说明CAST工艺更具脱氮潜力,但考虑到CAST工艺出水TN明显高于A2/O和氧化沟工艺,因而对其运行提出以下调控建议:1)DO浓度调控:依据现场测定和同步硝化-反硝化率测定实验,为获得良好的同步硝化-反硝化脱氮性能(较高同步硝化-反硝化率)及降低运行费用(减少供氧量),在CAST工艺好氧池内实行阶梯曝气,沿泥水混合物流动方向不断升增加曝气量,控制好氧池前段、中段和后段的DO质量浓度依次处于0.6~1.0、1.0~2.0和2~3 mg·L−1。2)污泥回流比调控:依据饱食-饥饿实验,为保证微生物将进水有机物尽可能多的以内碳源的形式储存,从而增强同步硝化-反硝化脱氮性能,将CAST工艺的回流比由当下的20%(饥饿时长为80 h)提升至45%(饥饿时长为36 h)。
-
1) CAST工艺好氧池内同步硝化-反硝化和沉淀过程中内源反硝化对总氮去除的贡献占据主导作用,贡献率分别为 (35.50±4.15)%和(62.86±4.13)%,而缺氧池反硝化脱氮贡献仅为(1. 64±0.05)%。
2) DO浓度对同步硝化-反硝化脱氮性能有重要影响,控制DO浓度在1.0~1.5 mg·L−1时,可获得最大同步硝化-反硝化脱氮效果。
3)饥饿时长对微生物吸收HAc有重要影响,当饥饿时长为36 h时,可获得最大HAc吸收量,达到每1 g VSS消耗0.173 g HAc,可为同步硝化-反硝化脱氮提供更多碳源。
某污水处理厂CAST工艺的脱氮性能评价
Evaluation of nitrogen removal performance of the CAST process in a wastewater treatment plant
-
摘要: 通过对某污水处理厂循环活性污泥法工艺(cyclic activated sludge technology, CAST)中选择池、缺氧池和好氧池中氮组分和污泥浓度进行测定,结合污泥活性、同步硝化-反硝化(simultaneous nitrification and denitrification, SND)速率及饱食-饥饿(feast-famine)批次实验,评价该处理工艺的脱氮性能。结果表明,好氧池内同步硝化-反硝化和沉淀过程中的内源反硝化(endogenous denitrification, ED)脱氮对总氮去除的贡献占据主导,分别为(35.50±4.15)%和(62.86±4.13)%,而缺氧池反硝化(DEN)脱氮贡献仅为(1.64±0.05)%;溶解氧(dissolved oxygen, DO)浓度对CAST工艺脱氮性能有极大影响,控制好氧池中DO浓度为1~1.5 mg·L−1时可获得最佳脱氮效果,CAST工艺的TN去除率可达84.51%;饱食-饥饿批次实验证明,饥饿时长为36 h时对乙酸(HAc)的吸收能力最强,可达每1 g VSS消耗0.173 g HAc,依此可推算出CAST工艺的最佳回流比为45%。Abstract: The nitrogen removal performance of CAST process in a wastewater treatment was evaluated by measuring the nitrogen components and suspended solids in the selective, anoxic and aerobic tanks, combined with the simultaneous nitrification and denitrification (SND) rate and feast-famine batch tests. The results showed that the contribution to total nitrogen(TN) removal through SND in aerobic tank and endogenous denitrification at sedimentation stage were dominated, which were (35.50±4.15)% and (62.86±4.13)%, respectively, while the denitrification in anoxic tank only accounted for (1.64±0.05)%. The Dissolved oxygen (DO) concentration had a great influence on the nitrogen removal performance of CAST process. The best nitrogen removal effect happened when the DO concentration in aerobic pool was controlled at 1~1.5 mg·L−1, and the TN removal rate of CAST process could reach 84.51%. The Feast-Famine batch tests proved that the maximum absorption of HAc occurred at the famine duration of 36 h, being up to 0.173 g HAc per 1 g VSS consumption. Based on this result, the optimal reflux ratio could be estimated to be 45%.
-
碳酸酐酶(CA)是一种普遍存在的含锌金属酶,其主要功能是催化CO2的水合反应,反应式如下所示:CO2+H2O↔HCO3−+H+,它是目前已知的催化最快的酶之一[1]. 由于游离CA在高温、强酸或强碱溶剂和化学杂质等条件下容易失活,限制了其大规模的工业应用. 因此,采用分子修饰[2]、定向进化[3]、酶固定化[4]等策略来提高其稳定性和活性. 在上述方法中,酶固定化因其操作简单、方便的特点而被广泛应用.
通过选择良好的载体和合适的固定化方法对游离CA进行固定是降低催化剂成本、提高稳定性和可重复使用性的有效途径[5]. 基于固定化CA的生物催化剂因其低耗能和高效率被认为是转化CO2的环境友好的候选方法. 固定化CA主要通过酶促反应吸收[6],CO2矿化生成金属碳酸盐[7]和多酶级联反应[8]来进行CO2转化. 吸收是目前应用最为广泛的CO2捕集技术之一[6]. 然而,吸收溶剂具有再生所需能量高、吸收速率慢和稳定性低等缺点[9]. CA作为促进剂可以提高CO2吸收反应速率,从而降低反应器尺寸和成本. 在某些情况下,CA还用于促进酶催化的CO2转化. CA参与的多酶级联反应用于生产甲酸盐和甲醇等高价值产品[7].
无论CA是以何种方式转化CO2,CO2转化实验始终以烟气为基础,而烟气中具有高温和其他化合物(NOx和SOx等)[10]. 这些可能会影响CA的活性和稳定性,进而减缓CA转化CO2实际工业应用中的反应速率.
1. 碳酸酐酶的性质和催化机理(Properties and catalytic mechanism of carbonic anhydrase)
1.1 碳酸酐酶的性质
CA广泛存在于细菌、真菌和藻类等生物体内,分为不同的类别(α、β、γ、δ、ζ、η和θ)[11]. 大多数CA从牛血红细胞中提取,少数CA从羊和猪等哺乳动物的肝脏以及重组细菌中获得[12]. CA是一种以锌离子作为活性位点的金属酶,在某些情况下,也可以选择其他金属原子,如Cd2+、Co2+、Fe2+或Mn2+来代替Zn2+[13-14],催化各种水解反应,包括羧酸、卤化物、尿素和可水解底物的水合作用[15]. 虽然相同的催化反应由不同的CAs进行,但它们既没有结构相似性,也没有其他相关性[16].
1.2 碳酸酐酶的催化CO2水合机理
在自然状态下,CO2的可逆水合反应十分缓慢,一级反应速率常数仅为5×10−2 s−1,反应步骤如下所示[17]:
(1)CO2水合生成H2CO3:
CO2(aq)+H2O↔H2CO3 (1) (2)H2CO3电离生成
HCO−3 CO2−3 H2CO3↔H++HCO−3 (2) HCO−3↔H++CO2−3 (3) 反应式(1)是CO2水合反应的限速步骤,也是生物系统中碳矿化和碳再利用技术的切入点[18]. 随着CA的引入,Kcat值可高达1.6×106 s–1[19],说明CA可以高效催化CO2的可逆水合反应. 与此同时,CO2水合反应的机制发生改变,变为以下两个步骤[20]:
第一步是CA中与活性中心的Zn2+相连的H2O去质子化形成中间产物EZnOH−,随后CO2被EZnOH−中的OH−水合形成EZn
HCO−3 HCO−3 HCO−3 HCO−3 第二步是EZnH2O的去质子化:EZnH2O通过酶分子中的质子转运体将质子(H+)转运至溶剂中并被还原成具有催化活性的EZnOH−.
2. 碳酸酐酶的固定化(Immobilization of carbonic anhydrase)
酶固定化的催化活性和稳定性在很大程度上取决于载体和固定化方法的选择,首先应在固定化过程中保持酶活性[21]. 近年来,各种材料已被用作固定化CA的载体,包括水凝胶[22]、壳聚糖[23]、二氧化硅[19]和金属有机框架(MOFs)[24]等. 酶可以通过多种方法固定在载体上,例如包埋、吸附、交联、共价结合、交联酶聚集体(CLEAs)[25]和纳米花型酶-无机杂化固定化酶(简称纳米花)[26]等. CA固定化方法优缺点如表1所示.
表 1 CA固定化方法优缺点Table 1. Advantages and disadvantages of CA immobilization methods固定化方法Immobilization methods 优点Advantages 缺点Disadvantages 参考文献Reference 吸附 易于操作;固定化酶构象变化小 酶易从载体中泄漏 [6] 共价结合 酶和载体之间紧密结合;提高了固定化酶的稳定性 引起酶构象的变化;酶的活性通常在固定化后降低 [7] 包埋 操作简单;可以减少酶浸出和变性 酶被限制在载体材料中,减弱了酶的表观活力 [6] 交联酶聚集体 无需载体;对酶纯度要求不高 机械强度不高 [21] 纳米花 固定化酶具有高比表面积和高酶活性 纳米花晶体的生长时间较长 [26] | Show Table DownLoad:
CSV
DownLoad:
CSV
吸附是一种简单有效的固定化方法,通过氢键、疏水键、离子键和范德华力的相互作用将酶固定在载体表面. 在多数情况下,与载体的温和相互作用可确保固定后酶的微小构象变化[27],这在很大程度上取决于载体的性质. Yin等[24]结合了磁性材料易于回收和可重复使用性,以及MOF具有高孔隙率、大比表面积和良好稳定性的优点,构建了核壳磁性ZIF-8@Fe3O4复合材料. CA通过吸附固定在ZIF-8@Fe3O4上. 游离CA和CA@ZIF-8@Fe3O4分别在40 ℃的1 mol·L−1 MDEA中储存9 d,CA@ZIF-8@Fe3O4的活性随着储存时间的延长而逐渐降低,但其活性始终高于等量游离CA. 包埋是一种将酶限制在载体材料中的固定化方法,可以有效防止酶浸出和变性[6]. Hsieh等[19]将二氧化硅缩合肽(R5)与硫化氢碳酸酐酶(SazCA)融合形成R5-SazCA,再把R5-SazCA包埋到二氧化硅纳米颗粒(R5-SazCA-SP)中. 在70 ℃培养3 h后,R5-SazCA和R5-SazCA-SP的残余活性分别为49%和60%,表明CA固定化后热稳定性得到改善. 增强的热稳定性通常归因于二氧化硅交联网络施加的空间约束,这有助于维持CA的立体结构.
共价结合是一种常用的固定化方法,可以显著增强和提高酶的稳定性. 改性载体表面的胺基、羟基、羧基或环氧基等官能团和CA的氨基酸残基之间可以形成共价键,从而使得酶和载体中间具有很强的相互作用[28]. Shamna等[29]将CA通过共价结合固定在胺官能化的铝-硅氧烷气凝胶珠(BCA-Al/Si-NH2)上. BCA-Al/Si-NH2杂化珠的动力学参数与游离酶相当. 固定化CA具有热稳定性、pH稳定性和储存稳定性,这是由于酶的多点附着和酶与载体之间的相互作用导致酶刚性增加,从而减少了构象变化. Sun等[30]将CA通过戊二醛(GA)共价连接到聚偏二氟乙烯(PVDF)膜和聚乙烯膜(PE)的表面,这些膜固定化CA之前通过聚乙烯亚胺(PEI)和多巴胺(DA)的共沉淀进行了修饰. PEI/PDA-PE的活性回收率高于PEI/PDA-PVDF,因为PE具有更大的孔结构和改性后更好的亲水性,从而引起更高的酶负载量和更好的酶亲和力. 此外,固定化后,CA的储存稳定性和可重复使用性得到很大提高.
考虑到载体材料的物理和化学性质、操作条件等,通常需要多种固定化方法的协同作用从而获得高效的固定化CA[6]. Chang等[31]将CA通过吸附-交联法固定在地质聚合物微球上,与游离CA相比,固定化CA的最适pH值从7.5增加到8.0,最适温度从25 ℃增加到30 ℃. Effendi等[11]将Sulfurihydrogenibium yellowstonense CA(SyCA)共价结合在新型聚丙烯腈(PAN)和聚对苯二甲酸乙二醇酯(PET)纳米纤维上,所得复合材料通过戊二醛(GA)进一步交联. 固定化粗SyCA在60 ℃下保留了超过100%的相对残余活性,这归因于粗SyCA的胺基和纳米纤维的羟基之间的共价结合,而且固定化粗SyCA和GA之间交联也会形成共价键. 此外,测定了固定化CA对SOx和NOx的耐受性. 固定化SyCA在50 mmol·L−1 HNO3和H2SO4存在下的残余酶活性分别为57.1%和61.6%.
CLEAs是一种无需载体的新型固定化酶制备技术,该方法包括沉淀和交联两个主要步骤[26]:(1)通过沉淀剂(盐、水溶性有机溶剂或非离子聚合物)在溶液中形成酶聚集体;(2)聚集体中酶分子通过交联剂交联,使它们在去除沉淀剂后不溶. 然而,对于工业应用来说,CLEA柔软且容易断裂,为了提高CLEA的机械性能,Jun等[32]使用纳米纤维共价连接bCA,然后交联并进行沉淀,以获得bCA沉淀涂层(EPC). EPC在剧烈摇动下868 d后仍保留其活性的65.3%,EPC的高稳定性可以通过酶沉淀和交联的结合来解释,使酶分子之间通过化学键紧密的链接,从而有效防止酶变性和浸出. 纳米花型酶-无机杂化固定化酶由2012年Ge等[33]首次提出,通过将酶与硫酸铜在磷酸盐缓冲溶液中共沉淀,从而形成典型的花瓣状结构. Wen等[22]对此固定化方法进一步研究,将双金属离子(Cu2+和Zn2+)代替单个金属离子来固定CA,然后将双金属基杂化纳米花(CANF)复合材料嵌入到聚乙烯醇(PVA)/壳聚糖(CS)水凝胶膜(PVA/CS@CANF)中. 水凝胶膜在8个循环后仍保持其原始活性的75%以上,在60 ℃下仍有超过80%的相对活性. 此外,PVA/CS@CANF膜转化CO2产生的CaCO3量是游离CA的9倍.
3. CA转化CO2的应用(Application of CA to CO2 conversion)
二氧化碳排放是造成温室效应的主要原因. 当前,CO2捕集、利用和封存(CCUS)技术已广泛应用于减少CO2排放到大气中. CCUS技术将CO2资源化,便于运输和长期储存[29]. 基于CA的生物酶法通过捕集大气或废气中的CO2,并进一步转化为高价值的工业化学品和燃料,如金属碳酸盐、甲醇和甲酸等.
3.1 CA在CO2矿化中的应用
CA催化的CO2水合反应生成的碳酸氢盐可进一步用作碳源,用于合成无机碳酸盐和促进藻类的生长. CA将CO2矿化生成CaCO3的原理为CA首先催化CO2水合形成碳酸氢根离子(4),然后与添加的Ca2+反应生成CaCO3(5),CaCO3易于分离和储存以供进一步利用. Shamna等[29]将CA共价连接在铝-硅氧烷气凝胶珠上,并进一步检测了其将CO2转化为CaCO3的能力:1 mg游离bCA和17 μg bCA-Al/Si-NH2生成CaCO3质量为32.4 mg和12 mg. 通过XRD表明CA转化的碳酸钙的晶相为方解石,且CaCO3的晶体结构不会因添加酶而改变. Jun等[32]使用纳米纤维共价固定bCA,然后进行交联并沉淀,以获得bCA沉淀涂层(EPC). CA通过催化系统中鼓泡的CO2的可逆水合生成碳酸氢盐,为微藻提供光合作用的碳源. 添加 EPC的系统与含有25 mmol·L−1 NaHCO3和不含碳源的相比,分别使微藻生长加速了134%和231%. Xu等[34]将CA与戊二醛(GA)交联并包埋在漂浮的海藻酸钙水凝胶珠中,并将珠子保留在微藻-大气界面便于直接从空气中捕集CO2. 与(22.7±0.5) mg·L−1每天的自然生长速率相比,游离CA和CA-GA珠粒分别将微藻的生长速率提高到(37±3)mg·L−1每天和(40±1)mg·L−1每天,而且珠子容易回收和重复使用,具有广阔的工业前景.
CO2+H2O→HCO−3+H+ (4) Ca2++HCO−3→CaCO3+H+ (5) 当前通过全细胞固定化构建了用于CO2捕集的新型全细胞催化剂,避免酶泄漏和细胞裂解从而提高酶的稳定性和可重复使用性[7]. Sharma等[35]将含CA的苦参棒状杆菌菌株的细胞共价固定到戊二醛功能化的角蛋白颗粒上. 含有144 U·mg−1 CA的菌株细胞裂解物形成65.12 mg CaCO3. 进行10次CO2转化后,固定化细胞和游离细胞CaCO3的相对产量分别为53.46%和22.15%. Moon等[36]将周质水生弧菌CA(hmCA)的全细胞催化剂通过包埋固定在聚氨酯泡沫上(pCA-PUF),经9次循环使用后,pCA-PUF的高相对活性没有明显下降,并且显示出良好的可重复使用性. 使用该固定化生物催化剂将设计的填充床反应器中的CO2捕集速率提高了80%. 微生物细胞表面展示是将某一蛋白或短肽(靶蛋白)与微生物细胞的外膜蛋白(载体蛋白)以融合蛋白的形式锚定在微生物细胞表面的技术. 大肠杆菌是生产重组蛋白最常用的宿主细胞之一[37]. Zhu等[38]以大肠杆菌为宿主细胞,通过将SazCA作为靶蛋白与冰核蛋白(INPN)(载体蛋白)锚定在细胞外膜上构建表面展示菌株. 工程菌株矿化CO2产生的CaCO3质量(241 mg)明显高于胞内表达菌株(173 mg). Tan等[39]将(Mesorhizobium loti CA)MlCA克隆到3个载体中且在大肠杆菌中表达,并将离心获得的全细胞样品固定在琼脂上. 结果表明,全细胞生物催化剂具有良好的稳定性,储存40 d后相对活性保持近100%. 此外,具有固定化全细胞生物催化剂的系统可在3.5 min内有效地将CO2100%转化为CaCO3. 通过表2展示了固定化CA矿化CO2性能的比较.
表 2 CA在二氧化碳捕集中的矿化性能Table 2. Mineralization performance of CA in carbon dioxide capture碳酸酐酶种类Types of carbonic anhydrase 矿化特性Mineralization properties CaCO3晶相CaCO3 crystal phase 参考文献Reference 牛CA(bCA) 固定化细胞比游离细胞的CaCO3产量增加了1.35倍;固定化细胞和游离细胞的CaCO3 相对产量在10个循环后分别为53.46%和22.15% 球霰石和方解石 [35] bCA 17 μg固定化CA产生12 mg CaCO3 方解石 [29] CA CA@ZIF-8获得的CaCO3的产量是游离 CA的22倍 球霰石 [40] 硫化氢碳酸酐酶(SazCA) 固定化CA转化CO2形成碳酸钙的时间比游离CA缩短了33% 球霰石和方解石 [19] SazCA 工程菌株矿化CO2产生的CaCO3质量(241 mg)高于细胞内表达菌株(173 mg) — [38] 周质水生弧菌CA(hmCA) 固定化CA将填充床反应器中的CO2捕集速率提高了80% — [36] CA PVA/CS@CANF膜转化CO2产生的CaCO3 量是游离CA的9倍 — [22] bCA 添加EPC的系统与含有25 mmol·L−1 NaHCO3和不含碳源的对照相比,分别使微藻生长加速了134%和231% — [32] bCA 与(22.7±0.5)mg·L−1·d−1的自然生长速率相比,游离CA和CA-GA珠粒分别将微球藻的生长速率提高到(37±3)mg·L−1·d−1和(40±1)mg·L−1·d−1 — [34] Sulfurihydrogenibium yellowstonense CA(SyCA) 固定化粗CA的CaCO3总产量是游离CA的5.8倍 — [11] CA 固定化CA产生的CaCO3质量是空白实验的5倍 — [31] bCA CA@ZIF-8和PVA/CS/CA@ZIF-8水凝胶膜获得的CaCO3产量分别为游离CA的20倍和32.6倍. 球霰石和方解石 [41] Mesorhizobium loti CA(MICA) 具有固定化全细胞生物催化剂的系统可在3.5 min内有效地将CO2100%转化为CaCO3 — [39] | Show TableDownLoad:
CSV
3.2 液体吸收法捕集CO2
到目前为止,已经开发了使用纯水、碳酸盐溶液(Na2CO3和K2CO3等)和醇胺溶液等吸收二氧化碳的工艺. 常用于捕集CO2的醇胺溶液主要有伯胺(一乙醇胺,MEA)、仲胺(二乙醇胺-DEA)、叔胺(N-甲基二乙醇胺-MDEA、二异丙醇胺-ADIP和三乙醇胺-TEA)和哌嗪(PZ)等[42]. MEA吸收速度快,不易挥发,但是溶剂在汽提塔中的解吸需要大量的能量,导致电费成倍增加,需要对其进行改进. 与MEA相比,K2CO3溶液具有低毒性,不易腐蚀,在高温下高效且低成本的再生等优点,而且MDEA再生能耗低、吸收CO2容量大. 但是K2CO3溶液和MDEA吸收速率较慢,添加CA可显著促进MDEA和K2CO3溶液吸收CO2[17].
当前,固定化CA已用于纯水中吸收二氧化碳研究. Chang等[31]将CA通过吸附-交联法固定在地质聚合物微球上. 采用水浴设定不同温度,pH计可监测反应溶液pH值随不同催化反应条件的变化,从而评价固定化CAs的催化性能. 当温度为30 ℃,气体流速为300 mL·min−1时,溶液的pH值明显下降. Xu等[43]将CA通过交联固定在聚多巴胺(PDA)/聚乙烯亚胺(PEI)修饰的聚偏氟乙烯(PVDF)膜上(简称CA-m-PVDF复合膜),在气液膜接触器中使用速率为0.25 m·s−1的水作为吸收剂,CA-m-PVDF复合膜具有较高的CO2通量值(2.5×10−3 mol·m−2·s−1),该CO2通量值比未添加CA时增加约160%.
MDEA吸收CO2的过程中形成不稳定的氨基甲酸盐和碳酸氢根离子,加热时释放出CO2达到解吸和再生的目的. Zhang等[44]将CA通过吸附固定到ZIF-L-1(MOFs)上,得到CA/ZIF-L-1复合材料. 由于ZIF-L-1和CA均含有Zn2+,两者之间存在协同作用,ZIF-L-1中的咪唑基团(mIm)可以作为亲核试剂参与CO2的水合生成碳酸氢盐(6),而且该反应能够促进CO2的液-固传质. 加入0.05 g·L−1 CA/ZIF-L-1后,MDEA中的CO2吸收率提高了2.5倍. Du等[45]进行了类似的研究,把CA包埋到不同粒径的ZIF-8(CA/ZIF-8)中,并将CO2吸收到MDEA中,MDEA的吸收率增加到2.4倍. Xu等[46]将CA固定在表面改性的磁性Fe3O4纳米颗粒(MNP)上,并用原位聚合合成MNP-CA纳米凝胶. 通过反应器测试了MNP-CA纳米凝胶加速MDEA水溶液中CO2吸收的有效性,MNP-CA纳米凝胶在30 ℃的CO2吸收率为不含CA时的170%. 而且MNP-CA纳米凝胶的加入使湿壁柱气相中的总传质系数(KG)在60 ℃时增加了4.61倍. 费潇瑶[47]使用共价结合法将氨基功能化介孔材料固定在CA上(CA/AFS)对用于MDEA溶液解吸CO2进行了研究,固定化CA的加入对CO2平衡解吸量基本没有影响,但CA/AFS的加入可以在未达到平衡状态之前,提高CO2的瞬时解吸量.
H2O+mIm+CO2↔mIm++HCO−3 (6) CO2在碳酸盐溶液中的总反应如下所示(7),CA可有效促进二氧化碳吸收到碳酸盐溶液中[9]. Fabbricino等[48]将嗜热细菌的碳酸酐酶(SspCA)作为膜锚定蛋白固定在大肠杆菌细胞的外膜(INPN-SspCA)上. 在碱性溶液(0.5 mol·L−1 Na2CO3/0.5 mol·L−1 NaHCO3)吸收CO2反应过程中,加入SspCA的膜细胞碎片用作生物催化剂,观察到压力值的迅速下降和比未添加催化剂时更高的CO2吸收率. Peirce等[49]报道了CA在高浓度碳酸盐溶液中容易失活,因此将工业级碳酸酐酶(NovoCA)固定在顺磁性Fe3O4纳米颗粒上进行改善,并分别在25 ℃和40 ℃的条件下,于0.5 mol·L−1 Na2CO3/NaHCO3缓冲液(pH=9.6)和10%wt K2CO3溶液的搅拌池反应器中进行CO2吸收测试[10]. 溶液中固定化CA的Kcat/Km均在5.52×10−1—5.52 L·mol−1·s−1之间. Qi等[50]通过湿壁柱测定表明,在20% wt K2CO3溶剂中加入2 g·L−1 CA后,CO2总传质系数(KG)提高了约5倍. 当CA浓度从零增加到2.5 g·L−1时,CO2捕集效率增加了4.6倍.
CO2−3+H2O+CO2↔2HCO−3 (7) Gladi等[17]探究了在湿壁柱中对4种溶液(MEA、AMP(2-氨基-2-甲基-1-丙醇)、MDEA和K2CO3溶液)添加0.2% wt CA后吸收CO2的影响. 研究表明,添加CA显著增加了MDEA和K2CO3的液侧传质膜系数,AMP略微增加,而MEA没有变化. MDEA的反应速率随温度升高而降低,而在较低的溶剂浓度(5% wt—15 %wt)下,温度和溶剂浓度对K2CO3中反应速率的影响较小,但在20% wt时,温度升高会显着提高反应速率. 此外,混合溶剂对CO2进行吸收比单一溶剂表现出更高的吸收效率,因为它们结合了单一溶剂的优点. Sahoo等[51]将CA共价固定到异质功能化载体(HFS)和锌络合物(Zn-Im)上(CA/Zn-Im:HFS),并将CA/Zn-Im:HFS加入到混合溶剂(30% wt MEA+7.5% wt PZ+15% wt K2CO3)中加快CO2的吸收和解吸. 含有CA/Zn-Im: HFS的混合溶剂(21.65 %wt)在20 min内CO2的吸收量高于纯混合溶剂(13.75 %wt). 含有CA/Zn-Im: HFS的混合溶剂比纯混合溶剂的CO2相对解吸率高1.57倍. 通过表3展示了CA在二氧化碳吸收中的催化性能.
表 3 CA在二氧化碳吸收中的催化性能Table 3. Catalytic performance of CA in carbon dioxide absorption吸收剂成分Absorbent ingredients 温度Temperature 催化特性Catalytic properties 参考文献References 100 mg的固定化CA(固载量为2 mg·g−1)+H2O 30 ℃ 反应进行30 min,CO2吸收量Gv=1.858×10−4 L [31] CA-m-PVDF复合膜+0.25 m·s−1 H2O 25 ℃ CA-m-PVDF复合膜CO2通量值为2.5×10−3 mol·m−2·s−1 [43] 0.05 g·L−1 CA/ZIF-L-1+20 mL 1 mol·L−1 MDEA 40 ℃ MDEA中的CO2吸收率提高了2.5倍 [44] CA/ZIF-8+MDEA 40 ℃ CO2吸收到含有不同粒径的ZIF-8的MDEA溶液中,MDEA溶液的吸收率增加到2.4倍 [45] 10 mg MNP-CA纳米凝胶+100 mL 1 mol·L−1 MDEA 30 ℃ 反应3 h后,MNP-CA纳米凝胶的CO2吸收率为不含CA时的170% [46] 0.3 kg m−3工业级碳酸酐酶(NovoCA)+2—3 mol·L−1 K2CO3(碳酸盐转化度0—40%) 25 ℃和40 ℃ 当加入的酶浓度低于0.018 kg·m−3时,Kcat/Km值介于0.50×102 L·mol−1·s−1和0.39×103 L·mol−1·s−1之间 [49] 0.2×10−2—1.5×10−2 kg NovoCA·m−3 Fe3O4纳米颗粒固定化CA+0.5 mol·L−1 Na2CO3/NaHCO3缓冲液(pH=9.6) 25 ℃和40 ℃ 固定化CA的Kcat/Km值介于4.87—8.06 L·mol−1·s−1之间 [10] 0.2×10−2—1.5×10−2 kg NovoCA·m−3 Fe3O4纳米颗粒固定化CA+10%wt K2CO3溶液(碳酸盐转化度0—40%) 25 ℃ 固定化CA的Kcat/Km值介于3.24—6.73 L·mol−1·s−1之间 [10] 嗜热细菌的全细胞固定(INPN-SspCA)+0.5 mol·L−1 Na2CO3/NaHCO3 25 ℃ INPN-SspCA的Kcat/Km值介于9.94×10−1—3.09 L·mol−1·s−1之间 [48] 2 g·L−1 CA+20% wt K2CO3 40 ℃ 与未添加CA相比,CO2总传质系数(KG)提高了约5倍 [50] 0.22 g·L−1 CA+30% wt K2CO3(pH~11—12) 50 ℃ CA的Kcat/Km值为5.3×108 L·mol−1·s−1 [52] 0.2% wt CA+30% MDEA/15% K2CO3 17—50 ℃ 添加CA显著增加了MDEA和K2CO3的液侧传质膜系数 [17] 0.2% wt CA+30% wt MEA + 7.5% wt PZ+15% wt K2CO3 30 ℃ 含有CA/Zn-Im: HFS的混合溶剂(21.65% wt)在20 min内CO2的吸收量高于纯混合溶剂(13.75% wt) [51] 0.2% wt CA+30% wt MEA +7.5% wt PZ+15% wt K2CO3 90 ℃ 含有CA/Zn-Im: HFS的混合溶剂比纯混合溶剂的CO2相对解吸率高1.57倍. [51] | Show TableDownLoad:
CSV
3.3 多酶复合级联催化CO2
CO2加氢提供了一种碳再生方法,可以通过特定的催化剂转化为甲酸盐、甲醛和甲醇等高价值化学品和燃料. 固定化酶催化因具有反应条件温和、选择性高、可多次催化和效率高等优点,在CO2加氢中引起了广泛关注. 但是,常压下CO2的低溶解度严重限制了酶促反应的速率[53]. CA不仅能够加速CO2的水合,还用于促进酶催化的CO2转化. Wang等[54]报道随着CA的加入,甲酸脱氢酶的底物从CO2转化为更易溶解的
HCO−3 甲酸本身是一种具有商业价值的化学品,它可用作燃料电池的原料和青贮饲料防腐剂,并且二氧化碳形成甲酸所需的能量较低[55]. Zhai等[56]把CA固定在聚乙烯亚胺(PEI)和聚多巴胺(PDA)的改性的二氧化硅微球(PDA/PEI-SiO2-CA)上,并将其添加到含有NADH和FDH的反应溶液中,从而将CO2转化为甲酸盐. 通过改性在SiO2表面引入氨基,氨基与CO2反应形成氨基甲酸酯和碳酸氢盐,因此CA固定在PDA/PEI-SiO2表面上可产生协同效应. 在PDA/PEI-SiO2-CA存在下,初始反应速率是空白对照的48.6倍,甲酸盐产量也高于空白对照. Zhang等[57]通过微生物转谷氨酰胺转胺酶(MTG)充当“交联介质”将CA和FateDH交联在一起形成交联酶. 在二氧化碳转化为甲酸的实验中,交联酶的催化效率可高达游离酶的5.8倍.
固定多酶体系转化CO2生产甲酸已得到广泛研究. 然而,价格昂贵的辅因子(NADH)的再生和循环利用仍然是CO2高效转化的主要问题[58]. Ren等[53]通过将CA、FateDH、GDH(谷氨酸脱氢酶)、PEI(阳离子聚电解质)和辅因子原位包埋到ZIF-8中,构建了纳米级多酶反应器(Co-IMR). NADH通过带正电的PEI和带负电的辅因子之间的离子交换被束缚在ZIF-8中,并通过嵌入ZIF-8中的GDH再生. Co-IMR在8次重复循环使用后仍保留其初次甲酸生产率的50%,表现出良好的重复使用性. 与游离多酶系统相比,Co-IMR在8次重复循环使用后,总累积甲酸产量增加了4.6倍. Chai等[55]进行了类似研究,制备了ZIF-8/CA&FDH薄膜. ZIF-8中的咪唑基团和CA的协同作用使得CO2对甲酸的整体催化活性提高了1.6倍. Wang等[59]将CA、FateDH、GDH包埋在聚多巴胺微胶囊上. 与游离多酶相比,聚多巴胺微胶囊中固定化多酶体系催化产生的HCOOH高4.5倍以上,基于NADH的甲酸产率为342%. Li等[60]使用胺官能化MIL-101(Cr)作为核心,其具有CO2气体吸附性能,再通过包埋将CA、FateDH和GDH 3种酶固定在外层HKUST-1上. 胺官能化的MIL-101(Cr)释放的CO2底物首先进入CA并水合为碳酸氢根离子,然后
HCO−3 甲醇是一种重要的化工原料及能源物质,由合成天然气制备的甲醇程序复杂,且成本较高[8]. 多酶级联催化CO2甲醇化作为一种生物催化手段近年来引起了较多关注. Wang等[61]把FateDH、FaldDH和ADH共固定在聚苯乙烯微粒上,从而将CO2转化为甲醇. 在没有添加CA的情况下,固定多酶体系11次重复循环使用后产生了48%的累计甲醇收率. 甲醇的收率较低,有待进一步改进. Ji等[62]通过将甲酸脱氢酶、甲醛脱氢酶、乙醇脱氢酶和谷氨酸脱氢酶四种酶和辅因子原位包埋在阳离子聚电解质掺杂的中空纳米纤维腔内,并通过在中空纳米纤维的外表面上组装CA加速CO2水合,进行了进一步研究. 对于涉及3种脱氢酶的CO2还原酶系统,实现了103.2%的最高甲醇收率,该反应系统10次重复循环使用后产生了940.5%的累积甲醇收率.
3.4 CA转化CO2的工业应用
基于CA的生物催化系统可以有效地促进CO2的捕集和分离,但只有少数案例与工业过程中的实际应用密切相关. 对于工业应用,CA必须长时间在高温、高离子强度、高pH值和高剪切应力的条件下保持较高活性和稳定性. Lalande等[63]设计并安装了含有固定化CA的CO2填充柱,用于回收和循环利用来自水泥熟料生产的CO2. 固定化CA催化CO2水合形成碳酸氢根离子与添加的CaCl2反应生成的CaCO3被用作波特兰水泥厂的原料. 2015年,在加拿大魁北克省[64]进行了基于CA的CO2捕集工艺的有史以来规模最大的测试. 该测试使用了添加定向进化CA的20% wt的K2CO3/KHCO3溶液,能够在2500 h内以80%的平均捕集效率每天从燃烧天然气的锅炉的燃烧气体中捕集1×104 kg CO2. 实验过程中仅产生少量无毒碱性废水,无需特殊处理直接排入市政下水道. 此外,从汽提塔装置中排出的CO2气体的纯度约99.3%,可用于生产燃料、塑料和化学品等产品. Reardon等[65]在威尔逊维尔的国家碳捕集中心(NCCC)将CA固定在吸收柱的内表面,以20% wt K2CO3和非挥发性碱性盐溶液(AKM-24)作为吸收剂. 固定化CA使传质速率提高了6—7倍,并在40 d 3460 h内保持了超过80%的烟气中CO2的捕集效率. 为了评估CA催化的CO2加速吸收到MDEA水溶液中,在威尔逊维尔的NCCC进行中试规模的CO2捕集实验. 捕集装置在连续6 d吸收温度25—35 ℃和87 ℃解吸温度之间循环,在稳态条件下每天能够捕集多达68.1 kg的二氧化碳[3]. 总而言之,与之前的无生物催化剂过程相比,CA介导的CO2捕集过程具有显著促进作用.
4. 结论与展望(Conclusions and prospects)
为了改善游离CA在高温、强酸或强碱溶剂和化学杂质等条件下容易失活的特性,使用合适的载体和固定化方法对CA进行固定. CO2可通过酶促反应吸收,或通过CA一步催化或级联催化转化为无机碳酸盐、甲酸和甲醇等高价值产品. 固定化CA转化CO2的应用取得了一定的成效,但是仍存在一些技术瓶颈,可总结为以下几个方面:(1)CA价格昂贵. (2)CA对烟气中存在的高温、SOx和NOx耐受性差. (3)固定化方法尚不完善和载体的酶负载量低导致固定化CA活性低.
针对上述问题,未来的研究方向应集中于:(1)从转基因生物中提取大量CA,降低CA生产成本. (2)进一步研究CAs相关基因的生理作用和代谢途径,以提高化学和热稳定性. (3)研发先进的固定化方法和制备具有良好的机械强度、稳定性和对酶亲和力高的载体,提高固定化CA活性.
-
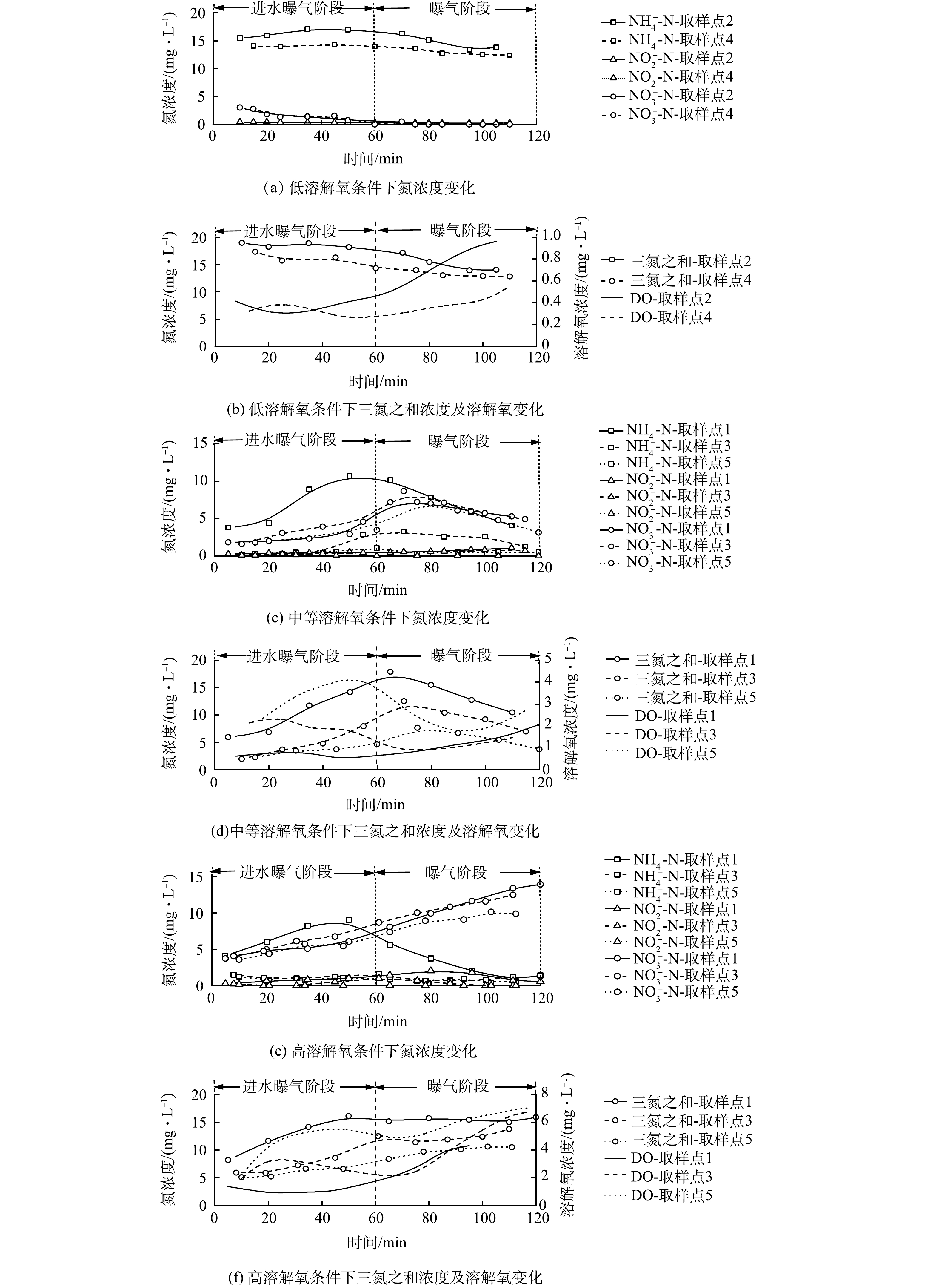
图 3 不同DO浓度下各取样点基质浓度变化
Figure 3. Variation of nitrogen concentration at each sampling point at different DO concentrations
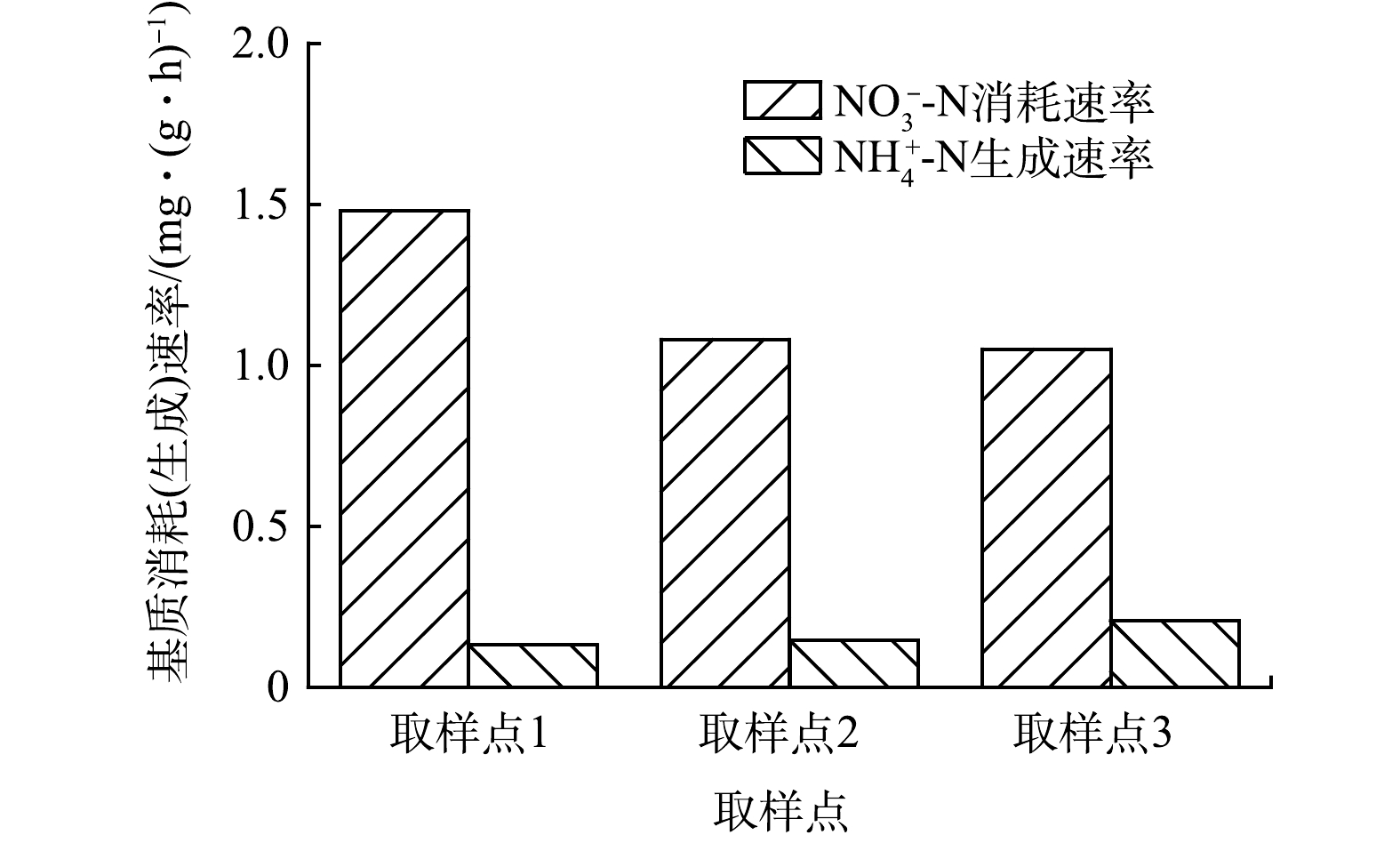
图 5 不同取样点内源反硝化过程中氮消耗(生成)速率
Figure 5. Rate of nitrogen consumption (production) during endogenous denitrification at different sampling points
表 1 进出水水质
Table 1. Quality of influent and effluent
mg·L−1 水样 COD NH4+-N NO3−- N TN PO43--P 进水 188±22 32.7±7.60 — 40.50±9.50 2.98±1.15 出水 20±3 1.60±1.30 6.74±3.59 11.30±2.20 0.07±0.06
下载: 导出CSV
表 2 CAST工艺典型周期各途径脱氮贡献占比
Table 2. Contribution of each pathway to nitrogen removal
脱氮方式 脱氮量/g 占比/% 反硝化脱氮 526.01±19.91 1.64±0.05 内源反硝化脱氮 20 093.13±1 291.55 62.86±4.13 同步硝化-反硝化脱氮 11 366.27±1 553.06 35.50±4.15 总脱氮 31 985.40±1 212.71 100
下载: 导出CSV
表 3 不同饥饿时长下活性污泥对HAc的吸收量
Table 3. Uptake of HAc by activated sludge at different famine durations
饥饿时长/h ΔMHAc0 ΔMNO−3 ΔMNO−2 ΔMHAc1 ΔMHAc2 5 335.16 79.35 74.87 91.31 243.84 24 394.68 101.39 88.34 128.47 266.2 36 468.91 103.59 94.3 124.77 344.14 48 407.25 102.71 93.03 124.46 282,79 72 376.14 96.54 84.47 121.76 254.38
下载: 导出CSV
表 4 CAST、A2/O和氧化沟工艺脱氮指标
Table 4. Denitrification indicators of CAST, A2/O and oxidation ditch processes
mg·L−1 工艺 进水COD 进水NH4+-N 进水TN 出水COD 出水NH4+-N 出水TN CAST 188±23 32.70±7.60 40.50±9.50 20±3 1.60±1.30 11.40±2.20 A2/O 249±38 36.12±5.38 44.36±4.87 18±3 0.32±0.26 8.30±1.26 氧化沟 643±136 35.12±4.89 47.50±5.33 21±3 1.07±0.57 8.47±1.01
下载: 导出CSV
-
[1] 邓伟斌. 南方某污水处理厂CAST工艺提标改造[J]. 中国给水排水, 2016, 32(16): 3-4. [2] 王俊, 王琴, 陈杰云, 等. CAST工艺处理小城镇污水的运行模式优化研究[J]. 中国给水排水, 2011, 27(19): 17-20. [3] MERVYN C, 朱明权. 循环式活性污泥法(CAST)的应用及其发展[J]. 中国给水排水, 1996, 12(6): 4-10. doi: 10.3321/j.issn:1000-4602.1996.06.001 [4] 孙剑辉, 闫怡新, 徐文刚. 循环式活性污泥法(CAST)的设计要点[J]. 中国给水排水, 2003, 19(1): 89-91. doi: 10.3321/j.issn:1000-4602.2003.01.028 [5] 孟栋. 利用剩余活性污泥合成聚羟基脂肪酸酯的研究进展[J]. 生物工程学报, 2019, 35(11): 2165-76. [6] BIAN D J, GUO H Y, ZHANG W H, et al. Simultaneous C and N removal in a hybrid CAST reactor[C]. 3rd International Conference on Bioinformatics and Biomedical Engineering, Beijing, 2009: 5145-5146. [7] WANG Y Y, ZHANG Z X, YAN M, et al. Impact of operating conditions on nitrogen removal using cyclic activated sludge technology (CAST)[J]. Journal of Science and Health Part A:Toxic/Hazardous Substances & Environmental Engineering, 2010, 45(3): 370-376. [8] WANG S P, YU J J, LIU Y H, et al, Achieving and maintaining of short-cut nitrification in a cyclic activated sludge system[J]. Water Science and Technology, 2011, 64(10): 2016-2022. [9] 国家环境保护局. 水和废水监测分析方法(第四版)[M]. 北京: 中国环境科学出版社, 2002. [10] 钱亮, 贺北平, 刘瑞东, 等. 西安市第四污水处理厂一期工程升级改造经验总结[J]. 中国给水排水, 2016, 32(2): 74-78. [11] 田敏, 崔涛, 吕恺, 等. 西安市第四污水处理厂A2/O工艺的脱氮性能评价[J]. 中国给水排水, 2020, 36(13): 1-6. [12] 徐凤, 李佩武, 刘艳辉. 曝气量对CAST工艺脱氮性能的影响[J]. 天津城市建设学院学报, 2010, 16(4): 281-284. [13] 彭党聪, 王志盈, 袁林江. 活性污泥系统非平衡增长理论及其应用[J]. 中国给水排水, 2001, 17(2): 19-21. doi: 10.3321/j.issn:1000-4602.2001.02.006 [14] MAJONE M, DIRCKS K, BEIM J. Aerobic storage under dynamic conditions in activated sludge processes. The state of the art[J]. Water Science & Technology, 1999, 39(1): 61-73. [15] 拉马尔奥, 王凤石. 废水处理概论[M]. 北京: 中国建筑工业出版社, 1982. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 2349
- HTML全文浏览数: 2349
- PDF下载数: 151
- 施引文献: 0













