










-
塑料因其优异的物理化学性能、低成本、使用便利成为全球最常用的基础材料之一[1]。2021年联合国环境规划署发布数据显示,目前全球累计生产的塑料有8.3×1010 t。在过去40年中,产量增两倍多,但总体回收率小于10%。海洋中现存近亿吨的塑料垃圾,且每年仍有约9×107~1.4×108 t塑料持续排放,给全球海洋生态系统造成的经济损失高达80亿美元,仅水产品损失就达31亿美元(UNEP,2021)。塑料污染是一个严重的问题,特别是尺寸小于5 mm的塑料[2]。微塑料(microplastics, MPs)是由大塑料碎裂化形成,或者原始生产的塑料微珠[3-5]。微塑料的大小、形状和颜色与天然沉积物和饲料的相似性可能会误导生物摄入。这会引发营养不良,甚至导致饥饿效应,表现为受影响生物的生长速度改变、健康状况降低和行为改变[5-9],进而可能影响渔业养殖产量和利润。形状不规则且边缘锋利的微塑料可磨损并破坏生物体中胃肠道黏膜的完整性[10]。此外,由于消化道酶促作用等,摄入的微塑料可能会释放单体和塑料添加剂,造成更多潜在危害[11-12]。另一方面,微塑料可作为某些环境污染物和致病菌的载体,摄入微塑料可促进它们转移到生物体内,并导致不良健康影响甚至死亡[11,13-15]。
红树林区海水养殖活动频繁,是粮食供应重要来源。微塑料赋存特征与海产品质量相关,是评估食品安全需要考虑的关键问题。虾等甲壳类动物会摄入其生长环境中的桡足类、鱼类幼虫,以及水生环境中的微塑料[16-18]。当虾被鱼类等更高营养级捕食者或人类食用时,可通过食物链转移和富集。虽然人类不食用虾的头部,但肌肉与消化组织微塑料赋存威胁水产品质量安全和人体健康。况且许多人在不去除肠道(虾线)的情况下将它们整个食用。人类食用被微塑料污染的虾形成直接的接触(传播)途径,对粮食安全和公共健康构成威胁。这些矛头指向如何研究微塑料在虾类海产品中赋存特征,以及在营养转移和积累中的潜在作用[18-19]。现有研究主要关注贝类和鱼类,对于虾类的报道比较有限。之前在几项研究中报道世界不同地区虾体内普遍检出微塑料[20-22]。这些研究中,选择虾是因为它们在食物链中发挥着重要作用,为从鱼类到人类的各种营养级生物提供食物。南美白对虾(Penaeus vannamei)是一种高度消费的海产品。2020年,世界对虾养殖年产量约为6×106 t,南美白对虾是其中养殖量最大的品种,约为5×106 t [23]。2020年,中国南美白对虾养殖产量同比增加4.41%,已占全球南美白对虾养殖产量的80%以上(《2020年全国渔业经济统计公报》)。10年来,南美白对虾的产量涨幅79.96%,2020年,福建省南美白对虾养殖产量为1.16×106 t(《2021年中国渔业年鉴》)。漳州是福建重要的南美白对虾养殖基地,养殖过程不可避免使用塑料养殖设施、渔具等。研究漳州红树林区海水养殖中南美白对虾微塑料的赋存特征,来源和影响因素对进一步评估水产品质量安全与养殖经济效益具有重要意义。
-
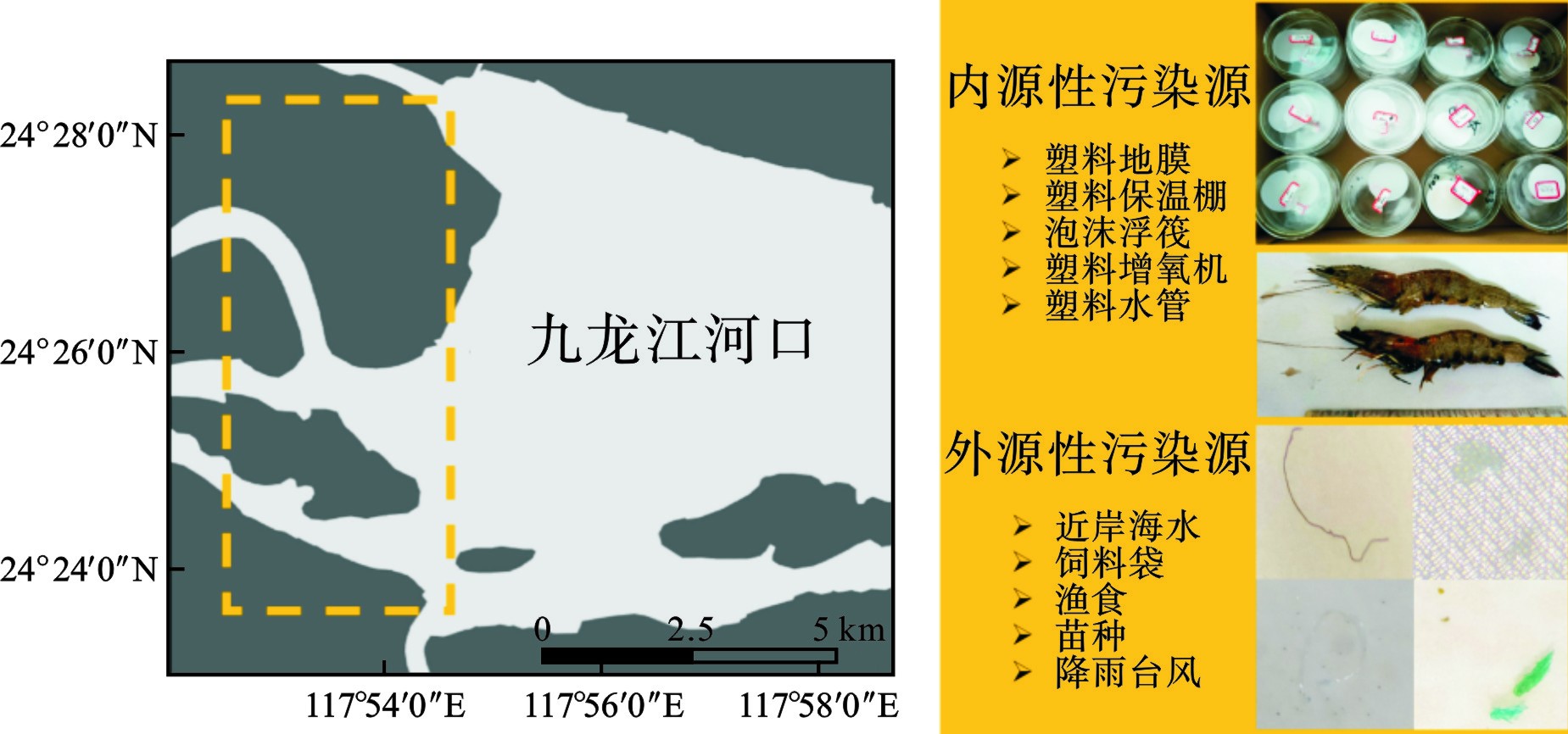
南美白对虾是对虾科、滨对虾属水生动物。体长最长可达24 cm,虾壳较薄,正常体色为青蓝色或浅青灰色,外观无斑纹,步足常呈白垩状[24]。本文选择南美白对虾为研究对象,因其是世界三大产量最高的虾类之一,是典型商业水产品。漳州是福建南美白对虾主要养殖基地,具有一定代表性,见图1。
-
光学显微镜(Moitc,SMZ168,中国)、烘箱(上海精宏,DHG-9140A,中国)、徕卡体式显微镜(Leica M165,德国)、傅里叶变换显微红外光谱仪(Thermo Fisher Scientific Nicolet iN10,美国)、电子分析天平(R&D,HR-250A,日本)、干燥箱(上海齐欣,DHG-9240A,中国)、超声清洗机(昆山舒美,KQ5200DE,中国)、游标卡尺,恒温振荡器(上海精宏,THZ-320,中国)、锥形瓶、搅拌器、烧杯、培养皿、解剖工具盒、pH试纸、锡箔纸、滤膜(Whatman,孔径0.70、1和50 μm,直径47 mm,英国)、10% 氢氧化钾溶液、超纯水、氯化钠、1.5 mol柠檬酸等。
-
2022年8月,从漳州市红树林区南美白对虾海水养殖基地3个不同养殖池(图1)采集南美白对虾样品共计900 g(每池各300 g),未洗涤状态直接装入铝膜袋储存在冷藏箱后第一时间带回厦门大学嘉庚学院生化大楼实验室。分别从3个不同养殖池随机选择3只虾,标记为养殖池1号的样品虾1~3,养殖池2号的样品虾4~6,养殖池3号的样品虾7~9。
-
为收集体液,每只虾先用超纯水清洗(先前通过0.70 μm孔径滤膜过滤),并根据SEVERINI[19]等描述的方法确定生物特征数据(长、宽、高、体重)。测量可量性状如下:
体长:由虾眼的前端到虾尾带有肌肉末端的直线长度
头长:由虾眼的前端到虾头壳末端的直线长度
体宽:虾身的最大宽度
体高:虾身的最大高度
体重:虾的质量(湿重)
然后,根据虾壳(虾壳与体液附着物)、肌肉,内脏组织(虾线与虾头)3大类来解剖。解剖过程避免破损,分离后分别装入锥形瓶中。在电子分析天平上分别测量重量并记录,−5 °C冷藏直至分析。
-
将25 mL的10% KOH溶液分别倒入锥形瓶中,立即用锡箔纸将瓶口覆盖, 防止空气中微塑料干扰。密封后的锥形瓶置于恒温振荡器中,设置振荡频率80 r/min,温度设置为65 ℃,振荡时间48 h,直到组织完全消化。在消化过程中消解液蒸发时加入额外的KOH溶液。样品冷却后,用1.5 mol柠檬酸中和到中性。每个样品通过玻璃纤维滤膜(0.70 μm孔径)真空过滤,然后放置在清洁过的培养皿中。消解过程注意观察瓶壁上是否黏附生物组织,将锥形瓶进行摇晃确保组织进入溶液中。使用饱和NaCl溶液(1.2 g/mL)作为浮选液分离微塑料,饱和NaCl溶液允许密度较低的颗粒分离,而未溶解的无机材料沉入底部。使用抽滤方式过滤上清液。玻璃纤维膜孔径1和50 μm,直径47 mm。最后,将每张滤膜置于干净的培养皿中,用锡箔纸覆盖,并在干燥柜中于50 °C干燥24 h。
-
(1)先使用普通光学显微镜分析每张滤膜是否存在微塑料,然后使用带数码显示的体式显微镜根据其颜色和形状识别拍照[25]。微塑料形状被分类为纤维、颗粒、泡沫、薄膜或碎片等[26],大小分为1~50和50 μm以上。根据微塑料的大小、颜色和形状进行识别、记录、拍照和分类。
(2)使用傅里叶变换红外光谱(FTIR)鉴定微塑料聚合物成分。采用中红外波段扫描多次,随后将所有光谱与聚合物参考光谱库进行比较,以验证聚合物类型。仅接受相似度为>70%的聚合物匹配[21],以确保结果的可靠性。通过FTIR分析验证微塑料聚合物类型后,重新计算丰度,排除具有相似形貌但未验证为微塑料观测物。
-
(1)分析和测量每只虾的微塑料尺寸、形状和颜色,并进行计数拍照。使用SPSS软件版本20.0(IBM, 美国)和Microsoft Excel 2022(微软,美国)对样本微塑料丰度进行统计分析。此外,还进行了One-way ANOVA方差分析,将生物学指标作为因变量,微塑料丰度作为自变量以进行微塑料丰度与生物指标Spearman相关性分析。
(2)在质量控制方面,实验前使用70%乙醇和纯净水彻底清洁所有实验桌台面和玻璃器皿。此外,为防止标本之间的微塑料交叉污染,每个标本上取出后须仔细冲洗镊子。为防止空气中的微塑料干扰,含有生物组织的锥形瓶或者放置滤膜的培养皿立即用锡箔纸覆盖。实验室期间穿着棉质实验服、口罩和丁腈手套。此外,用于制备这些溶液的反应溶液和蒸馏水在使用前通过0.7 μm孔径的滤膜过滤,以确保从试剂中消除微塑料。在样品处理分析过程中,使用对照空白评估实验室环境微塑料污染水平。
-
漳州南美白对虾样本生物学指标,见表1。
通过One-way ANOVA单因素分析,发现漳州南美白对虾样本生物学指标呈现显著差异(P<0.01),F值946.2。虽然同一时间投放的虾苗,且苗种来自同一个厂家。但是,养殖过程摄取的饲料量、饲料转化率不一。养殖环境微塑料赋存可影响其生长率。
Spearman相关性分析发现,漳州南美白对虾样本在体重和体长方面具有良好的相关性,Spearman相关系数为0.995 8。所有样品体长在13.7~14.8 cm之间变化,而体重在19.9 ~23.4 g之间,见图2。肌肉组织的重量在9.9~11.6 g之间。根据趋势线描述生长方程计算,其R² = 0.956 9,这意味着虾在生长时保持形状,即使在微塑料赋存的环境中。虾体长与虾重量呈一定正相关,其关系为y = − 2.4765x2 + 74.078x − 530.5。这是首次对南美白对虾生长体征相关因子与微塑料丰度相关性分析的研究。
图2显示内脏质量与微塑料丰度相关系数0.068 9、肌肉质量与微塑料丰度相关系数0.033 9、虾壳质量与微塑料丰度的相关性相关系数0.350 5,并没有发现其显著性关系。进一步分析整体质量以及虾体长与微塑料丰度关系,也没有发现其相关性特征。这可能因为样本数量不够,也可能因为养殖环境微塑料来源复杂,南美白对虾摄食行为与其体内微塑料赋存特征并不能直接从生物学特征上反映。整体上看,研究表明虾体长和重量呈现正相关关系,并未发现微塑料丰度与生物学特征的相关性特征。
-
从尺寸特征上看,见表2。
主要以小尺寸微塑料为主(1~50 μm),占比74.7%,这与其他学者采集的野生深水虾的尺寸特征不同。ESTER et al[27]观察到,从地中海西北部收集的深水虾(Aristeus antennatus)的胃中有13种微塑料,尺寸最长的高达6.6 mm[27]。本研究虾壳中检出30个小尺寸微塑料,占比最多(85.7%),50 μm以上微塑料检出5个。虾壳中检出的微塑料主要来自养殖水环境中赋存的微塑料或者饲料等。小尺寸的微塑料可能嵌入或吸附于虾壳,特别是虾脚和虾头容易携带微塑料。但是,虾类等甲壳类生物在生长发育过程需要蜕壳,这会去除大量的微塑料附着。蜕壳后的虾壳会漂浮在养殖水池表面,养殖户一般会打捞清除。
内脏中检出的微塑料最多,合计39个。各个样品中,最高检出9个,最低2个。其中,小颗粒微塑料(1~50 μm)是50 μm以上的颗粒数量的2倍多。文献[28]报道大西洋长臂虾(Palaemon varians)很容易摄入尺寸为0.1~9.9 µm的聚苯乙烯微珠,摄入的微塑料在消化器官通过复杂的过滤系统按大小分离,一些最小的颗粒(0.1 µm)可进入肠道。一般来说,虾线即消化系统是食用时会去除的部位,虾头内也有丰富的器官。由于本研究没有再细分器官,无法进一步区分具体什么器官的含量更高。但是,本研究所定义的虾头包括各种内脏器官(心脏、胃、肝等),小颗粒微塑料更容易被摄入并进入代谢系统。LUSHER et al[29]研究发现尺寸小于150 μm的微塑料在通过食物摄入后,主要通过易位进入循环组织和内脏,包括肾脏、大脑、肝脏和性腺。
最后,从肌肉中检出的微塑料尺寸特征上看,小颗粒1~50 和50 μm以上大颗粒的检出数量差异不大,合计分别是10和7个。因为微米尺寸级别上,微塑料已经可以通过新陈代谢进入各个器官组织,进而进入肌肉[29]。值得注意的是,肌肉是人类食用虾类主要部位,其微塑料检出预示着潜在水产品质量安全风险。
从微塑料丰度整体分布特征上看,见图3。
1号和3号养殖池的微塑料丰度较低,最高检出7个,最低5个。2号养殖池丰度最高(图3红色框标记),最高检出19个,最低16个,占比达到所有养殖池的检出量的58.2%。2号养殖池是较早投建的一批养殖池,靠近内陆(距离村居最近),其抽水管线路最长,且养殖池面最宽,养殖密度相对较大。由于投建时间较长,养殖池内铺设的塑料地膜和保温棚年久老化,可能是内环境微塑料来源。从器官分布特征上看,除虾2样品的肌肉和虾壳中未检出微塑料外,其余样品均在各个器官组织检出微塑料。虾6样品检出的微塑料最高,达到19个,其中内脏检出9个,虾壳检出7个,肌肉检出3个。虾壳检出微塑料说明养殖环境中普遍赋存微塑料。内脏组织的微塑料检出说明南美白对虾可通过摄食等生物行为摄入微塑料。肌肉组织微塑料检出说明摄食的微塑料并无法完全排泄出,可进一步代谢迁移。整体上看,内脏和虾壳中的微塑料检出量占比最高,肌肉相对较少(平均1.89 n/a),这与其他研究报道结论一致[18,20-21,30-31]。按照食用南美白对虾的习惯,去除掉虾头、虾壳、虾线等组织器官,单独食用肌肉是相对安全的。
-
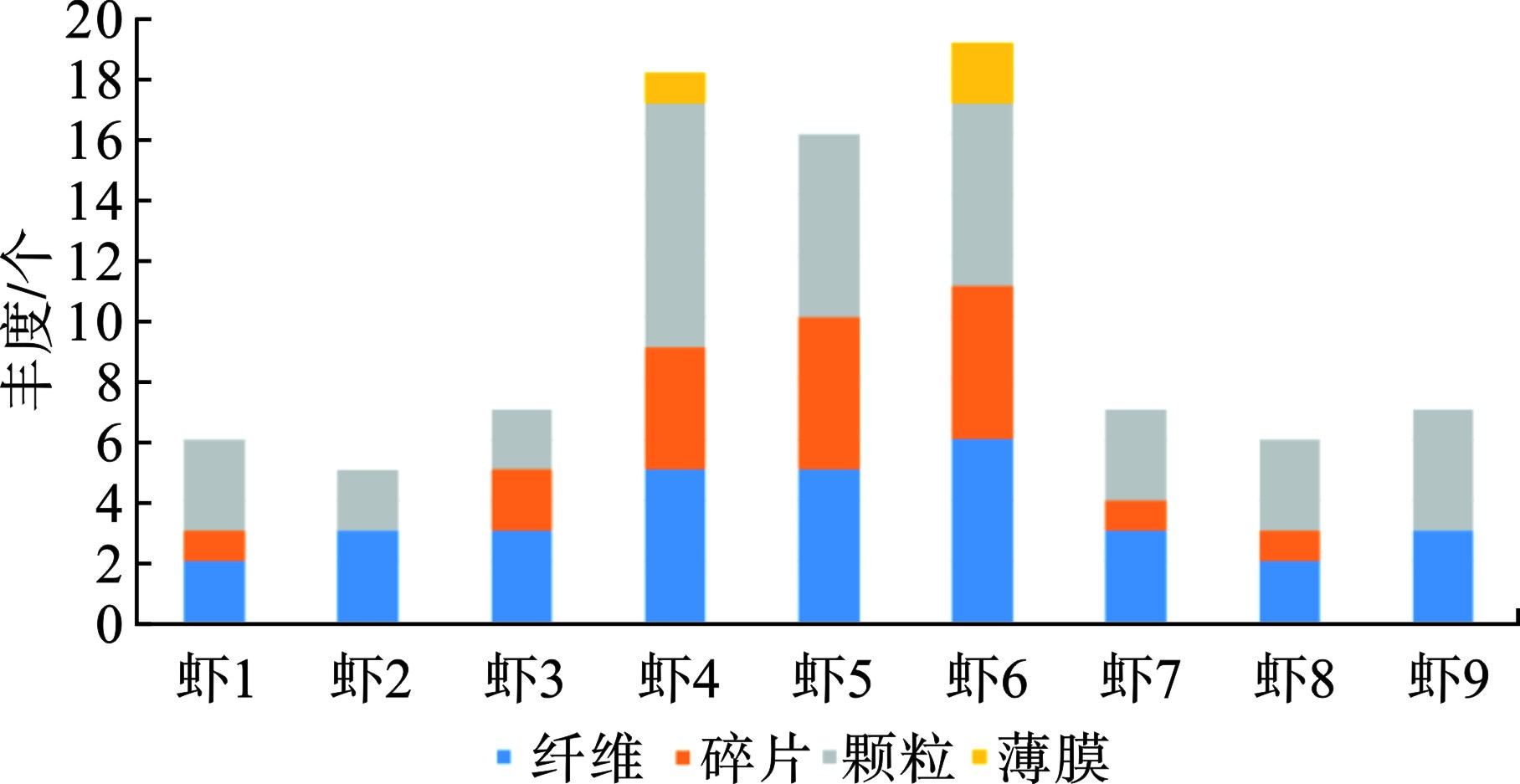
从形状特征上看,见图4。
图4可知,共检出4种形状的微塑料,分别是纤维32个、碎片19个、颗粒37个和薄膜3个。薄膜状只在2号养殖池采集的样本中检出,分别是虾4检出1个,虾6检出2个。薄膜状微塑料检出率较低,整体占比只有3.3%。只有2号养殖池的虾4和虾6样本检出4种形状,其他样本只检出2种或者3种形状。2号养殖池也是微塑料平均丰度最高的样本来源,这说明微塑料形状的多样性与微塑料丰度有一定的相关性,微塑料丰度越高可能检出更多种类的形状。不管是几号样本,均检出纤维状和颗粒状微塑料。纤维状和颗粒状合计检出69个,占总微塑料检出数量的75.8%。其高占比可归因于南美白对虾摄食习惯以及研究区域的人为活动。据报道,纤维状微塑料是全球海水中最丰富的形态[32]。本研究观察结果与之前报道的甲壳类动物(Crusta)[7]、蓝贻贝(Mytilus edulis)[33]和褐虾(Crangonidae)[33]的研究一致。这些研究显示这些物种的肠道中纤维状微塑料占主导地位[33-34]。HOSSAIN et al[21]确定纤维状微塑料在斑节对虾(Penaeus monodon)中占主导地位(57%),其次是颗粒(29%)和碎片(14%);而褐虾则以颗粒状(42%)为主,其次是纤维(32%)和碎片(26%)。渔业养殖过程中需要使用大量塑料渔绳和渔网,这可能是纤维状的主要来源。颗粒形状的微塑料在海洋环境中广泛赋存,样本中检出的颗粒状微塑料可能来自换水带来的周边近海表层水体,也可能是饲料或者塑料包装袋等来源。纤维和颗粒状微塑料更可能被南美白对虾误食。建议未来进一步观测其摄食行为和器官组织形态,以评估是否具有显著相关性。整体上看,漳州南美白对虾检出的4种形状微塑料具有一定代表性。薄膜是检出量最少的微塑料,其次是碎片。环境中塑料废物是养殖微塑料主要来源[35-36]。各种塑料材料和养殖设备,如塑料地膜、保温棚、泡沫浮阀、绳索、渔网、增氧机等是环境中微塑料的潜在来源(图1)。据调查,漳州红树林区海水养殖池塘是封闭或半封闭的,这导致微塑料的持续的保留和积累。此外,养殖池底部沉积物可作为微塑料的沉积区[37]。
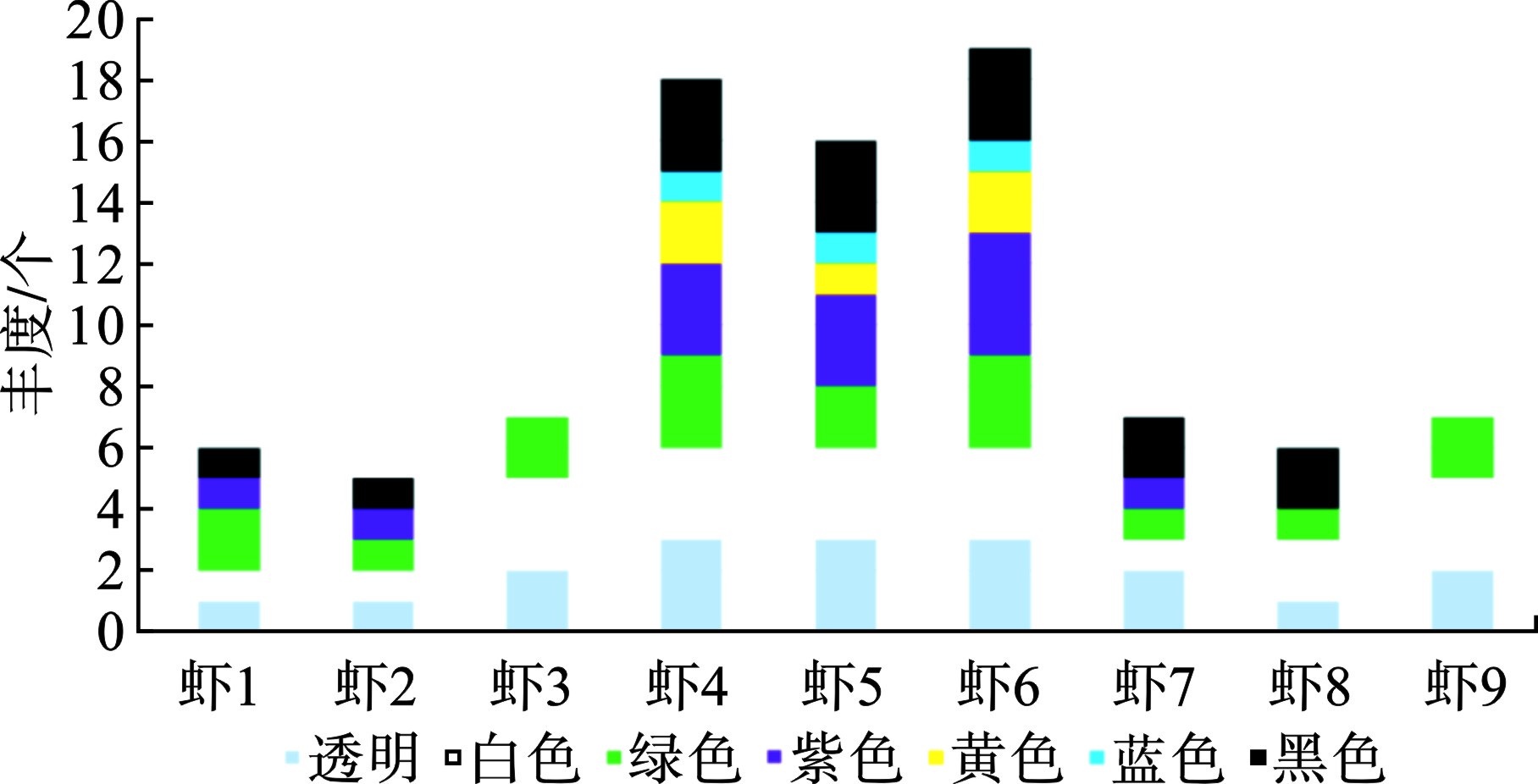
从颜色特征上看,见图5。
所有样本共检出7种颜色。占比最高的是白色和透明色,分别达22%和20%,其次是绿色(17个)和黑色(15个),最少的是蓝色(3个)。在从孟加拉国水域收集的老虎虾和棕色虾中也观察到类似的微塑料颜色特征[21]。微塑料可以直接被虾类摄食,因它们与饲料或天然沉积物相似;或者因为微塑料对食物(饲料)颗粒的粘附被间接食用。鉴于微塑料在视觉上与虾的天然食物和猎物相似,这些生物对微塑料的生物利用度相对较高[38]。并不是所有养殖池都检出多样性颜色的微塑料,比如1号养殖池虾3号样本只检出3种颜色,3号养殖池虾9样本检出3种颜色,只有2号养殖池的所有样本检出7种颜色微塑料。但是,不管几号养殖池,白色、绿色和透明颜色在所有养殖池的虾样本中都有检出。在虾类样品中检出透明的颜色,预示着其微塑料相对较新,养殖过程中使用的塑料保温棚可能是其来源。绿色微塑料比较引发关注,通过对现场养殖设备和养殖工具调查,发现养殖过程中需要定期通过吊挂绿色渔网观察南美白对虾进食情况,这些网袋可能是绿色微塑料来源。另据报道,褐虾主要摄取黄绿色的微塑料(50%),其次是紫蓝色(43%),半透明(15%)等[34]。
黑色微塑料也是海洋环境中常见的颜色。养殖池底部大面积铺设的塑料地膜年久老化脱落,可能是其来源。整体上看,本研究样本中微塑料颜色特征,预示着漳州红树林区南美白对虾体内微塑料来源更多来自内源性污染,受养殖设施和工具中使用的塑料制品影响。值得注意的是,微塑料颜色的可变性。由于塑料产品生产时使用着色剂,例如包装、渔具等[39]。着色剂可能老化脱落,透明颜色的微塑料在环境中可能因为氧化而变得不透明,显出白色。最后,紫色的塑料在养殖池中并无检出,但是样本中部分检出可能预示着养殖换水过程带来的外部污染。
-
本研究证实漳州红树林区海水养殖南美白对虾体内普遍检出各种形状、尺寸和颜色的微塑料。研究表明虾体长和重量的正相关关系,并未发现微塑料丰度与生物学特征的相关性。从尺寸特征上看,主要以小尺寸微塑料为主(1~50 μm),占比74.7%。内脏中检出的微塑料最多,39个;肌肉中检出的小颗粒(1~50 μm)和50 μm以上大颗粒数量差异不大。内脏和虾壳中的微塑料检出量占比最高,肌肉相对较少(平均1.89 n/a)。按照食用南美白对虾的习惯,去除掉虾头、虾壳、虾线等组织器官,单独食用肌肉是相对安全的。从形状特征上看,共检出4种形状的微塑料,纤维状和颗粒状微塑料占比75.8%。从颜色特征上看,占比最高的是白色和透明色,分别达22%和20%。漳州红树林区南美白对虾体内微塑料更多来自内源性污染,受养殖设施和工具中使用的塑料制品影响。封闭或半封闭的养殖池塘促进微塑料持续的保留和积累。微塑料的检出预示着潜在海产品质量安全风险,未来需要进一步研究其毒理以评估人体健康危害。
红树林海水养殖南美白对虾微塑料赋存特征、来源和影响因素
Occurrence characteristics, sources and influencing factors of microplastics in Penaeus vannamei from mangrove wetland
-
摘要: 微塑料污染威胁海水养殖与人体健康。南美白对虾(Penaeus vannamei)是一种高度消费海产品,文章首次对福建漳州红树林区南美白对虾微塑料赋存特征、来源和影响因素进行研究。结果证实南美白对虾体内普遍检出各种形状、尺寸和颜色的微塑料,未发现微塑料丰度与生物学特征的相关性。从尺寸特征上看,主要以小尺寸微塑料为主(1~50 μm),占比74.7%。内脏和虾壳中的微塑料检出量占比最高,肌肉相对较少(平均1.89 n/a),单独食用肌肉相对安全。共检出4种形状微塑料,纤维状和颗粒状占比75.8%。颜色占比最高的是白色和透明色,分别达22%和20%。漳州南美白对虾体内微塑料更多来自内源性污染,封闭或半封闭的养殖池塘促进微塑料积累。微塑料普遍检出预示着潜在海产品安全风险,建议进一步研究毒理以评估其对人体健康的影响。Abstract: Microplastic pollution threatens mariculture and human health. Penaeus vannamei is a highly consumed seafood. The characteristics, sources, and influencing factors of microplastics in Penaeus vannamei in the mangrove forest area of Zhangzhou, Fujian were studied at the first time. The results showed that microplastics of various shapes, sizes, and colors were commonly detected. No correlation between microplastic abundance and biological characteristics was found. From the perspective of size characteristics, small-size microplastics (1~50 μm) mainly accounted for 74.7%. The proportion of microplastics detected in viscera and shrimp shells was the highest, the microplastics in the muscle was relatively small (average 1.89 n/a), indicating that the muscle was relatively safe to eat alone. A total of 4 kinds of microplastics were detected, and fibrous and granular microplastics accounted for 75.8%. The highest proportion was white and transparent, accounting for 22 % and 20 % respectively. The source of microplastics in Penaeus vannamei of Zhangzhou could be from endogenous pollution. Closed or semi-closed aquaculture ponds promoted the accumulation of microplastics. The detection of microplastics indicated a potential risk to the quality and safety of seafood, and further toxicological studies were recommended to assess their human health.
-
Key words:
- Penaeus vannamei /
- microplastics /
- characteristics /
- mariculture /
- seafood safety /
- influencing factors
-
碳酸酐酶(CA)是一种普遍存在的含锌金属酶,其主要功能是催化CO2的水合反应,反应式如下所示:CO2+H2O↔HCO3−+H+,它是目前已知的催化最快的酶之一[1]. 由于游离CA在高温、强酸或强碱溶剂和化学杂质等条件下容易失活,限制了其大规模的工业应用. 因此,采用分子修饰[2]、定向进化[3]、酶固定化[4]等策略来提高其稳定性和活性. 在上述方法中,酶固定化因其操作简单、方便的特点而被广泛应用.
通过选择良好的载体和合适的固定化方法对游离CA进行固定是降低催化剂成本、提高稳定性和可重复使用性的有效途径[5]. 基于固定化CA的生物催化剂因其低耗能和高效率被认为是转化CO2的环境友好的候选方法. 固定化CA主要通过酶促反应吸收[6],CO2矿化生成金属碳酸盐[7]和多酶级联反应[8]来进行CO2转化. 吸收是目前应用最为广泛的CO2捕集技术之一[6]. 然而,吸收溶剂具有再生所需能量高、吸收速率慢和稳定性低等缺点[9]. CA作为促进剂可以提高CO2吸收反应速率,从而降低反应器尺寸和成本. 在某些情况下,CA还用于促进酶催化的CO2转化. CA参与的多酶级联反应用于生产甲酸盐和甲醇等高价值产品[7].
无论CA是以何种方式转化CO2,CO2转化实验始终以烟气为基础,而烟气中具有高温和其他化合物(NOx和SOx等)[10]. 这些可能会影响CA的活性和稳定性,进而减缓CA转化CO2实际工业应用中的反应速率.
1. 碳酸酐酶的性质和催化机理(Properties and catalytic mechanism of carbonic anhydrase)
1.1 碳酸酐酶的性质
CA广泛存在于细菌、真菌和藻类等生物体内,分为不同的类别(α、β、γ、δ、ζ、η和θ)[11]. 大多数CA从牛血红细胞中提取,少数CA从羊和猪等哺乳动物的肝脏以及重组细菌中获得[12]. CA是一种以锌离子作为活性位点的金属酶,在某些情况下,也可以选择其他金属原子,如Cd2+、Co2+、Fe2+或Mn2+来代替Zn2+[13-14],催化各种水解反应,包括羧酸、卤化物、尿素和可水解底物的水合作用[15]. 虽然相同的催化反应由不同的CAs进行,但它们既没有结构相似性,也没有其他相关性[16].
1.2 碳酸酐酶的催化CO2水合机理
在自然状态下,CO2的可逆水合反应十分缓慢,一级反应速率常数仅为5×10−2 s−1,反应步骤如下所示[17]:
(1)CO2水合生成H2CO3:
CO2(aq)+H2O↔H2CO3 (1) (2)H2CO3电离生成
HCO−3 CO2−3 H2CO3↔H++HCO−3 (2) HCO−3↔H++CO2−3 (3) 反应式(1)是CO2水合反应的限速步骤,也是生物系统中碳矿化和碳再利用技术的切入点[18]. 随着CA的引入,Kcat值可高达1.6×106 s–1[19],说明CA可以高效催化CO2的可逆水合反应. 与此同时,CO2水合反应的机制发生改变,变为以下两个步骤[20]:
第一步是CA中与活性中心的Zn2+相连的H2O去质子化形成中间产物EZnOH−,随后CO2被EZnOH−中的OH−水合形成EZn
HCO−3 HCO−3 HCO−3 HCO−3 第二步是EZnH2O的去质子化:EZnH2O通过酶分子中的质子转运体将质子(H+)转运至溶剂中并被还原成具有催化活性的EZnOH−.
2. 碳酸酐酶的固定化(Immobilization of carbonic anhydrase)
酶固定化的催化活性和稳定性在很大程度上取决于载体和固定化方法的选择,首先应在固定化过程中保持酶活性[21]. 近年来,各种材料已被用作固定化CA的载体,包括水凝胶[22]、壳聚糖[23]、二氧化硅[19]和金属有机框架(MOFs)[24]等. 酶可以通过多种方法固定在载体上,例如包埋、吸附、交联、共价结合、交联酶聚集体(CLEAs)[25]和纳米花型酶-无机杂化固定化酶(简称纳米花)[26]等. CA固定化方法优缺点如表1所示.
表 1 CA固定化方法优缺点Table 1. Advantages and disadvantages of CA immobilization methods固定化方法Immobilization methods 优点Advantages 缺点Disadvantages 参考文献Reference 吸附 易于操作;固定化酶构象变化小 酶易从载体中泄漏 [6] 共价结合 酶和载体之间紧密结合;提高了固定化酶的稳定性 引起酶构象的变化;酶的活性通常在固定化后降低 [7] 包埋 操作简单;可以减少酶浸出和变性 酶被限制在载体材料中,减弱了酶的表观活力 [6] 交联酶聚集体 无需载体;对酶纯度要求不高 机械强度不高 [21] 纳米花 固定化酶具有高比表面积和高酶活性 纳米花晶体的生长时间较长 [26] | Show Table DownLoad:
CSV
DownLoad:
CSV
吸附是一种简单有效的固定化方法,通过氢键、疏水键、离子键和范德华力的相互作用将酶固定在载体表面. 在多数情况下,与载体的温和相互作用可确保固定后酶的微小构象变化[27],这在很大程度上取决于载体的性质. Yin等[24]结合了磁性材料易于回收和可重复使用性,以及MOF具有高孔隙率、大比表面积和良好稳定性的优点,构建了核壳磁性ZIF-8@Fe3O4复合材料. CA通过吸附固定在ZIF-8@Fe3O4上. 游离CA和CA@ZIF-8@Fe3O4分别在40 ℃的1 mol·L−1 MDEA中储存9 d,CA@ZIF-8@Fe3O4的活性随着储存时间的延长而逐渐降低,但其活性始终高于等量游离CA. 包埋是一种将酶限制在载体材料中的固定化方法,可以有效防止酶浸出和变性[6]. Hsieh等[19]将二氧化硅缩合肽(R5)与硫化氢碳酸酐酶(SazCA)融合形成R5-SazCA,再把R5-SazCA包埋到二氧化硅纳米颗粒(R5-SazCA-SP)中. 在70 ℃培养3 h后,R5-SazCA和R5-SazCA-SP的残余活性分别为49%和60%,表明CA固定化后热稳定性得到改善. 增强的热稳定性通常归因于二氧化硅交联网络施加的空间约束,这有助于维持CA的立体结构.
共价结合是一种常用的固定化方法,可以显著增强和提高酶的稳定性. 改性载体表面的胺基、羟基、羧基或环氧基等官能团和CA的氨基酸残基之间可以形成共价键,从而使得酶和载体中间具有很强的相互作用[28]. Shamna等[29]将CA通过共价结合固定在胺官能化的铝-硅氧烷气凝胶珠(BCA-Al/Si-NH2)上. BCA-Al/Si-NH2杂化珠的动力学参数与游离酶相当. 固定化CA具有热稳定性、pH稳定性和储存稳定性,这是由于酶的多点附着和酶与载体之间的相互作用导致酶刚性增加,从而减少了构象变化. Sun等[30]将CA通过戊二醛(GA)共价连接到聚偏二氟乙烯(PVDF)膜和聚乙烯膜(PE)的表面,这些膜固定化CA之前通过聚乙烯亚胺(PEI)和多巴胺(DA)的共沉淀进行了修饰. PEI/PDA-PE的活性回收率高于PEI/PDA-PVDF,因为PE具有更大的孔结构和改性后更好的亲水性,从而引起更高的酶负载量和更好的酶亲和力. 此外,固定化后,CA的储存稳定性和可重复使用性得到很大提高.
考虑到载体材料的物理和化学性质、操作条件等,通常需要多种固定化方法的协同作用从而获得高效的固定化CA[6]. Chang等[31]将CA通过吸附-交联法固定在地质聚合物微球上,与游离CA相比,固定化CA的最适pH值从7.5增加到8.0,最适温度从25 ℃增加到30 ℃. Effendi等[11]将Sulfurihydrogenibium yellowstonense CA(SyCA)共价结合在新型聚丙烯腈(PAN)和聚对苯二甲酸乙二醇酯(PET)纳米纤维上,所得复合材料通过戊二醛(GA)进一步交联. 固定化粗SyCA在60 ℃下保留了超过100%的相对残余活性,这归因于粗SyCA的胺基和纳米纤维的羟基之间的共价结合,而且固定化粗SyCA和GA之间交联也会形成共价键. 此外,测定了固定化CA对SOx和NOx的耐受性. 固定化SyCA在50 mmol·L−1 HNO3和H2SO4存在下的残余酶活性分别为57.1%和61.6%.
CLEAs是一种无需载体的新型固定化酶制备技术,该方法包括沉淀和交联两个主要步骤[26]:(1)通过沉淀剂(盐、水溶性有机溶剂或非离子聚合物)在溶液中形成酶聚集体;(2)聚集体中酶分子通过交联剂交联,使它们在去除沉淀剂后不溶. 然而,对于工业应用来说,CLEA柔软且容易断裂,为了提高CLEA的机械性能,Jun等[32]使用纳米纤维共价连接bCA,然后交联并进行沉淀,以获得bCA沉淀涂层(EPC). EPC在剧烈摇动下868 d后仍保留其活性的65.3%,EPC的高稳定性可以通过酶沉淀和交联的结合来解释,使酶分子之间通过化学键紧密的链接,从而有效防止酶变性和浸出. 纳米花型酶-无机杂化固定化酶由2012年Ge等[33]首次提出,通过将酶与硫酸铜在磷酸盐缓冲溶液中共沉淀,从而形成典型的花瓣状结构. Wen等[22]对此固定化方法进一步研究,将双金属离子(Cu2+和Zn2+)代替单个金属离子来固定CA,然后将双金属基杂化纳米花(CANF)复合材料嵌入到聚乙烯醇(PVA)/壳聚糖(CS)水凝胶膜(PVA/CS@CANF)中. 水凝胶膜在8个循环后仍保持其原始活性的75%以上,在60 ℃下仍有超过80%的相对活性. 此外,PVA/CS@CANF膜转化CO2产生的CaCO3量是游离CA的9倍.
3. CA转化CO2的应用(Application of CA to CO2 conversion)
二氧化碳排放是造成温室效应的主要原因. 当前,CO2捕集、利用和封存(CCUS)技术已广泛应用于减少CO2排放到大气中. CCUS技术将CO2资源化,便于运输和长期储存[29]. 基于CA的生物酶法通过捕集大气或废气中的CO2,并进一步转化为高价值的工业化学品和燃料,如金属碳酸盐、甲醇和甲酸等.
3.1 CA在CO2矿化中的应用
CA催化的CO2水合反应生成的碳酸氢盐可进一步用作碳源,用于合成无机碳酸盐和促进藻类的生长. CA将CO2矿化生成CaCO3的原理为CA首先催化CO2水合形成碳酸氢根离子(4),然后与添加的Ca2+反应生成CaCO3(5),CaCO3易于分离和储存以供进一步利用. Shamna等[29]将CA共价连接在铝-硅氧烷气凝胶珠上,并进一步检测了其将CO2转化为CaCO3的能力:1 mg游离bCA和17 μg bCA-Al/Si-NH2生成CaCO3质量为32.4 mg和12 mg. 通过XRD表明CA转化的碳酸钙的晶相为方解石,且CaCO3的晶体结构不会因添加酶而改变. Jun等[32]使用纳米纤维共价固定bCA,然后进行交联并沉淀,以获得bCA沉淀涂层(EPC). CA通过催化系统中鼓泡的CO2的可逆水合生成碳酸氢盐,为微藻提供光合作用的碳源. 添加 EPC的系统与含有25 mmol·L−1 NaHCO3和不含碳源的相比,分别使微藻生长加速了134%和231%. Xu等[34]将CA与戊二醛(GA)交联并包埋在漂浮的海藻酸钙水凝胶珠中,并将珠子保留在微藻-大气界面便于直接从空气中捕集CO2. 与(22.7±0.5) mg·L−1每天的自然生长速率相比,游离CA和CA-GA珠粒分别将微藻的生长速率提高到(37±3)mg·L−1每天和(40±1)mg·L−1每天,而且珠子容易回收和重复使用,具有广阔的工业前景.
CO2+H2O→HCO−3+H+ (4) Ca2++HCO−3→CaCO3+H+ (5) 当前通过全细胞固定化构建了用于CO2捕集的新型全细胞催化剂,避免酶泄漏和细胞裂解从而提高酶的稳定性和可重复使用性[7]. Sharma等[35]将含CA的苦参棒状杆菌菌株的细胞共价固定到戊二醛功能化的角蛋白颗粒上. 含有144 U·mg−1 CA的菌株细胞裂解物形成65.12 mg CaCO3. 进行10次CO2转化后,固定化细胞和游离细胞CaCO3的相对产量分别为53.46%和22.15%. Moon等[36]将周质水生弧菌CA(hmCA)的全细胞催化剂通过包埋固定在聚氨酯泡沫上(pCA-PUF),经9次循环使用后,pCA-PUF的高相对活性没有明显下降,并且显示出良好的可重复使用性. 使用该固定化生物催化剂将设计的填充床反应器中的CO2捕集速率提高了80%. 微生物细胞表面展示是将某一蛋白或短肽(靶蛋白)与微生物细胞的外膜蛋白(载体蛋白)以融合蛋白的形式锚定在微生物细胞表面的技术. 大肠杆菌是生产重组蛋白最常用的宿主细胞之一[37]. Zhu等[38]以大肠杆菌为宿主细胞,通过将SazCA作为靶蛋白与冰核蛋白(INPN)(载体蛋白)锚定在细胞外膜上构建表面展示菌株. 工程菌株矿化CO2产生的CaCO3质量(241 mg)明显高于胞内表达菌株(173 mg). Tan等[39]将(Mesorhizobium loti CA)MlCA克隆到3个载体中且在大肠杆菌中表达,并将离心获得的全细胞样品固定在琼脂上. 结果表明,全细胞生物催化剂具有良好的稳定性,储存40 d后相对活性保持近100%. 此外,具有固定化全细胞生物催化剂的系统可在3.5 min内有效地将CO2100%转化为CaCO3. 通过表2展示了固定化CA矿化CO2性能的比较.
表 2 CA在二氧化碳捕集中的矿化性能Table 2. Mineralization performance of CA in carbon dioxide capture碳酸酐酶种类Types of carbonic anhydrase 矿化特性Mineralization properties CaCO3晶相CaCO3 crystal phase 参考文献Reference 牛CA(bCA) 固定化细胞比游离细胞的CaCO3产量增加了1.35倍;固定化细胞和游离细胞的CaCO3 相对产量在10个循环后分别为53.46%和22.15% 球霰石和方解石 [35] bCA 17 μg固定化CA产生12 mg CaCO3 方解石 [29] CA CA@ZIF-8获得的CaCO3的产量是游离 CA的22倍 球霰石 [40] 硫化氢碳酸酐酶(SazCA) 固定化CA转化CO2形成碳酸钙的时间比游离CA缩短了33% 球霰石和方解石 [19] SazCA 工程菌株矿化CO2产生的CaCO3质量(241 mg)高于细胞内表达菌株(173 mg) — [38] 周质水生弧菌CA(hmCA) 固定化CA将填充床反应器中的CO2捕集速率提高了80% — [36] CA PVA/CS@CANF膜转化CO2产生的CaCO3 量是游离CA的9倍 — [22] bCA 添加EPC的系统与含有25 mmol·L−1 NaHCO3和不含碳源的对照相比,分别使微藻生长加速了134%和231% — [32] bCA 与(22.7±0.5)mg·L−1·d−1的自然生长速率相比,游离CA和CA-GA珠粒分别将微球藻的生长速率提高到(37±3)mg·L−1·d−1和(40±1)mg·L−1·d−1 — [34] Sulfurihydrogenibium yellowstonense CA(SyCA) 固定化粗CA的CaCO3总产量是游离CA的5.8倍 — [11] CA 固定化CA产生的CaCO3质量是空白实验的5倍 — [31] bCA CA@ZIF-8和PVA/CS/CA@ZIF-8水凝胶膜获得的CaCO3产量分别为游离CA的20倍和32.6倍. 球霰石和方解石 [41] Mesorhizobium loti CA(MICA) 具有固定化全细胞生物催化剂的系统可在3.5 min内有效地将CO2100%转化为CaCO3 — [39] | Show TableDownLoad:
CSV
3.2 液体吸收法捕集CO2
到目前为止,已经开发了使用纯水、碳酸盐溶液(Na2CO3和K2CO3等)和醇胺溶液等吸收二氧化碳的工艺. 常用于捕集CO2的醇胺溶液主要有伯胺(一乙醇胺,MEA)、仲胺(二乙醇胺-DEA)、叔胺(N-甲基二乙醇胺-MDEA、二异丙醇胺-ADIP和三乙醇胺-TEA)和哌嗪(PZ)等[42]. MEA吸收速度快,不易挥发,但是溶剂在汽提塔中的解吸需要大量的能量,导致电费成倍增加,需要对其进行改进. 与MEA相比,K2CO3溶液具有低毒性,不易腐蚀,在高温下高效且低成本的再生等优点,而且MDEA再生能耗低、吸收CO2容量大. 但是K2CO3溶液和MDEA吸收速率较慢,添加CA可显著促进MDEA和K2CO3溶液吸收CO2[17].
当前,固定化CA已用于纯水中吸收二氧化碳研究. Chang等[31]将CA通过吸附-交联法固定在地质聚合物微球上. 采用水浴设定不同温度,pH计可监测反应溶液pH值随不同催化反应条件的变化,从而评价固定化CAs的催化性能. 当温度为30 ℃,气体流速为300 mL·min−1时,溶液的pH值明显下降. Xu等[43]将CA通过交联固定在聚多巴胺(PDA)/聚乙烯亚胺(PEI)修饰的聚偏氟乙烯(PVDF)膜上(简称CA-m-PVDF复合膜),在气液膜接触器中使用速率为0.25 m·s−1的水作为吸收剂,CA-m-PVDF复合膜具有较高的CO2通量值(2.5×10−3 mol·m−2·s−1),该CO2通量值比未添加CA时增加约160%.
MDEA吸收CO2的过程中形成不稳定的氨基甲酸盐和碳酸氢根离子,加热时释放出CO2达到解吸和再生的目的. Zhang等[44]将CA通过吸附固定到ZIF-L-1(MOFs)上,得到CA/ZIF-L-1复合材料. 由于ZIF-L-1和CA均含有Zn2+,两者之间存在协同作用,ZIF-L-1中的咪唑基团(mIm)可以作为亲核试剂参与CO2的水合生成碳酸氢盐(6),而且该反应能够促进CO2的液-固传质. 加入0.05 g·L−1 CA/ZIF-L-1后,MDEA中的CO2吸收率提高了2.5倍. Du等[45]进行了类似的研究,把CA包埋到不同粒径的ZIF-8(CA/ZIF-8)中,并将CO2吸收到MDEA中,MDEA的吸收率增加到2.4倍. Xu等[46]将CA固定在表面改性的磁性Fe3O4纳米颗粒(MNP)上,并用原位聚合合成MNP-CA纳米凝胶. 通过反应器测试了MNP-CA纳米凝胶加速MDEA水溶液中CO2吸收的有效性,MNP-CA纳米凝胶在30 ℃的CO2吸收率为不含CA时的170%. 而且MNP-CA纳米凝胶的加入使湿壁柱气相中的总传质系数(KG)在60 ℃时增加了4.61倍. 费潇瑶[47]使用共价结合法将氨基功能化介孔材料固定在CA上(CA/AFS)对用于MDEA溶液解吸CO2进行了研究,固定化CA的加入对CO2平衡解吸量基本没有影响,但CA/AFS的加入可以在未达到平衡状态之前,提高CO2的瞬时解吸量.
H2O+mIm+CO2↔mIm++HCO−3 (6) CO2在碳酸盐溶液中的总反应如下所示(7),CA可有效促进二氧化碳吸收到碳酸盐溶液中[9]. Fabbricino等[48]将嗜热细菌的碳酸酐酶(SspCA)作为膜锚定蛋白固定在大肠杆菌细胞的外膜(INPN-SspCA)上. 在碱性溶液(0.5 mol·L−1 Na2CO3/0.5 mol·L−1 NaHCO3)吸收CO2反应过程中,加入SspCA的膜细胞碎片用作生物催化剂,观察到压力值的迅速下降和比未添加催化剂时更高的CO2吸收率. Peirce等[49]报道了CA在高浓度碳酸盐溶液中容易失活,因此将工业级碳酸酐酶(NovoCA)固定在顺磁性Fe3O4纳米颗粒上进行改善,并分别在25 ℃和40 ℃的条件下,于0.5 mol·L−1 Na2CO3/NaHCO3缓冲液(pH=9.6)和10%wt K2CO3溶液的搅拌池反应器中进行CO2吸收测试[10]. 溶液中固定化CA的Kcat/Km均在5.52×10−1—5.52 L·mol−1·s−1之间. Qi等[50]通过湿壁柱测定表明,在20% wt K2CO3溶剂中加入2 g·L−1 CA后,CO2总传质系数(KG)提高了约5倍. 当CA浓度从零增加到2.5 g·L−1时,CO2捕集效率增加了4.6倍.
CO2−3+H2O+CO2↔2HCO−3 (7) Gladi等[17]探究了在湿壁柱中对4种溶液(MEA、AMP(2-氨基-2-甲基-1-丙醇)、MDEA和K2CO3溶液)添加0.2% wt CA后吸收CO2的影响. 研究表明,添加CA显著增加了MDEA和K2CO3的液侧传质膜系数,AMP略微增加,而MEA没有变化. MDEA的反应速率随温度升高而降低,而在较低的溶剂浓度(5% wt—15 %wt)下,温度和溶剂浓度对K2CO3中反应速率的影响较小,但在20% wt时,温度升高会显着提高反应速率. 此外,混合溶剂对CO2进行吸收比单一溶剂表现出更高的吸收效率,因为它们结合了单一溶剂的优点. Sahoo等[51]将CA共价固定到异质功能化载体(HFS)和锌络合物(Zn-Im)上(CA/Zn-Im:HFS),并将CA/Zn-Im:HFS加入到混合溶剂(30% wt MEA+7.5% wt PZ+15% wt K2CO3)中加快CO2的吸收和解吸. 含有CA/Zn-Im: HFS的混合溶剂(21.65 %wt)在20 min内CO2的吸收量高于纯混合溶剂(13.75 %wt). 含有CA/Zn-Im: HFS的混合溶剂比纯混合溶剂的CO2相对解吸率高1.57倍. 通过表3展示了CA在二氧化碳吸收中的催化性能.
表 3 CA在二氧化碳吸收中的催化性能Table 3. Catalytic performance of CA in carbon dioxide absorption吸收剂成分Absorbent ingredients 温度Temperature 催化特性Catalytic properties 参考文献References 100 mg的固定化CA(固载量为2 mg·g−1)+H2O 30 ℃ 反应进行30 min,CO2吸收量Gv=1.858×10−4 L [31] CA-m-PVDF复合膜+0.25 m·s−1 H2O 25 ℃ CA-m-PVDF复合膜CO2通量值为2.5×10−3 mol·m−2·s−1 [43] 0.05 g·L−1 CA/ZIF-L-1+20 mL 1 mol·L−1 MDEA 40 ℃ MDEA中的CO2吸收率提高了2.5倍 [44] CA/ZIF-8+MDEA 40 ℃ CO2吸收到含有不同粒径的ZIF-8的MDEA溶液中,MDEA溶液的吸收率增加到2.4倍 [45] 10 mg MNP-CA纳米凝胶+100 mL 1 mol·L−1 MDEA 30 ℃ 反应3 h后,MNP-CA纳米凝胶的CO2吸收率为不含CA时的170% [46] 0.3 kg m−3工业级碳酸酐酶(NovoCA)+2—3 mol·L−1 K2CO3(碳酸盐转化度0—40%) 25 ℃和40 ℃ 当加入的酶浓度低于0.018 kg·m−3时,Kcat/Km值介于0.50×102 L·mol−1·s−1和0.39×103 L·mol−1·s−1之间 [49] 0.2×10−2—1.5×10−2 kg NovoCA·m−3 Fe3O4纳米颗粒固定化CA+0.5 mol·L−1 Na2CO3/NaHCO3缓冲液(pH=9.6) 25 ℃和40 ℃ 固定化CA的Kcat/Km值介于4.87—8.06 L·mol−1·s−1之间 [10] 0.2×10−2—1.5×10−2 kg NovoCA·m−3 Fe3O4纳米颗粒固定化CA+10%wt K2CO3溶液(碳酸盐转化度0—40%) 25 ℃ 固定化CA的Kcat/Km值介于3.24—6.73 L·mol−1·s−1之间 [10] 嗜热细菌的全细胞固定(INPN-SspCA)+0.5 mol·L−1 Na2CO3/NaHCO3 25 ℃ INPN-SspCA的Kcat/Km值介于9.94×10−1—3.09 L·mol−1·s−1之间 [48] 2 g·L−1 CA+20% wt K2CO3 40 ℃ 与未添加CA相比,CO2总传质系数(KG)提高了约5倍 [50] 0.22 g·L−1 CA+30% wt K2CO3(pH~11—12) 50 ℃ CA的Kcat/Km值为5.3×108 L·mol−1·s−1 [52] 0.2% wt CA+30% MDEA/15% K2CO3 17—50 ℃ 添加CA显著增加了MDEA和K2CO3的液侧传质膜系数 [17] 0.2% wt CA+30% wt MEA + 7.5% wt PZ+15% wt K2CO3 30 ℃ 含有CA/Zn-Im: HFS的混合溶剂(21.65% wt)在20 min内CO2的吸收量高于纯混合溶剂(13.75% wt) [51] 0.2% wt CA+30% wt MEA +7.5% wt PZ+15% wt K2CO3 90 ℃ 含有CA/Zn-Im: HFS的混合溶剂比纯混合溶剂的CO2相对解吸率高1.57倍. [51] | Show TableDownLoad:
CSV
3.3 多酶复合级联催化CO2
CO2加氢提供了一种碳再生方法,可以通过特定的催化剂转化为甲酸盐、甲醛和甲醇等高价值化学品和燃料. 固定化酶催化因具有反应条件温和、选择性高、可多次催化和效率高等优点,在CO2加氢中引起了广泛关注. 但是,常压下CO2的低溶解度严重限制了酶促反应的速率[53]. CA不仅能够加速CO2的水合,还用于促进酶催化的CO2转化. Wang等[54]报道随着CA的加入,甲酸脱氢酶的底物从CO2转化为更易溶解的
HCO−3 甲酸本身是一种具有商业价值的化学品,它可用作燃料电池的原料和青贮饲料防腐剂,并且二氧化碳形成甲酸所需的能量较低[55]. Zhai等[56]把CA固定在聚乙烯亚胺(PEI)和聚多巴胺(PDA)的改性的二氧化硅微球(PDA/PEI-SiO2-CA)上,并将其添加到含有NADH和FDH的反应溶液中,从而将CO2转化为甲酸盐. 通过改性在SiO2表面引入氨基,氨基与CO2反应形成氨基甲酸酯和碳酸氢盐,因此CA固定在PDA/PEI-SiO2表面上可产生协同效应. 在PDA/PEI-SiO2-CA存在下,初始反应速率是空白对照的48.6倍,甲酸盐产量也高于空白对照. Zhang等[57]通过微生物转谷氨酰胺转胺酶(MTG)充当“交联介质”将CA和FateDH交联在一起形成交联酶. 在二氧化碳转化为甲酸的实验中,交联酶的催化效率可高达游离酶的5.8倍.
固定多酶体系转化CO2生产甲酸已得到广泛研究. 然而,价格昂贵的辅因子(NADH)的再生和循环利用仍然是CO2高效转化的主要问题[58]. Ren等[53]通过将CA、FateDH、GDH(谷氨酸脱氢酶)、PEI(阳离子聚电解质)和辅因子原位包埋到ZIF-8中,构建了纳米级多酶反应器(Co-IMR). NADH通过带正电的PEI和带负电的辅因子之间的离子交换被束缚在ZIF-8中,并通过嵌入ZIF-8中的GDH再生. Co-IMR在8次重复循环使用后仍保留其初次甲酸生产率的50%,表现出良好的重复使用性. 与游离多酶系统相比,Co-IMR在8次重复循环使用后,总累积甲酸产量增加了4.6倍. Chai等[55]进行了类似研究,制备了ZIF-8/CA&FDH薄膜. ZIF-8中的咪唑基团和CA的协同作用使得CO2对甲酸的整体催化活性提高了1.6倍. Wang等[59]将CA、FateDH、GDH包埋在聚多巴胺微胶囊上. 与游离多酶相比,聚多巴胺微胶囊中固定化多酶体系催化产生的HCOOH高4.5倍以上,基于NADH的甲酸产率为342%. Li等[60]使用胺官能化MIL-101(Cr)作为核心,其具有CO2气体吸附性能,再通过包埋将CA、FateDH和GDH 3种酶固定在外层HKUST-1上. 胺官能化的MIL-101(Cr)释放的CO2底物首先进入CA并水合为碳酸氢根离子,然后
HCO−3 甲醇是一种重要的化工原料及能源物质,由合成天然气制备的甲醇程序复杂,且成本较高[8]. 多酶级联催化CO2甲醇化作为一种生物催化手段近年来引起了较多关注. Wang等[61]把FateDH、FaldDH和ADH共固定在聚苯乙烯微粒上,从而将CO2转化为甲醇. 在没有添加CA的情况下,固定多酶体系11次重复循环使用后产生了48%的累计甲醇收率. 甲醇的收率较低,有待进一步改进. Ji等[62]通过将甲酸脱氢酶、甲醛脱氢酶、乙醇脱氢酶和谷氨酸脱氢酶四种酶和辅因子原位包埋在阳离子聚电解质掺杂的中空纳米纤维腔内,并通过在中空纳米纤维的外表面上组装CA加速CO2水合,进行了进一步研究. 对于涉及3种脱氢酶的CO2还原酶系统,实现了103.2%的最高甲醇收率,该反应系统10次重复循环使用后产生了940.5%的累积甲醇收率.
3.4 CA转化CO2的工业应用
基于CA的生物催化系统可以有效地促进CO2的捕集和分离,但只有少数案例与工业过程中的实际应用密切相关. 对于工业应用,CA必须长时间在高温、高离子强度、高pH值和高剪切应力的条件下保持较高活性和稳定性. Lalande等[63]设计并安装了含有固定化CA的CO2填充柱,用于回收和循环利用来自水泥熟料生产的CO2. 固定化CA催化CO2水合形成碳酸氢根离子与添加的CaCl2反应生成的CaCO3被用作波特兰水泥厂的原料. 2015年,在加拿大魁北克省[64]进行了基于CA的CO2捕集工艺的有史以来规模最大的测试. 该测试使用了添加定向进化CA的20% wt的K2CO3/KHCO3溶液,能够在2500 h内以80%的平均捕集效率每天从燃烧天然气的锅炉的燃烧气体中捕集1×104 kg CO2. 实验过程中仅产生少量无毒碱性废水,无需特殊处理直接排入市政下水道. 此外,从汽提塔装置中排出的CO2气体的纯度约99.3%,可用于生产燃料、塑料和化学品等产品. Reardon等[65]在威尔逊维尔的国家碳捕集中心(NCCC)将CA固定在吸收柱的内表面,以20% wt K2CO3和非挥发性碱性盐溶液(AKM-24)作为吸收剂. 固定化CA使传质速率提高了6—7倍,并在40 d 3460 h内保持了超过80%的烟气中CO2的捕集效率. 为了评估CA催化的CO2加速吸收到MDEA水溶液中,在威尔逊维尔的NCCC进行中试规模的CO2捕集实验. 捕集装置在连续6 d吸收温度25—35 ℃和87 ℃解吸温度之间循环,在稳态条件下每天能够捕集多达68.1 kg的二氧化碳[3]. 总而言之,与之前的无生物催化剂过程相比,CA介导的CO2捕集过程具有显著促进作用.
4. 结论与展望(Conclusions and prospects)
为了改善游离CA在高温、强酸或强碱溶剂和化学杂质等条件下容易失活的特性,使用合适的载体和固定化方法对CA进行固定. CO2可通过酶促反应吸收,或通过CA一步催化或级联催化转化为无机碳酸盐、甲酸和甲醇等高价值产品. 固定化CA转化CO2的应用取得了一定的成效,但是仍存在一些技术瓶颈,可总结为以下几个方面:(1)CA价格昂贵. (2)CA对烟气中存在的高温、SOx和NOx耐受性差. (3)固定化方法尚不完善和载体的酶负载量低导致固定化CA活性低.
针对上述问题,未来的研究方向应集中于:(1)从转基因生物中提取大量CA,降低CA生产成本. (2)进一步研究CAs相关基因的生理作用和代谢途径,以提高化学和热稳定性. (3)研发先进的固定化方法和制备具有良好的机械强度、稳定性和对酶亲和力高的载体,提高固定化CA活性.
-
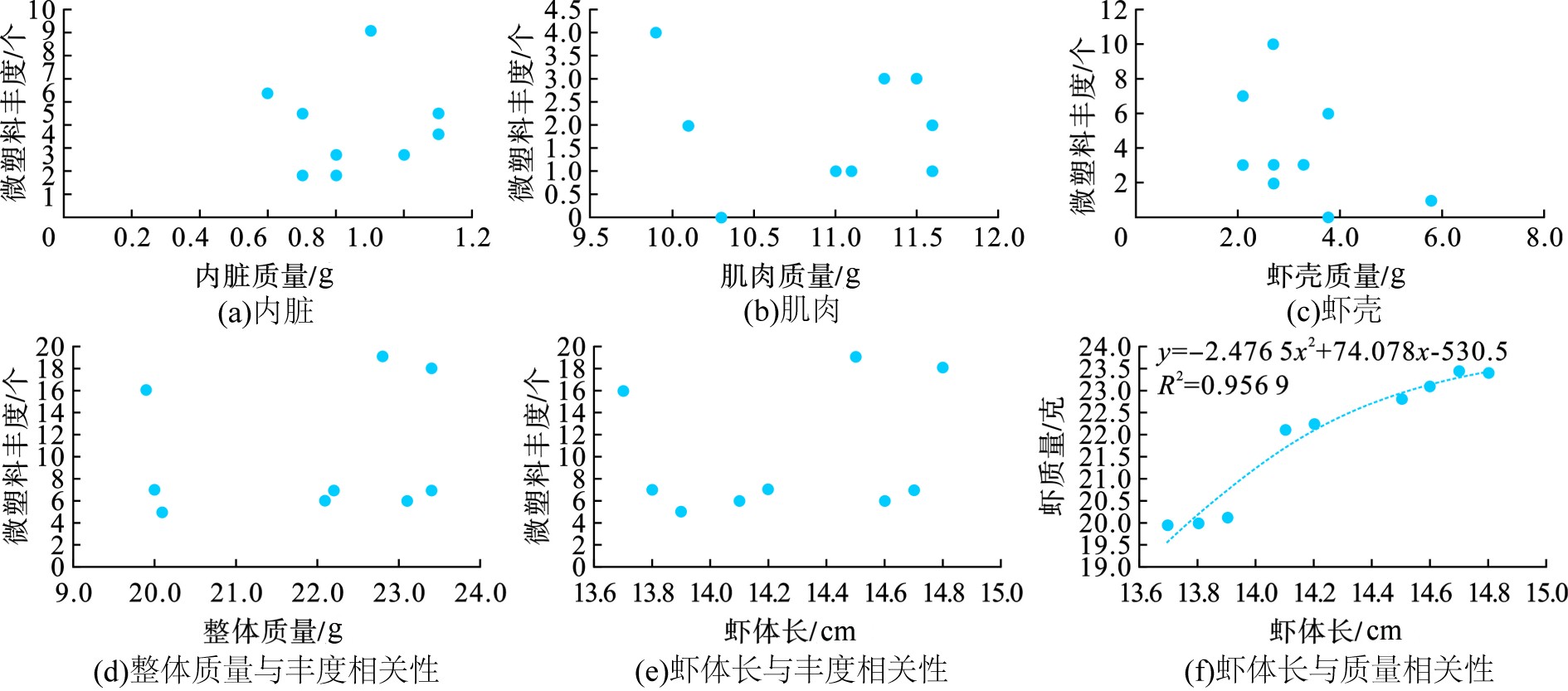
图 2 微塑料丰度与生物体征相关性分析
Figure 2. Correlation between abundance of microplastics and biological indicators
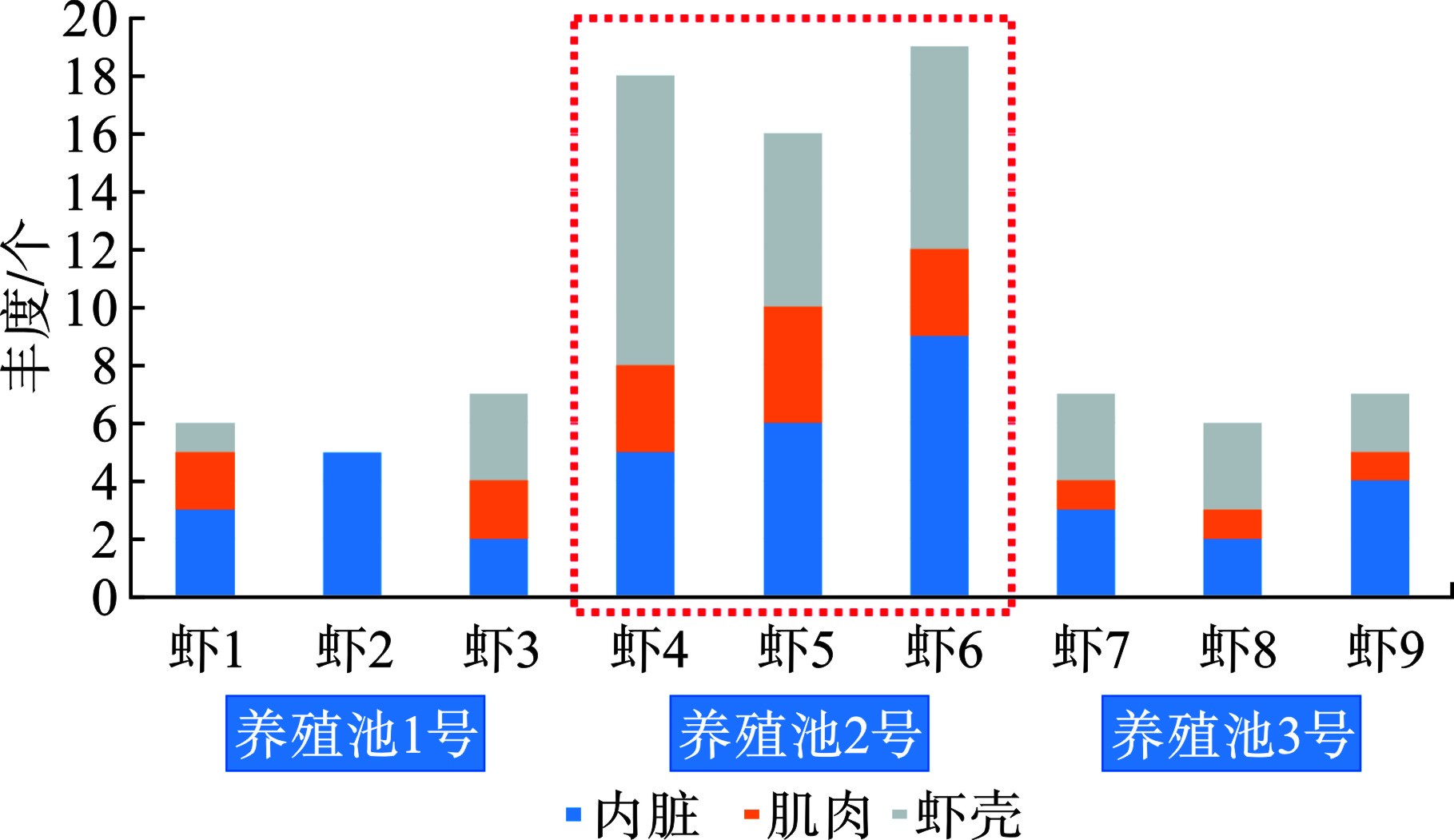
图 3 微塑料丰度与器官分布特征
Figure 3. Characteristics of microplastic abundance and organ distribution
表 1 样品生物学指标
Table 1. Biological indicators of samples
样本 体长/cm 头长/cm 体宽/cm 体高/cm 湿重/g 壳/g 肌肉/g 内脏/g 虾1 14.6 5.9 1.3 2.2 23.1 5.8 11.6 64.5 虾2 13.9 5.2 1.2 1.9 20.1 3.8 10.3 0.7 虾3 13.8 5.1 1.1 1.9 20.0 3.3 10.1 0.7 虾4 14.8 5.7 1.4 2.1 23.4 2.7 11.5 1.1 虾5 13.7 4.9 0.9 1.8 19.9 3.8 9.9 0.6 虾6 14.5 5.8 1.2 2.1 22.8 2.1 11.3 0.9 虾7 14.2 5.2 1.0 2.1 22.2 2.7 11.1 0.8 虾8 14.1 5.2 1.0 2.0 22.1 2.1 11.0 0.8 虾9 14.7 5.7 1.3 2.0 23.4 2.7 11.6 1.1
下载: 导出CSV
表 2 微塑料尺寸器官分布表
Table 2. Distribution of microplastic size in different organs
尺寸特征/μm 内脏/个 肌肉/个 虾壳/个 >50 11 7 5 1~50 28 10 30
下载: 导出CSV
-
[1] ARDUSSO M, LOPEZ A, BUZZI N S, et al. Journal pre-proof covid-19 pandemic repercussions on plastic and antiviral polymeric textile causing pollution on beaches and coasts of south america[J]. Science of The Total Environment, 2020, 763: 144365. [2] THOMPSON RC, OLSEN Y, MITCHELL RP, et al. Lost at sea: where is all the plastic?[J]. Science, 2004, 304(5672): 838. [3] NAPPER I E, BAKIR A, ROWLAND S J, et al. Characterisation, quantity and sorptive properties of microplastics extracted from cosmetics[J]. Marine Pollution Bulletin, 2015, 99(1-2): 178 − 185. [4] ANTHONY L A. Microplastics in the marine environment[J]. Marine Pollution Bulletin, 2011, 62: 1596 − 1605. [5] GRAHAM E R, THOMPSON J T. Deposit- and suspension-feeding sea cucumbers (echinodermata) ingest plastic fragments[J]. Journal of Experimental Marine Biology & Ecology, 2009, 368(1): 22 − 29. [6] MAZURAIS D, ERNANDE B, QUAZUGUEL P, et al. Evaluation of the impact of polyethylene microbeads ingestion in european sea bass (dicentrarchus labrax) larvae[J]. Marine Environmental Research, 2015, 112(DEC. PT. A): 78 − 85. [7] FIONA M, PHILLIP R C. Plastic contamination in the decapod crustacean nephrops norvegicus (linnaeus, 1758)[J]. Marine Pollution Bulletin, 2011, 62(6): 1207 − 1217. [8] WELDEN N A C, COWIE P R. Long-term microplastic retention causes reduced body condition in the langoustine, nephrops norvegicus[J]. Environmental Pollution, 2016, 218(nov.): 895 − 900. [9] WRIGHT S L, THOMPSON R C, GALLOWAY T S. The physical impacts of microplastics on marine organisms: a review[J]. Environmental Pollution, 2013, 178: 483 − 492. [10] BROWNE M A, DISSANAYAKE A, GALLOWAY T S, et al. Ingested microscopic plastic translocates to the circulatory system of the mussel, mytilus edulis (l)[J]. Environmental Science & Technology, 2008, 42(13): 5026 − 5031. [11] AHRENDT C, PEREZ-VENEGAS DJ, URBINA M, et al. Microplastic ingestion cause intestinal lesions in the intertidal fish girella laevifrons[J]. Marine pollution bulletin, 151(Feb. ), 2020: 110795.1 − 110795.6. [12] DELILAH L, ÅKE L, GÖRAN D. Environmental and health hazard ranking and assessment of plastic polymers based on chemical composition[J]. Science of the Total Environment(18), 2011(4): 038. doi: 10.1016/j.scitotenv [13] ROCHMAN C M, HOH E, KUROBE T, et al. Ingested plastic transfers hazardous chemicals to fish and induces hepatic stress[J]. Scientific Reports, 2013, 3: 3263. [14] WARDROP P, SHIMETA J, NUGEGODA D, et al. Chemical pollutants sorbed to ingested microbeads from personal care products accumulate in fish[J]. Environmental Science and Technology, 2016, 50(7): 4037 − 4044. [15] MARK A B, STEWART J N, TAMARA S G, et al. Microplastic Moves Pollutants and Additives to Worms, Reducing Functions Linked to Health and Biodiversity[J]. Current Biology, 2013, 23(2013): 2388 − 2392. [16] GUZZETTI E, SUREDA A, TEJADA S, et al. Microplastic in marine organism: environmental and toxicological effects[J]. Environmental Toxicology and Pharmacology, 2018, 64(12): 164 − 171. [17] NAIK R K, NAIK M M, D'COSTA P M, et al. Microplastics in ballast water as an emerging source and vector for harmful chemicals, antibiotics, metals, bacterial pathogens and hab species: a potential risk to the marine environment and human healt[J]. Marine Pollution Bulletin, 2019, 149: 110525. [18] CURREN E, LEAW C P, LIM P T, et al. Evidence of marine microplastics in commercially harvested seafood[J]. Frontiers in Bioengineering and Biotechnology, 2020, 8: 562760. [19] SEVERINI F, BUZZI N S, LOPEZ A F, et al. Chemical composition and abundance of microplastics in the muscle of commercial shrimp pleoticus muelleri at an impacted coastal environment (southwestern atlantic)[J]. Marine Pollution Bulletin(Dec. Pt. A), 2020: 161. [20] DANIEL D B, ASHRAF P M, THOMAS S N. Abundance, characteristics and seasonal variation of microplastics in indian white shrimps (fenneropenaeus indicus) from coastal waters off cochin, kerala, india[J]. Science of The Total Environment, 2020, 737(11): 139839. [21] HOSSAIN M S , RAHMAN M S , UDDIN M N , et al. Microplastic contamination in penaeid shrimp from the northern bay of bengal[J]. Chemosphere, 2020, 238: 124688. [22] GURJAR U R, XAVIER M, NAYAK B B, et al. Microplastics in shrimps: a study from the trawling grounds of north eastern part of arabian sea[J]. Environmental Science and Pollution Research., 2021, 28: 48494 − 48504. [23] FAO. The State of World Fisheries and Aquaculture Food and Agriculture[J]. Organization of the United Nations, Rome, 2021. [24] 张伟权. 世界重要养殖品种——南美白对虾生物学简介[J]. 海洋科学, 1990(3): 69 − 73. [25] HIDALGO-RUZ V, GUTOW L, THOMPSON RC, et al. Microplastics in the marine environment: a review of the methods used for identification and quantification[J]. Environmental Science & Technology, 2012, 46(6): 3060 − 3075. [26] LEI S, HUIWEN C, PRABHU K, et al. Using the Asian clam as an indicator of microplastic pollution in freshwater ecosystems[J]. Environmental Pollution, 2018: 234. [27] ESTER CC, MARIA C, ANNA SM, et al. Spatial occurrence and effects of microplastic ingestion on the deep-water shrimp aristeus antennatus[J]. Marine Pollution Bulletin, 2018: 133. [28] SABOROWSKI R, KOREZ P, RIESBECK S, et al. Shrimp and microplastics: a case study with the atlantic ditch shrimp palaemon varians[J]. Ecotoxicology and Environmental Safety, 2022, 234: 113394. [29] LUSHER A, HOLLMAN P, MENDOZA-HILL J. Microplastics in fisheries and aquaculture: Status of knowledge on their occurrence and implications for aquatic organisms and food safety[J]. FAO Fisheries and Aquaculture Technical Paper, 2017, 615: 1 − 126. [30] HARA J, FRIAS J, NASH R. Quantification of microplastic ingestion by the decapod crustacean nephrops norvegicus from irish waters[J]. Marine Pollution Bulletin, 2020: 152. [31] YAN M, LI W, CHEN X, et al. A preliminary study of the association between colonization of microorganism on microplastics and intestinal microbiota in shrimp under natural conditions[J]. Journal of Hazardous Materials, 2020, 408: 124882. [32] WALKINSHAW C, LINDEQUE P K, THOMPSON R, et al. Microplastics and seafood: lower trophic organisms at highest risk of contamination[J]. Ecotoxicology and Environmental Safety, 2019, 190: 110066. [33] DEVRIESE L I , DMMD V , MAES T , et al. Microplastic contamination in brown shrimp (crangon crangon, linnaeus 1758) from coastal waters of the southern north sea and channel area[J]. Marine Pollution Bulletin, 2015, 98(1-2): 179 − 187. [34] WITTE B D, DEVRIESE L, BEKAERT K, et al. Quality assessment of the blue mussel (mytilus edulis): comparison between commercial and wild types[J]. Marine Pollution Bulletin, 2014, 85(1): 146 − 155. [35] GBA B, BU B, AM A., et al. Identification of microplastics in fish ponds and natural freshwater environments of the carpathian basin, europe[J]. Chemosphere, 2019, 216: 110 − 116. [36] NICOLAS C O, PAULA S, JOANA L F, et al. Amberstripe scad decapterus muroadsi (carangidae) fish ingest blue microplastics resembling their copepod prey along the coast of rapa nui (easter island) in the south pacific subtropical gyre[J]. Science of The Total Environment, 2017, 586: 430 − 437. [37] CHEN M , JIN M , TAO P , et al. Assessment of microplastics derived from mariculture in xiangshan bay, china[J]. Environmental Pollution, 2018, 242(PT. B): 1146 − 1156. [38] LOTTE K, CASADO-COY N., VALLE C, et al. Plastic debris accumulation in the seabed derived from coastal fish farming[J]. Environmental Pollution, 2019, 257: 113336. [39] WANG J, PENG J, TAN Z, et al. Microplastics in the surface sediments from the beijiang river littoral zone: composition, abundance, surface textures and interaction with heavy metals[J]. Chemosphere, 2017, 171: 248 − 258. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 2342
- HTML全文浏览数: 2342
- PDF下载数: 9
- 施引文献: 0
