















-
随着大气中温室气体浓度的不断增加,气候变暖已成为当下全球面临的主要环境问题[1]。甲烷(CH4)的排放是自然生态系统中不可忽视的问题,其单位分子的增温潜力是CO2的25倍,平均存留时间为CO2的4倍,在自然生态系统碳循环过程中起着重要作用[2-3]。截止2016年,大气中甲烷含量已经达到1.32 mg·m−3。此外,全球生态系统还以每年540 Tg的速度继续向大气释放[4-5]。我国是世界上水库数量最多的国家。截止2017年底,我国已建成水库98 795座,总库容为9 035×108 m3,其容量是我国天然湖泊储水量的3倍左右,约占我国年河川年径流总量(2.8×1012 m3·a−1)的32%[6]。可见水库对于我国水资源的储存有着深远的影响。大量研究[7-8]表明,水库中甲烷的释放也是大气中甲烷浓度升高的一个重要来源。
水库甲烷排放是一个复杂的生物化学过程,产甲烷菌和甲烷氧化菌是影响甲烷产生与排放的主要微生物,且这2种微生物在自然环境中大量存在[9]。目前许多研究主要集中在海洋及淡水湖泊系统中[10],对底泥中甲烷功能菌对甲烷排放的影响还有待进一步研究。
三峡水库是世界上最大的人工水库,目前,对于甲烷排放的研究主要侧重于甲烷传输过程和排放量的测算[11],还未将甲烷排放量数据(宏观)与甲烷功能菌的生长与作用机制(微观)相结合进行深入的研究。因此,本研究拟从微观角度探讨了甲烷功能菌对水库中甲烷排放的影响,以期对大坝生态调度、温室气体排放的研究提供参考。
-
三峡水库是超大型水利工程,兼顾防洪、发电、航运等多种功能。水库东起湖北省宜昌市,西至重庆江津区,形状如同条带,属于河道型水库(图1)。水库正常蓄水水位高程为175 m,水域面积38×1010 m2,总库容393×108 m3,其中防洪库容为221.5×108 m3。三峡水库整体上属于亚热带季风气候,平均温度为18.4 ℃,年降水量为1 000~1 800 mm,分布均匀。三峡库区干流水质良好,除个别地区外,水质整体达到Ⅲ类水标准,总体优于支流水质[12]。大坝建成后,库区反季节的蓄水与泄水,改变了库区干流和支流天然水流条件,库区支流如澎溪河、香溪河、东溪河等的部分河段均出现了不同程度的富营养化现象。其中澎溪河是三峡库区每年水华爆发较为严重的支流。澎溪河作为三峡工程长江中游最大的支流之一,全长180 km,年平均径流量达34.1×108 m3。澎溪河流域属于亚热带季风性湿润气候,年平均气温为18.5 °C,四季气温变化差异大,夏季最高温度可达42 °C,而冬季最低温可低至−4.5 °C。澎溪河因受到回水影响流速较为平缓,部分河段为缓流水体。
-
1)采样方法。根据前期相关研究资料与实地考察,结合三峡水库调度运行方案和各库区支流水生态特征等[13-14],本研究选择了3个采样监测点(图2),分别位于高阳平湖(N31°6′14″,E108°40′26″)、黄石(N30°59′50″,E108°40′51″)、万州(N30°54′40″,E108°31′18″)。
采样时间为2019年7月和9月,采集表层底泥样本,每次采样在1~2 d内完成,主要用于理化指标的测定及向生物公司送检用于微生物分析,其中微生物种类为产甲烷菌和甲烷氧化菌。具体样本信息如表1所示。
采样时,乘坐采样船,到达河道深弘线处(航标船附近)进行采样,记录采样点位对应的具体位置(经纬度)。在现场使用气压计测定大气压,气温计测定气温,UNI-TUT363风速仪测定风速。用彼德森采泥器采集表层沉积物样品,装入聚乙烯密封袋中。为了保证采集样品的代表性和准确性,样品采集与保存均按照《水和废水监测分析方法(第四版)》中相关规范进行。所有样本采集3个平行样品。
2)甲烷通量。水体中甲烷浓度(p(CH4))根据亨利系数、水温、大气压、顶空瓶气液体积比2/3,通过安捷伦7820A气相色谱仪,利用顶空平衡法计算得出,具体方法参见文献[15];采用薄边界模型(TBL)估算法研究水-气界面温室气体通量。根据Fick定律,对于淡水水体,水-气界面气体交换通量(正为释放、负为吸收)可由式(1)计算得出。
式中:
FCH4 为CH4扩散通量,mmol·(m2·h)−1;kx 为气体交换系数,cm·h−1;Cwater 为气体在水中的浓度,mmol·L−1;Cair 为现场温度及压力下大气中CO2的饱和浓度,mmol·L−1。交换系数
kx 的估算用1989年J¨a HNE B等建立的数学经验公式(式(2))[16]。式中:
k600 为六氟化硫(SF6)气体的交换系数,cm·h−1;Sc 为t ℃下CO2的Schmidt常数。考虑采样点均属于已建成水电站的库区范围,本研究采用对于湖泊、水库生态系统的交换系数,选用MACINTYRE等建立的经验公式(式(3))[17]。
式中:
U10 为水面上方10 m处的风速,m·s−1。通常水体上方1 m风速
U1 可用式(4)进行换算[18]:对于淡水而言,可按式(5)进行计算Sc[19]。式中:
t 为水温,℃。监测得到的温室气体在水中的分压乘以亨利系数,可得到温室气体在水中的饱和浓度
Cwater ,可根据式(6)计算[20]。式中:
K0 为亨利系数,即气体溶解度,mol·(L·atm)−1;p(Gas) 为当前水温下的气体分压,μatm。3)引物设计。泥样经预处理后送往上海美吉生物有限公司(Majorbio Biotechnology Company, Shanghai, China)进行高通量测序。在生物公司内使用MPbio FastDNA®试剂盒提取DNA方法提取基因组DNA,以样品的DNA为模板,利用通用引物515F (GTGCCAGCMG CCGCGG)和806R (GGACTAVHVGGGTWTCTA AT)对细菌的16S rRNA基因序列进行扩增。PCR反应参数如下:95 ℃预变性3 min;94 ℃变性30 s,52 ℃退火30 s,72 ℃延伸1.5 min,共35个循环;72 ℃延伸10 min。采用特异性引物来提取mcrA与pmoA目的基因的产甲烷菌与甲烷氧化菌。其中,好氧甲烷氧化菌的PCR扩增参数为:94 ℃预变性5 min;94 ℃变性45 s,56 ℃退火1 min;72 ℃延伸1 min,共30个循环;72 ℃延伸1 min。产甲烷菌的PCR扩增参数为:94 ℃预变性90 s;5 ℃变性30 s,56 ℃退火30 s,72 ℃延伸90 s,共33个循环;最后72 ℃延伸3 min。PCR扩增产物用2%的琼脂糖凝胶电泳进行检测。利用生物公司的MiSeq PE300测序仪(Illumina Inc,USA)完成高通量测序。高通量测序所需引物序列表如表2所示。
-
甲烷通量数据在Excel 2016中进行标准化处理。经过生物公司高通量测序后的细菌数据使用Uparse软件平台(version 7.0
http://drive5.com/uparse/ )根据相似度水平对OTU划分,进行生物信息统计分析;利用IBM SPSS Statistic软件对产甲烷菌群落及甲烷氧化菌群落与甲烷排放通量进行皮尔逊相关性分析。运用Canoco软件对甲烷功能菌与甲烷排放通量进行冗余分析(RedundancyAnalysis,RDA)。 -
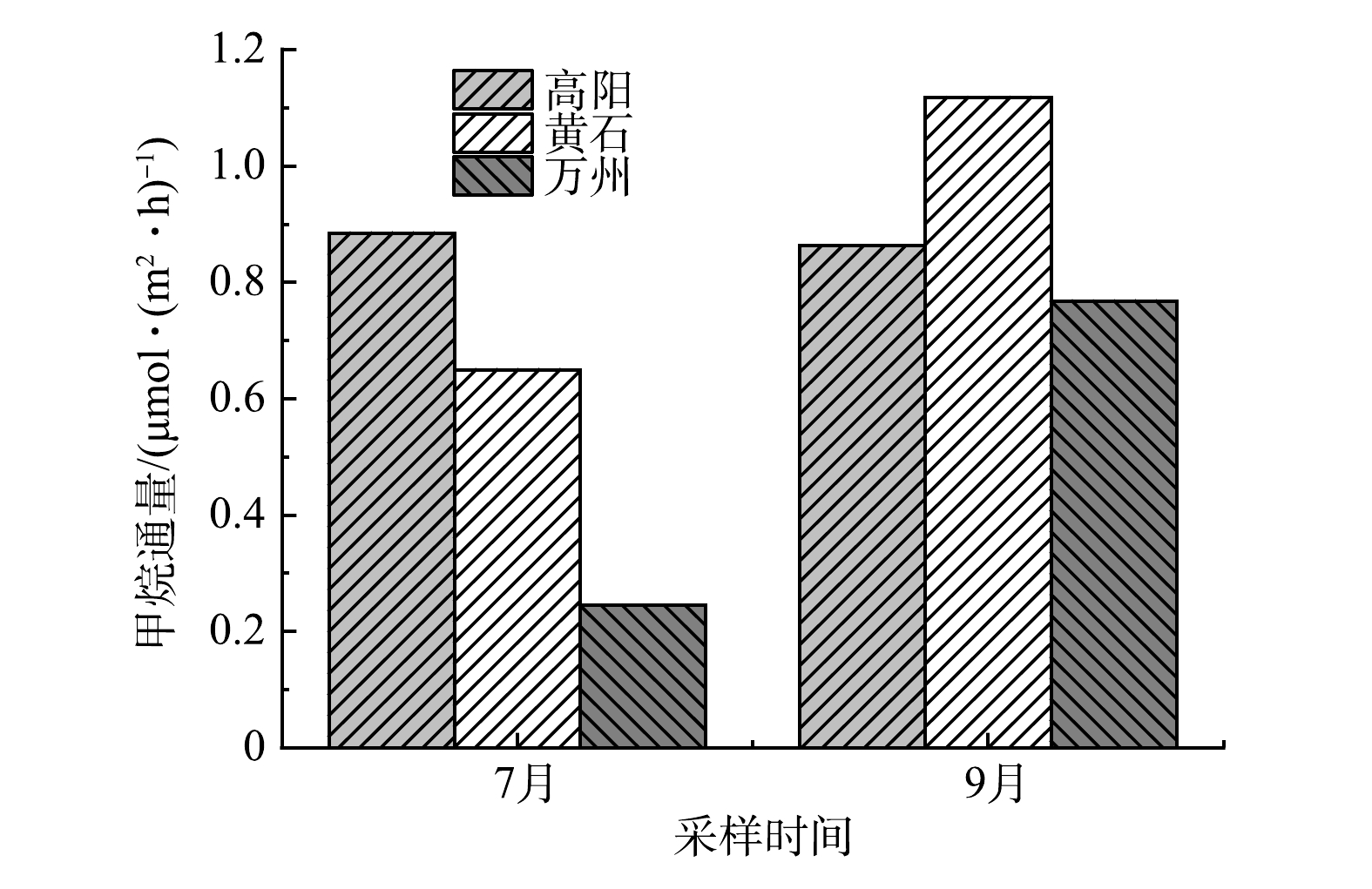
监测期内3个观测点甲烷通量变化如图3所示。高阳、黄石、万州平均甲烷通量为(0.874±0.011)、(0.884±0.234)、(0.507±0.262) μmol·(m2·h)−1,甲烷通量排序为支流黄石>支流高阳>干流万州。从整体上看,干流万州和支流高阳、黄石均向大气中释放甲烷,呈现甲烷“源”的特征。万州观测点在7月达到监测期内最低值,为0.245 μmol·(m2·h)−1;黄石观测点在9月达到监测期内最高值,为1.118 μmol·(m2·h)−1,两地通量在9月均有所上升。而高阳观测点的甲烷通量在7月和9月分别为0.885 μmol·(m2·h)−1和0.863 μmol·(m2·h)−1,通量有所下降,与万州、黄石观测点的通量变化特征不同。从变化幅度来看,支流高阳观测点甲烷通量变化幅度较小,是较稳定的甲烷“源”。而干流万州观测点的变化幅度最大,干流点位7月处于整个研究期间甲烷“源”的最小值。
大量研究表明,河流为明显的温室气体的“源”[22]。本次对三峡库区万州段7月和9月的甲烷通量研究结果表明,甲烷通量的排放表现为“源”,与前人的研究结果一致。通过比较可发现,干流万州点位总体甲烷排放通量低于两支流高阳、黄石点位,在7月万州观测点甲烷通量达到了监测期内的最低值。这与赵登忠[23]的研究结果一致,其研究区域清江流域水布垭水库中干流甲烷排放通量明显比支流甲烷排放通量低得多。两水库的环境特征、水位变化等条件有很强的相似性,可作为参考的依据。干流甲烷排放通量低于支流甲烷排放通量的原因,推测有以下2点。
1)夏季温度较高,相关微生物活性增强,沉积物中有机质分解作用增强。此外,干流沉积物中的甲烷氧化菌相比于支流含量较高,活性增强后氧化甲烷的能力增强,从而降低了向大气排放的甲烷通量[24]。赵登忠[25]的研究表明,甲烷排放通量在9月呈现最大值,这与本文结果一致。干流沉积物中的甲烷氧化菌含量较高,活性增强后氧化甲烷的能力增强,降低了甲烷的排放。
2)在部分支流区域,水体受干流水体顶托作用的影响,形成回水区域,导致水体交换作用减小,水体流速降低,处于准静止状态,水力学条件以及水中营养条件均适合浮游植物的生长,为水体中浮游植物的生长创造了条件[26-27]。浮游植物通过呼吸作用消耗水中的溶解氧,造成溶解氧降低;同时,若出现水华的情况,藻类浓度将会急剧升高,导致在流速缓慢的地方,河流底部溶解氧会出现厌氧情况,有利于甲烷的生成,从而增加了甲烷的释放。自三峡水库修建后,库区水质及生态系统发生变化,由于回水托顶加重了部分支流的污染,水域局部水体出现富营养化[9]。近年来澎溪河、香溪河等支流区域遭受到了严重的水华污染[28]。本研究中高阳、黄石支流属于澎溪河境内,且监测期为水华爆发期,导致甲烷大量产生排放,造成支流甲烷排放通量大于干流的现象。
但在研究期间,高阳观测点出现了与万州、黄石观测点相反的通量变化特征,其甲烷通量在9月由0.885 μmol·(m2·h)−1下降到0.863 μmol·(m2·h)−1,在月平均尺度上出现了甲烷吸收的现象。高阳观测点距离干流更远,受回水顶托影响相比于黄石观测点较小,因此,河流底部的厌氧情况相比黄石较好。有研究[29]表明,在水深较大的区域甲烷排放通量小于浅水区域。高阳观测点水深大于支流回水末端的黄石观测点,因此,甲烷排放通量小于黄石观测点。而甲烷氧化菌对甲烷的氧化过程作为阻止甲烷排放至大气的重要过程,故可推测,因高阳不同于黄石的水文环境,引起高阳观测点底泥中有某种甲烷氧化菌丰度发生改变,在氧化过程中对分解甲烷起到重要作用。
-
在属水平上将底泥中丰度前10的产甲烷菌、甲烷氧化菌与甲烷排放通量作皮尔逊相关性分析分析,结果如表3和表4所示(显著性水平取0.05)。结果表明,底泥中不存在与甲烷排放通量呈显著相关的产甲烷菌物种,而底泥中存在与甲烷排放通量呈显著相关的甲烷氧化菌物种。其中Methylobacter(甲基杆菌)、Methylosarcina(甲基八叠球菌属)与甲烷通量呈显著负相关(P=0.03),其余物种与甲烷排放通量关系不显著。
-
在属水平上将底泥中的产甲烷菌和甲烷氧化菌分别与甲烷排放通量作冗余分析,结果如图4和图5所示。在图4中,样本点WZ1909、HS1909产甲烷菌群落组成与甲烷排放通量呈显著正相关;样本点WZ1907、HS1907产甲烷菌群落组成与甲烷排放通量呈显著负相关。未分类属unclassified_p_Euryarchaeota与甲烷排放通量呈显著正相关,Methanosarcinaceae的未分类属environmental_samples_f_Methanosarcinaceae与甲烷排放通量呈正相关。在图5中,样本点HS1907、HS1909、GY1909甲烷氧化菌群落组成与甲烷排放通量呈显著正相关;样本点WZ1907甲烷氧化菌群落组成与甲烷排放通量呈显著负相关。Ⅰ型甲烷氧化菌Methylobacter、Methylosarcina与甲烷排放通量呈显著负相关,且Methylobacter、Methylosarcina丰度与Ⅱ型甲烷氧化菌Methylocystis (甲基孢囊菌属)呈负相关。
根据产甲烷菌群落与甲烷排放通量的冗余分析,可以得到甲烷八叠球菌科的未分类属environmental_samples_f_Methanosarcinaceae、未分类属unclassified_p_Euryarchaeota以及未命名属norank_c_environmental_samples对甲烷通量排放呈正相关。结合研究区域甲烷排放通量的变化,可以看出,这3种细菌是影响甲烷排放的微生物类群,当这几类物种丰度增加时,甲烷排放量会显著提高。WATANABE[30]等研究表明,甲烷八叠球菌属通过转录mcrA基因,参与了与土壤条件相适应的甲烷产生。IMACHI[31]等发现,在甲烷产生的同时,主要检测到与Euryarchaeota相关的潜在产甲烷类型。这与本研究的结果一致,说明这3类细菌是产甲烷的主要功能菌属。
-
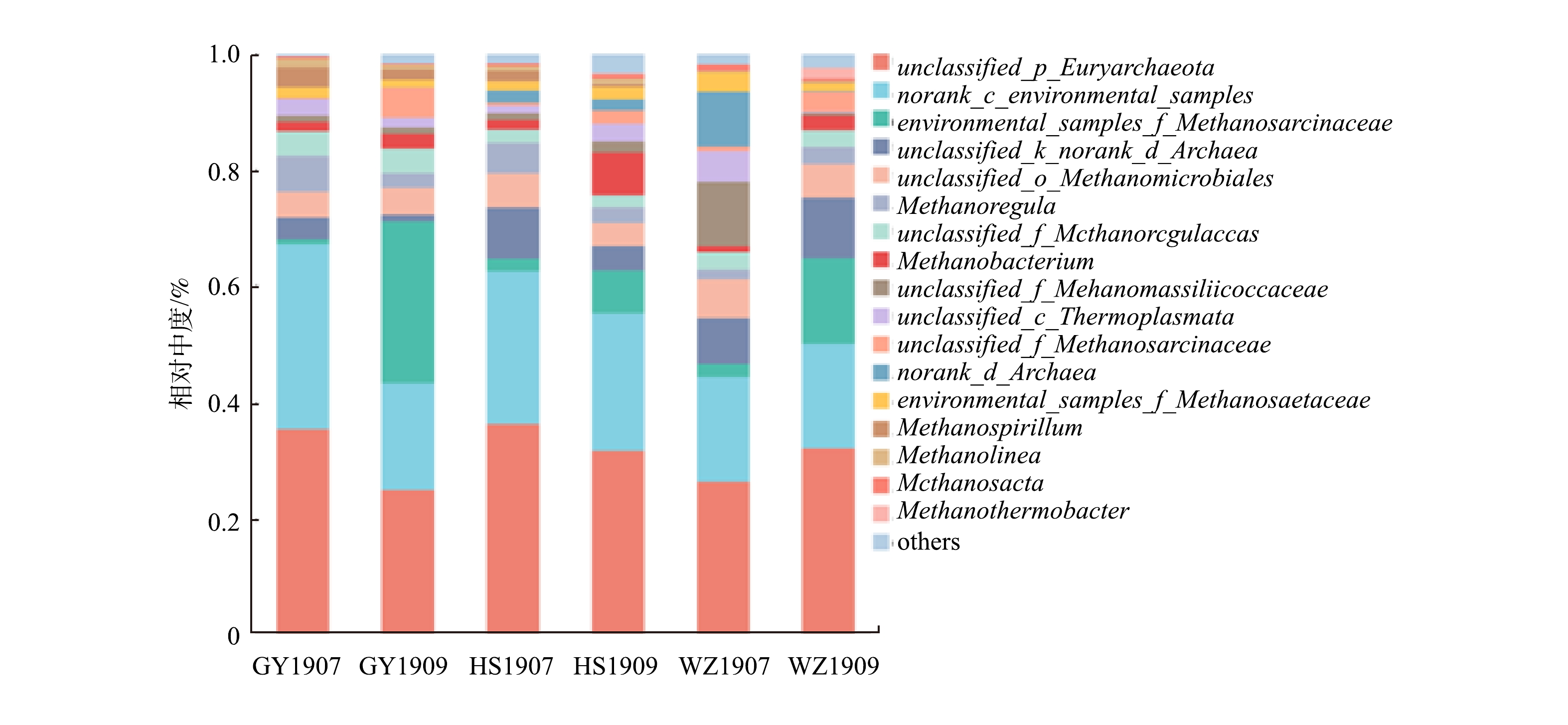
底泥中产甲烷菌与甲烷氧化菌在属水平上的群落组成如图6、图7所示。由图6可以看出,在属水平上,样品中的主要产甲烷菌种有unclassified_p_Euryarchaeota、norank_c_environmental_samples。Euryarchaeota的未分类属unclassified_p_Euryarchaeota在所有样本中为最大优势菌种,unclassified_p_Euryarchaeota占比在GY、HS中7月>9月,在WZ中7月<9月。未分类属environmental_samples_f_Methanosarcinaceae占比在9月占比较大,在7月占比较小,其中在GY1909产甲烷菌群落中为最大占比,达到了27.8%。
产甲烷菌是CH4循环的重要功能菌群,广泛分布于厌氧环境中,在有机物厌氧降解的最后一步将其转化为CH4[32]。本研究发现,未分类属unclassified_p_Euryarchaeota、未命名属norank_c_environmental_samples为研究区的优势菌种。因本研究是在属水平上进行分析,结果出现许多未分类和未命名菌属,尚无法准确了解其具体所属菌属类别。为进一步了解其特性,将部分重要菌属在NCBI上进行基因比对。将unclassified p Euryarchaeota的测序结果提交NCBI上进行BLAST比对,相似性最高的是一种未命名的产甲烷古菌,相似度达到100%,这种未命名菌株在LIU[33]等的文章中被记录过,菌株来自于中国浙江慈溪市沿海地区的潮汐沼泽,其沼泽被开垦用来进行水稻生产。而稻田淹水条件创造的厌氧环境为产甲烷菌提供了理想的生存场所[34],对比本文的采样点,河流底泥中大多为厌氧状态,两者的采样环境有一定的相似性。将norank_c_environmental_samples的测序结果提交NCBI上进行BLAST比对,相似性最高的是一种未命名的产甲烷古菌,相似度达到99.52%。这种未命名菌株在ZHUANG[35]等的文章中被记录,菌株来自于法国西地中海罗纳河以及摩洛哥穆卢亚河中的表层沉积物,采样地与本研究相似,具有一定的参考性。而在本研究区域,产甲烷菌unclassified_p_Euryarchaeota丰度变化在干、支流出现差异,推测原因是受干、支流的环境差异所影响。因8月份重庆雨季来临,降雨量大增,造成库区上游携带大量微生物进入研究区域,而其中含有大量产甲烷菌unclassified_p_Euryarchaeota,从而增加了该产甲烷菌丰度,对干流甲烷通量增加造成了影响。
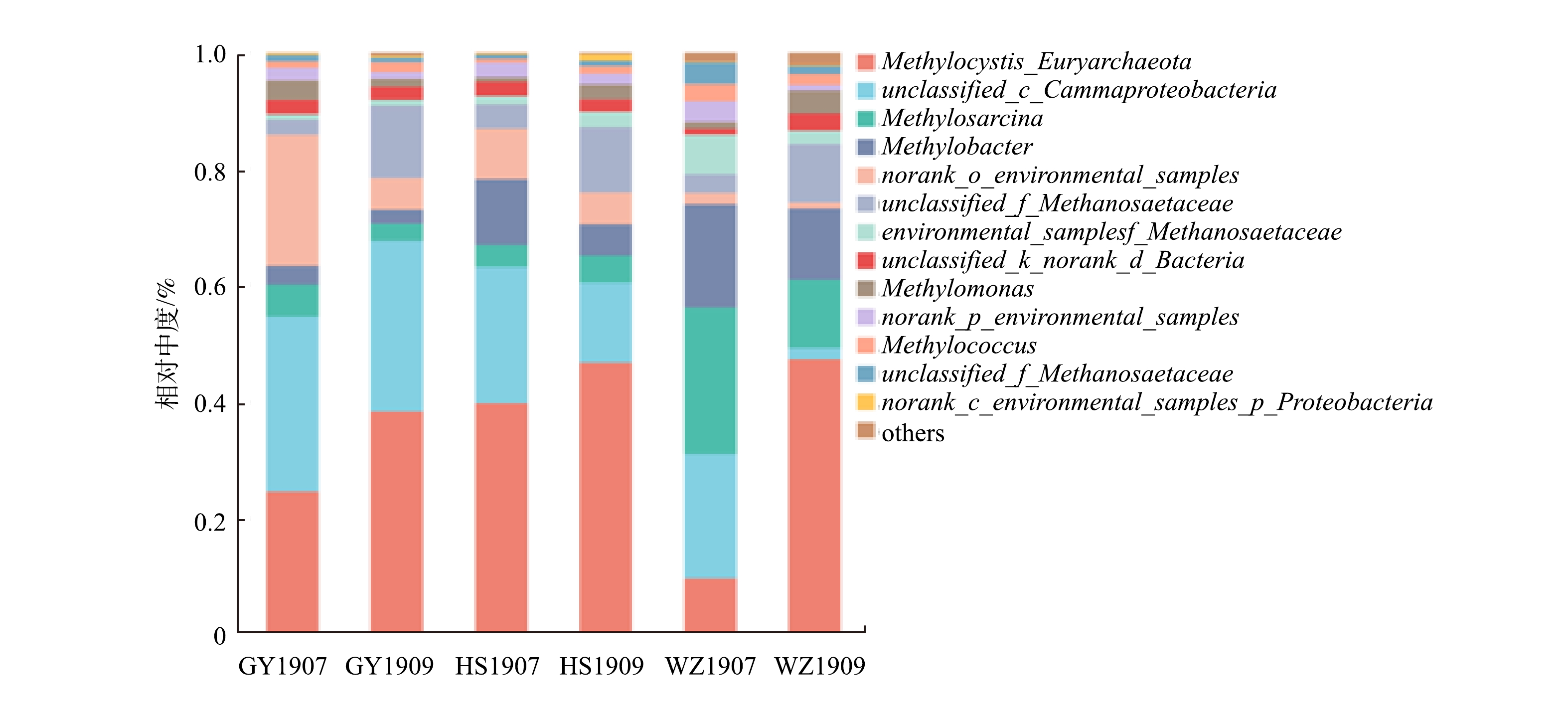
由图7可以看出,样品中的主要甲烷氧化菌种有Methylocystis、unclassified_c_ Gammaproteobacteria、Methylobacter、Methylosarcina。整体来看,研究区域底泥菌群优势菌的占比随月份有较大波动,Methylocystis在7月占比明显小于9月,其中在WZ1907占比小于所有样本点,为9.7%。而Methylobacter、Methylosarcina占比在7月明显大于9月,占比按地区排名为WZ>HS>GY。从地区上看,未命名菌种unclassified_c_ Gammaproteobacteria占比在HS和WZ从7月到9月均有一定程度的减小,而在GY变化不明显。未命名菌种norank_o_environmental_samples在HS、WZ占比很小,而在GY1907占比较大,达到22.5%。
甲烷氧化菌能消耗转化生态系统中约76%~90%的甲烷,是甲烷重要的生物汇,对保持大气甲烷浓度的平衡具有重要的意义[36]。好氧甲烷氧化菌在自然环境中分布广泛,能以甲烷作为碳源和能源,将90%的由产甲烷菌产生的甲烷氧化[37]。本研究的结果表明,Ⅱ型甲烷氧化菌Methylocystis为样品中的最大优势菌属;其次为未命名的γ变形菌unclassified_c_ Gammaproteobacteria以及Methylobacter、Methylosarcina,三者都属于Ⅰ型甲烷氧化菌。在王晓琳[38]的研究中,Ⅱ型甲烷氧化菌Methylocystis以及Ⅰ型甲烷氧化菌Methylobacter为研究区的优势菌属,这与本研究的结果较为一致。不过,也有研究表明,淡水湖泊生态系统中好氧甲烷氧化菌以Ⅰ型甲烷氧化菌Methylobacter、Methylosarcina等为主[39]。马若潺[40]的研究结果也表明,低氧浓度下Ⅰ型甲烷氧化菌占主要优势。这与本研究中Ⅱ型甲烷氧化菌Methylocystis为最大优势菌属结论有所差别。其原因可能是,对于菌种本身来说,溶解性有机碳浓度过高通常会伴随着更高的好氧甲烷氧化菌丰度,可能会出现更高的Ⅱ型好氧甲烷氧化菌比例[41]。而大量的研究表明,湖泊中Ⅰ型甲烷氧化菌主导甲烷氧化过程[42]。在本研究中,结合甲烷氧化菌群落与甲烷排放通量的冗余分析以及相关性分析结果,可以看出,当unclassified_c_Gammaproteobacteria、Methylobacter、Methylosarcina丰度减少时,甲烷排放通量增大。对比在干支流中这3种细菌的丰度变化可以发现,黄石、万州观测点这3种细菌比例减小,高阳观测点基本持平,这符合甲烷通量的变化规律。由此可推测,雨季来临后上游洪水所携带的这3类甲烷氧化菌较少,造成在干流点位的该甲烷氧化菌丰度减少。由于回水顶托现象,干流江水倒灌进支流黄石点位,造成支流黄石点位的该甲烷氧化菌丰度略微减少,从而对甲烷通量造成一定影响。
Ⅰ型好氧甲烷氧化菌对整个湖泊和水柱层的甲烷氧化活动发挥主导作用,对甲烷的循环有着非常显著的影响[43]。在美国康斯坦茨湖[44]以及我国湿地[45]的相关研究均证明,Ⅰ型甲烷氧化菌主导着我国湖泊沉积物中的甲烷氧化过程。有学者在帕文湖缺氧层中检测到了大量的Methylobacter属好氧甲烷氧化菌[46]。OSWALD[47]等运用放射性示踪实验揭示了甲烷氧化的光诱导作用下,看似缺氧的水层存在溶解氧,并且在同一区域发现了Methylobacter属的好氧甲烷氧化菌[48]。有研究者[49]对部分湖泊水样进行功能基因测序,发现Ⅰ型好氧甲烷氧化菌中的Methylobacter属占主导地位。本研究中样本点水华爆发严重,水体底部出现厌氧情况,溶解氧处于低浓度状态,导致Methylobacter属活性增大,从而加速了甲烷氧化,该结论在秦宇[43]的研究中也有类似结果。
-
1)监测期间内,三峡库区高阳、黄石、万州3个观测点平均甲烷通量为(0.874±0.011)、(0.884±0.234)、(0.507±0.262) μmol·(m2·h)−1,总体表现为甲烷排放源。干、支流甲烷通量表现为支流黄石>支流高阳>干流万州。产生此结果的原因是,温度与河流回水顶托作用导致了干、支流甲烷排放的差异。
2)在产甲烷菌群落中,部分未分类的产甲烷古菌unclassified_p_Euryarchaeota、environmental_samples_f_ Methanosarcinaceae以及未命名产甲烷菌对甲烷排放影响较大。当这类物种丰度增加时,可促进甲烷的产生并影响甲烷排放。
3)在甲烷氧化菌群落中,Methylobacter、Methylosarcina以及未分类甲烷氧化菌对甲烷排放影响较大。当该类细菌丰度增加时会加速甲烷氧化,从而减少水-气界面的甲烷排放。
三峡库区夏季万州段底泥甲烷功能菌群落对甲烷排放的影响
Effects of functional methane community in sediments of Wanzhou section of the Three Gorges Reservoir on methane emissions in summer
-
摘要: 三峡水库是世界上最大的人工水库,其潜在甲烷释放近年来备受关注。目前将甲烷排放量与甲烷功能菌的生长与作用机制相结合开展研究的报道较少。为探究三峡库区夏季万州段甲烷功能菌群落对甲烷排放的影响,分别于2019年7月和9月采集了三峡库区万州段底泥,利用16S rRNA基因高通量测序技术,对该区域的万州干流和高阳、黄石支流在属水平上的甲烷功能菌群落组成结构以及甲烷功能菌群落与甲烷排放通量间的关系进行了研究。结果表明:监测期间内高阳、黄石、万州平均甲烷通量为(0.874±0.011)、(0.884±0.234)、(0.507±0.262) μmol·(m2·h)−1,支流大于干流,总体表现为甲烷排放“源”。在产甲烷菌群落中,部分未分类产甲烷古菌unclassified_p_Euryarchaeota、environmental_samples_f_ Methanosarcinaceae以及未命名古菌对产甲烷影响较大,该类细菌可促进甲烷产生。在甲烷氧化菌群落中,Methylobacter、Methylosarcina以及未分类氧化菌对甲烷氧化有较大影响,当该类细菌占比增加时会加速甲烷氧化,从而减少水-气界面中的甲烷排放。除甲烷功能菌群以外,推测温度与河流回水顶托作用也是导致干、支流甲烷排放出现差异的重要因素。以上研究结果可对揭示水库甲烷排放与甲烷功能菌生长和作用机制的关系提供参考。Abstract: The Three Gorges Reservoir is the largest artificial reservoir in the world, and its potential methane release has attracted much attention in recent years. At present, there are few reports on combining methane emission with the growth and action mechanism of methane functional bacteria. In order to explore the impact of functional methane bacterial communities in the Wanzhou section of the Three Gorges reservoir area on methane emissions in summer, sediments from the Wanzhou section of the Three Gorges reservoir area were collected during July and September 2019. 16S rRNA gene high-throughput sequencing technologies were used to study the composition and structure of the methane functional bacteria community at the genus level in Wanzhou mainstream, Gaoyang and Huangshi tributaries in the region, as well as the relationship between the methane functional bacteria community and methane emission fluxes. The results showed that during the monitoring period, the average methane fluxes in Gaoyang, Huangshi, and Wanzhou were (0.874±0.011), (0.884±0.234), (0.507±0.262) μmol·(m2·h)−1, and the tributaries were higher than the main streams, and they were generally methane emission sources. In the methanogen community, some unclassified methanogenic archaea unclassified_p_Euryarchaeota, environmental_samples_f_Methanosarcinaceae and unnamed archaea had a greater impact on methanogenesis than other bacteria, and they could promote methane production. In the methanotrophic community, Methylobacter, Methylosarcina and unclassified oxidizing bacteria had a greater influence on methane oxidation than other bacteria. When their proportion increased, methane oxidation could be accelerated, thereby reducing methane emissions at the water-air interface. In addition to the functional methane flora, it was speculated that temperature and river backwater support were also important factors that could cause the difference in methane emissions between dry and branch streams. The research results lay the foundation for the growth and action mechanism of methane functional bacteria that reveal the temporal and spatial changes of methane emissions from reservoirs.
-
随着大气中温室气体浓度的不断增加,气候变暖已成为当下全球面临的主要环境问题[1]。甲烷(CH4)的排放是自然生态系统中不可忽视的问题,其单位分子的增温潜力是CO2的25倍,平均存留时间为CO2的4倍,在自然生态系统碳循环过程中起着重要作用[2-3]。截止2016年,大气中甲烷含量已经达到1.32 mg·m−3。此外,全球生态系统还以每年540 Tg的速度继续向大气释放[4-5]。我国是世界上水库数量最多的国家。截止2017年底,我国已建成水库98 795座,总库容为9 035×108 m3,其容量是我国天然湖泊储水量的3倍左右,约占我国年河川年径流总量(2.8×1012 m3·a−1)的32%[6]。可见水库对于我国水资源的储存有着深远的影响。大量研究[7-8]表明,水库中甲烷的释放也是大气中甲烷浓度升高的一个重要来源。
水库甲烷排放是一个复杂的生物化学过程,产甲烷菌和甲烷氧化菌是影响甲烷产生与排放的主要微生物,且这2种微生物在自然环境中大量存在[9]。目前许多研究主要集中在海洋及淡水湖泊系统中[10],对底泥中甲烷功能菌对甲烷排放的影响还有待进一步研究。
三峡水库是世界上最大的人工水库,目前,对于甲烷排放的研究主要侧重于甲烷传输过程和排放量的测算[11],还未将甲烷排放量数据(宏观)与甲烷功能菌的生长与作用机制(微观)相结合进行深入的研究。因此,本研究拟从微观角度探讨了甲烷功能菌对水库中甲烷排放的影响,以期对大坝生态调度、温室气体排放的研究提供参考。
1. 材料与方法
1.1 研究区域概况
三峡水库是超大型水利工程,兼顾防洪、发电、航运等多种功能。水库东起湖北省宜昌市,西至重庆江津区,形状如同条带,属于河道型水库(图1)。水库正常蓄水水位高程为175 m,水域面积38×1010 m2,总库容393×108 m3,其中防洪库容为221.5×108 m3。三峡水库整体上属于亚热带季风气候,平均温度为18.4 ℃,年降水量为1 000~1 800 mm,分布均匀。三峡库区干流水质良好,除个别地区外,水质整体达到Ⅲ类水标准,总体优于支流水质[12]。大坝建成后,库区反季节的蓄水与泄水,改变了库区干流和支流天然水流条件,库区支流如澎溪河、香溪河、东溪河等的部分河段均出现了不同程度的富营养化现象。其中澎溪河是三峡库区每年水华爆发较为严重的支流。澎溪河作为三峡工程长江中游最大的支流之一,全长180 km,年平均径流量达34.1×108 m3。澎溪河流域属于亚热带季风性湿润气候,年平均气温为18.5 °C,四季气温变化差异大,夏季最高温度可达42 °C,而冬季最低温可低至−4.5 °C。澎溪河因受到回水影响流速较为平缓,部分河段为缓流水体。
1.2 实验方法
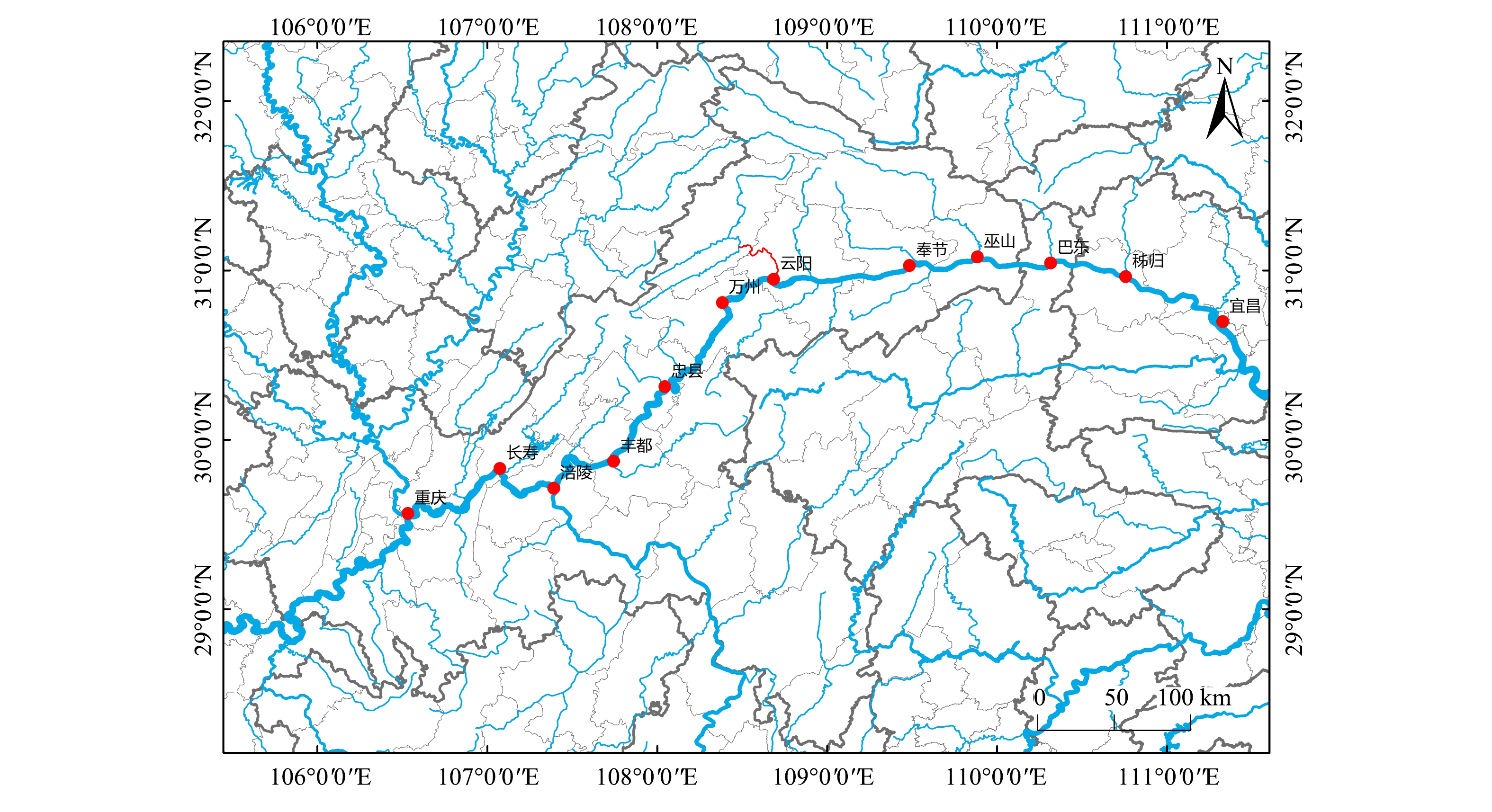
1)采样方法。根据前期相关研究资料与实地考察,结合三峡水库调度运行方案和各库区支流水生态特征等[13-14],本研究选择了3个采样监测点(图2),分别位于高阳平湖(N31°6′14″,E108°40′26″)、黄石(N30°59′50″,E108°40′51″)、万州(N30°54′40″,E108°31′18″)。
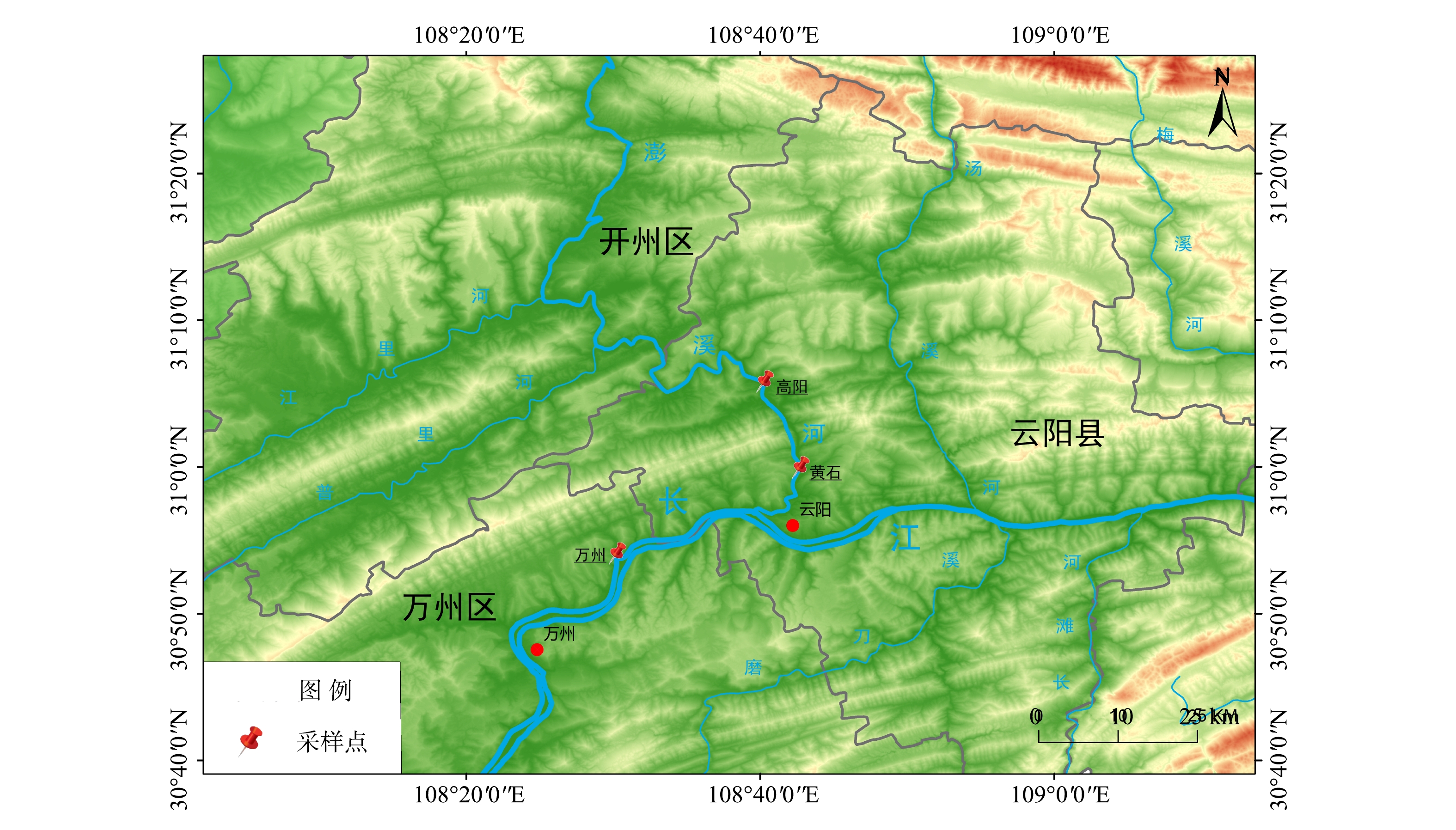 图 2 三峡库区万州段采样点位流域图Figure 2. Location map of sampling sites in Wanzhou section of the Three Gorges Reservoir area
图 2 三峡库区万州段采样点位流域图Figure 2. Location map of sampling sites in Wanzhou section of the Three Gorges Reservoir area采样时间为2019年7月和9月,采集表层底泥样本,每次采样在1~2 d内完成,主要用于理化指标的测定及向生物公司送检用于微生物分析,其中微生物种类为产甲烷菌和甲烷氧化菌。具体样本信息如表1所示。
表 1 样本信息表Table 1. Sample information table采样时间 样本名称 采样时间及地点 2019年7月 WZ1907 2019年7月万州泥样 GY1907 2019年7月高阳泥样 HS1907 2019年7月黄石泥样 2019年9月 WZ1909 2019年9月万州泥样 GY1909 2019年9月高阳泥样 HS1909 2019年9月黄石泥样 | Show Table DownLoad:
CSV
DownLoad:
CSV
采样时,乘坐采样船,到达河道深弘线处(航标船附近)进行采样,记录采样点位对应的具体位置(经纬度)。在现场使用气压计测定大气压,气温计测定气温,UNI-TUT363风速仪测定风速。用彼德森采泥器采集表层沉积物样品,装入聚乙烯密封袋中。为了保证采集样品的代表性和准确性,样品采集与保存均按照《水和废水监测分析方法(第四版)》中相关规范进行。所有样本采集3个平行样品。
2)甲烷通量。水体中甲烷浓度(p(CH4))根据亨利系数、水温、大气压、顶空瓶气液体积比2/3,通过安捷伦7820A气相色谱仪,利用顶空平衡法计算得出,具体方法参见文献[15];采用薄边界模型(TBL)估算法研究水-气界面温室气体通量。根据Fick定律,对于淡水水体,水-气界面气体交换通量(正为释放、负为吸收)可由式(1)计算得出。
FCH4=kx(Cwater−Cair) (1) 式中:
FCH4 kx Cwater Cair 交换系数
kx ¨a kx=k600(600Sc) (2) 式中:
k600 Sc 考虑采样点均属于已建成水电站的库区范围,本研究采用对于湖泊、水库生态系统的交换系数,选用MACINTYRE等建立的经验公式(式(3))[17]。
k600=0.45U1.6410 (3) 式中:
U10 通常水体上方1 m风速
U1 U10=1.22U1 (4) Sc=1897.8−114.28t+3.2902t2−0.03906t3 (5) 式中:
t 监测得到的温室气体在水中的分压乘以亨利系数,可得到温室气体在水中的饱和浓度
Cwater Cwater=K0·p(Gas) (6) 式中:
K0 p(Gas) 3)引物设计。泥样经预处理后送往上海美吉生物有限公司(Majorbio Biotechnology Company, Shanghai, China)进行高通量测序。在生物公司内使用MPbio FastDNA®试剂盒提取DNA方法提取基因组DNA,以样品的DNA为模板,利用通用引物515F (GTGCCAGCMG CCGCGG)和806R (GGACTAVHVGGGTWTCTA AT)对细菌的16S rRNA基因序列进行扩增。PCR反应参数如下:95 ℃预变性3 min;94 ℃变性30 s,52 ℃退火30 s,72 ℃延伸1.5 min,共35个循环;72 ℃延伸10 min。采用特异性引物来提取mcrA与pmoA目的基因的产甲烷菌与甲烷氧化菌。其中,好氧甲烷氧化菌的PCR扩增参数为:94 ℃预变性5 min;94 ℃变性45 s,56 ℃退火1 min;72 ℃延伸1 min,共30个循环;72 ℃延伸1 min。产甲烷菌的PCR扩增参数为:94 ℃预变性90 s;5 ℃变性30 s,56 ℃退火30 s,72 ℃延伸90 s,共33个循环;最后72 ℃延伸3 min。PCR扩增产物用2%的琼脂糖凝胶电泳进行检测。利用生物公司的MiSeq PE300测序仪(Illumina Inc,USA)完成高通量测序。高通量测序所需引物序列表如表2所示。
表 2 高通量测序引物序列表[21]Table 2. High-throughput sequencing primer sequence list目的基因 引物 引物序列 (5’-3’) 16Sr RNA (细菌) 515FmodF GTGCCAGCMGCCGCGG 806RmodR GGACTAVHVGGGTWTCTAAT pmoA (好氧甲烷氧化菌) A189F GGNGACTGGGACTTCTGG Mb661R GGTAARGACGTTGCNCCGG mcrA (产甲烷菌) MLf GCCCGGTGGTGTMGGATTCACACARTAYGCWACAGC MLr TTCATTGCRTAGTTWGGRTAGTT | Show TableDownLoad:
CSV
1.3 数据处理
甲烷通量数据在Excel 2016中进行标准化处理。经过生物公司高通量测序后的细菌数据使用Uparse软件平台(version 7.0
http://drive5.com/uparse/ )根据相似度水平对OTU划分,进行生物信息统计分析;利用IBM SPSS Statistic软件对产甲烷菌群落及甲烷氧化菌群落与甲烷排放通量进行皮尔逊相关性分析。运用Canoco软件对甲烷功能菌与甲烷排放通量进行冗余分析(RedundancyAnalysis,RDA)。2. 结果与讨论
2.1 甲烷排放通量特征
监测期内3个观测点甲烷通量变化如图3所示。高阳、黄石、万州平均甲烷通量为(0.874±0.011)、(0.884±0.234)、(0.507±0.262) μmol·(m2·h)−1,甲烷通量排序为支流黄石>支流高阳>干流万州。从整体上看,干流万州和支流高阳、黄石均向大气中释放甲烷,呈现甲烷“源”的特征。万州观测点在7月达到监测期内最低值,为0.245 μmol·(m2·h)−1;黄石观测点在9月达到监测期内最高值,为1.118 μmol·(m2·h)−1,两地通量在9月均有所上升。而高阳观测点的甲烷通量在7月和9月分别为0.885 μmol·(m2·h)−1和0.863 μmol·(m2·h)−1,通量有所下降,与万州、黄石观测点的通量变化特征不同。从变化幅度来看,支流高阳观测点甲烷通量变化幅度较小,是较稳定的甲烷“源”。而干流万州观测点的变化幅度最大,干流点位7月处于整个研究期间甲烷“源”的最小值。
大量研究表明,河流为明显的温室气体的“源”[22]。本次对三峡库区万州段7月和9月的甲烷通量研究结果表明,甲烷通量的排放表现为“源”,与前人的研究结果一致。通过比较可发现,干流万州点位总体甲烷排放通量低于两支流高阳、黄石点位,在7月万州观测点甲烷通量达到了监测期内的最低值。这与赵登忠[23]的研究结果一致,其研究区域清江流域水布垭水库中干流甲烷排放通量明显比支流甲烷排放通量低得多。两水库的环境特征、水位变化等条件有很强的相似性,可作为参考的依据。干流甲烷排放通量低于支流甲烷排放通量的原因,推测有以下2点。
1)夏季温度较高,相关微生物活性增强,沉积物中有机质分解作用增强。此外,干流沉积物中的甲烷氧化菌相比于支流含量较高,活性增强后氧化甲烷的能力增强,从而降低了向大气排放的甲烷通量[24]。赵登忠[25]的研究表明,甲烷排放通量在9月呈现最大值,这与本文结果一致。干流沉积物中的甲烷氧化菌含量较高,活性增强后氧化甲烷的能力增强,降低了甲烷的排放。
2)在部分支流区域,水体受干流水体顶托作用的影响,形成回水区域,导致水体交换作用减小,水体流速降低,处于准静止状态,水力学条件以及水中营养条件均适合浮游植物的生长,为水体中浮游植物的生长创造了条件[26-27]。浮游植物通过呼吸作用消耗水中的溶解氧,造成溶解氧降低;同时,若出现水华的情况,藻类浓度将会急剧升高,导致在流速缓慢的地方,河流底部溶解氧会出现厌氧情况,有利于甲烷的生成,从而增加了甲烷的释放。自三峡水库修建后,库区水质及生态系统发生变化,由于回水托顶加重了部分支流的污染,水域局部水体出现富营养化[9]。近年来澎溪河、香溪河等支流区域遭受到了严重的水华污染[28]。本研究中高阳、黄石支流属于澎溪河境内,且监测期为水华爆发期,导致甲烷大量产生排放,造成支流甲烷排放通量大于干流的现象。
但在研究期间,高阳观测点出现了与万州、黄石观测点相反的通量变化特征,其甲烷通量在9月由0.885 μmol·(m2·h)−1下降到0.863 μmol·(m2·h)−1,在月平均尺度上出现了甲烷吸收的现象。高阳观测点距离干流更远,受回水顶托影响相比于黄石观测点较小,因此,河流底部的厌氧情况相比黄石较好。有研究[29]表明,在水深较大的区域甲烷排放通量小于浅水区域。高阳观测点水深大于支流回水末端的黄石观测点,因此,甲烷排放通量小于黄石观测点。而甲烷氧化菌对甲烷的氧化过程作为阻止甲烷排放至大气的重要过程,故可推测,因高阳不同于黄石的水文环境,引起高阳观测点底泥中有某种甲烷氧化菌丰度发生改变,在氧化过程中对分解甲烷起到重要作用。
2.2 底泥中甲烷功能菌群落与甲烷排放通量相关性
在属水平上将底泥中丰度前10的产甲烷菌、甲烷氧化菌与甲烷排放通量作皮尔逊相关性分析分析,结果如表3和表4所示(显著性水平取0.05)。结果表明,底泥中不存在与甲烷排放通量呈显著相关的产甲烷菌物种,而底泥中存在与甲烷排放通量呈显著相关的甲烷氧化菌物种。其中Methylobacter(甲基杆菌)、Methylosarcina(甲基八叠球菌属)与甲烷通量呈显著负相关(P=0.03),其余物种与甲烷排放通量关系不显著。
表 3 产甲烷菌群落与甲烷排放通量相关性分析Table 3. Correlation analysis between methanogenic bacterial communities and methane emission flux产甲烷菌群落 (属) P 值 相关系数 r unclassified_p__Euryarchaeota 0.174 0.636 norank_c__environmental_samples 0.122 0.700 environmental_samples_f__Methanos-arcinaceae 0.514 0.337 unclassified_k__norank_d__Archaea 0.481 −0.362 unclassified_o__Methanomicrobiales 0.276 −0.533 Methanoregula 0.311 0.501 unclassified_f__Methanoregulaceae 0.715 0.192 Methanobacterium 0.115 0.709 unclassified_f__Methanomassiliicocc-aceae 0.073 −0.771 unclassified_c__Thermoplasmata 0.563 −0.300 | Show TableDownLoad:
CSV
表 4 甲烷氧化菌群落与甲烷排放通量相关性分析Table 4. Correlation analysis between methane oxidizing bacteria community and methane emission flux甲烷氧化菌群落 (属) P 值 相关系数 r Methylocystis 0.141 0.651 unclassified_c__Gammaproteobacteria 0.763 −0.159 Methylosarcina 0.033* −0.847 Methylobacter 0.031* −0.853 norank_o__environmental_samples 0.581 0.287 unclassified_f__Methylocystaceae 0.214 0.593 environmental_samples_f__Methylococ-caceae 0.080 -0.759 unclassified_k__norank_d__Bacteria 0.275 0.534 Methylomonas 0.333 0.482 norank_p__environmental_samples 0.110 −0.715 注:“*”表示P<0.05,显著相关。 | Show TableDownLoad:
CSV
2.3 底泥中甲烷功能菌群落与甲烷排放通量的冗余分析
在属水平上将底泥中的产甲烷菌和甲烷氧化菌分别与甲烷排放通量作冗余分析,结果如图4和图5所示。在图4中,样本点WZ1909、HS1909产甲烷菌群落组成与甲烷排放通量呈显著正相关;样本点WZ1907、HS1907产甲烷菌群落组成与甲烷排放通量呈显著负相关。未分类属unclassified_p_Euryarchaeota与甲烷排放通量呈显著正相关,Methanosarcinaceae的未分类属environmental_samples_f_Methanosarcinaceae与甲烷排放通量呈正相关。在图5中,样本点HS1907、HS1909、GY1909甲烷氧化菌群落组成与甲烷排放通量呈显著正相关;样本点WZ1907甲烷氧化菌群落组成与甲烷排放通量呈显著负相关。Ⅰ型甲烷氧化菌Methylobacter、Methylosarcina与甲烷排放通量呈显著负相关,且Methylobacter、Methylosarcina丰度与Ⅱ型甲烷氧化菌Methylocystis (甲基孢囊菌属)呈负相关。
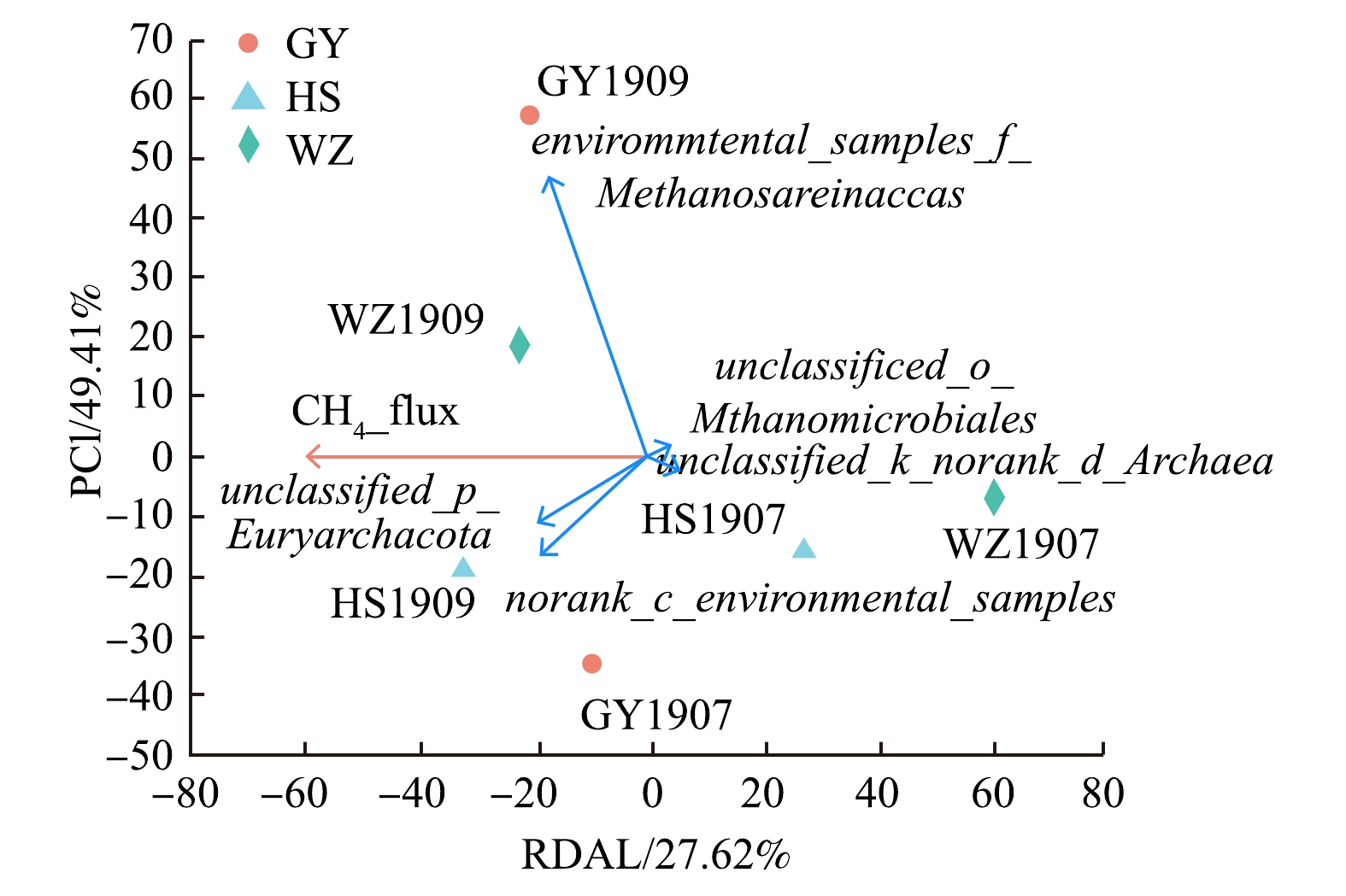 图 4 产甲烷菌群落与甲烷排放通量的冗余分析Figure 4. Redundant analysis of methanogenic bacterial community and methane emission flux
图 4 产甲烷菌群落与甲烷排放通量的冗余分析Figure 4. Redundant analysis of methanogenic bacterial community and methane emission flux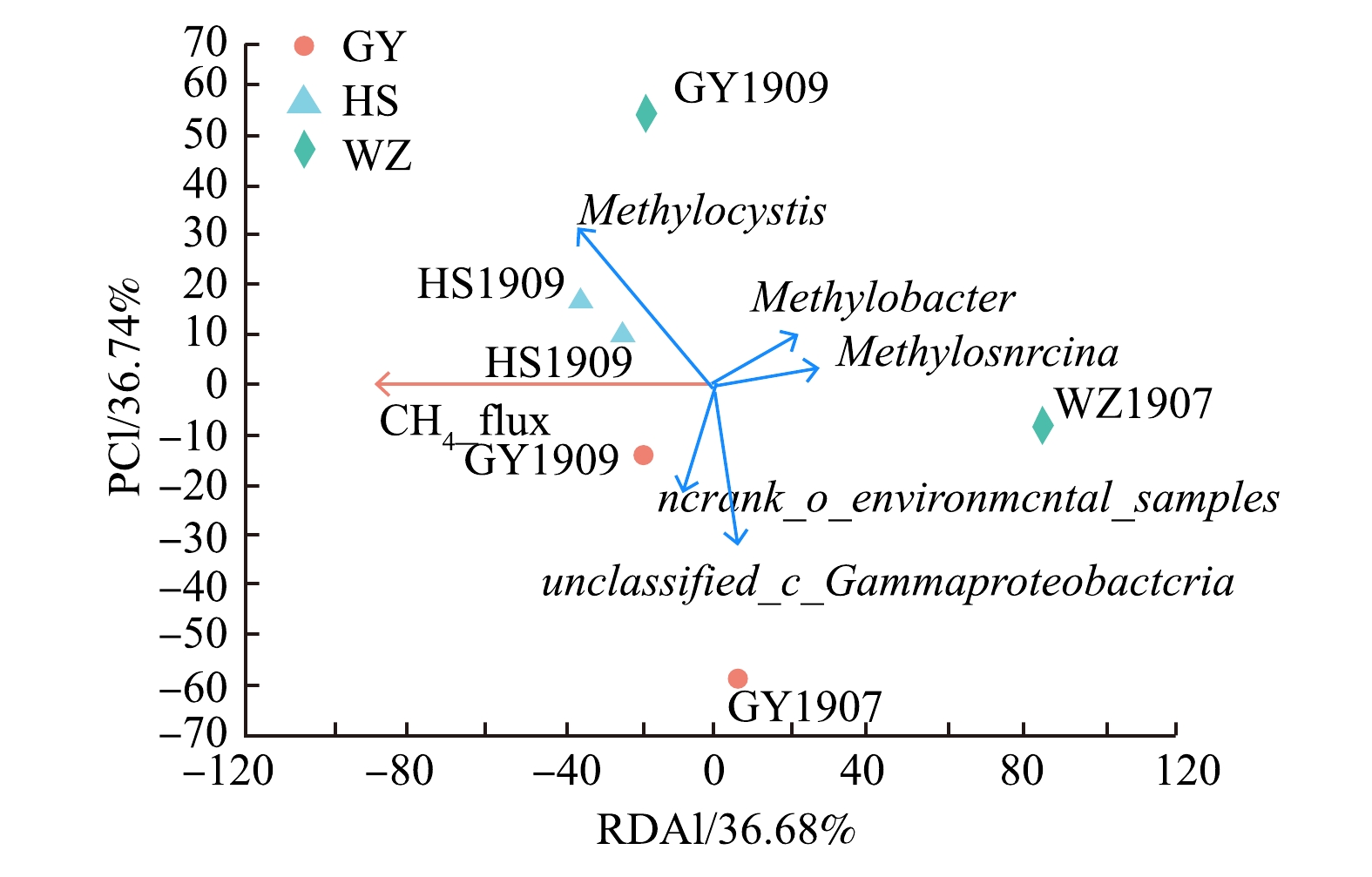 图 5 甲烷氧化菌群落与甲烷排放通量的冗余分析Figure 5. Redundant analysis of methane oxidizing bacteria community and methane emission flux
图 5 甲烷氧化菌群落与甲烷排放通量的冗余分析Figure 5. Redundant analysis of methane oxidizing bacteria community and methane emission flux根据产甲烷菌群落与甲烷排放通量的冗余分析,可以得到甲烷八叠球菌科的未分类属environmental_samples_f_Methanosarcinaceae、未分类属unclassified_p_Euryarchaeota以及未命名属norank_c_environmental_samples对甲烷通量排放呈正相关。结合研究区域甲烷排放通量的变化,可以看出,这3种细菌是影响甲烷排放的微生物类群,当这几类物种丰度增加时,甲烷排放量会显著提高。WATANABE[30]等研究表明,甲烷八叠球菌属通过转录mcrA基因,参与了与土壤条件相适应的甲烷产生。IMACHI[31]等发现,在甲烷产生的同时,主要检测到与Euryarchaeota相关的潜在产甲烷类型。这与本研究的结果一致,说明这3类细菌是产甲烷的主要功能菌属。
2.4 甲烷功能菌群落组成分析
底泥中产甲烷菌与甲烷氧化菌在属水平上的群落组成如图6、图7所示。由图6可以看出,在属水平上,样品中的主要产甲烷菌种有unclassified_p_Euryarchaeota、norank_c_environmental_samples。Euryarchaeota的未分类属unclassified_p_Euryarchaeota在所有样本中为最大优势菌种,unclassified_p_Euryarchaeota占比在GY、HS中7月>9月,在WZ中7月<9月。未分类属environmental_samples_f_Methanosarcinaceae占比在9月占比较大,在7月占比较小,其中在GY1909产甲烷菌群落中为最大占比,达到了27.8%。
产甲烷菌是CH4循环的重要功能菌群,广泛分布于厌氧环境中,在有机物厌氧降解的最后一步将其转化为CH4[32]。本研究发现,未分类属unclassified_p_Euryarchaeota、未命名属norank_c_environmental_samples为研究区的优势菌种。因本研究是在属水平上进行分析,结果出现许多未分类和未命名菌属,尚无法准确了解其具体所属菌属类别。为进一步了解其特性,将部分重要菌属在NCBI上进行基因比对。将unclassified p Euryarchaeota的测序结果提交NCBI上进行BLAST比对,相似性最高的是一种未命名的产甲烷古菌,相似度达到100%,这种未命名菌株在LIU[33]等的文章中被记录过,菌株来自于中国浙江慈溪市沿海地区的潮汐沼泽,其沼泽被开垦用来进行水稻生产。而稻田淹水条件创造的厌氧环境为产甲烷菌提供了理想的生存场所[34],对比本文的采样点,河流底泥中大多为厌氧状态,两者的采样环境有一定的相似性。将norank_c_environmental_samples的测序结果提交NCBI上进行BLAST比对,相似性最高的是一种未命名的产甲烷古菌,相似度达到99.52%。这种未命名菌株在ZHUANG[35]等的文章中被记录,菌株来自于法国西地中海罗纳河以及摩洛哥穆卢亚河中的表层沉积物,采样地与本研究相似,具有一定的参考性。而在本研究区域,产甲烷菌unclassified_p_Euryarchaeota丰度变化在干、支流出现差异,推测原因是受干、支流的环境差异所影响。因8月份重庆雨季来临,降雨量大增,造成库区上游携带大量微生物进入研究区域,而其中含有大量产甲烷菌unclassified_p_Euryarchaeota,从而增加了该产甲烷菌丰度,对干流甲烷通量增加造成了影响。
由图7可以看出,样品中的主要甲烷氧化菌种有Methylocystis、unclassified_c_ Gammaproteobacteria、Methylobacter、Methylosarcina。整体来看,研究区域底泥菌群优势菌的占比随月份有较大波动,Methylocystis在7月占比明显小于9月,其中在WZ1907占比小于所有样本点,为9.7%。而Methylobacter、Methylosarcina占比在7月明显大于9月,占比按地区排名为WZ>HS>GY。从地区上看,未命名菌种unclassified_c_ Gammaproteobacteria占比在HS和WZ从7月到9月均有一定程度的减小,而在GY变化不明显。未命名菌种norank_o_environmental_samples在HS、WZ占比很小,而在GY1907占比较大,达到22.5%。
甲烷氧化菌能消耗转化生态系统中约76%~90%的甲烷,是甲烷重要的生物汇,对保持大气甲烷浓度的平衡具有重要的意义[36]。好氧甲烷氧化菌在自然环境中分布广泛,能以甲烷作为碳源和能源,将90%的由产甲烷菌产生的甲烷氧化[37]。本研究的结果表明,Ⅱ型甲烷氧化菌Methylocystis为样品中的最大优势菌属;其次为未命名的γ变形菌unclassified_c_ Gammaproteobacteria以及Methylobacter、Methylosarcina,三者都属于Ⅰ型甲烷氧化菌。在王晓琳[38]的研究中,Ⅱ型甲烷氧化菌Methylocystis以及Ⅰ型甲烷氧化菌Methylobacter为研究区的优势菌属,这与本研究的结果较为一致。不过,也有研究表明,淡水湖泊生态系统中好氧甲烷氧化菌以Ⅰ型甲烷氧化菌Methylobacter、Methylosarcina等为主[39]。马若潺[40]的研究结果也表明,低氧浓度下Ⅰ型甲烷氧化菌占主要优势。这与本研究中Ⅱ型甲烷氧化菌Methylocystis为最大优势菌属结论有所差别。其原因可能是,对于菌种本身来说,溶解性有机碳浓度过高通常会伴随着更高的好氧甲烷氧化菌丰度,可能会出现更高的Ⅱ型好氧甲烷氧化菌比例[41]。而大量的研究表明,湖泊中Ⅰ型甲烷氧化菌主导甲烷氧化过程[42]。在本研究中,结合甲烷氧化菌群落与甲烷排放通量的冗余分析以及相关性分析结果,可以看出,当unclassified_c_Gammaproteobacteria、Methylobacter、Methylosarcina丰度减少时,甲烷排放通量增大。对比在干支流中这3种细菌的丰度变化可以发现,黄石、万州观测点这3种细菌比例减小,高阳观测点基本持平,这符合甲烷通量的变化规律。由此可推测,雨季来临后上游洪水所携带的这3类甲烷氧化菌较少,造成在干流点位的该甲烷氧化菌丰度减少。由于回水顶托现象,干流江水倒灌进支流黄石点位,造成支流黄石点位的该甲烷氧化菌丰度略微减少,从而对甲烷通量造成一定影响。
Ⅰ型好氧甲烷氧化菌对整个湖泊和水柱层的甲烷氧化活动发挥主导作用,对甲烷的循环有着非常显著的影响[43]。在美国康斯坦茨湖[44]以及我国湿地[45]的相关研究均证明,Ⅰ型甲烷氧化菌主导着我国湖泊沉积物中的甲烷氧化过程。有学者在帕文湖缺氧层中检测到了大量的Methylobacter属好氧甲烷氧化菌[46]。OSWALD[47]等运用放射性示踪实验揭示了甲烷氧化的光诱导作用下,看似缺氧的水层存在溶解氧,并且在同一区域发现了Methylobacter属的好氧甲烷氧化菌[48]。有研究者[49]对部分湖泊水样进行功能基因测序,发现Ⅰ型好氧甲烷氧化菌中的Methylobacter属占主导地位。本研究中样本点水华爆发严重,水体底部出现厌氧情况,溶解氧处于低浓度状态,导致Methylobacter属活性增大,从而加速了甲烷氧化,该结论在秦宇[43]的研究中也有类似结果。
3. 结论
1)监测期间内,三峡库区高阳、黄石、万州3个观测点平均甲烷通量为(0.874±0.011)、(0.884±0.234)、(0.507±0.262) μmol·(m2·h)−1,总体表现为甲烷排放源。干、支流甲烷通量表现为支流黄石>支流高阳>干流万州。产生此结果的原因是,温度与河流回水顶托作用导致了干、支流甲烷排放的差异。
2)在产甲烷菌群落中,部分未分类的产甲烷古菌unclassified_p_Euryarchaeota、environmental_samples_f_ Methanosarcinaceae以及未命名产甲烷菌对甲烷排放影响较大。当这类物种丰度增加时,可促进甲烷的产生并影响甲烷排放。
3)在甲烷氧化菌群落中,Methylobacter、Methylosarcina以及未分类甲烷氧化菌对甲烷排放影响较大。当该类细菌丰度增加时会加速甲烷氧化,从而减少水-气界面的甲烷排放。
-
图 2 三峡库区万州段采样点位流域图
Figure 2. Location map of sampling sites in Wanzhou section of the Three Gorges Reservoir area
图 4 产甲烷菌群落与甲烷排放通量的冗余分析
Figure 4. Redundant analysis of methanogenic bacterial community and methane emission flux
图 5 甲烷氧化菌群落与甲烷排放通量的冗余分析
Figure 5. Redundant analysis of methane oxidizing bacteria community and methane emission flux
表 1 样本信息表
Table 1. Sample information table
采样时间 样本名称 采样时间及地点 2019年7月 WZ1907 2019年7月万州泥样 GY1907 2019年7月高阳泥样 HS1907 2019年7月黄石泥样 2019年9月 WZ1909 2019年9月万州泥样 GY1909 2019年9月高阳泥样 HS1909 2019年9月黄石泥样
下载: 导出CSV
表 2 高通量测序引物序列表[21]
Table 2. High-throughput sequencing primer sequence list
目的基因 引物 引物序列 (5’-3’) 16Sr RNA (细菌) 515FmodF GTGCCAGCMGCCGCGG 806RmodR GGACTAVHVGGGTWTCTAAT pmoA (好氧甲烷氧化菌) A189F GGNGACTGGGACTTCTGG Mb661R GGTAARGACGTTGCNCCGG mcrA (产甲烷菌) MLf GCCCGGTGGTGTMGGATTCACACARTAYGCWACAGC MLr TTCATTGCRTAGTTWGGRTAGTT
下载: 导出CSV
表 3 产甲烷菌群落与甲烷排放通量相关性分析
Table 3. Correlation analysis between methanogenic bacterial communities and methane emission flux
产甲烷菌群落 (属) P 值 相关系数 r unclassified_p__Euryarchaeota 0.174 0.636 norank_c__environmental_samples 0.122 0.700 environmental_samples_f__Methanos-arcinaceae 0.514 0.337 unclassified_k__norank_d__Archaea 0.481 −0.362 unclassified_o__Methanomicrobiales 0.276 −0.533 Methanoregula 0.311 0.501 unclassified_f__Methanoregulaceae 0.715 0.192 Methanobacterium 0.115 0.709 unclassified_f__Methanomassiliicocc-aceae 0.073 −0.771 unclassified_c__Thermoplasmata 0.563 −0.300
下载: 导出CSV
表 4 甲烷氧化菌群落与甲烷排放通量相关性分析
Table 4. Correlation analysis between methane oxidizing bacteria community and methane emission flux
甲烷氧化菌群落 (属) P 值 相关系数 r Methylocystis 0.141 0.651 unclassified_c__Gammaproteobacteria 0.763 −0.159 Methylosarcina 0.033* −0.847 Methylobacter 0.031* −0.853 norank_o__environmental_samples 0.581 0.287 unclassified_f__Methylocystaceae 0.214 0.593 environmental_samples_f__Methylococ-caceae 0.080 -0.759 unclassified_k__norank_d__Bacteria 0.275 0.534 Methylomonas 0.333 0.482 norank_p__environmental_samples 0.110 −0.715 注:“*”表示P<0.05,显著相关。
下载: 导出CSV
-
[1] OCKO I B, SUN T Y, SHINDELL D, et al. Acting rapidly to deploy readily available methane mitigation measures by sector can immediately slow global warming[J]. Environmental Research Letters, 2021, 16(5): 054042. doi: 10.1088/1748-9326/abf9c8 [2] SHEN L, LIU S, ZHU Q, et al. Distribution and diversity of nitrite-dependent anaerobic methane-oxidising bacteria in the sediments of the Qiantang River[J]. Environmental Microbiology, 2014, 67(2): 341-349. [3] 刘洋, 陈永娟, 王晓燕, 等. 水库与河流沉积物中好氧甲烷氧化菌群落差异性研究[J]. 中国环境科学, 2018, 38(5): 1844-1854. doi: 10.3969/j.issn.1000-6923.2018.05.030 [4] KIRSCHKE S, BOUSQUET P, CIAIS P, et al. Three decades of global methane sources and sinks[J]. Nature Geoscience, 2013, 6(10): 813-823. doi: 10.1038/ngeo1955 [5] ROLAND F A, DARCHAMBEAU F, MORANA C, et al. Emission and oxidation of methane in a meromictic, eutrophic and temperate lake (Dendre, Belgium)[J]. Chemosphere, 2017, 168: 756-764. doi: 10.1016/j.chemosphere.2016.10.138 [6] 孙志禹, 陈永柏, 李翀, 等. 中国水库温室气体研究(2009—2019): 回顾与展望[J]. 水利学报, 2020, 51(3): 253-267. [7] GUÉRIN F, ABRIL G, RICHARD S, et al. Methane and carbon dioxide emissions from tropical reservoirs: Significance of downstream rivers[J]. Geophysical Research Letters, 2006, 33(21): 493-495. [8] ROSA L P, SANTOS M A D, MATVIENKO B, et al. Scientific errors in the Fearnside comments on greenhouse gas emissions (GHG) from hydroelectric dams and response to his political claiming[J]. Climatic Change, 2006, 75(1/2): 91-102. [9] 秦宇, 张渝阳, 李哲, 等. 三峡澎溪河水华期间水体CH4浓度及其通量变化特征初探[J]. 环境科学, 2018, 39(4): 1578-1588. [10] 唐千, 薛校风, 王惠, 等. 湖泊生态系统产甲烷与甲烷氧化微生物研究进展[J]. 湖泊科学, 2018, 30(3): 597-610. doi: 10.18307/2018.0302 [11] LEI D, LIU J, ZHANG J W. Methane oxidation in the water column of Xiangxi Bay, Three Gorges Reservoir[J]. Clean—Soil, Air, Water, 2019, 47(9): 1-9. [12] 卓海华, 吴云丽, 刘旻璇, 等. 三峡水库水质变化趋势研究[J]. 长江流域资源与环境, 2017, 26(6): 925-936. doi: 10.11870/cjlyzyyhj201706015 [13] 李哲, 姚骁, 何萍, 等. 三峡水库澎溪河水-气界面CO2、CH4扩散通量昼夜动态初探[J]. 湖泊科学, 2014, 26(4): 576-584. doi: 10.18307/2014.0412 [14] 李哲, 张呈, 刘靓, 等. 三峡水库澎溪河CO2、CH4气泡释放通量初探[J]. 湖泊科学, 2014, 26(5): 789-798. doi: 10.18307/2014.0518 [15] 秦宇, 杨博逍, 李哲, 等. 夏季金沙江下游水-气界面CO2、CH4通量特征初探[J]. 湖泊科学, 2017, 29(4): 991-999. [16] B. JÄHNE, P. LIBNER, R. FISCHER, et al. Investigating the transfer processes across the free aqueous viscous boundary layer by the controlled flux method[J]. Tellus B, 1989, 41B(2): 177-195. doi: 10.1111/j.1600-0889.1989.tb00135.x [17] MACINTYRE S, WANNINKHOF R, CHANTON J P. Trace gas exchange across the air-water interface in freshwater and coastal marine environments[J]. New York:P. A. Matson and Harriss Blackwell Science, 1995: 52-97. [18] MOREL, F M M. Principles of aquatic chemistry[J]. Limnology and Oceanography, 1985, 30(2): 450-450. doi: 10.4319/lo.1985.30.2.0450 [19] ROEHM C L, PRAIRIE Y T, DEL GIORGIO P A. The p(CO2)dynamics in lakes in the boreal region of northern Québec, Canada[J]. Global Biogeochemical Cycles, 2009, 23(3): 7-10. [20] ANDERSEN C B. Understanding carbonate equilibria by measuring alkalinity in experimental and natural systems[J]. Journal of Geoscience Education, 2002, 50(4): 389-403. doi: 10.5408/1089-9995-50.4.389 [21] 张曦. 三峡库区万州段沉积物细菌群落对CH4排放的影响[D]. 重庆: 重庆交通大学, 2020. [22] 秦宇, 王紫薇, 李哲, 等. 三峡水库澎溪河水-气界面CO2与CH4通量特征及影响因素初探[J]. 地球环境学报, 2019, 10(2): 177-189. [23] 赵登忠, 谭德宝, 汪朝辉, 等. 清江流域水布垭水库温室气体交换通量监测与分析研究[J]. 长江科学院院报, 2011, 28(10): 197-204. doi: 10.3969/j.issn.1001-5485.2011.10.035 [24] 谌佳伟, 葛继稳, 冯亮, 等. 神农架大九湖泥炭湿地甲烷通量特征及其与土壤微生物群落组成的关系[J]. 地球科学, 2020, 45(3): 1082-1092. [25] 赵登忠, 程学军, 汪朝辉, 等. 清江流域典型发电水库甲烷源汇时空变化规律研究[J]. 水力发电学报, 2014, 33(5): 128-137. [26] 赵炎, 曾源, 吴炳方, 等. 三峡水库香溪河支流水域温室气体排放通量观测[J]. 水科学进展, 2011, 22(4): 546-553. [27] 吴学谦, 操满, 傅家楠, 等. 三峡水库夏季干流、支流(草堂河)水体的二氧化碳分压及扩散通量[J]. 上海大学学报(自然科学版), 2015, 21(3): 311-318. [28] 李哲, 方芳, 郭劲松, 等. 三峡小江(澎溪河)藻类功能分组及其季节演替特点[J]. 环境科学, 2011, 32(2): 392-400. [29] 赵玮, 朱孔贤, 黄文敏, 等. 三峡水库湖北段甲烷排放通量时空特征及其影响因素分析[J]. 水生生物学报, 2013, 37(4): 776-781. doi: 10.7541/2013.95 [30] WATANABE T, KIMURA M, ASAKAWA S. Distinct members of a stable methanogenic archaeal community transcribe mcrA genes under flooded and drained conditions in Japanese paddy field soil[J]. Soil Biology and Biochemistry, 2008, 40(1): 276-285. [31] IMACHI H, AOI K, TASUMI E, et al. Cultivation of methanogenic community from subseafloor sediments using a continuous-flow bioreactor[J]. The ISME Journal:Multidisciplinary Journal of Microbial Ecology, 2011, 5(12): 1913-1925. [32] 李思琦, 臧昆鹏, 宋伦. 湿地甲烷代谢微生物产甲烷菌和甲烷氧化菌的研究进展[J]. 海洋环境科学, 2020, 39(3): 488-496. doi: 10.12111/j.mes20200325 [33] LIU Y, WANG P, CROWLEY D, et al. Methanogenic abundance and changes in community structure along a rice soil chronosequence from east China[J]. European Journal of Soil Science, 2016, 67(4): 443-455. doi: 10.1111/ejss.12348 [34] CONRAD R, ERKEL C, LIESACK W. Rice Cluster I methanogens, an important group of Archaea producing greenhouse gas in soil[J]. Current Opinion in Biotechnology, 2006, 17(3): 262-267. doi: 10.1016/j.copbio.2006.04.002 [35] ZHUANG G C, HEUER V B, LAZAR C S, et al. Relative importance of methylotrophic methanogenesis in sediments of the Western Mediterranean Sea[J]. Geochimica Et Cosmochimica Acta, 2018, 224: 171-186. doi: 10.1016/j.gca.2017.12.024 [36] 沈李东. 湿地亚硝酸盐型厌氧氨氧化和厌氧甲烷氧化微生物生态学研究[D]. 浙江: 浙江大学, 2014. [37] MER J L, ROGER P. Production, oxidation, emission and consumption of methane by soils: A review[J]. European Journal of Soil Biology, 2001, 37(1): 25-50. doi: 10.1016/S1164-5563(01)01067-6 [38] 王晓琳, 曹爱新, 周传斌, 等. 垃圾填埋场甲烷氧化菌及甲烷减排的研究进展[J]. 生物技术通报, 2016, 32(5): 16-25. [39] 贠娟莉, 王艳芬, 张洪勋. 好氧甲烷氧化菌生态学研究进展[J]. 生态学报, 2013, 33(21): 6774-6785. [40] 马若潺, 魏晓梦, 何若. 低氧生境中好氧甲烷氧化菌的缺氧耐受机理及种群结构研究进展[J]. 应用生态学报, 2017, 28(6): 2047-2054. [41] YANG Y Y, CHEN J F, TONG T L, et al. Eutrophication influences methanotrophic activity, abundance and community structure in freshwater lakes[J]. Science of the Total Environment, 2019, 662: 863-872. doi: 10.1016/j.scitotenv.2019.01.307 [42] OSWALD K, MILUCKA J, BRAND A, et al. Aerobic gammaproteobacterial methanotrophs mitigate methane emissions from oxic and anoxic lake waters[J]. Limnology and Oceanography, 2016, 61(S1): S101-S118. doi: 10.1002/lno.10312 [43] 秦宇, 黄璜, 李哲, 等. 内陆水体好氧甲烷氧化过程研究进展[J]. 湖泊科学, 2021, 33(4): 1004-1017. [44] YUN J L, ZHUANG G Q, MA A Z, et al. Community structure, abundance, and activity of methanotrophs in the Zoige wetland of the Tibetan Plateau.[J]. Microbial ecology, 2012, 63(4): 835-843. doi: 10.1007/s00248-011-9981-x [45] LIU Y, ZHANG J X, ZHAO L, et al. Aerobic and nitrite-dependent methane-oxidizing microorganisms in sediments of freshwater lakes on the Yunnan Plateau[J]. Applied Microbiology and Biotechnology, 2015, 99(5): 2371-2381. doi: 10.1007/s00253-014-6141-5 [46] CORINNE B P, DIDIER J, ERIC D B, et al. Identification of microbial communities involved in the methane cycle of a freshwater meromictic lake[J]. FEMS Microbiology Ecology, 2011, 77(3): 533-545. doi: 10.1111/j.1574-6941.2011.01134.x [47] OSWALD K, MILUCKA J, BRAND B, et al. Light-dependent aerobic methane oxidation reduces methane emissions from seasonally stratified lakes[J]. PLoS ONE, 2017, 10(7). [48] KADNIKOV V V, SAVVICHEV A S, MARDANOV A V, et al. Microbial communities involved in the methane cycle in the near-bottom water layer and sediments of the meromictic subarctic Lake Svetloe[J]. Antonie van Leeuwenhoek, 2019, 112(12): 1801-1814. doi: 10.1007/s10482-019-01308-1 [49] BLEES J, NIEMANN H, WENK C B, et al. Micro-aerobic bacterial methane oxidation in the chemocline and anoxic water column of deep south-Alpine Lake Lugano (Switzerland)[J]. Limnology and Oceanography, 2014, 59(2): 311-324. doi: 10.4319/lo.2014.59.2.0311 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 5687
- HTML全文浏览数: 5687
- PDF下载数: 93
- 施引文献: 0














