













































-
由于我国养殖和种植规模巨大,养殖业产生的畜禽粪污年产量高达3.8×109 t[1],作物秸秆每年产生的数量约为9.0×108 t [2] 。如对它们不进行妥善处理,将会造成水体、大气和土壤的污染,影响农村环境卫生,破坏生态环境,并对人畜健康造成威胁。但是,农业废弃物含有大量养分和有机质资源,如将这些养分以有机肥的形式归还土壤,不仅可有效减少化肥用量和过量化肥施用导致的环境压力,同时也可为“化肥零增长”和“农业可持续发展”等国家战略需求提供支撑[3]。好氧堆肥具有可就地处理、成本低、耗时短、有机物分解较彻底、产品无害化等优势[3],然而该过程会产生一系列的二次污染。例如,堆肥原料中碳氮化合物的转化会产生大量的CH4、CO2和N2O等温室气体,加剧温室效应;NH3大量挥发至大气中,与SO2以及氮氧化物结合形成灰霾,加快PM2.5的形成速度[4];恶臭气体H2S[5]挥发到空气中会严重影响空气质量,危害人群和动物的健康。上述各种气体的产生不仅会导致严重的环境污染,同时也会造成堆肥产品碳氮养分的损失。有研究者指出,由氨挥发造成的堆肥氮素的损失高达50%以上[6]。因此,如何减少堆肥过程中的氨挥发和温室气体排放是目前亟待解决的问题。
目前,研究者多通过调整堆肥物料特性、改良堆肥工艺、优化供气方式和添加外源添加剂[7]等方式实现氨气减排和氮素的保留。其中,外源添加剂是较便捷且高效的手段[8]。常见的外源添加剂主要包括物理添加剂、化学添加剂和微生物添加剂3类。物理添加剂主要包括生物炭、沸石、彭韧土、麦饭石等吸附剂,这类吸附剂具有丰富的孔隙结构和带负电荷的吸附位点,有利于NH3和
NH+4 的吸附[7];化学添加剂有磷酸、磷石膏、凹凸棒土、氢氧化镁和过磷酸钙等,可以通过降低pH和增加NH+4 -N的化学固定实现NH3减排[9];微生物添加剂是由一种或者多种菌种组成的复合菌剂,可以通过加速升温,延长高温期,减少养分的损失,减少温室气体和恶臭氮硫化合物的排放[10]。酸性添加剂一般具有较低pH;在部分添加剂中,其中含有的阳离子如Fe3+和Al3+会在溶液中水解形成酸[9]。酸性添加剂分为无机酸,有机酸和酸性盐,由于其可以通过降低物料pH,造成物料酸化,实现NH3减排,因此日益受到研究者的重视。胡雨彤等[11]发现,在牛粪和锯末堆肥中加入0.2 mol·L−1的硫酸会起到有效的保氮作用。还有研究者[12-13]发现,硫磺粉和磷酸一铵的加入可降低物料pH,从而减少NH+4 -N向氨气的转化。陶勇等[14]将不同比例苹果渣和1%的柠檬酸混合加入猪粪和秸秆中进行好氧堆肥,发现苹果渣中的果酸可以降低堆肥的pH,从而降低氮素损耗;添加10%的苹果渣处理的保氮效果最好,添加5%的苹果渣处理有利于NH+4 -N的积累。但是,由于有机酸容易降解,其氨气减排效果会比无机酸弱[15]。过磷酸钙来源于磷矿石的酸解,是一种含有丰富的磷、钙、硫和微量元素的酸性化学磷肥,在堆肥中有良好的固氮和保氨效果,堆肥腐熟后施入土壤后还会增加土壤磷的生物有效性[16]。糠醛渣是以玉米和秸秆等农副产品的下脚料为原料制取糠醛后残留的固体废弃物[17],具有较高的碳氮比和较低的pH。本实验以牛粪和小麦秸秆为原料,采用户外条垛式发酵工艺,通过添加过磷酸钙和糠醛渣以改变肥料的pH,探究其对NH3和N2O温室气体排放的影响,旨在为实现好氧堆肥过程中有害气体的减排提供依据。
-
堆肥实验在甘肃省白银市某商品化堆肥生产车间进行,堆肥方式采用条垛式,堆肥时间为2020年6月15日至2020年7月31日,共计45 d。该实验区年平均气温约为9 ℃,7—8月平均气温28 ℃,年均降雨量181 mm,全年多风。堆肥原料为牛粪和小麦秸秆,新鲜牛粪由肥料厂提供,小麦秸秆由附近奶牛场提供,堆置前将秸秆切成2~5 cm的碎料。添加剂为过磷酸钙和糠醛渣,供试材料理化性质见表1。
-
堆肥实验采用牛粪和小麦秸秆混合堆肥,各堆体由790 kg牛粪和228 kg小麦秸秆组成,调节其含水率在60%左右,C/N于25左右。共设置3个条跺,其中无添加剂为对照组(CK),添加物料干重10%的过磷酸钙和添加物料干重10%的糠醛渣的堆体分别记为处理SP和FR。每个条跺尺寸设计为长6 m、底部宽1.8 m、高1.2 m,自然堆置。采样时将堆体分为3部分,即每2 m设置1个采样点,每个采样点按离地面高度分为上、中、下3层(分别距离地面约100、60和20 cm)。各条垛之间距离保持在5 m以外,防止物料相互掺杂渗透。
-
本次堆肥期间,每3 d采集1次固体样品,在每个采样点分层取样,每层采集300 g样品,将采集的3层样品充分混匀后分为3份,1份鲜样收集在自封袋后立即密封保存于4 ℃冰箱,用于硝态氮、铵态氮和发芽指数测定。1份进行自然风干并粉碎过1.000 mm筛,用于测定总氮、pH;1份自然风干后粉碎过0.149 mm筛,用于测定有机氮及其组分。每次采样均在翻抛之前进行。
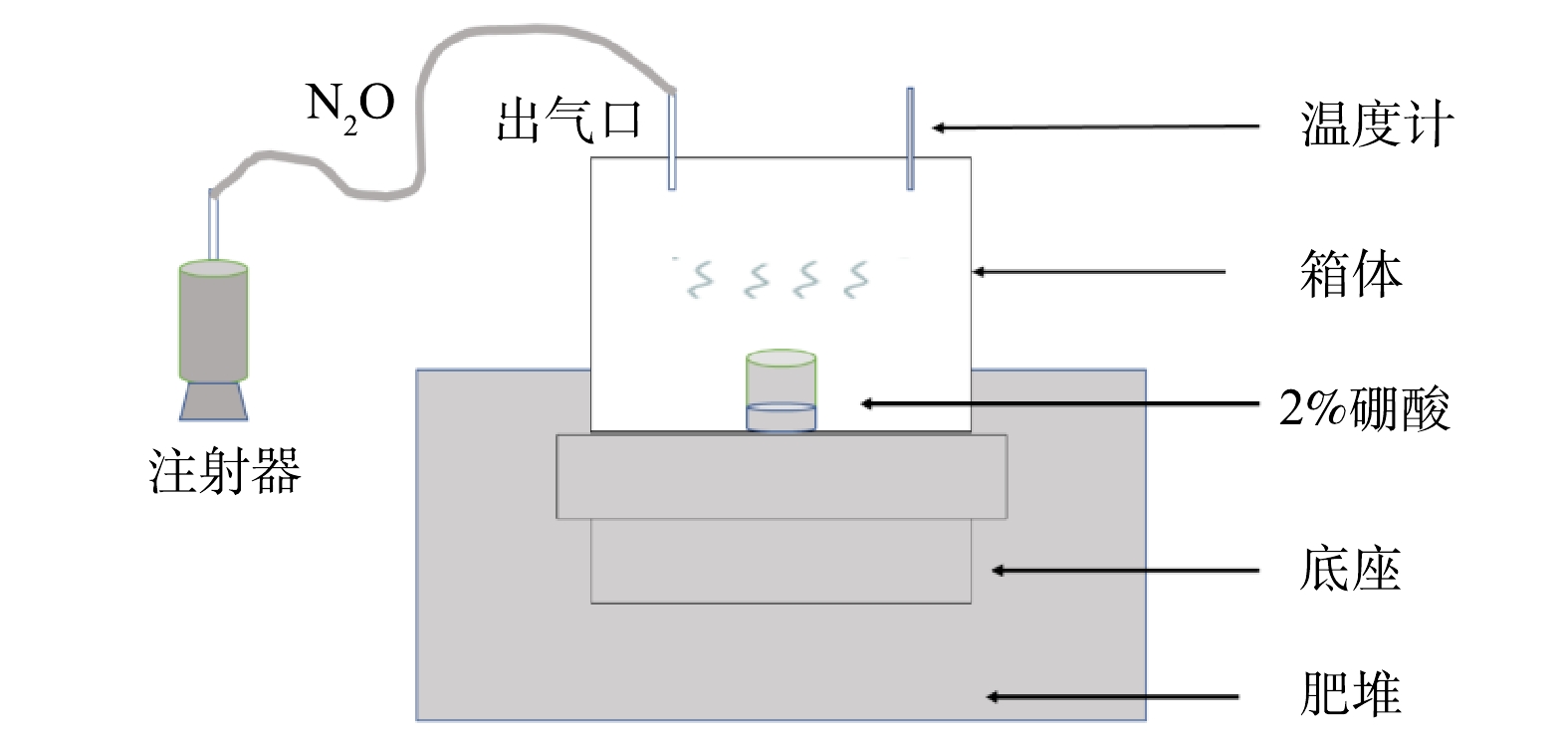
本实验研究的气体主要是氨气和氧化亚氮,采用静态箱法,在翻堆之前收集。静态箱由不锈钢材料制成,分为箱体和底座2部分。箱体为边长50 cm正方体;底座是高为20 cm、底边边长为50 cm,带有宽度约为2 cm、深度约1 cm的水槽,采集气体时将箱体放置在底座上,并在水槽中注水用来密封,以防止外界空气与箱内气体接触。静态箱上方有2个孔,一个用来插入注射器收集气体,另一个用来插入温度计测量箱体内气体温度。每个肥堆放置3个静态箱,以其采集到的气体为重复。
氧化亚氮采集:用连接了三通阀的200 mL注射器每隔5 min吸取1次,注入集气袋中,共计20 min,分别收集采样0、5、10、15、20 min时的气体样。每次采集气体的同时要记录集气箱内气体的温度变化。氨气采集:将盛2%硼酸的烧杯置于静态箱内吸收氨气。吸收结束后,将烧杯用保鲜膜封口,带回室内滴定[18]。图1为气体采集示意图。
-
温度测定:用100 ℃水银温度计在每天8:00、14:00、20:00进行测量,取3个时间段温度的平均值作为当日堆体温度。每个时间段测温层次与取样层次一致,每层均匀测温4次,取其平均值作为当前时刻堆体温度,并记录该时刻堆肥大棚内的环境温度取平均值作为当天的环境温度。
pH采用pH计法测定;将20粒小白菜种子均匀放入垫有2滤纸的培养皿中,加入10 mL浸提液(水肥比10∶1),在培养箱中25 ℃培养48 h,以去离子水为对照,根据种子发芽率和种子根长计算发芽指数;氨气采用2%的硼酸吸收,0.005 mol·L−1的硫酸进行滴定,氨气挥发速率和累积挥发量计算采用参考文献[18]的计算方法,当时间间隔为1 d时,未测定日期的数据通过前后2次测量数据的算术平均值得出,当时间间隔为2 d时,求出这段时间的算术均值,之后乘以这段时间的长度,得到这段时间的累积挥发量;氧化亚氮浓度利用Agilent 7890B气相色谱仪测定,采用微电子捕获器(ECD),工作温度300 ℃,尾吹气为N2,N2流量为20 mL·min−1,氧化亚氮排放通量计算公式参考文献[19],当时间间隔为1 d时,未测定日期的数据通过前后2次测量数据的算术平均值得出,当时间间隔为2 d时,求出这段时间的算术均值,之后乘以这段时间的长度,得到这段时间的累积挥发量;有机氮采用Bremner酸解法[20]测定。
小白菜种子发芽指数(GI)依照式(1)计算; NH3排放速率依照式(2)计算;N2O的排放速率依照式(3)计算。
式中:F表示NH3排放速率,mg·(m2·h)–1;C表示标定过的硫酸浓度,mol·L−1;V和V0(空白)表示滴定消耗的硫酸体积,mL;M表示NH3的分子量,17.03 g·mol−1;L2表示静态箱与肥堆接触面积,m2;T为NH3的吸收时间。
式中:F表示N2O排放速率,mg·(m2·h)−1;ρ为标准状况下N2O密度,1.978 kg·m−3;H表示箱内气体的高度,m;
dcdt 表示箱内气体浓度随时间的变化率,μL·(L·min)−1;T为箱内气体温度,℃。 -
1)温度的变化。如图2(a)所示,在好氧堆肥初期,堆体内部有机物质的分解和微生物的活动会产生大量的热,造成堆体温度的迅速上升。在堆肥第2天,各堆体均上升到了50 ℃以上;在第3天,各处理的温度都达到了55 ℃;在第5天均接近70 ℃。CK、SP和FR在高温期(>55 ℃)的持续时间分别为27、26和34 d,最高温度分别达到了68.9、70.0和68.8 ℃,符合我国畜禽废弃物无害化的标准[21]。27 d后,CK和SP处理温度开始下降至55 ℃以下,而FR处理的温度直到33 d后才开始低于55 ℃。这说明,添加糠醛渣延长了堆肥的高温期,而CK和SP的温度的变化趋势基本一致。
2) pH的变化。如图2(b)所示,随着堆体温度的升高,各处理的pH开始逐渐上升,在第9天达到最大值。这是由于微生物分解含氮化合物使其剧烈矿化,铵态氮大量积累所致[22-23];随着氨气的挥发和酸性物质的形成,pH会相应缓慢下降后逐渐趋于稳定[23]。在整个堆肥进程中,添加过磷酸钙可以明显降低堆体的pH;而FR处理的pH除第1天略低于CK外,在堆肥其他时间始终高于CK。这说明,添加糠醛渣后可提高堆体的pH,但提高的幅度并不大。
3)发芽指数。种子发芽指数(GI)是检验有机肥有无毒害的重要指标。当GI值大于50%时,可认为堆肥基本腐熟,当其大于80%时即可认为堆肥完全腐熟。图2(c)是本次堆肥中小白菜种子的发芽指数变化情况。堆肥刚开始时,由于肥料未腐熟,各处理GI值较低;从第18天开始,SP和FR处理的GI值大于50%;至第36天,SP处理GI值达到了90%;至堆肥结束,各处理GI值则分别达到了94.23%、110.36%和104.56%,这说明肥料已无毒害,可能是添加剂中携带的营养物质促进了种子的萌发。
-
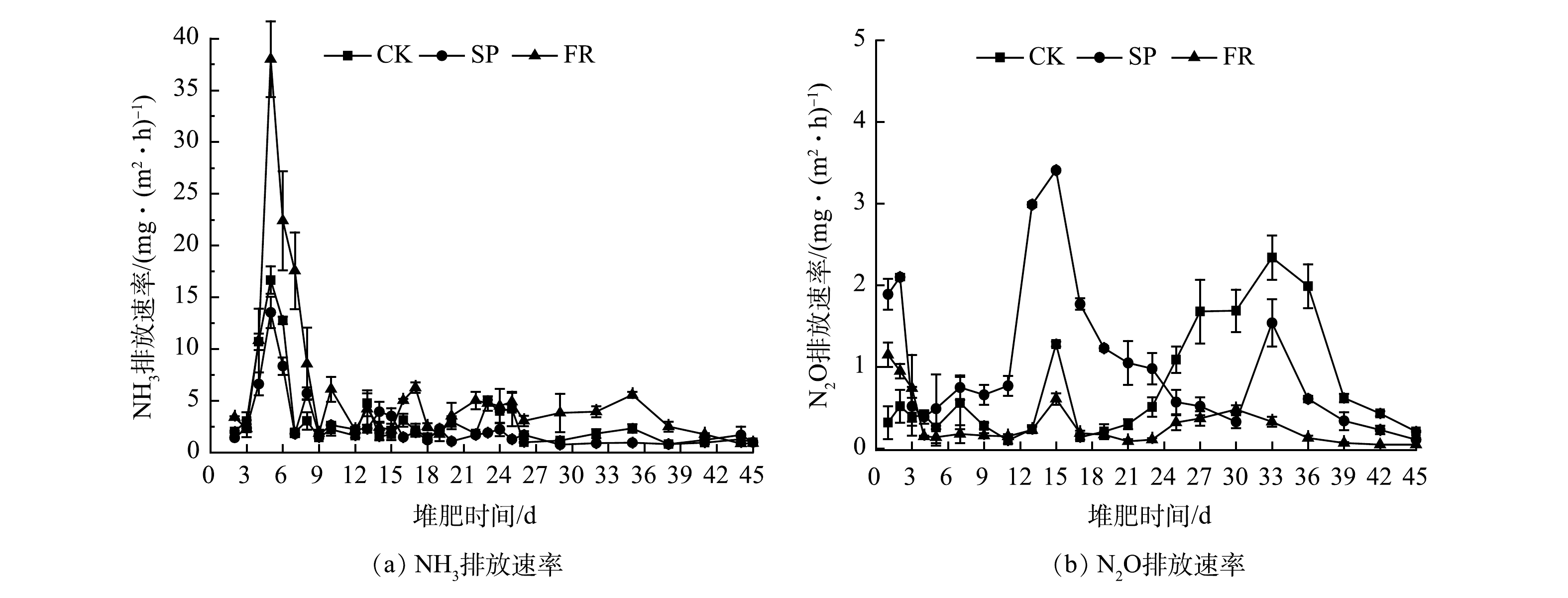
1)氨气排放速率。如图3(a)所示,各处理下氨气的大量挥发都主要集中在堆肥前9天,这与pH和温度的变化有关。堆肥初期的氨化细菌类代谢能力强,分解含氮有机物时可产生的大量
NH+4 -N[24],导致pH升高,这是氨挥发的直接原因[18]。随着堆体温度的升高,NH3的溶解度随之降低[25],NH3在水相中由NH3·H2O的形态转化为气态挥发到大气中,且温度越高挥发量越大[26]。与CK相比,SP处理抑制了氨气的排放速率,尤其是明显降低了前6天的氨挥发,最高降幅为34.87%;FR处理则增加了氨气的排放速率,尤其是0~9天,最大增幅高达1.28倍。然而,温度上升与氨排放并不呈正相关。这是因为,超高温堆肥过程中的高温会通过抑制蛋白酶和脲酶的活性、降低氨化细菌丰度来减少氨气的排放;同时,其还可以通过加速腐殖化进程,促进含氮腐殖物质的形成,从而减少氮素的流失[27]。2)氧化亚氮排放速率。如图3(b)所示,N2O主要来源于
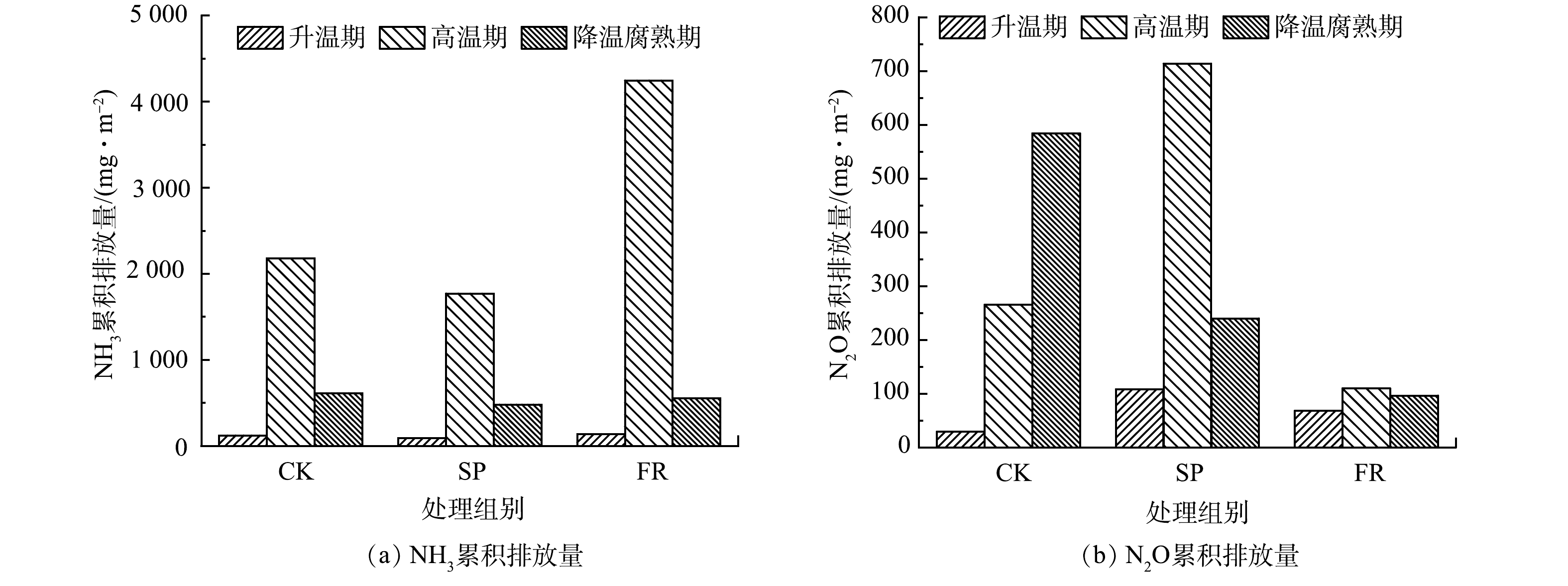
NH+4 -N的不完全硝化、厌氧条件下NO−x -N的硝化和硝酸盐的反硝化[28],反硝化作用对N2O的排放占据主导地位[29]。反硝化细菌的活性与反应速率和NO−3 -N含量呈正相关,在堆肥降温期数量最多[30],所以该时期会出现N2O排放高峰。SP处理明显增加了升温期和高温期的N2O排放速率,这与杨岩等[19]、罗一鸣等[31]的研究结果相似,这可能与该时期大量NH+4 -N的不完全硝化有关。此外,耐高温甲烷氧化菌发生氨氧化反应[29]和硝化作用[32]也会促使堆肥高温期产生N2O气体,说明添加过磷酸钙可能有利于这些特异微生物的生存。随着堆体温度的降低和NO−3 -N的积累,反硝化作用开始进行,SP和FR处理下N2O的排放速率与CK相比有所下降,这可能是因为过磷酸钙和糠醛渣的加入抑制了NO−3 -N的产生。另外,还有研究表明,堆肥过程中N2O的排放与pH呈负相关关系[29],即,在微酸性环境下会产生更多的N2O。这或许又可以作为解释SP处理N2O高于CK的原因之一,FR处理由于有较高的pH,故产生的N2O排放量较低。3)氨气累积排放量。通常,根据堆体温度的变化可以将好氧堆肥过程分为升温期、高温期、降温期和腐熟期,不同时期氨气排放速率的不同决定了该时期累积排放量的不同。如图4(a)所示,CK、SP和FR在升温期和高温期的氨气累积挥发量分别占整个堆肥期间氨气挥发总量的78.8%、79.3%和88.6%。由于CK处理与SP处理之间温度的差异不大,所以是因SP处理下较低的pH抑制了
NH+4 -N向NH3转化,从而导致NH3的总累计挥发量减少了19.65%;与之相反,FR处理因其具有较高的温度和pH,从而导致NH3的总累计挥发量增加了69.59%。4)氧化亚氮累积排放量。如图4(b)所示,与氨气累积排放量同理,SP处理下,因其较高的N2O排放速率,造成升温期和高温累积排放量分别提高了2.72倍和1.68倍,随着降温期和腐熟期N2O排放速率降低,累积排放量下降了58.95%;FR处理下的N2O累积排放量在仅在升温期提高了1.3倍,其他时期N2O累积排放量均有所下降。至堆肥结束,SP处理下N2O总累积排放量增加了20.8%,FR处理下N2O总累积排放量减少了68.79%。
-
1)总氮的变化。堆肥总氮(TN)的变化情况如图5(a)所示。在堆肥的0~6 d,各处理TN质量分数逐渐上升到最大值,这可能是由堆体内含碳有机物的降解率高于氨气的排放率所致[33] 。之后,由于氨气挥发等氮素损失,各处理下TN呈下降趋势;经过高温期的降解和反应,肥堆的体积和质量不断下降,当下降速率远超过氮素减少的速率时就会产生浓缩效应,使TN质量分数开始上升。至堆肥结束,各处理下TN质量分数与堆肥初期相比分别增加了 9.29%、5.44%和14.40%。在整个堆肥过程中,SP处理的TN质量分数始终低于CK,但FR处理的TN却始终高于CK。
2)酸解有机氮的变化。氮素由有机氮和无机氮组成,有机氮是主要的氮组分,占总氮的90%以上,酸解总氮(THN)占有机氮的50%以上[19]。如图5(b)所示,由于微生物的代谢活动,有机氮被分解,CK、SP和FR下的酸解总氮持续下降,降幅分别为13.87%、7.98%和5.53%;添加剂的加入减缓了有机氮的降解,这可能是由于微生物优先分解添加剂携带的营养物质。随着氨氧化微生物和其他氮同化微生物活性的恢复,有机氮的合成再次启动[26],THN质量分数开始上升;另外,浓缩效应也会造成THN呈上升趋势。至堆肥结束,CK、SP和FR处理的THN与堆肥初期相比增加了5.08%、7.85%和9.07%。在整个堆肥过程中,SP处理中THN质量分数始终低于CK;FR处理在前15 d低于CK,这可能是由前期其氨气排放过高所致。通过对CK、SP和FR处理的THN和
NH+4 -N进行Pearson相关分析,得到三者相关系数分别为−0.768、−0.639和−0.733,显著性水平分别为0.001、0.008和0.001,这说明THN与NH+4 -N之间存在显著的负相关(P<0.01)。3)铵态氮的变化。如图5(c)所示,各处理
NH+4 -N变化趋势基本一致。在堆肥前6 d,原料中含氮有机物大量分解,导致各处理NH+4 -N的不断累积升高并在第6天达到最大值;随着氨气的排放和硝化作用的加强,以及NH+4 -N参与堆体内含氮有机物的合成,NH+4 -N质量分数不断下降。添加过磷酸钙后,NH+4 -N质量分数提高了40%~80%。一方面,这是因为过磷酸钙的主要成分是Ca(H2PO4)2和CaSO4· 12H2O [5,16],磷酸根离子可以让部分NH+4 与堆肥物料中的众多阳离子(如Ca2+)结合,形在结晶和复合体留在肥料中[36];另一方面,可能是因为较低的pH抑制了其硝化作用。添加糠醛渣则降低了堆体中NH+4 -N质量分数,这可能与较高氨气排放量和硝化作用消耗较多NH+4 -N有关。4)硝态氮的变化。
NO−3 -N的变化如图5(d)所示。堆肥前24 d,高浓度的氨积累和高温抑制了硝化反应[28],各处理下硝态氮基本稳定在11~14 mg·kg −1。随着堆体温度的下降,硝化反应开始并渐趋剧烈。硝化过程是NH+4 在硝化细菌作用下转化为硝酸盐的过程,该过程分为两步,首先是NH+4 在氨氧化细菌(AOB)的氧化作用下氧化为NO−2 ,然后被亚硝酸盐氧化细菌(NOB)硝化为NO−3 [34]。当温度降低后,pH开始影响硝化细菌的活性。这是因为,AOB和NOB的最适pH分别为7.0~8.5和6.0~7.5[35]。当pH小于6.5时,不利于AOB的生存,硝化速率下降;当pH大于8.0时,有利于AOB的生长,氨氧化速率上升。所以,pH的升高有利于提高NH+4 -N的氧化[34]。当堆肥进入降温期后,CK、SP和FR处理的pH的范围分别为8.0、6.5和8.5上下,此时CK和FR中的氨氧化反应剧烈,增加了硝化反应产生的NO−3 -N。而SP处理则由于较低的pH限制了氨氧化菌(AOB) 的活性,减弱了氨氧化反应,从而抑制硝化过程,NO−3 -N质量分数较低。 -
1)3个肥堆的高温期保持时间均满足好氧堆肥的腐熟要求;添加糠醛渣使高温期延长了7 d,添加过磷酸钙则降低了肥料的pH;发芽指数计算结果表明,添加剂的加入不会对植物生长产生毒性。
2)添加过磷酸钙后,氨气的排放速率和排放量均有所下降,但在高温期会产生大量的氧化亚氮,导致氧化亚氮的排放量上升;添加糠醛渣后则可延长堆体的高温期时长,并略微提高肥料的pH,这造成氨气排放较高,但其对氧化亚氮的减排效果明显。
3)添加过磷酸钙后可促进有机氮向铵态氮的转化,并抑制铵态氮向硝态氮的转化;添加糠醛渣可提高堆体总氮和有机氮含量,并促进铵态氮向硝态氮的转化。
添加过磷酸钙和糠醛渣对好氧堆肥过程中氨气排放和氮素转化的影响
Effects of superphosphate and furfural residue addition on NH3 emissions and nitrogen conversion during the aerobic composting
-
摘要: 针对好氧堆肥过程中产生的二次污染问题,以牛粪和小麦秸秆为原料,探究了添加过磷酸钙(SP)和糠醛渣(FR)对户外条垛式好氧堆肥过程中氨气排放和氮素转化的影响。结果表明,添加过磷酸钙降低了堆体的pH,减少了19.65%的氨气排放量,但是其N2O排放量增加了20.8%;添加糠醛渣后堆肥的高温期延长了7 d,增加了69.59%的氨气排放量,但是其N2O排放量减少了68.79%。添加过磷酸钙后,
NH+4 -N增加了40%~80%,这可能是在升温期和高温期该处理的N2O排放高于对照的主要原因;酸解总氮降低了2.21%~13.71%,这说明添加过磷酸钙促进了有机态氮向无机氮的转化。添加糠醛渣降低了NH+4 -N质量分数,增加了总氮的质量分数。添加过磷酸钙通过降低pH减少NH3排放;添加糠醛渣有利于提高总氮质量分数和减少N2O的排放。本研究结果可为好氧堆肥中氨气和氧化亚氮的减排提供参考。Abstract: Aiming at the secondary pollution caused by the aerobic composting process, this experiment used cow dung and wheat straw as raw materials to study the effects of adding superphosphate (SP) and furfural residue (FR) on ammonia emissions and nitrogen conversion in the outdoor aerobic composting process. The results showed that the addition of SP reduced the pH of the pile and reduced the NH3 emissions by 19.65%, but the N2O emissions increased by 20.8%. The high temperature period of the compost after adding FR was prolonged by 7 days and increased the NH3 emissions by 69.59%, but its N2O emissions decreased by 68.79%. After adding FR, theNH+4 -N content increased by 40%~80%, which might be the main reason why the N2O emission of this treatment was higher than that of the control group during the heating period and the high temperature period. The content of total acidolysis nitrogen decreased by 2.21%~13.71%, indicating that the addition of SP promoted the conversion of organic nitrogen to inorganic nitrogen. However, the FR treatment reduced the mass fraction ofNH+4 -N and increased the mass fraction of total nitrogen. In summary, the addition of SP reduces the NH3 emission by lowering the pH; the addition of FR was beneficial to increase the mass fraction of total nitrogen and reduced the emission of N2O. This experiment can provide the theoretical basis for reducing NH3 and N2O emission during the aerobic composting.-
Key words:
- aerobic composting /
- superphosphate /
- furfural residue /
- ammonia emissions /
- nitrous oxide emissions /
- nitrogen
-
由于我国养殖和种植规模巨大,养殖业产生的畜禽粪污年产量高达3.8×109 t[1],作物秸秆每年产生的数量约为9.0×108 t [2] 。如对它们不进行妥善处理,将会造成水体、大气和土壤的污染,影响农村环境卫生,破坏生态环境,并对人畜健康造成威胁。但是,农业废弃物含有大量养分和有机质资源,如将这些养分以有机肥的形式归还土壤,不仅可有效减少化肥用量和过量化肥施用导致的环境压力,同时也可为“化肥零增长”和“农业可持续发展”等国家战略需求提供支撑[3]。好氧堆肥具有可就地处理、成本低、耗时短、有机物分解较彻底、产品无害化等优势[3],然而该过程会产生一系列的二次污染。例如,堆肥原料中碳氮化合物的转化会产生大量的CH4、CO2和N2O等温室气体,加剧温室效应;NH3大量挥发至大气中,与SO2以及氮氧化物结合形成灰霾,加快PM2.5的形成速度[4];恶臭气体H2S[5]挥发到空气中会严重影响空气质量,危害人群和动物的健康。上述各种气体的产生不仅会导致严重的环境污染,同时也会造成堆肥产品碳氮养分的损失。有研究者指出,由氨挥发造成的堆肥氮素的损失高达50%以上[6]。因此,如何减少堆肥过程中的氨挥发和温室气体排放是目前亟待解决的问题。
目前,研究者多通过调整堆肥物料特性、改良堆肥工艺、优化供气方式和添加外源添加剂[7]等方式实现氨气减排和氮素的保留。其中,外源添加剂是较便捷且高效的手段[8]。常见的外源添加剂主要包括物理添加剂、化学添加剂和微生物添加剂3类。物理添加剂主要包括生物炭、沸石、彭韧土、麦饭石等吸附剂,这类吸附剂具有丰富的孔隙结构和带负电荷的吸附位点,有利于NH3和
NH+4 NH+4 NH+4 NH+4 过磷酸钙来源于磷矿石的酸解,是一种含有丰富的磷、钙、硫和微量元素的酸性化学磷肥,在堆肥中有良好的固氮和保氨效果,堆肥腐熟后施入土壤后还会增加土壤磷的生物有效性[16]。糠醛渣是以玉米和秸秆等农副产品的下脚料为原料制取糠醛后残留的固体废弃物[17],具有较高的碳氮比和较低的pH。本实验以牛粪和小麦秸秆为原料,采用户外条垛式发酵工艺,通过添加过磷酸钙和糠醛渣以改变肥料的pH,探究其对NH3和N2O温室气体排放的影响,旨在为实现好氧堆肥过程中有害气体的减排提供依据。
1. 材料与方法
1.1 实验原料
堆肥实验在甘肃省白银市某商品化堆肥生产车间进行,堆肥方式采用条垛式,堆肥时间为2020年6月15日至2020年7月31日,共计45 d。该实验区年平均气温约为9 ℃,7—8月平均气温28 ℃,年均降雨量181 mm,全年多风。堆肥原料为牛粪和小麦秸秆,新鲜牛粪由肥料厂提供,小麦秸秆由附近奶牛场提供,堆置前将秸秆切成2~5 cm的碎料。添加剂为过磷酸钙和糠醛渣,供试材料理化性质见表1。
表 1 堆肥材料理化性质Table 1. Physical and chemical properties of composting materials供试材料 含水率/% pH TC/% TN/% TP/% TK/% C/N 小麦秸秆 7.1 7.3 80.7 1.1 0.2 1.1 76.9 牛粪 45.7 7.6 38.2 2.2 0.9 2.0 17.7 过磷酸钙 0.9 3.5 — — — — — 糠醛渣 10.1 2.91 81.35 0.97 0.87 2.03 81.35 | Show Table DownLoad:
CSV
DownLoad:
CSV
1.2 实验设计
堆肥实验采用牛粪和小麦秸秆混合堆肥,各堆体由790 kg牛粪和228 kg小麦秸秆组成,调节其含水率在60%左右,C/N于25左右。共设置3个条跺,其中无添加剂为对照组(CK),添加物料干重10%的过磷酸钙和添加物料干重10%的糠醛渣的堆体分别记为处理SP和FR。每个条跺尺寸设计为长6 m、底部宽1.8 m、高1.2 m,自然堆置。采样时将堆体分为3部分,即每2 m设置1个采样点,每个采样点按离地面高度分为上、中、下3层(分别距离地面约100、60和20 cm)。各条垛之间距离保持在5 m以外,防止物料相互掺杂渗透。
1.3 实验样品采集
本次堆肥期间,每3 d采集1次固体样品,在每个采样点分层取样,每层采集300 g样品,将采集的3层样品充分混匀后分为3份,1份鲜样收集在自封袋后立即密封保存于4 ℃冰箱,用于硝态氮、铵态氮和发芽指数测定。1份进行自然风干并粉碎过1.000 mm筛,用于测定总氮、pH;1份自然风干后粉碎过0.149 mm筛,用于测定有机氮及其组分。每次采样均在翻抛之前进行。
本实验研究的气体主要是氨气和氧化亚氮,采用静态箱法,在翻堆之前收集。静态箱由不锈钢材料制成,分为箱体和底座2部分。箱体为边长50 cm正方体;底座是高为20 cm、底边边长为50 cm,带有宽度约为2 cm、深度约1 cm的水槽,采集气体时将箱体放置在底座上,并在水槽中注水用来密封,以防止外界空气与箱内气体接触。静态箱上方有2个孔,一个用来插入注射器收集气体,另一个用来插入温度计测量箱体内气体温度。每个肥堆放置3个静态箱,以其采集到的气体为重复。
氧化亚氮采集:用连接了三通阀的200 mL注射器每隔5 min吸取1次,注入集气袋中,共计20 min,分别收集采样0、5、10、15、20 min时的气体样。每次采集气体的同时要记录集气箱内气体的温度变化。氨气采集:将盛2%硼酸的烧杯置于静态箱内吸收氨气。吸收结束后,将烧杯用保鲜膜封口,带回室内滴定[18]。图1为气体采集示意图。
1.4 分析方法
温度测定:用100 ℃水银温度计在每天8:00、14:00、20:00进行测量,取3个时间段温度的平均值作为当日堆体温度。每个时间段测温层次与取样层次一致,每层均匀测温4次,取其平均值作为当前时刻堆体温度,并记录该时刻堆肥大棚内的环境温度取平均值作为当天的环境温度。
pH采用pH计法测定;将20粒小白菜种子均匀放入垫有2滤纸的培养皿中,加入10 mL浸提液(水肥比10∶1),在培养箱中25 ℃培养48 h,以去离子水为对照,根据种子发芽率和种子根长计算发芽指数;氨气采用2%的硼酸吸收,0.005 mol·L−1的硫酸进行滴定,氨气挥发速率和累积挥发量计算采用参考文献[18]的计算方法,当时间间隔为1 d时,未测定日期的数据通过前后2次测量数据的算术平均值得出,当时间间隔为2 d时,求出这段时间的算术均值,之后乘以这段时间的长度,得到这段时间的累积挥发量;氧化亚氮浓度利用Agilent 7890B气相色谱仪测定,采用微电子捕获器(ECD),工作温度300 ℃,尾吹气为N2,N2流量为20 mL·min−1,氧化亚氮排放通量计算公式参考文献[19],当时间间隔为1 d时,未测定日期的数据通过前后2次测量数据的算术平均值得出,当时间间隔为2 d时,求出这段时间的算术均值,之后乘以这段时间的长度,得到这段时间的累积挥发量;有机氮采用Bremner酸解法[20]测定。
小白菜种子发芽指数(GI)依照式(1)计算; NH3排放速率依照式(2)计算;N2O的排放速率依照式(3)计算。
stringUtils.convertMath(!{formula.content}) (1) stringUtils.convertMath(!{formula.content}) (2) 式中:F表示NH3排放速率,mg·(m2·h)–1;C表示标定过的硫酸浓度,mol·L−1;V和V0(空白)表示滴定消耗的硫酸体积,mL;M表示NH3的分子量,17.03 g·mol−1;L2表示静态箱与肥堆接触面积,m2;T为NH3的吸收时间。
stringUtils.convertMath(!{formula.content}) (3) 式中:F表示N2O排放速率,mg·(m2·h)−1;ρ为标准状况下N2O密度,1.978 kg·m−3;H表示箱内气体的高度,m;
dcdt 2. 结果与讨论
2.1 堆肥过程中基本指标的变化
1)温度的变化。如图2(a)所示,在好氧堆肥初期,堆体内部有机物质的分解和微生物的活动会产生大量的热,造成堆体温度的迅速上升。在堆肥第2天,各堆体均上升到了50 ℃以上;在第3天,各处理的温度都达到了55 ℃;在第5天均接近70 ℃。CK、SP和FR在高温期(>55 ℃)的持续时间分别为27、26和34 d,最高温度分别达到了68.9、70.0和68.8 ℃,符合我国畜禽废弃物无害化的标准[21]。27 d后,CK和SP处理温度开始下降至55 ℃以下,而FR处理的温度直到33 d后才开始低于55 ℃。这说明,添加糠醛渣延长了堆肥的高温期,而CK和SP的温度的变化趋势基本一致。
2) pH的变化。如图2(b)所示,随着堆体温度的升高,各处理的pH开始逐渐上升,在第9天达到最大值。这是由于微生物分解含氮化合物使其剧烈矿化,铵态氮大量积累所致[22-23];随着氨气的挥发和酸性物质的形成,pH会相应缓慢下降后逐渐趋于稳定[23]。在整个堆肥进程中,添加过磷酸钙可以明显降低堆体的pH;而FR处理的pH除第1天略低于CK外,在堆肥其他时间始终高于CK。这说明,添加糠醛渣后可提高堆体的pH,但提高的幅度并不大。
3)发芽指数。种子发芽指数(GI)是检验有机肥有无毒害的重要指标。当GI值大于50%时,可认为堆肥基本腐熟,当其大于80%时即可认为堆肥完全腐熟。图2(c)是本次堆肥中小白菜种子的发芽指数变化情况。堆肥刚开始时,由于肥料未腐熟,各处理GI值较低;从第18天开始,SP和FR处理的GI值大于50%;至第36天,SP处理GI值达到了90%;至堆肥结束,各处理GI值则分别达到了94.23%、110.36%和104.56%,这说明肥料已无毒害,可能是添加剂中携带的营养物质促进了种子的萌发。
 图 2 堆肥过程中的温度、pH和发芽指数的变化Figure 2. Changes of temperature, pH value and germination index during composting
图 2 堆肥过程中的温度、pH和发芽指数的变化Figure 2. Changes of temperature, pH value and germination index during composting2.2 堆肥过程中含氮气体(NH3和N2O)的变化
1)氨气排放速率。如图3(a)所示,各处理下氨气的大量挥发都主要集中在堆肥前9天,这与pH和温度的变化有关。堆肥初期的氨化细菌类代谢能力强,分解含氮有机物时可产生的大量
NH+4 2)氧化亚氮排放速率。如图3(b)所示,N2O主要来源于
NH+4 NO−x NO−3 NH+4 NO−3 NO−3 3)氨气累积排放量。通常,根据堆体温度的变化可以将好氧堆肥过程分为升温期、高温期、降温期和腐熟期,不同时期氨气排放速率的不同决定了该时期累积排放量的不同。如图4(a)所示,CK、SP和FR在升温期和高温期的氨气累积挥发量分别占整个堆肥期间氨气挥发总量的78.8%、79.3%和88.6%。由于CK处理与SP处理之间温度的差异不大,所以是因SP处理下较低的pH抑制了
NH+4 4)氧化亚氮累积排放量。如图4(b)所示,与氨气累积排放量同理,SP处理下,因其较高的N2O排放速率,造成升温期和高温累积排放量分别提高了2.72倍和1.68倍,随着降温期和腐熟期N2O排放速率降低,累积排放量下降了58.95%;FR处理下的N2O累积排放量在仅在升温期提高了1.3倍,其他时期N2O累积排放量均有所下降。至堆肥结束,SP处理下N2O总累积排放量增加了20.8%,FR处理下N2O总累积排放量减少了68.79%。
2.3 堆肥过程中氮素的变化
1)总氮的变化。堆肥总氮(TN)的变化情况如图5(a)所示。在堆肥的0~6 d,各处理TN质量分数逐渐上升到最大值,这可能是由堆体内含碳有机物的降解率高于氨气的排放率所致[33] 。之后,由于氨气挥发等氮素损失,各处理下TN呈下降趋势;经过高温期的降解和反应,肥堆的体积和质量不断下降,当下降速率远超过氮素减少的速率时就会产生浓缩效应,使TN质量分数开始上升。至堆肥结束,各处理下TN质量分数与堆肥初期相比分别增加了 9.29%、5.44%和14.40%。在整个堆肥过程中,SP处理的TN质量分数始终低于CK,但FR处理的TN却始终高于CK。
2)酸解有机氮的变化。氮素由有机氮和无机氮组成,有机氮是主要的氮组分,占总氮的90%以上,酸解总氮(THN)占有机氮的50%以上[19]。如图5(b)所示,由于微生物的代谢活动,有机氮被分解,CK、SP和FR下的酸解总氮持续下降,降幅分别为13.87%、7.98%和5.53%;添加剂的加入减缓了有机氮的降解,这可能是由于微生物优先分解添加剂携带的营养物质。随着氨氧化微生物和其他氮同化微生物活性的恢复,有机氮的合成再次启动[26],THN质量分数开始上升;另外,浓缩效应也会造成THN呈上升趋势。至堆肥结束,CK、SP和FR处理的THN与堆肥初期相比增加了5.08%、7.85%和9.07%。在整个堆肥过程中,SP处理中THN质量分数始终低于CK;FR处理在前15 d低于CK,这可能是由前期其氨气排放过高所致。通过对CK、SP和FR处理的THN和
NH+4 NH+4 3)铵态氮的变化。如图5(c)所示,各处理
NH+4 NH+4 NH+4 NH+4 NH+4 NH+4 NH+4 NH+4 4)硝态氮的变化。
NO−3 NH+4 NH+4 NO−2 NO−3 NH+4 NO−3 NO−3  图 5 堆肥过程中的总氮、酸解有机氮、铵态氮和硝态氮含量变化Figure 5. Changes of TN、THN、
图 5 堆肥过程中的总氮、酸解有机氮、铵态氮和硝态氮含量变化Figure 5. Changes of TN、THN、NH+4 NO−3 3. 结论
1)3个肥堆的高温期保持时间均满足好氧堆肥的腐熟要求;添加糠醛渣使高温期延长了7 d,添加过磷酸钙则降低了肥料的pH;发芽指数计算结果表明,添加剂的加入不会对植物生长产生毒性。
2)添加过磷酸钙后,氨气的排放速率和排放量均有所下降,但在高温期会产生大量的氧化亚氮,导致氧化亚氮的排放量上升;添加糠醛渣后则可延长堆体的高温期时长,并略微提高肥料的pH,这造成氨气排放较高,但其对氧化亚氮的减排效果明显。
3)添加过磷酸钙后可促进有机氮向铵态氮的转化,并抑制铵态氮向硝态氮的转化;添加糠醛渣可提高堆体总氮和有机氮含量,并促进铵态氮向硝态氮的转化。
-
图 2 堆肥过程中的温度、pH和发芽指数的变化
Figure 2. Changes of temperature, pH value and germination index during composting
图 5 堆肥过程中的总氮、酸解有机氮、铵态氮和硝态氮含量变化
Figure 5. Changes of TN、THN、
NH+4 -N andNO−3 -N content during composting表 1 堆肥材料理化性质
Table 1. Physical and chemical properties of composting materials
供试材料 含水率/% pH TC/% TN/% TP/% TK/% C/N 小麦秸秆 7.1 7.3 80.7 1.1 0.2 1.1 76.9 牛粪 45.7 7.6 38.2 2.2 0.9 2.0 17.7 过磷酸钙 0.9 3.5 — — — — — 糠醛渣 10.1 2.91 81.35 0.97 0.87 2.03 81.35
下载: 导出CSV
-
[1] 严铠, 刘仲妮, 成鹏远, 等. 中国农业废弃物资源化利用现状及展望[J]. 农业展望, 2019, 15(7): 62-65. doi: 10.3969/j.issn.1673-3908.2019.07.014 [2] 于法稳, 杨果. 农作物秸秆资源化利用的现状、困境及对策[J]. 社会科学家, 2018(2): 33-39. doi: 10.3969/j.issn.1002-3240.2018.02.005 [3] 尹瑞, 张鹤, 邱慧珍, 等. 不同碳氮比牛粪玉米秸秆堆肥的碳素转化规律[J]. 甘肃农业大学学报, 2019, 54(5): 68-78. [4] 巨晓棠, 谷保静, 蔡祖聪. 关于减少农业氨排放以缓解灰霾危害的建议[J]. 科技导报, 2017, 35(13): 11-12. [5] 宋修超, 郭德杰, 成卫民, 等. 工厂化条件下外源添加剂对猪粪堆肥过程中NH3和H2S的减排效果[J]. 农业环境科学学报, 2021, 40(9): 2014-2020. doi: 10.11654/jaes.2021-0221 [6] 史春梅, 王继红, 李国学, 等. 不同化学添加剂对猪粪堆肥中氮素损失的控制[J]. 农业环境科学学报, 2011, 30(5): 1001-1006. [7] 曹玉博, 张陆, 王选, 等. 畜禽废弃物堆肥氨气与温室气体协同减排研究[J]. 农业环境科学学报, 2020, 39(4): 923-932. doi: 10.11654/jaes.2020-0104 [8] 张涛, 李国学. 探究畜禽粪污资源化技术革新 促进养殖业绿色可持续发展[J]. 畜牧业环境, 2019(3): 29-31. [9] CAO Y B, WANG X, BAI Z H, et al. Mitigation of ammonia, nitrous oxide and methane emissions during solid waste composting with different additives: A meta-analysis[J]. Cleaner Production, 2019, 235: 626-635. doi: 10.1016/j.jclepro.2019.06.288 [10] 赵彬涵, 孙宪昀, 黄俊, 等. 微生物在有机固废堆肥中的作用与应用[J]. 微生物学通报, 2021, 48(1): 223-240. [11] 胡雨彤, 时连辉, 刘登民, 等. 添加硫酸对牛粪堆肥过程及其养分变化的影响[J]. 植物营养与肥料学报, 2014, 20(3): 718-725. [12] 顾文杰, 张发宝, 徐培智, 等. 堆肥反应器中硫磺对牛粪好氧堆肥的保氮效果研究[J]. 植物营养与肥料学报, 2011, 17(1): 224-230. doi: 10.11674/zwyf.2011.0131 [13] 顾文杰, 张发宝, 徐培智, 等. 添加无机磷肥对农业固体废弃物好氧堆肥的影响[J]. 广东农业科学, 2010, 37(4): 101-105. doi: 10.3969/j.issn.1004-874X.2010.04.034 [14] 陶勇, 湛含辉, 唐铄松. 添加苹果渣对猪粪好氧堆肥理化性状的影响[J]. 中国土壤与肥料, 2019(5): 135-140. doi: 10.11838/sfsc.1673-6257.18416 [15] CAO Y B, WANG X, LIU L, et al. Acidification of manure reduces gaseous emissions and nutrient losses from subsequent composting process[J]. Environmental Management, 2020, 264(24): 110454. [16] 吴娟. 过磷酸钙对猪粪堆肥碳、氮转化及减缓有机质降解技术机制研究[D]. 北京: 中国农业大学, 2017. [17] 王素芬, 苏东海, 周凌云. 废物糠醛渣的农业利用研究进展[J]. 河北农业科学, 2009, 13(11): 97-99. doi: 10.3969/j.issn.1088-1631.2009.11.037 [18] 周海瑛, 邱慧珍, 杨慧珍, 等. C/N比对好氧堆肥中NH3挥发损失和含氮有机物转化的影响[J]. 干旱地区农业研究, 2020, 38(2): 69-77. doi: 10.7606/j.issn.1000-7601.2020.02.10 [19] 杨岩, 孙钦平, 李妮, 等. 添加过磷酸钙对蔬菜废弃物堆肥中氨气及温室气体排放的影响[J]. 应用生态学报, 2015, 26(1): 161-167. [20] BREMNER J M. Organic forms of nitrogen[M]//BLACK C A. Methods of Soil Analysis, Part 2. Madison: Wisconsin, 1965: 1238-1255. [21] 中华人民共和国农业行业标准. 有机肥料行业标准 NY 525-2012[S]. 北京: 中国农业出版社, 2012. [22] NAKHSHINIEV B, PERERA C, BIDDINIKA M K, et al. Reducing ammonia volatilization during composting of organic waste through addition of hydrothermally treated lignocellulose[J]. International Biodeterioration 2014, 96: 58-62. [23] 王友玲, 邱慧珍, PHILIP G, 等. 通风方式对牛粪堆肥氨气排放与氮素转化的影响[J]. 农业机械学报, 2020, 51(11): 313-320. doi: 10.6041/j.issn.1000-1298.2020.11.034 [24] GAJALAKSHMI S, ABBASI S A. Solid waste management by composting: State of the art[J]. Environmental Science and Technology, 2008, 38(5): 311-400. [25] 黄懿梅, 苟春林, 来航线, 等. 两种添加剂对猪粪玉米秸秆堆肥氮素转化和堆肥质量的影响[J]. 干旱地区农业研究, 2005(6): 116-122. [26] 付丽丽, 阚培赢, 刘娟, 等. 颗粒粒径对玉米芯混合鸡粪堆肥氨气减排的影响[J]. 辽宁石油化工大学学报, 2019, 39(5): 36-39. doi: 10.3969/j.issn.1672-6952.2019.05.007 [27] CUI P, LIAO H P, BAI Y D, et al. Hyperthermophilic composting reduces nitrogen loss via inhibiting ammonifiers and enhancing nitrogenous humic substance formation[J]. Science of the Total Environment, 2019, 692: 98-106. doi: 10.1016/j.scitotenv.2019.07.239 [28] 王成. 堆肥过程N2O减排控制与木质纤维素降解微生物学机理研究[D]. 杭州: 浙江大学, 2014. [29] 吴伟祥, 李丽劼, 吕豪豪, 等. 畜禽粪便好氧堆肥过程氧化亚氮排放机制[J]. 应用生态学报, 2012, 23(6): 1704-1712. [30] 刘学玲. 猪粪高温堆肥中氮转化复合微生物菌剂及其保氮机理的研究[D]. 杨凌: 西北农林科技大学, 2012. [31] 罗一鸣, 李国学, FRANK S, 等. 过磷酸钙添加剂对猪粪堆肥温室气体和氨气减排的作用[J]. 农业工程学报, 2012, 28(22): 235-242. doi: 10.3969/j.issn.1002-6819.2012.22.033 [32] MAEDA K, TOYODA S SHIMOJIMA R, et al. Source of nitrous oxide emissions during the cow manure composting process as revealed by isotopomer analysis of and amoA abundance in betaproteobacterial ammonia-oxidizing bacteria[J]. Applied and Environmental Microbiology, 2010, 76(5): 1555-1562. doi: 10.1128/AEM.01394-09 [33] 唐尙柱, 赵晓海, 斯鑫鑫, 等. 不同镁/磷盐添加剂对蓝藻堆肥的氮素损失控制效果[J]. 农业环境科学学报, 2021, 40(2): 428-435. doi: 10.11654/jaes.2020-0940 [34] 杨宏, 姚仁达. pH和硝化细菌浓度对氨氮氧化速率的影响[J]. 环境工程学报, 2017, 11(5): 2660-2665. doi: 10.12030/j.cjee.201512155 [35] 王淑莹, 李论, 李凌云, 等. 快速启动短程硝化过程起始pH值对亚硝酸盐积累的影响[J]. 北京工业大学学报, 2011, 37(7): 1067-1072. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 5622
- HTML全文浏览数: 5622
- PDF下载数: 55
- 施引文献: 0































