


-
随着我国污水处理规模的迅速发展,污泥产量也逐年剧增。截至2015年底,我国脱水污泥年产量超过3×107 t,预计2020年污泥产量将再增加1倍[1-2]。因此,污泥的处置是我国目前面临的一大难题。污泥堆肥可以杀死有害细菌,增加和稳定其中的腐殖质,降低所含重金属的毒性效应[3-4]。因此,腐熟污泥农用是一种消纳污泥的有效途径[5-6]。我国颁布的土地改良泥质、农用泥质和园林绿化泥质标准规定了污泥使用量、污泥累计使用量、连续使用年限和施用频率的要求,但针对腐熟污泥在具体土壤类型上如何施用并没有相关要求[7]。石灰性土壤作为广泛分布在我国干旱和半干旱地区的土壤类型,其碱性特征使其容易和重金属形成难溶性氢氧化物等沉淀[8]。因此,该类土壤对重金属污染表现出“不敏感性”特征,该特征极易导致污泥过量施用而造成土壤质量的降低[9]。
土壤微生物是土壤生态系统中最活跃的成分,污泥施用造成土壤pH,阳离子交换量,电导率和团聚体稳定性等理化性质发生变化[10-13],这些变化又对土壤微生物活性、微生物种类、数量、分布等多样性产生影响[14-17]。其中需要关注的根际土壤微生物多样性是表征外源物质引起土壤质量变化的敏感指标[18],该多样性的变化值得重视。
已有研究[19]表明,石灰性土壤中微生物学特征及其微生物群落结构对调节土壤养分供应有重要作用,污泥添加同样会对石灰性土壤微生物量以及碳、氮等微生物学性质产生影响[20],且上述影响除与土壤性质和作物系统有关外[20-21],不同污泥的施用量和施用时期长短也是影响土壤微生物性质及其多样性的重要因素[22-24]。目前,污泥施用对根际和非根际石灰性土壤中细菌多样性差异的研究还较少,尤其是过量施用污泥对土壤微生物特征变化的影响尚缺乏研究。因此,开展不同用量腐熟污泥对石灰性土壤细菌多样性特征影响研究,可为其在石灰性土壤中施用的生物学质量评价和合理施用策略提供参考。
-
实验地点位于河南省洛阳市(北纬34°41′,东经112°27′),该地区属于干旱半干旱气候,年均气温和降雨量分别为15.6 ℃和460 mm,土壤类型属于典型石灰性土壤,其基本理化性质和重金属含量如表1和表2所示。供试作物为夏玉米(夏玉米品种为郑丹958),夏玉米在每年6月种植,10月收获。
供试污泥取自洛阳市某污泥处理厂,该污泥配以秸秆等辅料,按照秸秆与污泥重量比1∶5均匀混合,通过好氧高温堆肥后制得腐熟污泥,其重金属含量与《农用污泥中污染物控制标准》(GB 4284-2018)所规定的值相比,除As符合B级污泥限值外,其余重金属符合A级污泥限值。
-
根据污泥的施用量不同,实验设置5个处理,分别为0、3.75、7.5、37.5、75 t·hm−2等5个污泥施用量。其中,5个污泥施用量分别对应非根际土壤样品为A01,A03,A05,A07,A09;对应的根际土壤样品为A02,A04,A06,A08,A10。污泥的施用量参考我国污泥土地利用标准(GJ/T 309-2009)进行设计,每个小区面积30 m2,随机排列,并重复3次。实验从2015年10月开始,连续2 a种植冬小麦-夏玉米,并且在冬小麦季把污泥和180、90、90 kg·hm−2的N,P2O5,KO2肥作为底肥一次性施入土壤,采用人工撒施和多次翻耕的方式使污泥和土壤均匀混合。期间进行正常的灌溉和管理。2017年10月,夏玉米收获后,采集0~20 cm非根际土壤(没有作物生长的0~20 cm表层土壤)和根际土壤(根际土壤采用“抖根法”,先将玉米根系从土壤中挖出,抖掉与根系结合松散的土壤,收集与根际紧密结合在0~4 mm范围的土壤作为根际土壤)。采集的新鲜土样分成2份,其中一份放入50 mL灭菌的离心管后,于−80 ℃冰箱保存用于细菌多样性分析;另一份风干用于测定土壤其他参数。
-
微生物多样性分析主要是在编码核糖体RNA的核酸序列保守区开展研究,其中细菌主要基于的是16S保守区。本研究采用细菌16S rDNA(V3+V4)扩增利用的引物为338F(5′-ACTCCTACGGGAGGCAGCA-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′),通过PCR扩增后形成测序文库,然后利用Illumina HiSeq 2500进行测序,上述测序结果覆盖了土壤样品中98%以上细菌的真实组成,而且OTUs丰度稀释曲线随着测序量增加趋向平坦,说明测序数量足够,能够反应出样品中物种组成特征。
-
根据PE reads之间的Overlap关系,将Hiseq测序得到的双端序列数据拼接(Merge)成一条序列Tags,同时对Reads的质量和Merge的效果进行质控过滤。使用FLASHE. v1.27、Trimmomatic. v0.33和Uchime v4.2等软件进行质控过滤后,得到优化序列(Tags);将优化序列进行聚类,划分OTU,并根据OTU的序列组成得到其物种分类;通过Alpha多样性分析研究样品内部的物种多样性,统计样品在97%相似度水平下的Ace、Chao、Shannon及Simpson指数;根据距离矩阵获得相应距离下的样品PCA(主成分分析);利用16S功能基因和冗余分析(RDA)分别对样品功能基因丰度和细菌群落组成与土壤参数之间的相关关系进行分析。
-
供试污泥和采集的土壤样品风干过筛后分析以下指标:全氮,全磷,全钾、有机质、pH、电导率。上述指标测定方法参考文献中的方法[25]。重金属测定:取0.500 g通过100目筛样品,置于聚氟坩埚中,加入10 mL HF,8 mL HNO3和3 mL HClO4后静置12 h,以除去有机质和硅酸,然后在加热板上加热,温度逐渐升高至200 ℃,稳定温度加热至白烟产生且坩埚中残存少量浅白色黏稠状物质为止。完全冷却后,加2%的HNO3 2~3 mL溶解残存物质,用25 mL容量瓶定容待用,空白和标准样品同时消化,样品中的Zn、Cu、Pb、Cr、Cd利用等离子发射光谱法(美国,PE optima5300DV)测定。As采用双道原子荧光光度计(北京,吉田AFS-9120)测定。每个样品重复3次,使用SPSS 19.0软件进行单因素方差分析和相关性分析。
-
石灰性土壤的主要理化性质受污泥施用的影响如表3所示。随着污泥用量的增加,根际和非根际土壤pH呈现降低趋势,但土壤中电导率、有机质、总氮和总磷有明显增加的趋势。与对照相比,当污泥施用量超过7.5 t·hm−2时,污泥施用对上述参数的影响会显著增加。污泥施用也增加了土壤中Pb、Zn、Cu、Cr、Cd、As等重金属的含量,当污泥施用量为75 t·hm−2时,土壤中Zn、Cu、Cd的含量与其他处理相比具有显著差异(P<0.05),而土壤中As在7.5,37.5,75 t·hm−2的污泥施肥量条件下并没有显示出显著差异(P>0.05)。和非根际土壤相比,根际土壤中有机质、电导率、As含量高于对应样品中的非根际土壤含量。
-
把序列之间相似性高于97%的Tags进行聚类,获得OTU。图1显示了各污泥施用水平下根际和非根际石灰性土壤OTU丰度(图1中不同小写字母表示各样品平均值具有显著差异(P<0.05))。非根际土壤样品中细菌的OTU丰度大小依次为A09 < A01 < A03 < A05 < A07。由此可见,当污泥施用量为3.75~37.5 t·hm−2时,非根际土壤中细菌OTU丰度随污泥施用量增加而增加;当污泥用量为75 t·hm−2时,样品A09中OTU的丰度与A03,A05,A07相比显著降低,并且低于A01丰度。根际土壤中细菌OTU的丰度大小顺序与非根际土壤有所不同,其OTU丰度依次为A10 < A08 < A02 < A04 < A06。由此可见,当污泥施用量在37.5 t·hm−2时,根际土壤中OTU丰度便开始降低,而且A10样品和其他各样品之间的OTU丰度相比具有显著差异。这说明污泥施用对根际土壤细菌OTU丰度的影响更加敏感,而且当污泥用量超过一定范围内会显著降低根际-非根际土壤细菌的OTU丰度。
Venn图可以表示不同样品之间共有、特有的OTU数目,直观地表现出个样品之间OTU的重合情况(图2)。结合OTU所代表的物种,可以找出不同样品中的共有微生物。不同样品用不同颜色表示,交叠部分数字为2个样品之间共有的OTU个数。非根际土壤样品A01、A03、A05、A07、A09中总OTU数量为1 896个,共有的数量为993个,占总数52.37%,各样品中特有OTU数量分别为4、11、11、9、12个。这说明污泥施用于石灰性土壤中,改变了非根际土壤中细菌OTU丰度,但随着污泥用量的增加其对土壤中特有微生物的数量影响不大。根际土壤中共有的OTU数量为882,占总共OTU数量1 895比例为46.54%,因此,污泥施用对根际土壤OTU的影响大于非根际土壤。其中A02、A04、A06、A08、A10样品中特有OTU数量分别为8、10、4、13、34个,除A08样品外,随着污泥用量的增加,根际土壤中各处理特有OTU的数量呈增加趋势。污泥用量在75 t·hm−2时,A10和A09样品中特有微生物数量多于其他样品,且根际土壤比非根际土壤增加了183%。
-
石灰性土壤施用不同量污泥后,对细菌多样性指数(Shannon、Simpson)和丰富度指数(Ace,Chao)的影响如表4所示。在污泥施用量为75 t·hm−2的条件下,根际和非根际土壤的Shannon指数与其他各处理相比有显著差异(P<0.05),其他各处理之间无显著差异(P>0.05)。当污泥用量为3.2~37.5 t·hm−2时,同一污泥施用水平下,根际土壤的Shannon指数与非根际土壤无显著差异(P>0.05)。非根际和根际土壤的Simpson指数在污泥施用量为37.5 t·hm−2时最低,在污泥施用量为75 t·hm−2时最高,该指数值分别为0.004 2和0.003 2,0.009 4和0.009 8,说明37.5 t·hm−2的污泥施用量有利于细菌多样性指数的增加,当污泥用量达到75 t·hm−2时会显著降低细菌多样性。不同污泥施用量的Ace,Chao指数在非根际和根际土壤变化趋势一致,都是污泥施用量为75 t·hm−2时最小,说明75 t·hm−2的污泥施用量不但降低了细菌多样性指数,而且降低细菌的丰富度指数。此外,根际样品A02和A04的Ace,Chao指数比对应的非根际土壤高,但非根际样品A05,A07,A09的Ace,Chao指数大于对应的根际土壤,说明随着污泥施用量的增加会对根际细菌丰富度指数产生不利影响。
-
PCA主成分分析可将样品细菌多样性差异反映在二维坐标图上,主成分轴用来解释差异的综合信息,第1主成分轴解释最多的差异,第2主成分轴解释第2多的差异。污泥施用后对石灰性土壤中根际和非根际土壤细菌主成分分析表明,随着污泥施用量的增加,土壤中各处理细菌群落结构距离越远,说明对其群落组成造成的差异越大。对非根际土壤细菌群落结构而言,主成分1的贡献率为56.48%,主成分2的贡献率为24.08%,两者累计为80.56%(图3)。在第1主成分轴(PC1)上,污泥施用后非根际石灰性土壤上细菌群落结构差异较大,且随着污泥用量的增加土壤中细菌群落结构差异变大。在第2主成分轴(PC2)上,污泥用量较低的A01和A03样品细菌群落结构相近,随着污泥用量增加A05,A07,A09样品之间细菌群落结构相近。
根际土壤细菌群落结构多样性的主成分1占66.15%,主成分2占23.36%,二者累计贡献率为89.51%。在第1主成分轴上A02、A04、A06、A08之间的差异较小,但与A10之间的差异最大。在第2主成分轴上,A02与A04之间的差异最小,与A10之间的差异最大。当污泥施用量增加时,在主成分1对根际土壤细菌会产生较大的影响,在主成分2对非根际土壤产生较大影响。
-
根际和非根际土壤细菌在门水平分类上排名前10丰度的物种如图4所示。非根际土壤中细菌门类群集中于变形菌门(Proteobacteria)、酸杆菌门(Acidobacteria)、拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)(相对丰度>5%)。土壤中变形菌门和酸杆菌门细菌是土壤微生物的重要成员,在土壤物质循环和生态环境构建过程其道非常重要作用。其中,各处理变形菌门的丰度都超过48%,因为变形菌门适合养分含量高的土壤环境,且与土壤有机质和全氮含量显著相关[26]。污泥施用增加了土壤中有机质和养分(表3),使得变形菌门的丰度增量范围为1.63%~4.05%。污泥施用量在3.75~37.5 t·hm−2时拟杆菌门丰度增加明显,但污泥施用降低了酸杆菌门丰度(降低幅度为31.58%~34.59%)和放线菌门的丰度。同时,当污泥施用量为75 t·hm−2时,放线菌门的丰度降低达51.75%,这将不利于放线菌抵御病原菌和防治病害作用的发挥[27]。与非根际土壤相比较,根际土壤细菌在门水平上排名前10的种类相同,但其丰度有所变化,根际土壤中细菌门类群集中于变形菌门(Proteobacteria)、拟杆菌门(Bacteroidetes)、酸杆菌门(Acidobacteria)。当污泥用量增加时,变形菌门丰度增加不明显。与对照相比,当污泥用量为75 t·hm−2时,拟杆菌门有明显的增加,丰度增加达1.45倍;酸杆菌门和放线菌门丰度降低49.74%和80.57%。其中酸杆菌门的降低不但与土壤pH的降低有关,还与污泥施用导致土壤氮素增加也有密切关系[28]。由此可见,随着污泥用量的增加,对根际和非根际土壤细菌门的丰度变化会产生不同影响。
-
由上述研究结果可知,污泥施用量达75 t·hm−2时对根际和非根际土壤中细菌多样性产生较大影响。因此,通过KEGG(代谢途径分析数据库)分析空白对照和污泥施用量为75 t·hm−2的处理可知,污泥过量施用可造成根际和非根际土壤微生物群落功能基因在代谢途径上的差异。KEGG功能分析表明(见图5),2个处理的前十大比例的代谢途径相同,但污泥施用为75 t·hm−2时,明显降低了A09样品中碳水化合物代谢、脂类代谢、氨基酸代谢以及膜运输代谢功能,说明过量的污泥施用已经抑制了土壤系统中C、N的循环过程,并显著增加了代谢途径有辅酶因子和维生素代谢、核苷酸代谢、翻译和信号传导功能,而这些功能的增加可能源于放线菌门数量减少所带来的环境胁迫响应。样品A10的碳水化合物代谢、脂类代谢、能量代谢、外源性物质的生物降解和代谢以及膜运输代谢显著降低,但辅酶因子和维生素的代谢、核苷酸代谢、翻译功能显著增加。不施用污泥时,样品A02与A01相比只有异源生物降解代谢,翻译,膜运输,信号传导功能增加,其余功能降低。上述代谢差异为判断污泥添加对土壤功能质量影响提供了依据。
-
污泥施用不但对土壤理化性质变化产生影响,而且会造成重金属积累。因此,为了明确上述不同参数对细菌群落组成影响大小,本研究选择土壤pH、电导率(EC)、有机质(OM)、总氮(TN)、总磷(TP)以及污泥添加对重金属影响较为显著的Cd,Zn,Cu,Pb作为环境变量,通过冗余分析(RDA)探查上述参数与纲分类水平下非根际土样品(组1)和根际样品(组2)细菌群落组成的相关性(图6)。第1排序轴解释了细菌群落变化的34.03%,第2排序轴解释了细菌群落变化的20.73%,2个排序轴累积贡献率达54.76%。从图中可以看出,土壤TN、Cd、Cu、TP与细菌群落的相关性大,其中TN、Cd、Cu和土壤Shannon和Simpson多样性指数达到显著相关(表5),EC、Pb、Zn与细菌群落的关系次之,而土壤中pH和OM对细菌群落组成影响较小。上述结果的原因在于,3.5~37.5 t·hm−2的污泥施用量并没有对土壤pH和OM造成显著影响,但已明显影响了土壤中TN、TP、Cd、Cu等含量。
-
微生物多样性是土壤质量的重要参数,而且土壤微生物对其环境改变极为敏感,其多样性变化与土壤退化有密切关系[29]。适量地投放污泥有利于土壤质量的维持,但过量或长期施用污泥会导致土壤质量的降低。ANDRES等[30]通过向土壤中施用污泥,初始阶段均可增加土壤微生物生物量,但1 a以后污泥对土壤微生物活性已消失。长达30 a在大豆、生菜、芥菜上施用污泥的研究表明,污泥长期施用对土壤微生物有负面影响[31]。本研究连续2 a施用75 t·hm−2污泥后,根际和非根际土壤中细菌OTU丰度、多样性Shannon指数和丰富度Ace,Chao指数均会显著降低。因此,针对不同土壤类型开展污泥施用后的微生物多样性变化研究,并根据研究结果采取“隔年间歇式”污泥施用模式,可做到既利用了污泥养分,又能保证土壤质量。
有研究表明,土壤细菌群落多样性的变化与土壤pH、有机质、氮、磷有关,其中pH的变化被认为是影响土壤微生物多样性的主要因素[32-33]。和对照相比,研究中污泥用量为37.5 t·hm−2时,pH才有显著差异(表2),但由于石灰性土壤上pH变幅较小,使得其不是影响细菌多样性变化的一个重要因素,本研究的RDA分析也印证了该结论。由于重金属对细菌的毒害作用,会显著降低其多样性[34]。较高含量的Cu,Pb,Zn重金属污泥,施入土壤会造成土壤中微生物种群由细菌向真菌转化[35]。富含Zn的污泥施用于沙壤土中会导致了所有微生物群落结构的变化(即细菌,真菌,古生菌,放线菌和根瘤菌),其中真菌群落受其影响最大[36]。本研究污泥的施用虽然增加了土壤中重金属积累,但由于我国污泥重金属含量有降低趋势[37],以及石灰性土壤对重金属的“不敏感性”等原因,使得除Cd,Cu外,其他重金属和与细菌群落的相关性并不大。
选择根际土壤进行微生物多样性研究,更能灵敏地反映出外源物质施用对土壤质量的影响[38]。石灰性土壤上通过对细菌OTU丰度和PCA主成分分析,也表明污泥施用对根际土壤细菌的影响更敏感。另外,上述结果只是污泥连续施用2 a后的结论。为了更加全面的准确了解污泥对该类施用对该类土壤微生物多样性的影响,选择多年根际土壤中细菌和真菌的变化研究,才能准确评价污泥农用对微生物多样性的影响,同时也为推荐污泥施用量提供依据。
-
1)污泥在石灰性土壤上施用会改变根际和非根际土壤中细菌丰富度和多样性指数,而且在2 a连续施用量为37.5 t·hm−2的条件下,不利于根际土壤中细菌多样性增加。当污泥施用量达到75 t·hm−2时,会显著降低根际和非根际土壤中细菌的多样性指数。因此,考虑到根际微生物对外源物质响应的灵敏性,此类土壤中连续2 a的污泥施用量宜控制在37.5 t·hm−2以下。
2)污泥施用造成土壤理化性质的变化是影响土壤细菌群落变化的一个主要因素,另外重金属种类对微生物群落组成的影响也存在差异。本研究在2 a连续施用污泥后,土壤中TN、Cd和Cu是影响细菌群落的主要参数。
3)污泥施用量超过75 t·hm−2时,将会抑制根际和非根际土壤中C、N的循环代谢功能,降低酸杆菌门和放线菌门丰度。这说明此污泥施用量下,土壤质量已经开始下降。
污泥施用对根际和非根际石灰性土壤中细菌多样性的影响
Effects of sludge addition on bacterial diversity in rhizosphere and non-rhizosphere calcareous soil
-
摘要: 为研究污泥施用对石灰性土壤细菌多样性的影响,在连续2年种植夏玉米大田系统中,施用不同量的腐熟污泥,采集根际和非根际土壤进行16S rDNA-V3-V4区高通量测序,分析污泥施用对石灰性土壤细菌丰富度、多样性指数、群落结构、功能基因在代谢途径的影响,同时对细菌群落和环境因素之间的相关性进行分析。结果表明:污泥用量分别在3.5~37.5 t·hm−2和3.5~7.5 t·hm−2有利于非根际和根际土壤OTU数量增加;但污泥在75 t·hm−2过量施用时,根际和非根际土壤细菌的Shannon多样性指数显著降低,Simpson指数显著增加,Ace,Chao丰富度指数显著降低。主成分和门水平物种丰度分析表明:污泥施用量不同会造成根际和非根际细菌群落结构差异,并且当污泥施用量为75 t·hm−2时,明显降低了非根际土壤放线菌门的丰度;污泥施用量在3.75~37.5 t·hm−2时,非根际土壤中拟杆菌门丰度会明显增加。与对照相比,当污泥用量为75 t·hm−2时,根际土壤拟杆菌门有明显的增加,增加丰度达1.45倍,但酸杆菌门和放线菌门丰度降低了49.74%和80.57%。冗余分析(RDA)表明,土壤TN、Cd、Cu、TP是影响土壤细菌群落最主要的因素,其中TN、Cd、Cu和细菌Shannon、Simpson多样性指数呈现显著相关性(P< 0.05)。由此可见,连续过量施入污泥会对根际和非根际石灰性土壤中细菌多样性造成不利影响,而且上述微生物多样性变化可作为污泥合理施用的判断依据。Abstract: In order to study the effects of sludge application on bacterial diversity of calcareous soil, different amounts of compost sludge were applied in a 2 consecutive year-summer maize field system. The high-throughput sequencing of 16S rDNA-V3-V4 region was carried out for collected rhizosphere and non-rhizosphere soil. Effects of sludge application on bacterial richness, diversity index, community formation and functional genes were analyzed. The correlation between bacterial community and environmental factors was also determined. The results showed that the amounts of OTU in these two kinds of soil increased at sludge addition amounts of 3.5~37.5 t·hm−2 in non-rhizosphere soil and 3.5~7.5 t·hm−2 in rhizosphere soil, respectively. However, at sludge addition amounts above 75 t·hm−2, Shannon index of bacterial diversity in rhizosphere and non-rhizosphere soil decreased significantly, Simpson index increased significantly, Ace and Chao indices for bacterial abundance also decreased significantly. Principal component and phylum-level species abundance analysis showed that different sludge addition amount could cause the differences in rhizosphere and non-rhizosphere bacterial community structure, and the abundance of actinobacteria in non-rhizosphere soil was significantly reduced at the sludge addition amount of 75 t·hm−2. The abundance of Bacteroidetes in non-rhizosphere soil increased significantly at the sludge addition amounts of 3.75~37.5 t·hm−2. Compared with the control, at the sludge addition amount of 75 t·hm−2, the abundance of Bacteroidetes in rhizosphere soil increased by 1.45 times, but the abundances of Acidobacteria and Actinobacteria decreased by 49.74% and 80.57%, respectively. Redundancy analysis (RDA) showed that soil TN, Cd, Cu and TP were the most important factors affecting soil bacterial community, among which TN, Cd, Cu were significantly correlated with Shannon and Simpson diversity indexes (P<0.05). Therefore, the continuous and excessive sludge application can adversely affect bacterial diversity in rhizosphere and non-rhizosphere calcareous soil, Moreover, the changes of microbial diversity indices mentioned above can be used as a basis for the rational sludge application.
-
Key words:
- sludge /
- calcareous soil /
- rhizosphere /
- non-rhizosphere /
- bacterial diversity
-
生物炭(biochar,BC)是生物质在限氧条件下热解(<700 ℃)形成的固态的、难熔的、高度芳香化的富含碳的材料[1],被广泛用于固碳[1-2]、土壤改良[3-4]、水体和土壤污染修复[5]等领域. 溶解性黑碳(dissolved black carbon,DBC)是BC中可溶于水的具有稠环芳烃结构的组分[6]. BC的大规模施用导致DBC释放[7],并通过灌溉、渗透、地表径流在环境中迁移[8]. 相比于BC,DBC具有较小的芳香微域结构和更多的羧基和羟基官能团[8-9],有更高的反应活性和更强的环境迁移能力[9]. 据估计,DBC全球河流通量每年可达2650万t,占河流进入海洋溶解有机碳通量的10%,是陆地BC输入海洋的一个重要来源[6, 10]. DBC的芳香微域结构是有机物和重金属吸附/络合的高能位点[9],会影响污染物的吸附、迁移、转化等关键环境过程[8]. DBC富含的羧基和羟基官能团是其在氧化-还原反应中的重要电子来源[8],参与有机污染物的光化学转化过程[10-11]. 原料和热解温度(heating temperature,HTT)是影响BC性质和用途的关键因素,也必然决定了DBC的结构特征和环境行为[12]. 研究发现,木质素含量较高的稻壳制备的BC芳香性和产量显著高于低木质素含量的甘蔗所制备的BC[3]. 另外,相比于低温(<500 ℃)制备的BC,高温(>500 ℃)BC往往具有更高的芳香性、灰分和pH[5]. 因此,深入理解水环境中不同原料和HTT的DBC与有机污染物的相互作用,有助于更好地预测BC施用环境中污染物的环境行为和评估污染物的生态风险.
光化学转化是影响水环境中有机污染物环境归趋的重要过程[13]. 水环境中光敏剂的分子结构(芳香性结构和分子大小)和环境条件(pH和盐离子)等是影响光降解的重要因素[14]. 与水环境中天然溶解有机质(natural organic matter,NOM)相比,人工来源的DBC稠环度高,芳香性强,分子量较小,这使其具有更高的光电转化效率[8, 15],更易产生羟基自由基(·OH)、单线态氧(1O2)和激发三重态DBC(3DBC*)等活性中间体(reactive intermediates,RIS)[10]. DBC的这些特性导致其对水环境中有机污染物的光降解过程的影响较NOM更为复杂[16]. 研究发现,DBC促进了塑化剂(如邻苯二甲酸二乙酯)[16]、杀虫剂(如吡虫啉)[12]、个人护理品(如阿替洛尔)[15]和性激素(如17-β雌二醇)[10]等多种有机污染物的光降解. 然而,腐殖酸(humic acid, HA)(DBC的重要组成部分)也可能会因遮光效应抑制有机物的光降解[13]. 可见,DBC参与的水环境中有机污染物的降解过程仍然存在不确定性.
新污染物是指由人类活动造成的,但尚无法律法规和标准规定或规定不完善的,危害人体健康和生态环境的污染物,主要包括环境内分泌干扰物、全氟化合物、微塑料和抗生素(antibiotics, ATs)四大类[17],而我国对于新污染物的毒理学风险评估、替代技术开发的方面还有局限性. ATs是一类由微生物分泌、化学合成或半合成的化合物,因其对微生物生长代谢的抑制而被广泛应用于医疗、畜牧业、水产养殖等领域[18]. 四环素(tetracycline, TC)因其广谱抗菌性和低廉的价格成为使用量最大的ATs之一. 2015年,全球TC使用量达6万t[19],这也导致TC在地表水、地下水和饮用水等水环境中被广泛检出,含量为ng·L−1—μg·L−1[20-21]. TC对水生动物(水生植物、浮游动物和鱼类)具有较强的生物急性毒性,能够抑制藻类和水生植物叶绿素的合成[22-23]、促进氧化应激反应的产生,影响浮游动物和鱼类的生长发育[22],同时TC还能够破坏水体微生物群落结构[21],损害水生生态系统的结构和功能[24]. 另外,ATs的长期暴露能够促进抗生素抗性基因的传播扩散,增加人体对ATs的抗药性[25],对人体健康产生极大的威胁[18, 26]. ATs在水环境中的光降解过程引起了广泛关注[13]. ATs的光降解主要受ATs种类[15]、环境条件(pH、盐离子)[27]、光敏剂(NOM、DBC)[10]等影响. 目前,水环境中DBC存在时,ATs的光降解研究以往主要集中在单一HTT制备的DBC对ATs光降解的影响上[13, 28],很少涉及不同原料和HTT的DBC对TC光降解的影响.
本研究以不同HTT(300—600 ºC)制备的湿地植物芦苇(Phragmites australis)和芦竹(Arundo donax)BC为原料提取DBC,探究HTT对DBC理化性质及结构特征的影响,系统研究了不同HTT的DBC对水环境中典型ATs TC的光降解的影响,并探讨了水中常见离子和pH对TC光降解过程的影响,以期阐明DBC影响TC光降解的主要机制. 研究结果将为评估DBC的环境行为及其对共存有机污染物的环境归趋的预测和生态风险的评估提供理论依据.
1. 材料与方法(Materials and methods)
1.1 实验材料
本研究所用BC均是实验室前期以湿地植物芦苇和芦竹为原料,利用限氧慢速热解法在300—600 ℃条件下制备[29]. DBC的提取步骤如下[10, 13]:将0.2 g BC置于20 mL超纯水中,室温条件下130 r·min−1振荡12 h,过0.45 μm醋酸纤维膜,收集滤液. 将滤膜上的BC重新分散在20 mL超纯水中,用水浴超声仪(KQ-500DE,昆山舒美,中国)在40 kHz频率下水浴超声30 min,过0.45 μm醋酸纤维膜后再次收集滤液. 重复上述超声提取过程两次,合并收集到的滤液,即为DBC溶液[10, 15]. 芦苇DBC标记为LW300、LW400和LW500,芦竹DBC标记为LZ300、LZ400、LZ500和LZ600,数字代表BC的HTT. 将DBC置于4 ℃冰箱中避光保存. TC(纯度>99%)购于生工生物工程(上海)股份有限公司,理化性质见表1.
CAS 分子式Molecular formula 溶解度Solubility pKa 结构式Structural formula 64-75-5 C22H25ClN2O8 19.7 g·L−1(25 ℃) pKa1=3.30pKa2=7.68pKa3=9.68 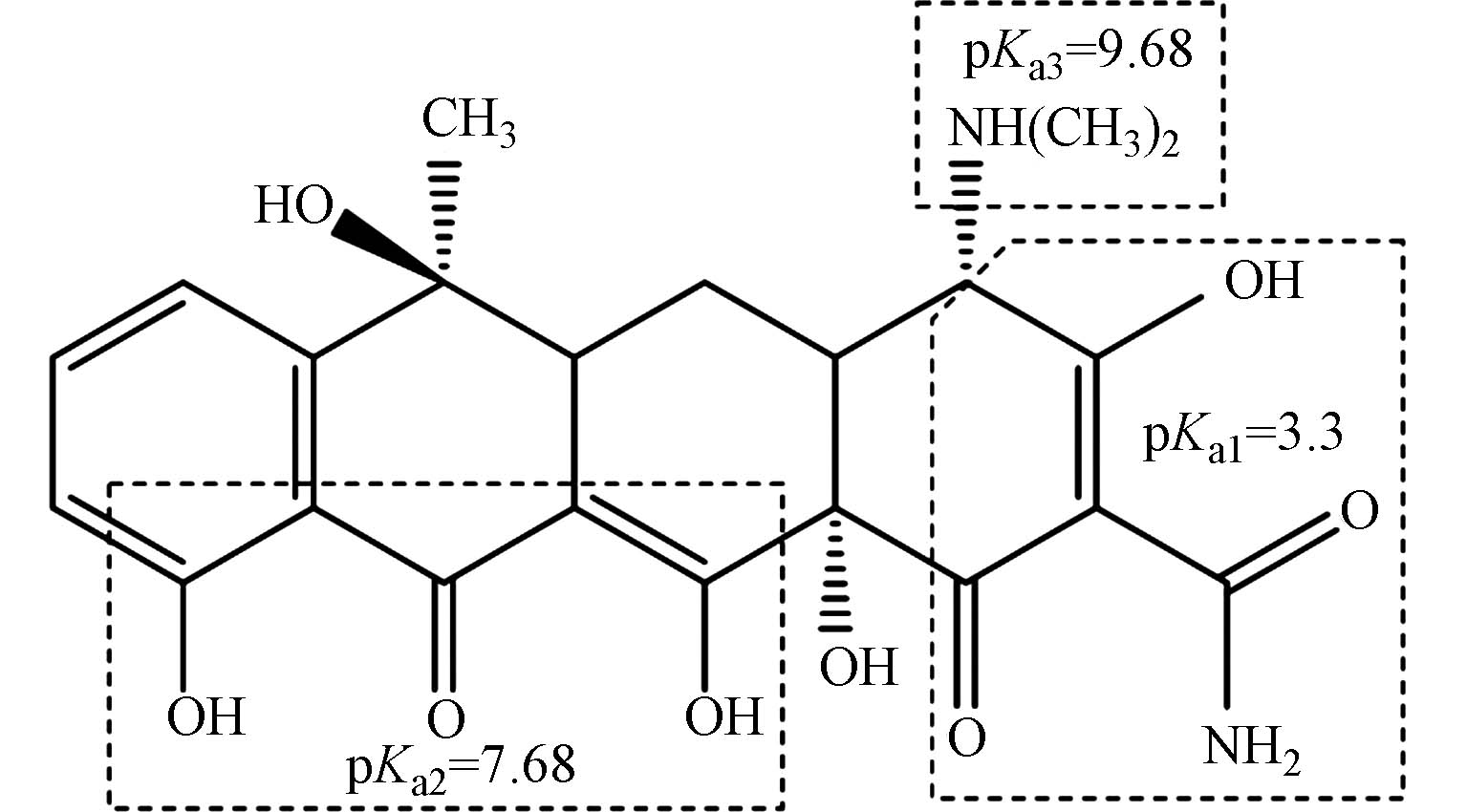 | Show Table
| Show Table DownLoad:
CSV
DownLoad:
CSV
1.2 DBC的表征
总有机碳分析仪(TOC-Vcpn,Shimadzu,日本)和pH计(AB150,Thermo Fisher Scientific,美国)测定DBC的总有机碳(TOC)含量和pH. 紫外可见分光光度计(UV-9000S,上海元析,中国)测定DBC紫外-可见吸收光谱,扫描波长为100—900 nm. 荧光分光光度计(F-4600,日立,日本)分析DBC的荧光特性,测定发射波长(Em)为200—550 nm,激发波长(Ex)为220—450 nm. 傅里叶变换红外光谱仪(FTIR,Spectrum Two,PerkinElmer,英国)测定DBC的官能团,扫描区域为4000—500 cm−1,分辨率 4 cm−1 .
1.3 DBC对TC的光降解实验
1.3.1 不同种类DBC对TC光降解影响
在模拟太阳光照射下,于100 mL石英管(含20 mL溶液)中进行DBC暴露下的TC光解实验[27]. 以测得TOC作为DBC原始质量浓度(表2),配置100 mg·L−1(以C的质量计)的DBC储备液. 向石英管中分别加入0.02、0.2、2 mL的DBC储备液和1 mL的TC储备液(400 mg·L−1),用超纯水定容至20 mL. 将石英管放在于以汞灯为光源的光化学反应仪(BL-GHX-V,上海比朗,中国)中,波长为290—400 nm,光功率密度为30 W·m−2,石英管以25 r·min−1匀速旋转. 反应启动后,分别在0、10、20、40、60 min时取600 µL反应溶液,过0.22 µm聚醚砜滤膜后用高效液相色谱仪(HPLC 1260,安捷伦,美国)对TC定量分析. HPLC的检测条件为:C18反向色谱柱(3.5 μm,4.6 mm×250 mm),紫外检测器,波长设为355 nm,柱温为30 ℃,流动相为1:3的乙腈和草酸(0.01 mol·L−1)溶液,流速为 1 mL·min−1,进样量为20 μL.
表 2 DBC的基本物理化学性质Table 2. Basic physicochemical properties of DBCDBC TOC/(mg·L−1) pH SUVA254/L·(mgC·m-1)−1 E2/E3 E4/E6 LW300 123 4.25 0.18 3.92 7.17 LW400 152 4.74 0.12 5.01 5.82 LW500 98.9 5.96 0.16 3.67 3.88 LZ300 520 5.66 0.06 6.23 5.48 LZ400 832 5.77 0.02 7.28 15.8 LZ500 451 7.75 0.17 3.81 5.09 LZ600 101 10.2 0.27 4.61 2.00 注:SUVA254为100倍的254 nm处的吸光度和DBC TOC的比值,E4/E6为465 nm处吸光度和665 nm处吸光度的比值,E2/E3为254 nm处吸光度和365 nm处吸光度的比值. Note: SUVA254 is the ratio of 100 times the absorbance at 254 nm to DBC TOC, E4/E6 is the ratio of the absorbance at 465 nm to 665 nm, E2/E3 is the ratio of the absorbance at 254 nm to 365 nm. | Show TableDownLoad:
CSV
1.3.2 TC初始浓度对光解的影响
向含有2 mL DBC储备液的石英管中分别加入0、0.25、0.5、1、2 mL的TC储备液,用超纯水定容至20 mL,按照1.3.1节中条件光解、检测.
1.3.3 常见离子对TC降解影响
选择水环境中常见的阴离子(NO3−、HCO3−、SO42-、Cl−)和阳离子(Fe3+、Ca2+)进行光解实验[27]. 向含有2 mL DBC储备液和1 mL TC储备液的石英管中分别加入0、0.002、0.02、0.2、2 mL的离子储备液(100 mg·L−1),用超纯水定容至20 mL,按照1.3.1节中条件光解、检测.
1.3.4 初始pH对TC光解的影响
使用0.1 mol·L−1的HCl和NaOH将含有DBC(10 mg·L−1)和TC(20 mg·L−1)混合溶液的pH分别调整为3、5、7、9和11. 按照1.3.1节中条件光解、检测.
1.4 活性中间体对TC光解的影响
为确定DBC产生的不同RIS在TC光降解中的作用,选择常用的猝灭剂异丙醇(IPA)、2,4,6-三甲基酚(TMP)和叠氮化钠(NaN3)分别验证·OH、3DBC*以及·OH和1O2在光降解过程中的作用[10, 12]. 向含有2 mL DBC储备液和1 mL TC储备液的石英管中分别添加20 μL IPA、2 mL 80 mg·L−1的NaN3和2 mL 0.545 g·L−1的TMP,用超纯水定容至20 mL,按照1.3.1节中条件光解、检测.
1.5 数据分析
采用Excel 2016和Origin 2018进行实验结果的处理分析及图表绘制. 使用SPSS 22.0进行单因素方差分析(ANOVA),结合Duncan 检验(P < 0.05)进行显著性差异分析. 利用Origin 2018绘制三维荧光光谱图并对荧光区域积分进行定量分析. 用二级动力学模型(1/C-1/C0=kt)模拟TC光降解动力学[13],其中C和C0分别是t时刻和0时刻的TC浓度,k是二级速率常数.
2. 结果与讨论(Results and discussion)
2.1 DBC的理化性质及结构特征
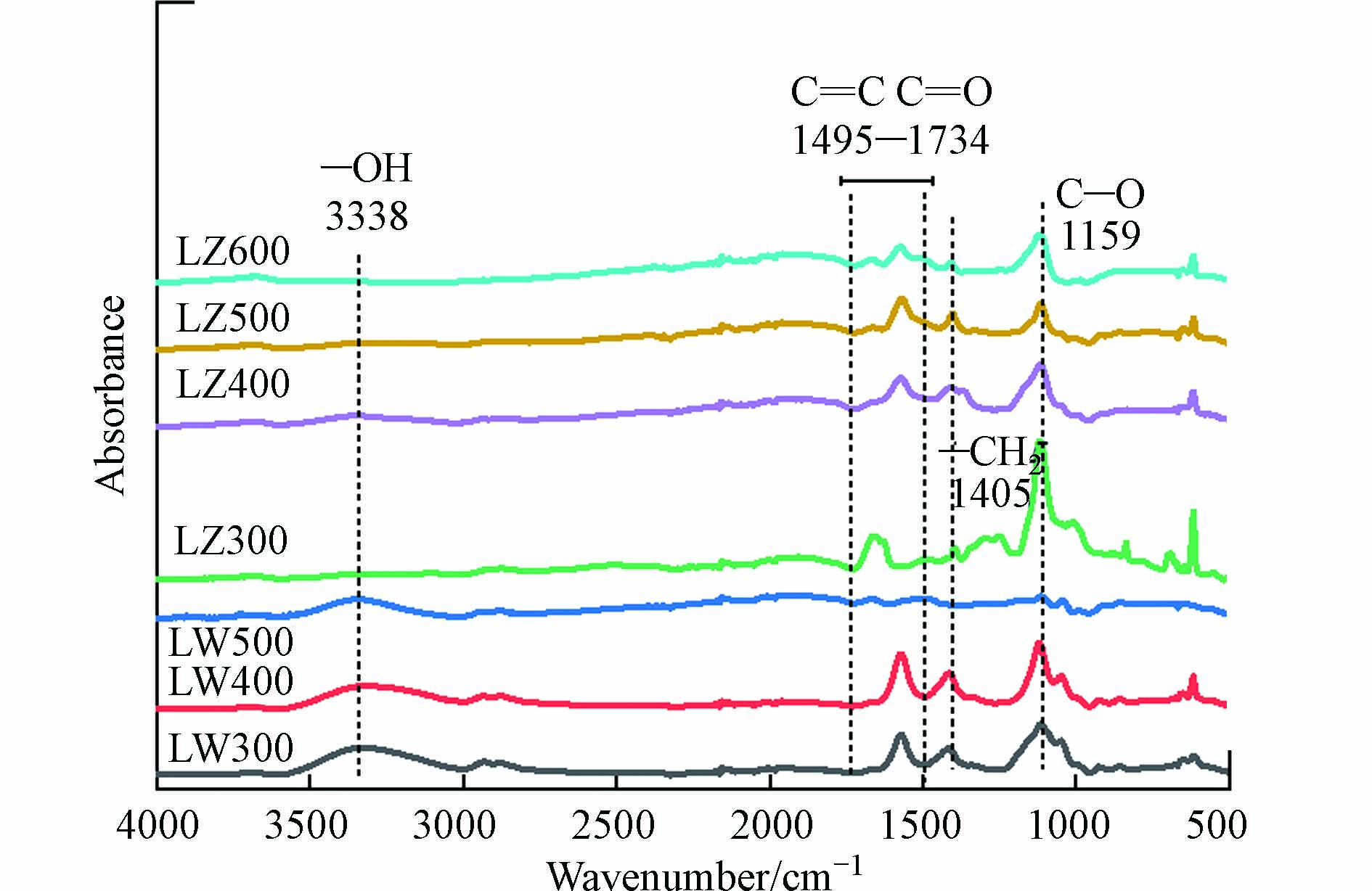
DBC的物理化学性质如表2所示. 随着HTT升高,芦苇和芦竹DBC的TOC含量均呈先升高后降低的趋势. 这是由于在较低HTT下(300—400 ℃),BC炭化程度低,溶解性物质含量较高[3, 30];当HTT升至500 ℃时,纤维素和木质素进一步分解聚合,产生更多稳定性较强的芳烃类物质[3]. 另外,芦竹DBC的TOC含量(101—832 mg·L−1)高于芦苇DBC(98.9—152 mg·L−1),这可能是芦竹生物质较芦苇含有更多的纤维素和半纤维素[31],导致相同HTT的芦竹BC稳定性较低,含有更多的可溶性物质. 随着HTT升高,芦苇和芦竹DBC的pH分别由4.25和5.66上升至5.96和10.2,这是因为HTT升高导致BC中矿物质富集,且矿物质的溶解使得DBC的溶液的pH增加[3]. E4/E6是有机质的芳香性指标,与芳香性呈反比[12]. 随着HTT的升高,芦苇和芦竹DBC的E4/E6由7.17和5.48降低至3.88和2.00(除LZ400外),说明高HTT的DBC芳香性更高. 这是因为BC形成时,生物质发生脱水和裂解反应导致不稳定有机物分解,而聚合反应增加了芳香性物质的含量[5]. 值得注意的是,LZ400的E4/E6大幅升高,这可能是因为纤维素分解为酸醛等可溶性高的物质[32],导致芳香性组分在水相总有机质中占比下降. SUVA254是有机质的芳香性指标,与芳香性呈正比[12]. 随着HTT升高SUVA254先降低后升高,这表明芳香性在400 ℃时降低,但高温DBC芳香性更高这与E4/E6的结果一致. 相同HTT下,芦苇DBC(0.12—0.18 L·(mg·C·m-1)−1)比芦竹DBC(0.02—0.17 L·(mg·C·m-1)−1)的芳香性更强,这可能是由于芦苇生物质的木质素含量高,产生的BC芳香性物质更多[31]. E2/E3是有机质分子量大小的指标,与其分子量呈反比[12]. 两类DBC的E2/E3随HTT增加先升高后降低,表明DBC分子量先降低后上升,这与芦苇和芦竹热解形成BC的过程中脂肪烃分解和芳香烃的形成有关[30]. 另外,相同HTT下,芦竹DBC的E2/E3比值(3.81—7.28)高于芦苇DBC(3.67—5.01),表明芦竹DBC平均分子量更小,这与TOC的含量变化规律一致,表明小分子量物质可能更易从BC溶出,形成DBC[33].
DBC的FTIR图谱如图1所示. 对于芦竹DBC,其在1495—1734 cm−1(C=C、C=O)、1405 cm−1(—CH2—)和1159 cm−1(C—O)处有明显的吸收峰[34].
随着HTT的升高,C—O的峰大幅度降低,—CH2—的峰也变小,表明不稳定的脂肪族化合物逐渐被分解,而C=C和C=O虽有小幅度降低,但仍有较大的振动峰存在,表明芦竹DBC仍具有较高的芳香性组分[34]. 对于芦苇DBC,在3338 cm−1(酚羟基,—OH)[29, 35]、1495—1734 cm−1(C=C、C=O)、1405 cm−1(—CH2—)和1159 cm−1(C—O)处有明显的吸收峰. 随着HTT升高,—CH2—和C—O的峰逐渐消失,表明不稳定的脂肪烃逐渐分解,C=C和C=O的峰虽也消失,但酚—OH的峰却无明显变化,说明芦苇DBC中的芳香性组分虽有分解,但仍有较高含量的芳香性组分存在. 总体来说,随着HTT的升高,DBC中脂肪族化合物的含量降低,芳香性组分含量增加,这与DBC的芳香性增加的结果一致(表2).
DBC发色基团的含量对于其光化学反应的发生至关重要[36]. 因此对DBC进行了三维荧光光谱(3D-EEM)分析(图2).
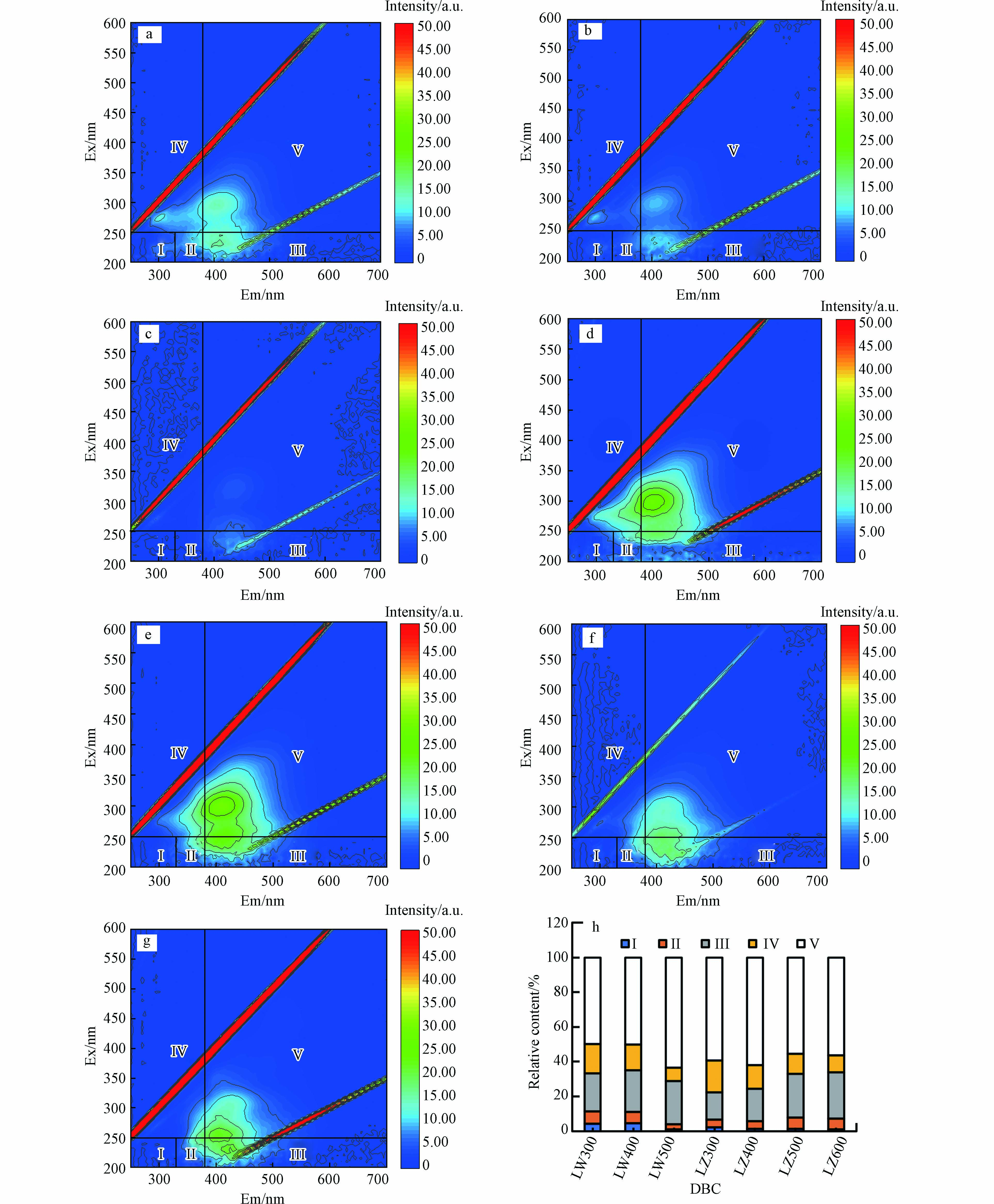 图 2 DBC的荧光组分及相对含量Figure 2. Fluorescence components and their relative contents in DBC(a)LW300、(b)LW400、(c)LW500、(d)LZ300、(e)LZ400、(f)LZ500和(g)LZ600三维荧光谱图;(h)DBC的5种组分的相对含量3D-EEM of (a) LW300, (b) LW400, (c) LW500, (d) LZ300, (e) LZ400, (f) LZ500 and (g) LZ600; (h) The relative contents of DBC five components
图 2 DBC的荧光组分及相对含量Figure 2. Fluorescence components and their relative contents in DBC(a)LW300、(b)LW400、(c)LW500、(d)LZ300、(e)LZ400、(f)LZ500和(g)LZ600三维荧光谱图;(h)DBC的5种组分的相对含量3D-EEM of (a) LW300, (b) LW400, (c) LW500, (d) LZ300, (e) LZ400, (f) LZ500 and (g) LZ600; (h) The relative contents of DBC five componentsLW300、LW400、LZ300、LZ400和LZ500在Ⅲ(Ex:200—250 nm和Em:380—550 nm)、Ⅳ(Ex: 200—250 nm和Em: 330—380 nm)和Ⅴ(Ex: 250—400 nm和Em: 380—550 nm)处有明显的荧光峰,它们分别代表富里酸(fulvic acid, FA)类、溶解性微生物代谢产物和HA类物质. LW500和LZ600仅在Ⅲ和Ⅴ处有荧光峰,表明存在FA类和HA类物质. 所有DBC在Ⅰ(Ex:200—250 nm和Em:280—330 nm)和Ⅱ(Ex:200—250 nm和Em:330—380 nm)区域的荧光峰不明显,表明存在芳香性类蛋白质物质. 这些结果表明FA类和HA类物质是DBC的主要组分. 随HTT升高,芦苇DBC FA和HA相对含量分别由21.9%和49.7%增加至24.9%和63.6%,溶解性微生物代谢产物含量由17.1%降至7.5%,芦竹DBCFA类含量由15.6%增加到26.6%,溶解性微生物代谢产物和HA的含量分别由59.3%和18.2%减少到56.4%和9.7%. FA类和HA类物质的总含量随HTT升高呈上升趋势(71.6%—88.5%). NOM中的类FA和HA类物质是重要的光敏剂,吸收光子后会生成激发三重态HA(3HA*)和FA(3FA*),可通过能量转换或者电子转移降解有机污染物[37]. 因此,含有FA类和HA类组分的DBC也可能促进水体中ATs等有机污染物的光降解,这需要进一步的研究.
2.2 DBC对TC光降解的影响
不同种类DBC对TC光降解的影响如图3所示. 黑暗条件下,DBC对TC浓度无明显影响(图3a),表明非光化学过程对TC降解作用不明显. 光照条件下,DBC不存在时,TC的直接光降解率较低(6.2%),但DBC的存在显著增加了TC的光解率(16.3%—97.0%)(图3b). 用二级动力学方程拟合光解速率k,在不添加DBC时,k为8.3×10−5,添加DBC后,k分别升高为1.7×10−4、1.8×10−4、2.6×10−4、8.3×10−4、8.4×10−4、9.0×10−4和8.8×10−2(图3c),表明DBC促进了TC的光解速率,且高温DBC促进效果更强. 相同HTT条件下,芦竹DBC对k的促进效果高于芦苇DBC. 7种DBC中,LZ600对TC的光解的促进最显著,且LZ600在10 mg·L−1的添加量下,TC光解率最高(图3d). 另外,当TC的初始浓度从5 mg·L−1增加至40 mg·L−1时,TC的光解速率从0.32 L·(mg·min)−1升高至1.45 L·(mg·min)−1,然后降低至0.15 L·(mg·min)−1(R2>0.91). 其中,TC浓度为20 mg·L−1时,其光解速率最快. 因此,选用10 mg·L−1 LZ600和20 mg·L−1的TC进行后续实验. ZHOU等[10]也发现竹子DBC能够将17-β雌二醇的一阶速率常数提高1个数量级,这与上述DBC增加k值的结果一致,说明DBC能够作为一种高效的光敏剂促进TC的光降解.
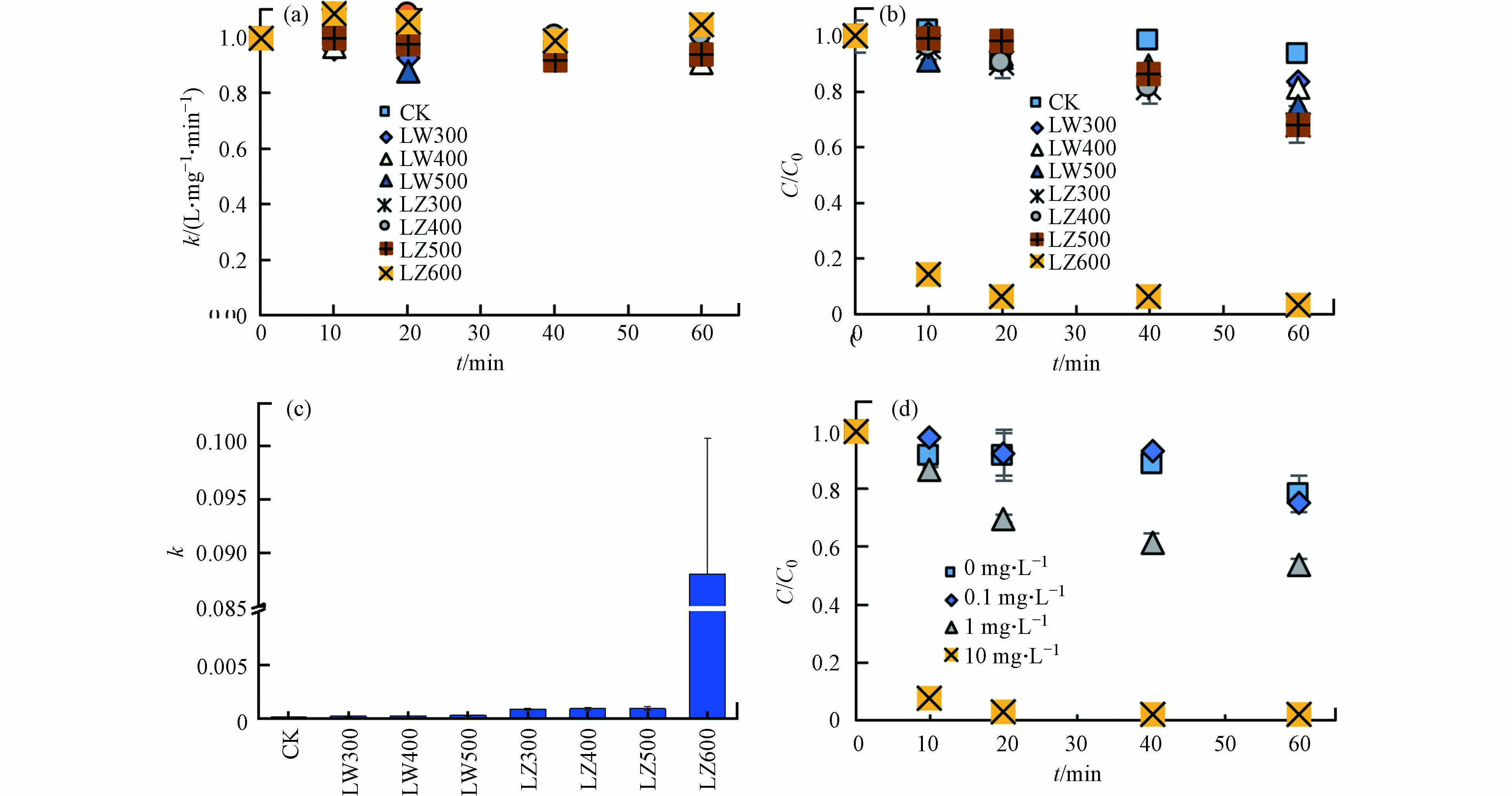 图 3 光照和黑暗条件下不同DBC和不同浓度的LZ600对TC降解的影响Figure 3. Effects of DBC and LZ600 concentrations on TC photodegradation under light and dark conditions光照(a)和黑暗(b)条件下不同DBC对TC降解的影响;(c)不同DBC条件下的TC光解的二阶速率常数(R2>0.93);(d)不同浓度LZ600对TC光解的影响Effect of different DBC on TC degradation under light (a) and dark (b) conditions; (c) Second-order rate constant of TC photolysis under different DBC conditions (R2 > 0.93); (d) Effects of different concentrations of LZ600 on TC photolysis
图 3 光照和黑暗条件下不同DBC和不同浓度的LZ600对TC降解的影响Figure 3. Effects of DBC and LZ600 concentrations on TC photodegradation under light and dark conditions光照(a)和黑暗(b)条件下不同DBC对TC降解的影响;(c)不同DBC条件下的TC光解的二阶速率常数(R2>0.93);(d)不同浓度LZ600对TC光解的影响Effect of different DBC on TC degradation under light (a) and dark (b) conditions; (c) Second-order rate constant of TC photolysis under different DBC conditions (R2 > 0.93); (d) Effects of different concentrations of LZ600 on TC photolysis2.3 环境条件对LZ600介导的TC光降解的影响
实际水环境中存在多种阴离子(如NO3−、HCO3−、SO42−、Cl−)和阳离子(如Fe3+、Ca2+),可能影响DBC的光敏性及污染物的光转化过程[38-43]. LZ600存在的条件下,与不添加离子的对照组相比(99.6%),当溶液中存在NO3−、HCO3−、SO42−、Cl−、Fe3+和Ca2+时,TC的光降解率降为76.0%—80.0%、76.3%—81.0%、71.5%—77.4%、66.8%—69.0%、16.0%—97.3%和1.2%—54.8%(图4a—g).
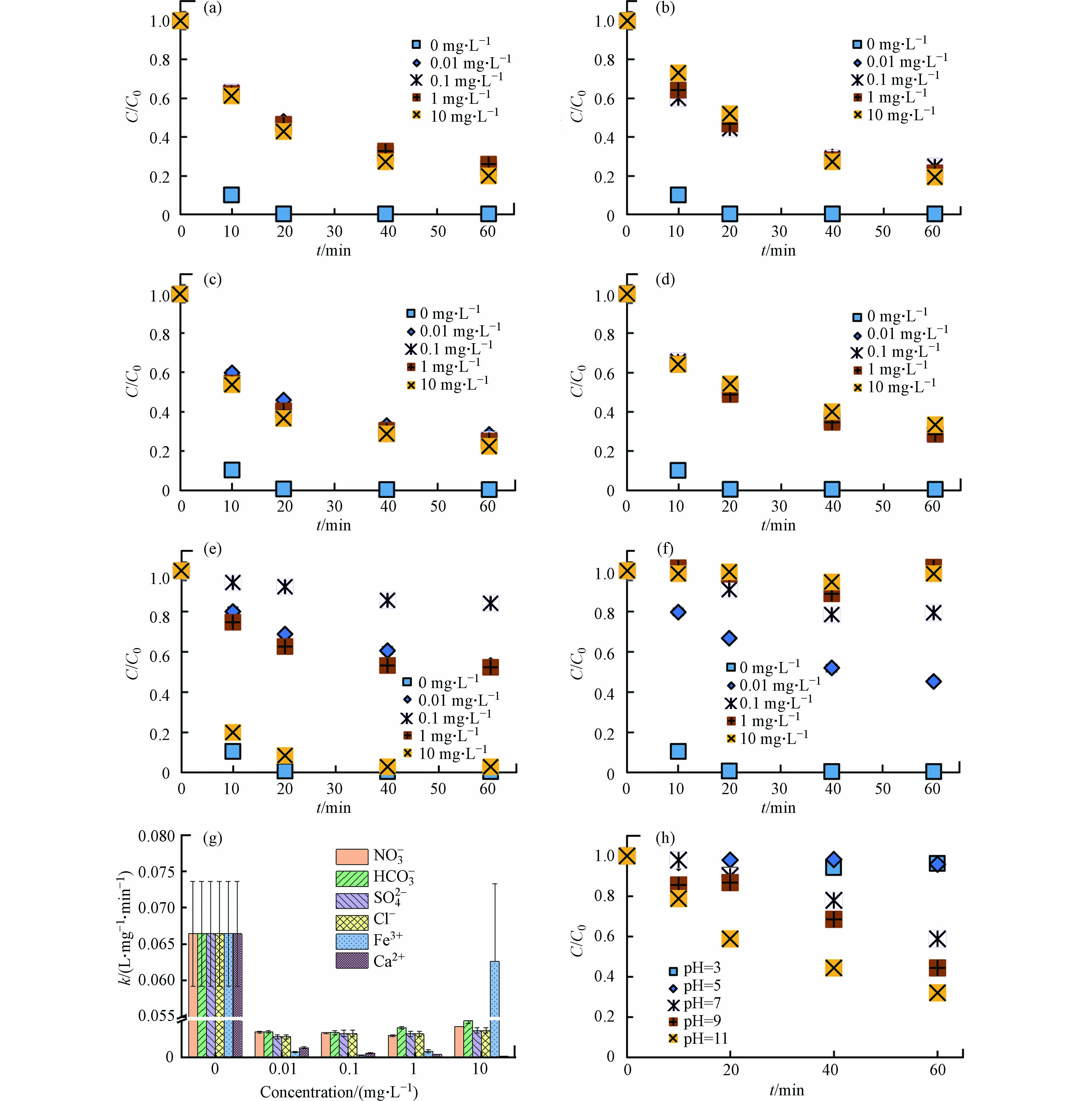 图 4 不同离子和pH条件下LZ600对TC光降解的影响Figure 4. Effect of LZ600 on TC photodegradation under different ambient ions and pH(a)NO3-、(b)HCO3-、(c)SO42-、(d)Cl-、(e)Fe3+和(f)Ca2+的不同浓度梯度下DBC对TC的光降解的影响;(g)不同离子条件下TC光解的二阶速率常数(R2>0.94);(h)LZ600存在下,不同pH对TC光降解的影响The effect of DBC on TC photodegradation under different concentration gradients of (a) NO3- (b) HCO3- (c) SO42- (d) Cl- (e) Fe3+ and (f) Ca2+; (g) Second-order rate constant of TC photolysis under different ionic conditions (R2 > 0.94); (h) Effect of different pH on TC photodegradation in the presence of LZ600
图 4 不同离子和pH条件下LZ600对TC光降解的影响Figure 4. Effect of LZ600 on TC photodegradation under different ambient ions and pH(a)NO3-、(b)HCO3-、(c)SO42-、(d)Cl-、(e)Fe3+和(f)Ca2+的不同浓度梯度下DBC对TC的光降解的影响;(g)不同离子条件下TC光解的二阶速率常数(R2>0.94);(h)LZ600存在下,不同pH对TC光降解的影响The effect of DBC on TC photodegradation under different concentration gradients of (a) NO3- (b) HCO3- (c) SO42- (d) Cl- (e) Fe3+ and (f) Ca2+; (g) Second-order rate constant of TC photolysis under different ionic conditions (R2 > 0.94); (h) Effect of different pH on TC photodegradation in the presence of LZ600另外,不添加离子时,k为6.6×10−2 L·(mg·min)−1,添加离子后,k降为3.4×10−3—3.9×10−3、3.4×10−3—4.2×10−3、3.5×10−3—3.7×10−3、2.2×10−3—2.7×10−3、2.5×10−4—2.6×10−2和9.8×10−4—1.3×10−3 L·(mg·min)−1 (图4g). 结果表明,环境中常见的离子对DBC介导的TC光降解有一定抑制效果,抑制效果受离子种类和浓度的影响. ZHANG等[27]研究表明HCO3−和Cl−能够与DBC产生的RIS反应,生成活性小于RIS的次级活性中间体,抑制抗生素抗性基因的光解. MACK和ZHOU等[40-41]发现,NO3−和SO42−可以抑制DBC对光子的吸收而抑制DBC光解. OU等[42]发现,Ca2+能够与DBC形成内球配合体,促进DBC沉降进而抑制其光反应. Fe3+在0.01—0.1 mg·L−1添加量下显著抑制DBC的光解,这是由于Fe3+与DBC竞争光子抑制了RIS的生成[43]. 但当添加量为1—10 mg·L−1时,Fe3+对DBC光解的抑制效果被缓解,这可能是因为Fe3+与水分子和DBC产生了光敏剂Fe(OH)2+和Fe-DBC剂促进了RIS的产生[43-44]. 简而言之,离子通过影响DBC对光子的吸收以及RIS的活性影响TC的光降解.
pH会影响DBC的表面活性和TC的电子密度[42, 45],因此pH可能会对TC的降解产生影响. 不同pH(3、5、7、9和11)对DBC影响的TC光降解如图4h. 随着pH增加,TC光解率从3.7%增加到67.9%,k从5.0×10−5增加到2.3×10−3 L·(mg·min)−1. 这表明碱性条件下,LZ600更有助于促进TC的光降解. 这与李聪鹤等[46]研究一致,在pH为11的条件下,溶解性有机质对磺胺甲恶唑的降解效果最显著. 这可能是由于pH的增加有助于增加DBC的稳定性[42]及促进RIS的产生[46],同时高pH条件下,TC去质子化程度增大,电子密度增加,更易受到RIS的攻击[46-47].
2.4 DBC影响TC降解的可能机制
DBC是水环境重要的光敏剂,会在光照下发生化学转化[15]. 为探究光解前后LZ600结构组成的变化,测定了光降解后LZ600组成及含量(图5a). 与原始LZ600相比(图2g),光照后LZ600组分Ⅰ—Ⅴ的荧光强度显著降低(14.1%—35.0%),其中FA类和HA类对荧光强度变化的贡献最大(27.1%和54.2%),表明HA类和FA类物质在光照下发生了明显的转化. 先前的研究表明,FA和HA的添加能够产生大量的RIS,将ATs的降解速率提高34%[13]. 因此,DBC导致的TC光解的增强可能归因于FA类和HA类物质的光转化而产生的强氧化性的RIS [15].
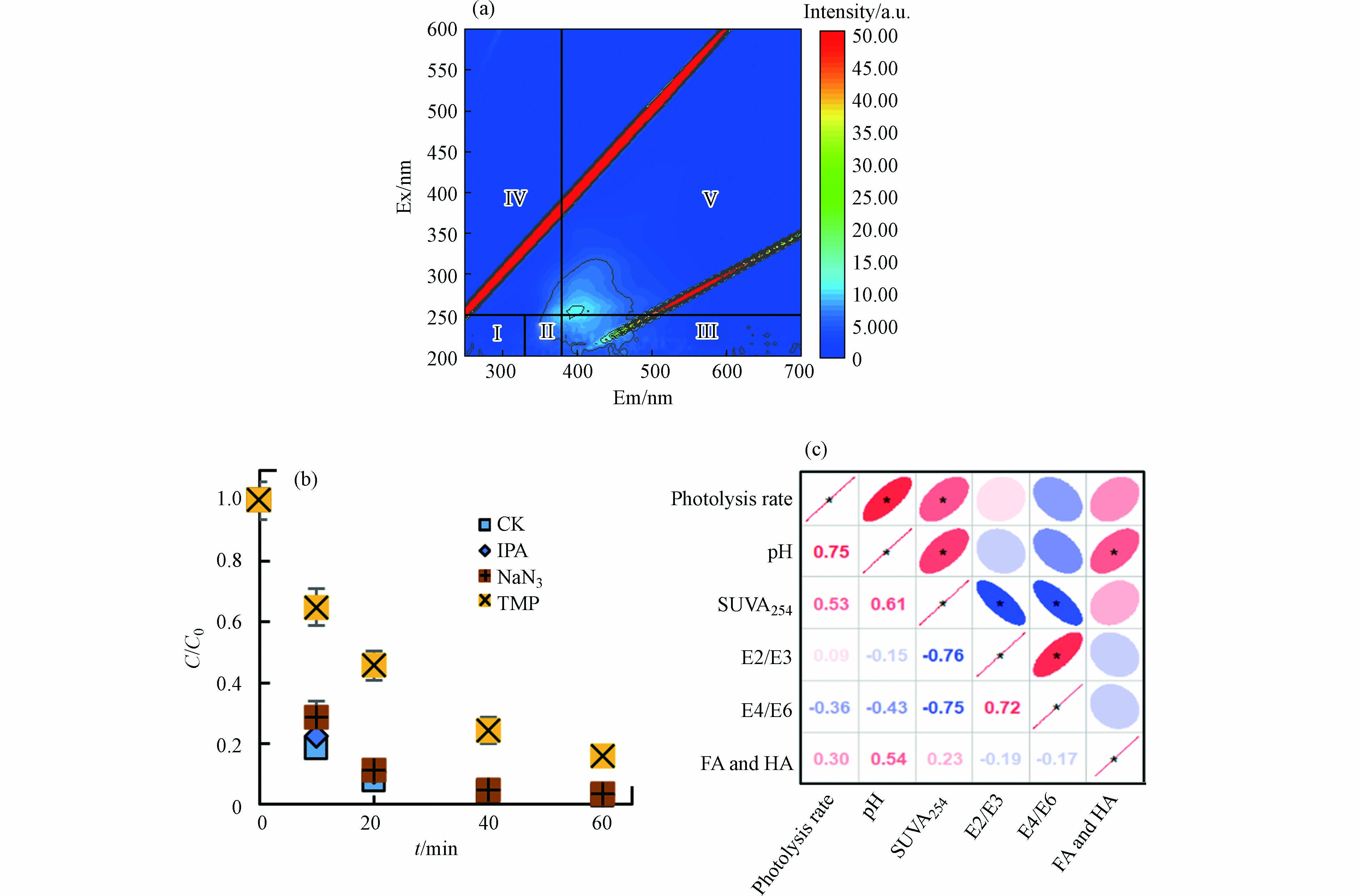 图 5 光降解对DBC性质及猝灭剂对TC光解的影响Figure 5. Effect of photolysis on LZ600 and quenching experiment(a)LZ600光降解后的三维荧光光谱;(b)活性中间体猝灭剂存在下DBC对TC光降解的影响;(c)TC光降解率的相关性分析(红色为正相关,蓝色为负相关)(a) 3D-EEM of LZ600 after photodegradation; (b) The effect of DBC on TC photodegradation in the presence of active intermediate quencher; (c) Correlation analysis of TC photodegradation rate (red is positive correlation, blue is negative correlation)
图 5 光降解对DBC性质及猝灭剂对TC光解的影响Figure 5. Effect of photolysis on LZ600 and quenching experiment(a)LZ600光降解后的三维荧光光谱;(b)活性中间体猝灭剂存在下DBC对TC光降解的影响;(c)TC光降解率的相关性分析(红色为正相关,蓝色为负相关)(a) 3D-EEM of LZ600 after photodegradation; (b) The effect of DBC on TC photodegradation in the presence of active intermediate quencher; (c) Correlation analysis of TC photodegradation rate (red is positive correlation, blue is negative correlation)为了进一步探究DBC在光照下产生的不同RIS(·OH、1O2和3DBC*)在光解中的作用,进行了RIS猝灭实验. 如图5b所示,添加TMP(3DBC*猝灭剂)后,DBC对TC的光解率从97.0%降至84.0%,NaN3(·OH和1O2猝灭剂)和IPA(·OH猝灭剂)的加入对TC光解影响较小. 这说明TC的光降解的增强主要归因于3DBC*. 另外,TMP使得k从1.1×10−2 L·( mg·min) −1降至1.1×10−3 L·(mg·min)−1,IPA使k从1.1×10−2 L·( mg·min) −1降至8.9×103 L·(mg·min)−1,NaN3使k从1.1×10−2 L·( mg·min) −1降至6.8×103 L·(mg·min)−1,NaN3与IPA的k差值为2.1×10−3 L·(mg·min)−1. 这进一步说明DBC对TC的降解由3DBC*起主导作用,·OH和1O2也促进了光解,但贡献较小[10].
不同HTT下DBC的芳香性、分子量及FA和HA类物质的总含量均不同,为探究这些性质是否会影响DBC对TC光解的效果,进行了TC光解率与DBC的SUVA254、E2/E3、E4/E6、pH和HA和FA总含量的相关性分析(图5c). 结果表明TC光解率与pH呈显著正相关,说明pH很大程度上影响了DBC对TC的光解,可能是因为低pH时,DBC表面会形成致密的层,不易发生光反应[42];高pH时,DBC中HA和FA产生的小分子酸更容易被消耗,使化学平衡不断向RIS生成方向进行,增加了RIS含量[48]. 除pH外,DBC的芳香性(SUV254)也与TC的光解率呈显著正相关,这是因为芳香性物质是DBC吸收光子的主要组分,芳香性的升高会增加DBC光子的吸收进而促进RIS的产生[15]. 除pH和芳香性外,HA和FA含量与TC的光解率也呈正相关,也可能会影响DBC对TC的光解效果,但不是主要原因,DBC的分子量与光解率的相关性较低,对TC的光降解影响较小. 结果进一步说明,DBC促进TC的光降解主要是由于DBC具有的稠环芳烃类结构导致的RIS(3DBC*)的产生引起的(图6).
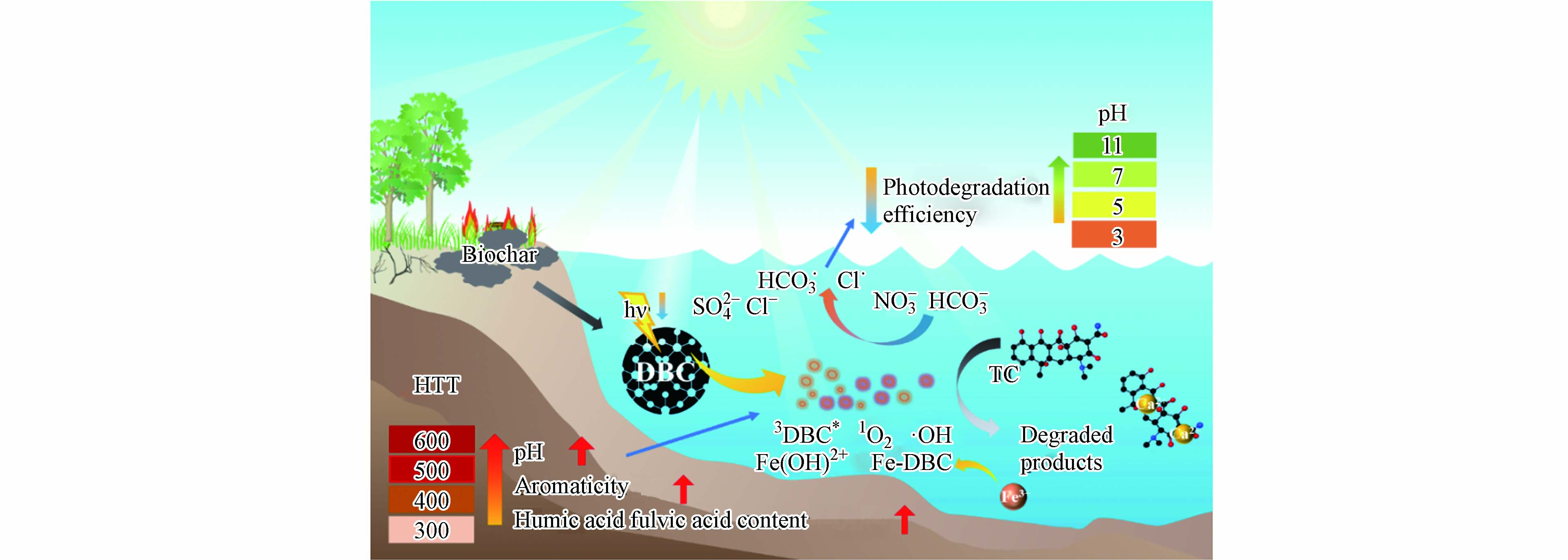 图 6 DBC影响TC降解的可能机制Figure 6. Potential mechanisms of DBC promoting TC photodegradation
图 6 DBC影响TC降解的可能机制Figure 6. Potential mechanisms of DBC promoting TC photodegradation3. 结论(Conclusion)
本研究针对不同热解温度和生物质类型的DBC对水环境中ATs光降解影响效果不明确的问题,重点评估了不同原料和热解温度条件下制备的DBC的理化性质及结构特征,探究了不同制备条件的DBC对ATs光降解的影响和机制及关键水环境条件对ATs光降解的影响. 研究发现:
(1)随着热解温度升高,DBC的TOC先上升后下降,分子量先降低后上升,芳香性逐渐增强,类FA和类HA相对含量增加.
(2)DBC对TC的光降解均有促进作用. 随着热解温度升高,芦苇和芦竹DBC对TC光降解的促进作用增加,在相同热解温度下,芦竹DBC对TC光降解的促进效果较芦苇DBC更高.
(3)水中常见阴离子(NO3−、HCO3−、SO42−、Cl−)和阳离子(Fe3+、Ca2+)均对TC的光降解有抑制作用,抑制效果受离子种类和浓度的影响. DBC在碱性条件下对TC光降解的促进作用较酸性条件更高.
(4)TC的光降解主要由DBC中的富里酸和腐殖酸类物质光转化产生的·OH、1O2和3DBC*等活性中间体诱发,其中TC的光降解由3DBC*主导,·OH、1O2对TC光降解的贡献较小.
-
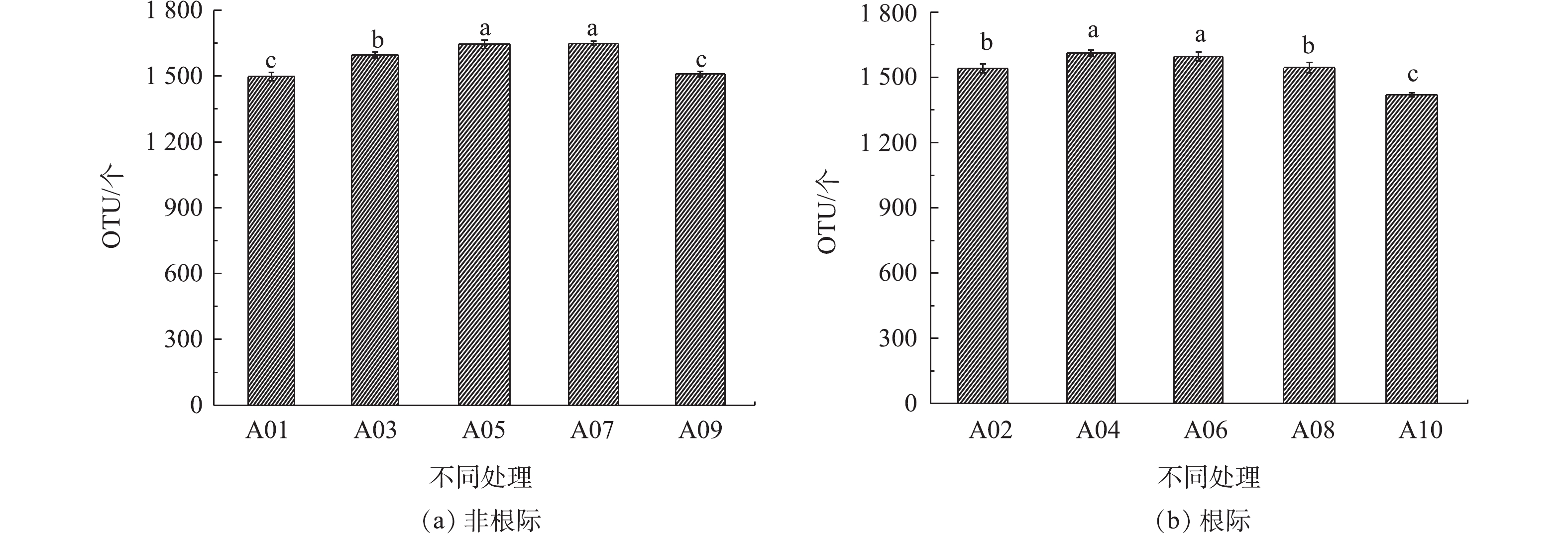
图 1 污泥施用对非根际和根际土壤OTU丰度影响
Figure 1. Effects of sludge application on OTU abundance in non-rhizosphere and rhizosphere soil
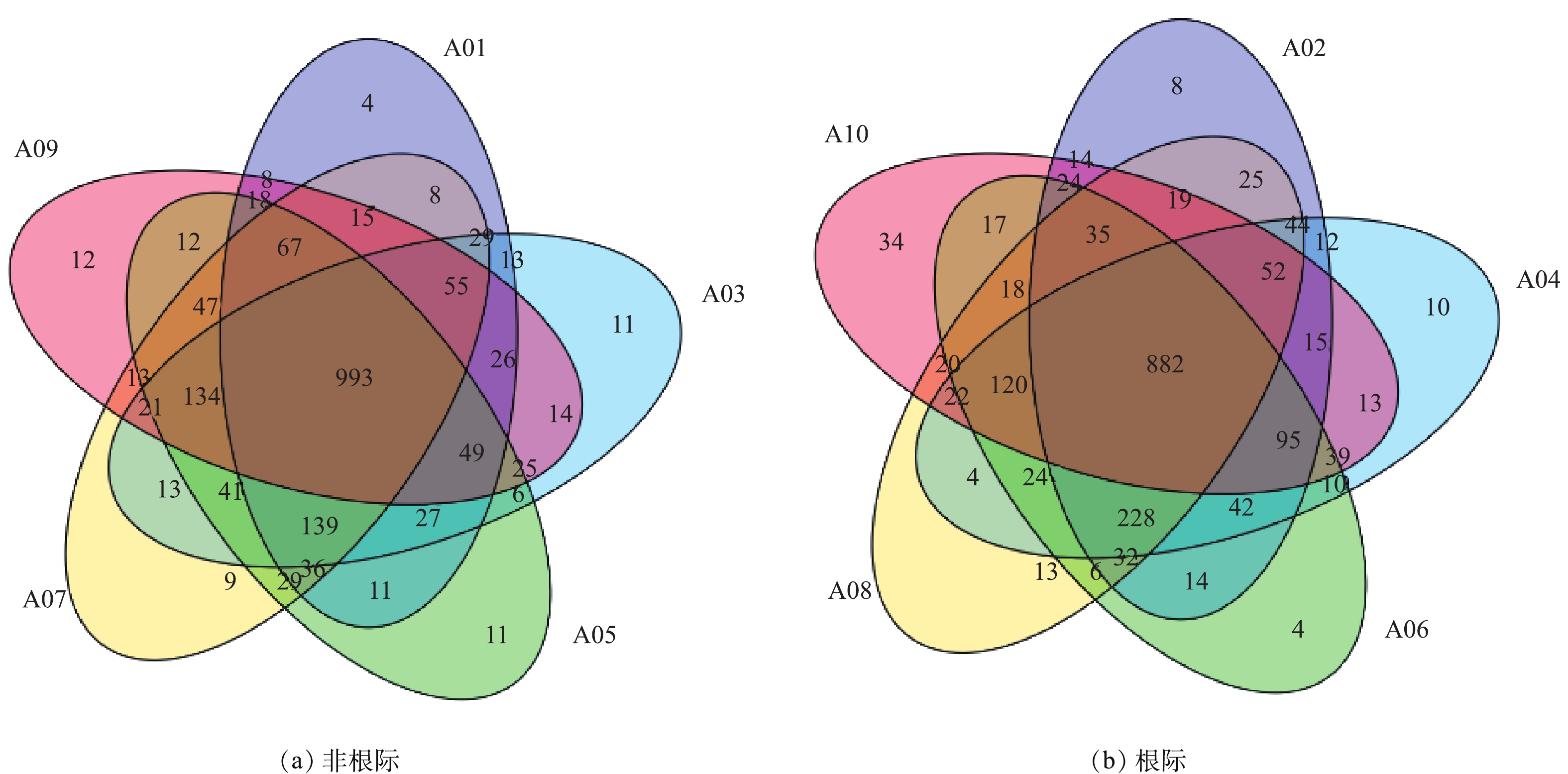
图 2 污泥施用后非根际和根际土壤细菌的Venn图
Figure 2. Venn diagram of bacteria in non-rhizosphere and rhizosphere soil after sludge application
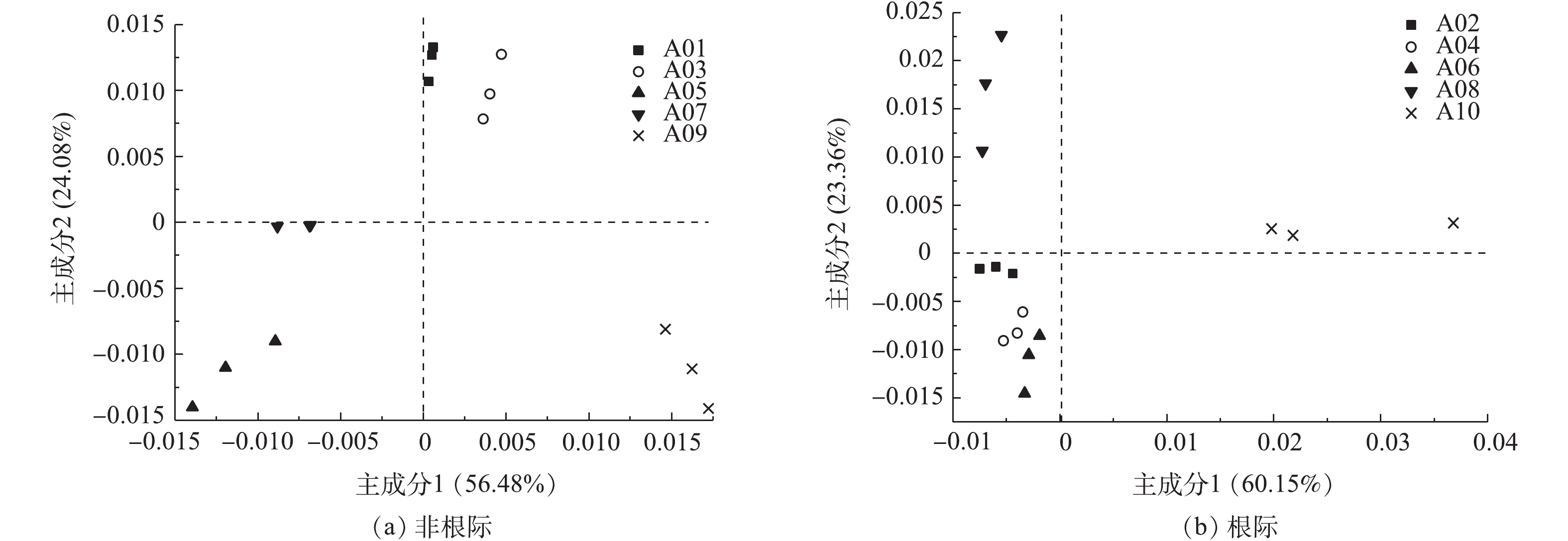
图 3 非根际和根际土壤细菌的主成分分析
Figure 3. Principal component analysis of bacteria in non-rhizosphere and rhizosphere soil
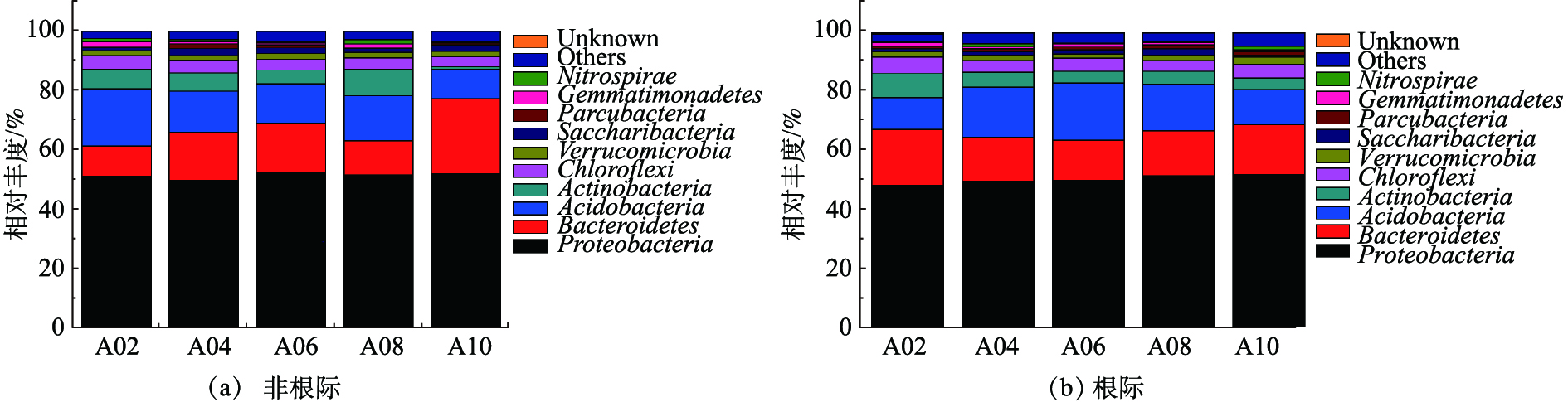
图 4 门水平上非根际和根际土壤细菌的相对丰度
Figure 4. Relative abundance of bacteria in non-rhizosphere and rhizosphere soil at phylum level
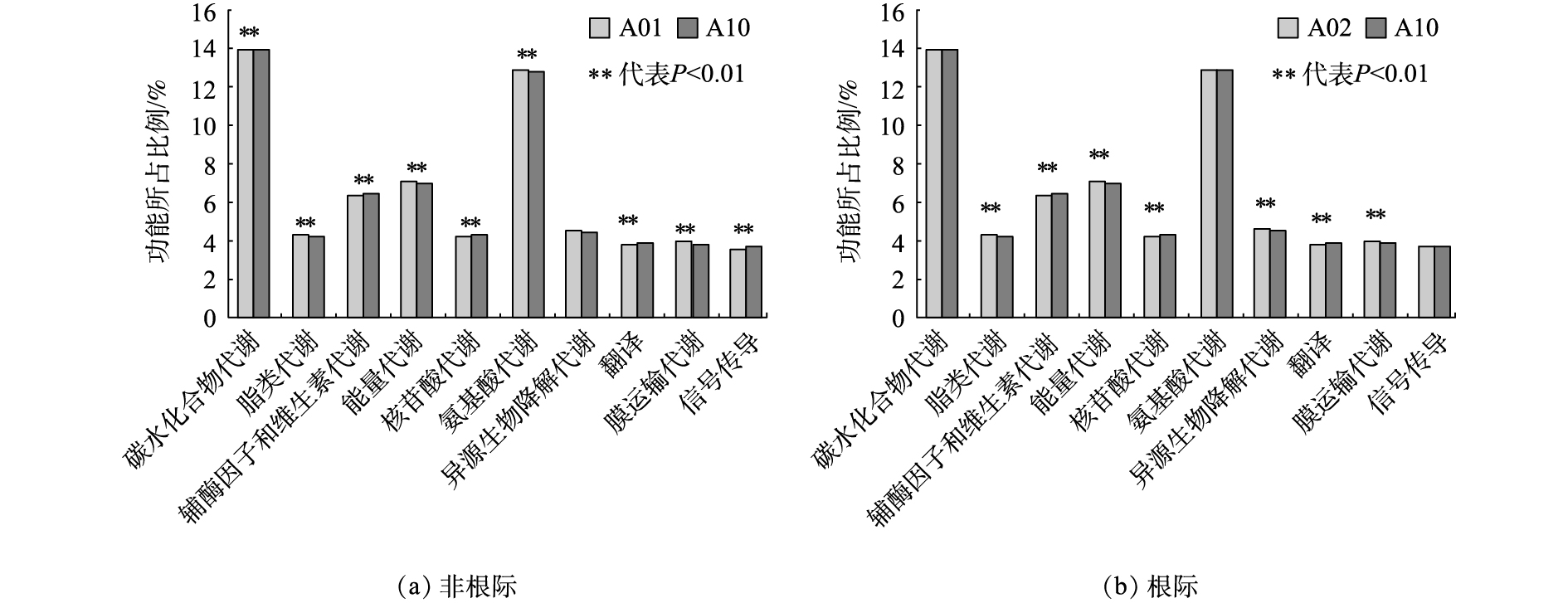
图 5 非根际和根际土壤细菌KEGG功能分析
Figure 5. Analysis of KEGG function of bacteria in non-rhizosphere and rhizosphere soil
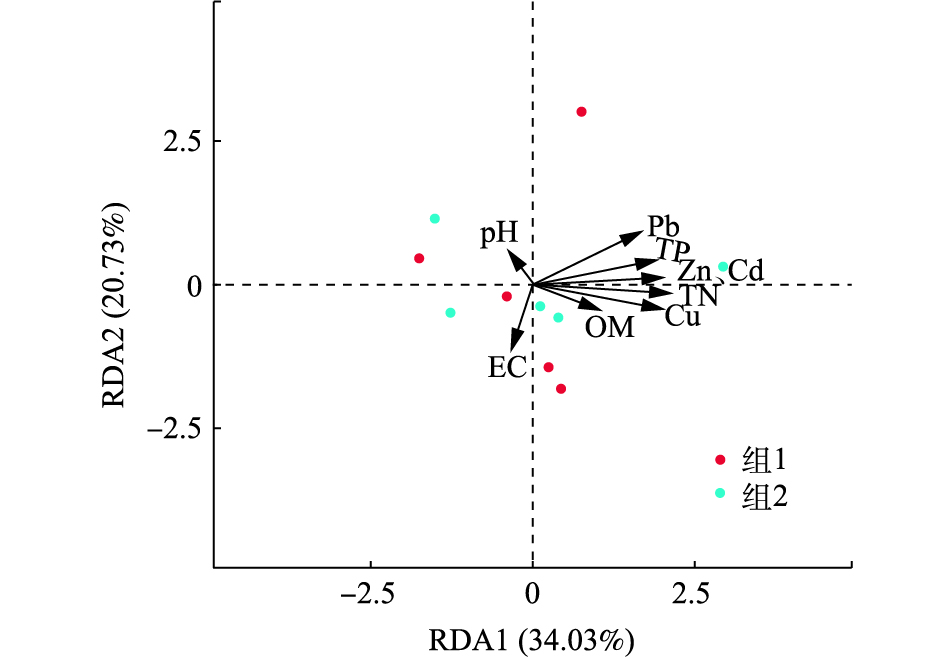
图 6 土壤细菌群落与土壤性质的RDA分析
Figure 6. RDA analysis of soil bacterial community and soil properties
表 1 土壤和腐熟污泥基本理化性质
Table 1. Basic physic-chemical properties of soil and compost sludge
供试材料 有机质/% 总氮/(g·kg−1) 总磷/(g·kg−1) 总钾/(g·kg−1) pH 碱解氮/(g·kg−1) 有效磷/(g·kg−1) 有效钾/(g·kg−1) 土壤 0.54±0.03 0.48±0.03 0.78±0.04 1.94±0.19 8.01±0.03 0.032±0.002 0.015±0.001 0.031±0.002 污泥 41.64±3.55 20.23±2.67 10.21±0.60 2.21±0.04 8.04±0.02 2.047±0.01 0.058±0.002 0.110±0.003 注:数据为平均数和标准差分析。
下载: 导出CSV
表 2 污泥和土壤中重金属的含量
Table 2. Heavy metals content of soil and compost sludge
供试材料 Pb Zn Cu Cr Cd As Hg 土壤 2.35±0.36 53.98±2.46 52.79±1.89 76.43±0.82 0.49±0.01 28.05±1.12 0.029±0.001 污泥 5.53±0.23 269.08±9.33 206.39±9.01 173.99±10.5 1.19±0.16 48.08±2.23 0.035±0.001 注:数据为平均数和标准差分析。
下载: 导出CSV
表 3 污泥施用对非根际和根际土壤各参数影响
Table 3. Effects of sludge application on parameters of non-rhizosphere and rhizosphere soil
参数 非根际土壤 根际土壤 A01 A03 A05 A07 A09 A02 A04 A06 A08 A10 pH 8.18±0.02a 8.16±0.01a 8.13±0.01a 8.08±0.01b 7.96±0.02c 7.96±0.02a 7.95±0.01a 7.92±0.01ab 7.86±0.01b 7.76±0.02c EC/(μS·cm−1) 69.67±1.12d 74.00±1.00cd 76.67±1.15c 82.48±2.19b 92.33±2.31a 120.33±10.79c 124.67±3.06c 156±13.23b 175.33±13.05b 208.673±12.06a 有机质/(g·kg−1) 5.57±0.37b 5.80±0.02b 6.33±0.20b 9.54±0.41a 10.40±0.54a 8.48±0.32b 8.55±1.01b 10.00±0.69b 12.38±0.59a 14.11±1.36a 总氮/(g·kg−1) 0.49±0.02c 0.52±0.02c 0.64±0.01b 0.66±0.02b 0.83±0.03a 0.36±0.02c 0.40±0.02c 0.52±0.01b 0.55±0.04b 0.72±0.03a 总磷/(g·kg−1) 0.77±0.03d 0.79±0.02cd 0.88±0.04bc 0.90±0.02b 1.02±0.06a 0.64±0.03c 0.66±0.02c 0.73±0.04bc 0.76±0.02b 0.88±0.05a Pb/(mg·kg−1) 5.21±0.17c 5.51±0.12bc 5.67±0.16ab 5.89±0.14ab 6.00±0.16a 4.39±0.22c 4.65±0.14c 4.78±0.20bc 5.10±0.076ab 5.25±0.07a Zn/(mg·kg−1) 53.86±3.35d 57.95±2.97d 68.60±1.85c 83.73±3.26b 96.30±4.57a 50.67±2.7d 55.58±5.53d 65.85±2.28c 80.16±1.44b 93.83±2.61a Cu/(mg·kg−1) 54.19±4.03c 59.31±2.31c 84.03±4.58b 89.20±1.73b 115.77±4.51a 49.05±2.39c 52.96±2.96c 78.35±4.05b 81.72±3.17b 105.24±6.91a Cr/(mg·kg−1) 81.28±2.23c 86.83±1.54bc 91.86±2.66ab 92.15±2.24ab 94.96±4.48a 74.59±2.13b 78.93±1.44b 85.74±2.22a 88.14±1.82a 90.70±2.29a Cd/(mg·kg−1) 0.41±0.01d 0.43±0.01d 0.49±0.02c 0.60±0.02b 0.74±0.03a 0.36±0.01d 0.39±0.01cd 0.45±0.02c 0.56±0.03b 0.69±0.04a As/(mg·kg−1) 28.76±1.22c 32.37±1.65b 35.70±0.75a 36.17±0.39a 38.22±1.31a 33.73±5.83b 34.68±4.95b 41.05±3.76ab 43.42±2.89ab 46.94±1.76a Hg/(mg·kg−1) — — — — — — — — — — 注:数据为平均数和标准差分析,每行根际和非根际样品中不同小写字母表示各处理平均值具有显著差异(P<0.05),“—”表示未检出。
下载: 导出CSV
表 4 污泥施用对石灰性土壤细菌多样性和丰度影响
Table 4. Effects of sludge application on bacterial diversity and abundance in calcareous soil
污泥施用量/(t·hm−2) Shannon Simpson Ace Chao 非根际 根际 非根际 根际 非根际 根际 非根际 根际 0 (6.36±0.07)a (6.41±0.12)a (0.004 3±0.000 1)c (0.004 5±0.000 1)c (1 166.06±15.89)c (1 670.20±15.53)b (1 645.37±17.32)c (1 700.84±21.52)b 3.5 (6.38±0.04)a (6.39±0.07)a (0.005 1±0.000 1)b (0.004 8±0.000 2)c (1 727.57±17.55)b (1 754.11±18.59)a (1 750.75±21.00)b (1 759.19±20.59)a 7.5 (6.36±0.12)a (6.37±0.10)a (0.005 1±0.000 1)b (0.005 7±0.000 2)b (1 789.14±17.93)a (1 721.38±17.62)ab (1 785.98+18.95)ab (1 743.80±10.00)ab 37.5 (6.42±0.09)a (6.51± 0.09)a (0.004 2±0.000 1)c (0.003 2±0.000 1)d (1 825.15±26.45)a (1 758.09±29.94)a (1 822.54±9.85)a (1 779.62±22.39)a 75 (6.15±0.04)b (5.98±0.03)b (0.009 4±0.000 1)a (0.009 8±0.000 1)a (1 657.49±13.80)c (1 599.57±21.53)c (1 657.17±17.23)c (1 637.26±14.00)c 注:数据为平均数和标准差分析,每列中不同小写字母表示各处理平均值的显著性差异 (P<0.05)。
下载: 导出CSV
表 5 土壤参数与微生物多样性指数之间的相关分析
Table 5. Correlation coefficients between soil properties and microbial diversity index
多样性指数 pH 有机质 总氮 总磷 EC Pb Zn Cu Cd Shannon 0.428 −0.457 −0.635* −0.550 −0.343 −0.277 −0.591 −0.630* −0.648* Simpson −0.470 0.469 0.642* 0.550 0.324 0.255 0.601 0.667* 0.654* Ace 0.268 −0.175 −0.081 −0.029 2 −0.265 0.198 −0.052 −0.061 7 −0.134 Chao 0.244 −0.164 −0.183 −0.142 −0.195 0.098 −0.124 −0.149 7 −0.223 注:*显著相关 (P<0.05)。
下载: 导出CSV
-
[1] 余亚伟, 杨雨浛, 张成, 等. 施用污泥堆肥品对土壤和植物总汞及甲基汞的影响[J]. 环境科学, 2017, 38(1): 405-410. [2] 韩鹏, 刘和, 丁春华, 等. 蒸汽爆破对城市污泥预处理和厌氧发酵产酸的影响[J]. 中国环境科学, 2017, 37(1): 238-244. [3] 吴冠男, 谢震宇, 张文妍. 我国城市污泥农用的潜力浅析[J]. 广东化工, 2014, 41(16): 141-142. doi: 10.3969/j.issn.1007-1865.2014.16.078 [4] ZAFARI A, KIANMEHR M H. Factors affecting mechanical properties of biomass pellet from compost[J]. Environmental Technology, 2014, 35(1/2/3/4): 478-486. [5] NOGUEIROL R C, DE MELO W J, BERTONCINI E I, et al. Concentrations of Cu, Fe, Mn, and Zn in tropical soils amended with sewage sludge and composted sewage sludge[J]. Environmental Monitoring and Assessment, 2013, 185(4): 2929-2938. doi: 10.1007/s10661-012-2761-3 [6] ZHAO S L, SHANG X J, DUO L. Accumulation and spatial distribution of Cd, Cr, and Pb in mulberry from municipal solid waste compost following application of EDTA and (NH4)2SO4[J]. Environmental Science and Pollution Research, 2013, 20(2): 967-975. doi: 10.1007/s11356-012-0992-z [7] 魏亮亮, 孔祥娟, 辛明, 等. 国内外污泥处理处置标准指标分析及对我国相关标准研究的建议[J]. 黑龙江大学自然科学学报, 2014, 31(6): 790-800. [8] AMUNDSON R, BERHE A A, HOPMANS J W, et al. Soil and human security in the 21st century[J]. Science, 2015, 348(6235): 647-653. [9] 王海燕, 姚金玲, 武雪芳, 等. 国内外城市污水处理厂污泥标准对比研究与建议[J]. 给水排水, 2010, 36(10): 27-32. doi: 10.3969/j.issn.1002-8471.2010.10.007 [10] SUNDERMEIER A P, ISLAM K R, RAUT Y, et al. Continuous no-till impacts on biophysical carbon sequestration[J]. Soil Science Society of America Journal, 2011, 75(5): 1779-1788. doi: 10.2136/sssaj2010.0334 [11] LI P P, CHENG X, XUE B, et al. Evaluation of composted sewage sludge application to soil[J]. Ieri Procedia, 2013, 5: 202-208. doi: 10.1016/j.ieri.2013.11.093 [12] FLORENCIO I, MARIA J M, MARIA D S, et al. Influence of organic matter transformations on the bioavailability of heavy metals in a sludge based compost[J]. Journal of Environmental Management, 2012, 95(Supplement): 104-109. [13] CESAR N, JENNIFER N K, TERESA H, et al. Soil aggregation in a semiarid soil amended with composted and non-composted sewage sludge: A field experiment[J]. Geoderma, 2014, 219-220: 24-31. doi: 10.1016/j.geoderma.2013.12.017 [14] 张鑫, 党岩, 冯丽娟, 等. 施用城市污泥堆肥对土壤微生物群落结构变化的影响[J]. 环境工程学报, 2014, 8(2): 716-722. [15] LIU X, GUO K L, HUANG L, et al. Responses of absolute and specific enzyme activity to consecutive application of composted sewage sludge in a fluventic ustochrept[J]. Plos One, 2017, 12(5): 1-16. [16] 孙玉焕, 骆永明, 杨志海. 污泥施用对土壤微生物群落结构多样性的影响[J]. 安徽农业科学, 2008, 36(12): 5126-5127. doi: 10.3969/j.issn.0517-6611.2008.12.141 [17] XIANG D H, DONG X, LIAN X. Changes in soil microbial functional diversity and biochemical characteristics of tree peony with amendment of sewage sludge compost[J]. Environmental Science Pollution Research, 2015, 22(15): 11617-11625. doi: 10.1007/s11356-015-4407-9 [18] GARCIA-GIL C, PLAZA C, SENESI N, et al. Effects of sewage sludge amendment on humic acids and microbiological properties of semiarid Mediterranean soil[J]. Biology and Fertility of Soils, 2004, 39(5): 320-328. doi: 10.1007/s00374-003-0709-z [19] 周然. 石灰性土壤微生物生物量C/P比的影响因素[D]. 北京: 中国农业大学, 2005. [20] 黄林, 乔俊辉, 郭康莉, 等. 连续施用无害化污泥对沙质潮土土壤肥力和微生物学性质的影响[J]. 中国土壤与肥料, 2017(5): 80-86. doi: 10.11838/sfsc.20170514 [21] 刘恋, 张建峰, 姜慧敏, 等. 无害化污泥施用对土壤有机质、微生物量碳和氮含量的影响[J]. 农业环境科学学报, 2014, 33(5): 978-984. doi: 10.11654/jaes.2014.05.022 [22] SANTOS J A, NUNES L A P L, MELO W J, et al. Tannery sludge compost amendment rates on soil microbial biomass of two different soils[J]. European Journal of Soil Biology, 2011, 47(2): 146-151. doi: 10.1016/j.ejsobi.2011.01.002 [23] SELIVANOVSKAYA S Y, LATYPOVA V Z. Effects of composted sewage sludge on microbial biomass, activity and pine seedlings in nursery forest[J]. Waste Management, 2006, 26(11): 1253-1258. doi: 10.1016/j.wasman.2005.09.018 [24] SASTRE I, VICENTE M A, LOBO M C. Influence of the application of sewage sludges on soil microbial activity[J]. Bioresource Technology, 1996, 57(1): 19-23. doi: 10.1016/0960-8524(96)00035-1 [25] 鲍士旦. 土壤农化分析[M]. 3版. 北京: 中国农业出版社, 2008. [26] LIU J J, SU Y Y, YU Z H, et al. Soil carbon content drives the biogeographical distribution of fungal communities in the black soil zone of north-east China[J]. Soil Biology and Biochemistry, 2015, 83: 29-39. doi: 10.1016/j.soilbio.2015.01.009 [27] TOKALA R K, STRAP J L, JUNG C M, et al. Novel plant-microbe-rhizosphere interaction involving Streptomyces lydicus WYEC108 and the pea plant(Pisum sativum)[J]. Applied and Environmental Microbiology, 2002, 68(5): 2161-2171. doi: 10.1128/AEM.68.5.2161-2171.2002 [28] 王光华, 刘俊杰, 于镇华, 等. 土壤酸杆菌门细菌生态学研究进展[J]. 生物技术通报, 2016, 32(2): 14-20. [29] 马维伟, 李广, 石万里, 等. 甘肃尕海湿地退化过程中植物生物量及物种多样性变化动态[J]. 草地学报, 2016, 24(5): 960-966. doi: 10.11733/j.issn.1007-0435.2016.05.005 [30] ANDRES P, MATEOS E, TARRASON D, et al. Effects of digested, composted and thermally dried sewage sludge on soil microbiota and meaofauna[J]. Applied Soil Ecology, 2011, 48(2): 236-242. doi: 10.1016/j.apsoil.2011.03.001 [31] KATANDA Y, MUSHONGA C, BANGANGYI F, et al. Effects of heavy metals contained in soil irrigated with a mixture of sewage sludge and effluent for thirty years on soil microbial biomass and plant growth[J]. Physics and Chemistry of the Earth, 2007, 32(15/16/17/18): 1185-1194. [32] 邵颖, 曹四平, 刘长海. 基于高通量测序的南泥湾湿地土壤细菌多样性分析[J]. 干旱区资源与环境, 2019, 33(2): 158-163. [33] FENG Y, GROGAN P, CAPORASO J G, et al. pH is a good predictor of the distribution of anoxygenic purple phototrophic bacteria in Arctic soils[J]. Soil Biology and Biochemistry, 2014, 74(6): 193-200. [34] BHATTACHARYA T, CHAKRABORTY S, BANERJEE D K. Heavy metal uptake and its effect on macronutrients, chlorophyll, protein, and peroxidase activity of Paspalum distichum grown on sludge-dosed soils[J]. Environmental Monitoring Assessment, 2011, 169(1/2/3/4): 15-26. [35] KAO P H, HUANG C C, HSEU Z Y. Response of microbial activities to heavy metals in a neutral loamy soil treated with biosolid[J]. Chemosphere, 2006, 64(1): 63-70. doi: 10.1016/j.chemosphere.2005.11.039 [36] MACDONALD C A, SINGH B K, PECK J A, et al. Long-term exposure to Zn-spliked sewage sludge alters soil community structure[J]. Soil Biology and Biochemistry, 2007, 39(10): 2576-2586. doi: 10.1016/j.soilbio.2007.04.028 [37] 陈同斌, 黄启飞, 高定, 等. 中国城市污泥的重金属含量及其变化趋势[J]. 环境科学学报, 2003, 23(5): 561-569. doi: 10.3321/j.issn:0253-2468.2003.05.001 [38] 陈泽斌, 高熹, 王定斌, 等. 生物炭不同施用量对烟草根际土壤微生物多样性的影响[J]. 华北农学报, 2018, 33(1): 224-232. doi: 10.7668/hbnxb.2018.01.032 期刊类型引用(4)
1. 钟俏君,韦柳江,庄炜东,杨小霞,赵志娟. 施用堆肥污泥对土壤细菌群落结构及朝天椒生长的影响. 南宁师范大学学报(自然科学版). 2024(01): 161-168 .  百度学术
百度学术
2. 周冉,张志斌,张晓蕊,孙杰,张彦浩. 干湿交替对南四湖底泥真菌群落影响研究. 山东建筑大学学报. 2022(05): 65-69+120 . 百度学术
3. 王启震,常会庆,吴杰,徐晓峰. 施用高量堆肥污泥提高小麦–玉米轮作模式下酸性水稻土pH和碳组分活性. 植物营养与肥料学报. 2021(02): 253-262 . 百度学术
4. 常会庆,王启震,吴杰,陈鲜妮. 腐熟污泥施用对土壤有机-无机碳组分的影响. 农业机械学报. 2020(08): 295-302 . 百度学术
其他类型引用(3)
-

 点击查看大图
点击查看大图
计量
- 文章访问数: 3177
- HTML全文浏览数: 3177
- PDF下载数: 44
- 施引文献: 7
