

























-
多环芳烃(PAHs)是一种难降解性有机污染物,其环境归趋受到人们的广泛重视与关注[1-2]. 其中,苯并[a]芘(BaP)是环境中主要的PAHs种类,我国每年约有470—900 t的BaP排放到环境中. BaP作为强致癌类物质的代表,是我国环境监测的常规项目[3]. 人工湿地(CWs)可以利用基质吸附、植物吸收、微生物降解等物理、化学、生物协同作用有效降低BaP浓度. 研究表明,CWs对BaP的去除率约为78%—92%,是环境中BaP一个重要的汇[4]. 与基质吸附相比,植物生物修复可以将汇入CWs中的BaP进行捕获、吸附或吸收,这一去除过程更为彻底,体现了CWs的“绿色”特征[5].
植物对环境中BaP的吸收是一个复杂而系统的过程,涉及不同的生物和化学等过程[6]. 研究表明,植物具有高比表面积和脂肪含量的侧根可有效吸附BaP[7],根系吸附的BaP不能被植物完全吸收,BaP只有在穿过质膜进入细胞质通道时才能被根吸收[8]. 植物通过释放根系分泌物造成基质上有机质分解速率改变的现象被称为植物激发效应[9],其对于地下环境有机质周转以及全球碳循环的影响至关重要[10],但当前尚缺乏CWs中根系分泌物激发效应的系统研究.
本研究以BaP为研究对象,系统分析典型湿地水生植物菖蒲中BaP的去除、吸收和归趋规律及其对CWs去除BaP的影响,并通过根箱试验,深入探究根系分泌物存在情况下湿地基质-水界面BaP的组分转化,明确根系分泌物对BaP赋存状态与环境归趋的影响机制.
-
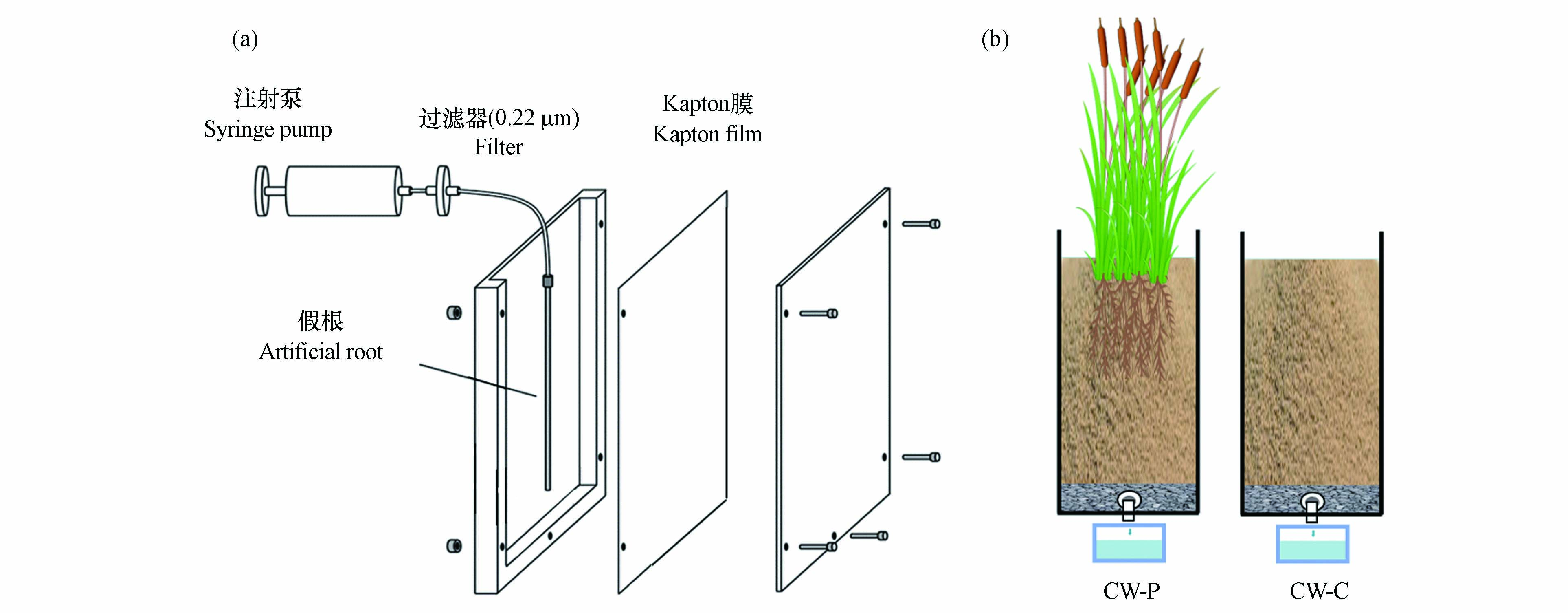
在植物生长间用装有10.00% Hogland溶液的1 L水桶中培养表面灭菌的菖蒲幼苗种. 对菖蒲的根系分泌物进行收集,进行冻干保存[11];使用微孔圆柱形根取样器(rhizon CSS 19.21.24F)作为假根来运输根系分泌物,构建一个微观间歇潜流根箱系统(图1a);将具有BaP生物降解能力的细菌菌株 M. vanbaalenii PYR-1(DSM No.7251)添加到根箱中;石英砂均匀填充到根箱,以获得1.46 g·cm−3的体积密度. 根系分泌物的输送速率为15.00 μmol ·cm−2·d−1C,输送量为1 mL·d−1. 设计两种处理组:A组(M.vanbaalenii PYR-1和不含根系分泌物)和B组(根系分泌物和M.vanbaalenii PYR-1). 构建两个间歇潜流CWs系统:CW-P(种植菖蒲CWs)和CW-C(无植物的对照CWs)(图1b),内径为15.00 cm,高度为50.00 cm. 所有试验装置结构均填充了砂质石英(Φ:(2.00±1.00)mm,孔隙度:0.40),高达45.00 cm. BaP浓度为10.00 μg·L−1,水力停留时间(HRT)为2 d.
-
按照HRT的频率收集废水和基质样品. 采用固相萃取法对废水中的BaP进行萃取[12]. 从每个试验装置中取3份1.00 g(干重)等分基质试样,用Tenax TA、己烷和丙酮与超声提取的混合物以及水浴中的碱性水解,依次处理基质中BaP的不稳定、稳定吸附和结合残余组分[13]. 在试验结束后回收植物的茎部和根部,采用二氯甲烷与超声提取技术提取植物组织中的BaP[14]. 应用气相色谱/质谱(GC/MS,岛津QP2020)系统来识别和测量样品中的BaP. 根据Cheng等[13]提供的模型模拟溶解性有机质(DOM)的根部诱导生成率变化,如下式:
式(1)中,bDOM是DOM的基质缓冲力,DDOM是DOM在基质水中的扩散系数,kDOM是DOM分解的一阶速率常数,σ(x)是距离根部x(cm)处DOM的根部诱导生成率(mg·L−1·d−1).
根据Sivaram等[15]提供的方法计算根系富集因子RCF、茎叶富集因子SCF以及迁移因子TF,如下式:
式(2)中,Cr为根中BaP浓度,Cw为水中BaP浓度.
式(3)中,Cs为茎中BaP浓度,Cw为水中BaP浓度.
-
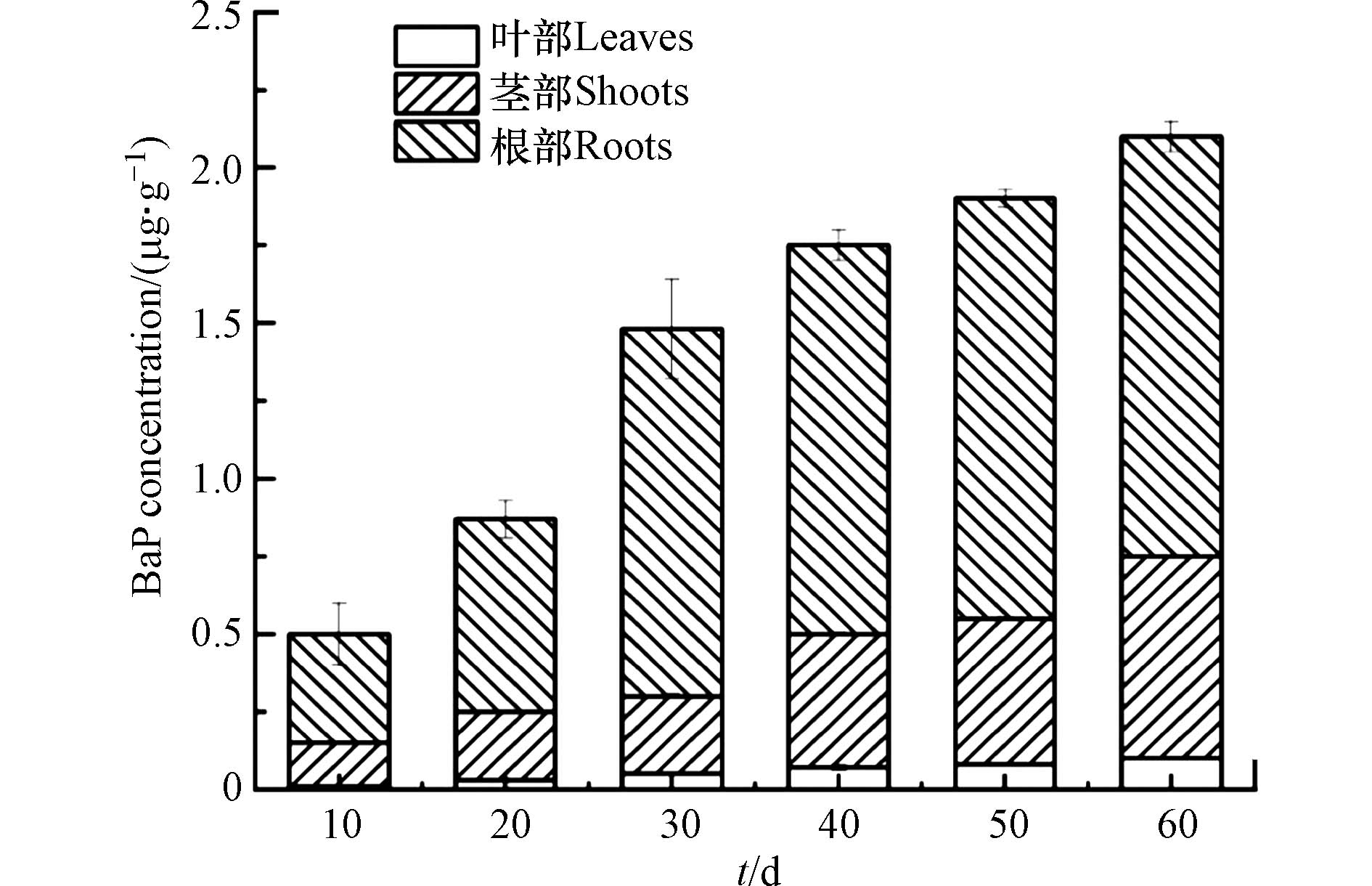
分析了BaP在菖蒲中不同部位(根、茎和叶)的分布,结果如图2所示. 随着暴露时间的延长,植物组织中的污染物浓度增加,菖蒲中积累的BaP含量达到(2.13±0.05)μg·g−1,且根中观察到较高浓度的BaP. 植物积累模式的显著差异可能是由于植物的生理特性的不同造成的,例如,植物脂肪含量、植物生物量、解毒和代谢系统、生长和蒸腾速率等[16],Jiao等[7]指出,具有高比表面积和脂肪含量的侧根对PAHs的吸附能力显著高于节根. 吸附在根表面的PAHs可以迁移到细胞壁的孔隙中,在那里它们可以与外部溶液自由交换. Kang等[14]观察到,黑麦草细胞壁中的苯丙氨酸迅速在前25 h增加到60 mg·kg−1以上,是其他细胞器的4倍. 这表明苯丙氨酸在吸附到其他细胞器之前先被吸附到细胞壁上.
根系富集因子(RCF)、茎叶富集因子(SCF)和迁移因子(TF)对菖蒲去除BaP的影响存在显著差异(表1). 菖蒲的RCF值是13.00,SCF值为3.30,TF值为3.60,结果表明在菖蒲植物中RCF对BaP的去除率影响最大. TF也可用于指示BaP从根到地上部组织的易位能力,菖蒲较高的TF值说明在根际富集的BaP更容易转移到根中,在根中被吸收和积累,然后与水一起从根转移到茎、叶和贮藏器官,BaP一旦被吸收并从根部转移到叶片,就可以通过挥发过程进行释放. 此外,为进一步研究植物累积行为特征,采用因子
× 因子方式表示不同组织累积BaP相互作用的影响. 在菖蒲中RCF× SCF为6.20,RCF× TF为14.60,SCF× TF为2.40,表明菖蒲在吸收BaP的过程中表现出RCF和SCF之间的显著双向相互作用. -
为了确定根系分泌物在基质中的影响范围,测定了DOM浓度的变化,结果如图3a所示. 根系分泌物导致DOM浓度随离假根距离呈降低趋势,结合DOM一级动力学模型分析,该系统中根系分泌物的影响小于4.00 cm(图3b).
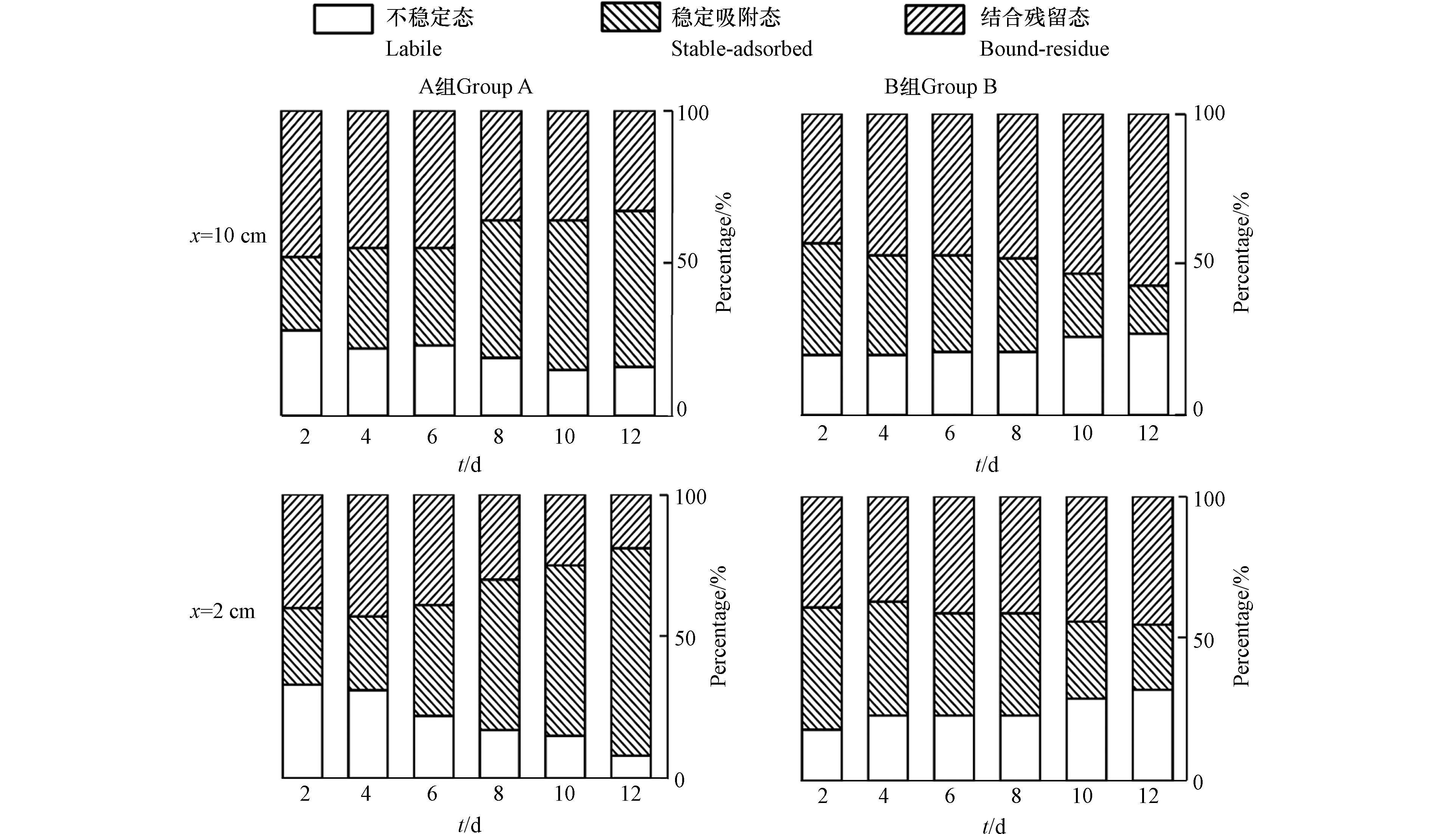
为了厘清BaP的生物可利用性和植物吸收、微生物降解、结合态的形成及它们各自对BaP消减过程的贡献,研究了BaP在湿地基质环境中赋存形态的变化,结果如图4所示. 结果表明,菖蒲根系分泌物显著影响湿地基质环境中BaP赋存形态. BaP结合残留态组分随着时间和与假根距离的延长而增加(图4c,图4d),说明菖蒲根系分泌物可以争夺BaP分子所占据的活性吸附位点和区域,导致BaP溶解于水中或在基质颗粒上松散吸附. BaP稳定吸附组分随着时间和与假根距离的延长而减少. 一般认为在老化作用下,BaP可能从松散吸附状态转变为更持久的结合相[17]. BaP活性态组分在菖蒲根系分泌物作用范围随着时间的延长而增加,作为在基质-水界面上能够快速解吸的生物可利用态,表明菖蒲根系分泌物的存在提高了生物对BaP的利用. 综上所述,菖蒲根系分泌物可以影响BaP在湿地基质环境中的归趋,菖蒲根系分泌物和微生物的共同作用加速了BaP的赋存形态转化,提升了湿地基质对BaP的去除. Keiluweit等[18]应用纳米二次离子质谱、STXM/NEXAFS成像揭示了根系泌碳背后的生物-非生物耦合机制,并发现根系分泌物会破坏基质矿物质保护作用,导致有机物生物可利用性提高,增强微生物接触. 本研究中,根系分泌物会使得基质上BaP从结合残留态组分转化成为活性态组分,影响BaP在湿地基质环境中的归趋,根系分泌物和微生物的共同作用加速了BaP的赋存形态转化,提升了湿地基质对BaP的去除.
-
研究了菖蒲植物对CWs中BaP去除的作用,结果如图5所示. 系统稳定运行后,种植菖蒲的CW-P中的BaP浓度减少了96.38 %,而CW-C的BaP出水浓度仅减少16.91 %,菖蒲CWs的BaP去除率较对照系统提升79.47%. CW-P基质根际区中吸附的BaP浓度为(0.89±0.028)μg·g−1,显著高于基质非根际区吸附的BaP((0.07±0.02)μg·g−1). Zhalnina 等[19]也得到相似的实验现象,发现植物可以调控根际微生物,增强有机物的降解作用,促进植物吸收有机物,保持植物生长良好. 因此,菖蒲植物对CWs中BaP的消散有促进作用,植物对BaP的吸收和根际微生物的降解可能共同导致根区BaP的急剧损失.
研究了基质中BaP赋存形态变化,结果如图6所示. 在CW-P根际区域,基质中BaP的不稳定组分从78.00%增加至82.00%,在非根际区域,基质中BaP的不稳定组分从37.00%增加至40.00%,而CW-C中的不稳定组分从22.00%降低至18.00%,表明BaP在根际的生物有效性较高,且菖蒲极大地增加了基质中BaP不稳定组分.
BaP结合残留组分是在老化作用下形成的,这种解毒过程在受根系活动影响较小的非根际区域或未种植植物的系统中尤为明显. CW-P中非根际区域的结合残留组分从21.00%增加至25.00%,根际区域从2.00%降低至1.00%,CW-C中结合残留组分占53.00%—58.00%的比例. 在整个试验周期,BaP在根际区域中的结合残留组分始终低于非根际区域中的结合残留组分以及未种植植物的系统,这表明结合残留组分BaP的形成受到根际效应的抑制.
作为基质中的过渡组分,BaP的稳定吸附组分随根际和非根际的区域而变化. 在CW-P非根际区域中,基质中BaP的稳定吸附组分从35.00%增加至42.00%,在根际区域中变化幅度较小. 稳定吸附部分被认为是对基质颗粒的可逆吸附. 在老化作用下,BaP可能从松散吸附状态转变为更持久的结合相,或作为不稳定组分溶解在水中.
-
(1) 菖蒲可累积大量BaP,且大多累积于根部组织中,较高的易位因子表明BaP可以更容易从根到地上部组织进行迁移.
(2) 菖蒲大幅提升了CWs对BaP的去除效果,去除率较对照系统提升79.47%.
(3) 菖蒲的根系分泌物可以提高BaP的生物可利用性,有效促进基质中不稳定态BaP转化.
典型人工湿地系统中植物对苯并[a]芘的去除作用及机制
Removal of benzo[a]pyrene by plants in typical constructed wetland system and its mechanism
-
摘要: 人工湿地(CWs)可有效去除环境中的苯并[a]芘(BaP),湿地植物对BaP的吸收是受生物、化学等因素影响的复杂过程,其对CWs中BaP环境归趋的影响机制还不明确. 本文系统分析了典型湿地植物菖蒲中BaP的去除、吸收和归趋规律. 结果表明,菖蒲可累积大量的BaP,含量达到(2.13±0.05)μg·g−1;根部组织中观察到较高浓度的BaP,BaP可以更容易地从菖蒲的根部到地上部组织进行迁移. 菖蒲大幅提升了CWs对BaP的去除效果,菖蒲CWs的BaP去除率较对照系统提升79.47%. 菖蒲的根系分泌物可以提高BaP的生物可利用性,加速BaP的组分净化和转化.Abstract: Constructed wetlands (CWs) can effectively remove benzo[a]pyrene (BaP) from the environment. The absorption of BaP by wetland plants is a complex process affected by biological, chemical and other factors, and its impact mechanism on the environmental fate of BaP in CWs is still unclear. In this paper, the removal, absorption and fate of BaP in typical wetland plant Acorus calamus were systematically analyzed. The results showed that Acorus calamus could accumulate a large amount of BaP. The content of BaP in Acorus calamus could reach (2.13±0.05) μg·g−1. Higher concentrations of BaP were observed in the root tissues, and BaP could migrate more easily from the root to the shoot tissues. The BaP removal rate of CWs with Acorus calamus was 79.47% higher than that of the control CWs, indicating the presence of Acorus calamus can greatly improve the removal effeciency of CWs on BaP. The root exudates of Acorus calamus can enhance the bioavailability of BaP and subsequently accelerate the purification and transformation of BaP components.
-
Key words:
- BaP /
- environmental fate /
- constructed wetlands /
- plant /
- rhizosphere
-
我国甘肃、新疆、青海等西北地区,干旱少雨,地表蒸发量大,属严重缺水区域[1]. 尤其是黄土塬上的偏远村镇,没有可利用的地表水和地下水,资源性缺水更加严重[2-3]. 这些村镇不具备修建大型水利工程的条件,收集雨水是当地村民唯一的饮水方式,而水窖是最常用的雨水储存设施[4].
与河、库等地表水相比,雨水是一种水质较好的优质水源[5],但在收集饮用时同样面临着一定的水质污染风险. 集流面材料是影响雨水安全集蓄的一个关键因素. 常见的集流面有瓦屋面、沥青屋面和混凝土地面等. 其中,沥青屋面释放的污染物最多,获得的水质最差;混凝土地面应用最多,释放的碱性物质会显著提高雨水pH[6]. 此外,水窖周围环境是影响窖水水质的另一重要因素. 已有研究表明,建在厕所和牲畜棚旁边的窖水含有更多的污染物质[7].
新污染物是指具有环境稳定性、生物累积性和生物毒性的有毒有害化学物质,主要包括持久性有机污染物、内分泌干扰物、抗生素和微塑料四大类. 不同于常规污染物,新污染物浓度水平一般较低,但由于生物累积和生物放大作用,对人体和生态系统潜在威胁较大.
邻苯二甲酸酯(PAEs)和全氟化合物(PFCs)是两类典型的新污染物. PAEs是一种环境激素类内分泌干扰物,能够抑制雄性激素的生成,对人类的呼吸系统、生殖系统和内分泌系统均有损害. PFCs广泛应用于聚四氟乙烯生产,是重要的防水防油材料,也是灭火泡沫的主要成分. PFCs对人体肝脏功能、脂肪代谢和遗传发育均有不良影响[8]. 已有研究表明,两种污染物广泛存在于全球大气、土壤及水环境介质中[9-12]. 可以推断,在西北高海拔黄土塬区村镇,PAEs和PFCs两种新污染物同样存在,窖水也面临着相应的的污染风险. 然而,针对西北村镇窖水这种分散型饮用水源,还未有新污染物方面的报道.
窖水作为西北高海拔村镇唯一可用的饮用水源,其水质安全对当地村民身体健康有着重要意义. 本研究基于西北黄土塬区村镇饮水现状,以我国甘肃某县为代表区域,采集冬夏两季窖水进行水质分析,研究了15种目标PAEs和17种PFCs在冬夏两季窖水中的浓度水平和组成,讨论了两种新污染物来源,并利用模型计算了采样点覆盖区域内人群通过饮用窖水摄入PAEs和PFCs的健康风险,为西北高海拔黄土塬区村镇窖水的饮用处理提供了基础数据支撑.
1. 材料与方法(Materials and methods)
1.1 研究区域及样品采集
研究区域所在县隶属甘肃省,为典型的黄土高原丘陵沟壑区,总体地势西北高、东南低,海拔1136—2089 m,常年干旱少雨,是典型的资源型缺水地区[13]. 该区以农业和畜牧业为主导产业,对水资源量的需求较高. 集雨水窖是当地村户最主要的饮用水渠道,雨季将雨水收集储存起来,仅依靠窖水的自然沉淀,未经其它处理,就直接进行生活饮用.
本次调研覆盖该县整个纵向区域,于2021年12月和2022年7月分两次,自北向南每间隔约10 km取水窖样品. 冬季窖水采样时距上次降雨约4个月,夏季窖水采样时正是雨天,具体采样点分布如图1所示,冬季共采集8个(a—h)窖水样品,所有窖水样品均进行了常规指标的检测,用于检测新污染物的窖水b、d、e、f用玻璃瓶密封,其它用聚乙烯瓶密封. 夏季共采集10个(1—10)窖水样品,所有样品均采用聚乙烯瓶密封. 1和a、2和b、4和c、6和d、7和f、8和g、9和h分别对应同一窖水采样点. 每批样品运回实验室后置于4℃冷库中保存,并在一周内完成所有样品的检测分析. 这些样品分布在不同的自然村落中,所有水窖材质均为混凝土水窖,集雨面均为混凝土地面,水窖8(g)是供水规模较大的集体水窖,其余各水窖皆为单户水窖.
1.2 药品及试剂
15种PAEs标准样品购自中国上海阿拉丁公司,如表1所列;17种PFCs标准样品,如表2所列,以及9种PFCs同位素内标(13C4PFBA、13C4PFHxA、13C4PFOA、13C4PFNA、13C4PFDA、13C4PFUdA、13C2PFDoA、18O2PFHxS 和13C4PFOS) 购自加拿大惠灵顿公司;色谱纯乙腈、色谱纯甲醇、色谱纯正己烷、色谱纯二氯甲烷均购自加拿大飞世尔科技公司;色谱级乙酸铵购自中国上海的麦克林公司;超纯水使用日本奥加诺株式会社的Milli-Q系统生成(电阻率>18.2 MΩ·cm).
表 1 检测的15种PAEs类型及化学式Table 1. The type and chemical formulas of 15 PAEs in this study化合物Compouds 英文名称English name 英文缩写Abbreviation 化学式Chemical formula CAS编号 CAS number 邻苯二甲酸二甲酯 Dimethyl phthalate DMP C10H10O4 131-11-3 邻苯二甲酸二乙酯 Diethyl phthalate DEP C12H14O4 84-66-2 邻苯二甲酸二异丁酯 Diisobutyl phthalate DIBP C16H20O4 84-69-5 邻苯二甲酸二丁酯 Dibutyl phthalate DBP C16H22O4 84-74-2 邻苯二甲酸二甲氧乙酯 Dimethylglycol phthalate DMEP C14H16O6 117-82-8 双(4-甲基-2-戊基)邻苯二甲酸酯 Bis(4-Methyl-2-pentyl)phthalate BMPP C20H30O4 146-50-9 双(2-乙氧基)邻苯二甲酸酯 Bis(2-ethoxyethyl) phthalate DEEP C16H22O6 605-54-9 邻苯二甲酸二戊酯 Di-n-pentyl phthalate DPP C18H26O4 131-18-0 邻苯二甲酸二己酯 Di-n-hexyl phthalate DNHP C20H30O4 84-75-3 邻苯二甲酸苄酯 Mono-Benzyl phthalate BBP C15H12O4 2528-16-7 双(2-正丁氧基乙酯)邻苯二甲酸酯 Bis(2-butoxyethyl) phthalate DBEP C20H30O6 117-83-9 邻苯二甲酸二环己基酯 Dicyclohexyl phthalate DCHP C20H26O4 84-61-7 邻苯二甲酸二辛酯 Di-n-octyl phthalate DNOP C24H38O4 117-84-0 双(2-乙基己基)邻苯二甲酸酯 Di(2-ethylhexyl)phthalate DEHP C24H38O4 117-81-7 邻苯二甲酸二异壬酯 Diisononyl phthalate DINP C26H42O4 68515-48-0 | Show Table DownLoad:
CSV
表 2 检测的17种PFCs类型及化学式Table 2. The type and chemical formulas of 17 PFCs in this study
DownLoad:
CSV
表 2 检测的17种PFCs类型及化学式Table 2. The type and chemical formulas of 17 PFCs in this study化合物Compouds 英文名称English name 英文缩写Abbreviation 化学式 Chemical formula CAS编号 Chemical formula 对应内标Inner standard 全氟丁酸 Perfluorobutanoic acid PFBA C3F7COOH 375-22-4 13C4PFBA 全氟戊酸 Perfluoropentanoic acid PFPeA C4F9COOH 2706-90-3 13C4PFOA 全氟己酸 Perfluorohexanoic acid PFHxA C5F11COOH 307-24-4 13C4PFHxA 全氟庚酸 Perfluoroheptanoic acid PFHpA C6F13COOH 375-85-9 13C4PFHxA 全氟辛酸 Perfluorooctanoic acid PFOA C7F15COOH 206-397-9 13C4PFOA 全氟壬酸 Perfluorononanoic acid PFNA C8F17COOH 375-95-1 13C4PFNA 全氟癸酸 Perfluorodecanoic acid PFDA C9F19COOH 335-76-2 13C4PFDA 全氟十一酸 Perfluoroundecanoic acid PFUnDA C10F21COOH 218-165-4 13C4PFUdA 全氟十二酸 Perfluorododecanoic acid PFDoDA C11F23COOH 307-55-1 13C2PFDoA 全氟十三酸 Perfluorotridecanoic acid PFTrDA C12F25COOH 276-745-2 13C2PFDoA 全氟十四酸 Perfluorotetradecanoic acid PFTeDA C13F27COOH 376-06-7 13C2PFDoA 全氟十六酸 Perfluorohexadecanoic acid PFHxDA C15F31COOH 67905-19-5 13C2PFDoA 全氟十八酸 Perfluorooctadecanoic acid PFODA C17F35COOH 16517-11-6 13C2PFDoA 全氟丁烷磺酸 Perfluorobutanesulfonic acid PFBS C4F9SO3H 375-73-5 18O2PFHxS 全氟己烷磺酸 Perfluorohexanesulfonic acid PFHxS C6F13SO3H 355-46-4 18O2PFHxS 全氟辛烷磺酸 Perfluorooctanesulfonic acid PFOS C8F17SO3H 1763-23-1 13C4PFOS 全氟癸烷磺酸 Perfluorodecanesulfonic acid PFDS C10F21SO3H 335-77-3 13C4PFOS | Show TableDownLoad:
CSV
1.3 样品分析及实验方法
温度、溶解氧(DO)、pH、总溶解固体(TDS)、氧化还原电位(ORP)采用美国哈希公司的HQ40d多参数分析仪在现场测定;浊度采用哈希2100Q便携式浊度仪在现场测定;总有机碳(TOC)和总氮(TN)采用日本岛津公司的 TOC-L 总有机碳分析仪测定.
样品中的PAEs用C18固相萃取柱提取,富集倍数为100倍. 将500 mL各样品经0.45 μm尼龙滤膜过滤后置于1 L玻璃烧杯中备用. 萃取前,依次用5 mL甲醇、5 mL二氯甲烷、5 mL正己烷和5 mL超纯水活化C18小柱. 活化后,开始进行萃取,水样通过C18小柱的流速控制在每秒1滴. 水样全部通过小柱且淋洗完成后,利用真空泵抽干残余水分,然后用5 mL正己烷洗脱,取适量洗脱液,用外标法在日本岛津公司的TQ8030三重四极杆气相色谱质谱联用仪上进行检测. 色谱柱:Agilent DB-5MS UI 30 m×0.25 mm×0.25 μm, Intuvo 气相色谱柱 (货号:122-5532UI-INT);GC 部分:进样口温度 260℃,不分流进样,进样时间1 min;总流量50 mL·min−1,吹扫流量3 mL·min−1,柱流量1.2 mL·min−1,载气为恒压72.8 kPa,程序升温为: 初始温度60℃保持1 min,以10 ℃·min−1升至220 ℃保持1 min,再以7 ℃·min−1 升至260℃,保持10 min,总时间为33.71 min;MS部分:离子源温度230℃,接口温度为260℃,溶剂延迟时间7 min;各物质定性离子对和定量离子对分别为:DMP 163.0>133.1、163.0>77.1 ; DEP 177.0>149.0、177.0>93.1 ; DIBP 149.0>65.0、149.0>93.1;DBP 149.0>65.1、149.0>93.1;DMEP 149.0>65.1、149.0>93.1;BMPP 149.0>65.1、149.0>93.1;DEEP 176.0>149.0、176.0>104.1;DPP 149.0>65.1、149.0>93.1;DNHP 149.0>65.1、149.0>93.1;BBP 206.0>149.0、206.0>93.1;DBEP为193.0>149.0、193.0>93.1;DCHP 167.0>149.1、167.0>93.0;DNOP 149.0>65.1、149.0>93.1;DEHP 167.0>149.0、167.0>93.0;DINP 293.0>149.0、293.0>93.0.
样品中的全氟化合物(PFCs) 用WAX固相萃取柱提取. 样品各取400 mL加入2ng内标后不经过滤直接萃取,以检测窖水中PFCs的总浓度并考察浊度对PFCs浓度的影响. 萃取前,WAX小柱依次用4 mL甲醇、4 mL氨水甲醇和4 mL超纯水活化. 水样全部通过小柱且淋洗完成后,利用真空泵抽干残余水分,然后依次用4 mL甲醇、4mL 0.1% 氨水甲醇洗脱,将洗脱液氮吹至近干,用甲醇复溶至200 μL,浓缩倍数为2000倍,最后在电喷雾离子源负离子模式(ESI-)下采用内标法在美国安捷伦公司1290 Infinity HPLC系统与Agilent 6460三重四极杆质谱系统联用的高效液相色谱串联质谱仪上检测. 检测方法在本课题组已有研究中给出,采用其所用的定性定量参数,流速改为0.2 mL·min−1[14].
1.4 质量控制
为了保证测定结果的可靠,窖水样品中PAEs和PFCs的测定均设置有1个平行,1个方法空白、3个溶剂空白和1个现场空白(在现场用相同采样瓶,在相同时间装入娃哈哈纯净水),并进行了3个窖水样品和3个超纯水空白的加标回收.
PAEs的空白加标回收率为87.3%—103.7%,是在3份超纯水样品中加入1 μg·L−1混合标准样品测得的. PAEs的基质加标回收率为82.5%—101.6%,是在3份窖水样品中加入1 μg.L−1 混合标准样品测得的. 以信噪比S/N>3为检出限,PAEs检出限范围为0.09—0.19 μg·L−1,其中DIBP、DMEP、DBEP 3种物质的空白值达到了各自检出限分别为0.27、0.14、0.10 μg·L−1,在计算浓度时需进行差减扣除. 各物质相对标准偏差范围为1.27%—9.79%. PFCs检出限范围为0.01—0.09 ng·L−1,所有空白均未达到检出限. 各物质相对标准偏差范围为2.50%—12.82%. PFCs检测的空白加标回收率为80.5%—119.7%,是在3份超纯水样品中加入20 ng混合标准样品测得的. PFCs检测的基质加标回收率为75.3%—117.1%,是在3份窖水样品中加入20 ng混合标准样品测得的,检测中PAEs和PFCs的校准曲线回归系数(r2)均超过0.99.
1.5 健康风险评价模型
采用US EPA 提供的健康风险评价模型来评价窖水中PAEs,该评价模型包括致癌和非致癌两个部分[15-16]. 非致癌风险用风险值HI来表示,计算公式如下:
HI=CDIRfD (1) 式中,CDI是由于暴露造成的污染物长期日摄入剂量,
mg⋅(kg⋅d)−1 mg⋅(kg⋅d)−1 致癌风险则用R表示,是长期日摄入剂量与致癌斜率因子的乘积,计算公式如下:
R=CDI×SF (2) 式中,SF是致癌斜率因子,
(kg⋅d)⋅mg−1 饮用水途径暴露造成的污染物长期日摄入剂量计算公式如下:
CDI=C×U×EF×EDBW×AT (3) 式中,C为水中目标污染物的质量浓度,mg·L−1;U为日饮用水量,L·d−1;EF为暴露频率,d·a−1;ED为暴露持续时间,a;BW为平均体重,kg;AT为平均暴露时间,d.
以上公式中的参数取值来自文献[17]和中国人群暴露参数手册(成人卷)[18] : 夏、冬两季U分别取3.1 L·d−1和1.9 L·d−1(甘肃省);EF取365 d·a−1;ED对于非致癌物取30 a,对于致癌物取70 a;BW取61.8 kg(甘肃省); AT对于非致癌物取10950 d,对于致癌物取25550 d.
根据人群的PFCs饮水途径每日摄入量(estimated daily intake, EDI)来评估甘肃该县村民每日通过饮水摄入PFCs的健康风险. EDI计算公式如下:
EDI=Cf×VwaterBW 式中,Cf为窖水样品中PFCs的测定浓度,ng·L−1;Vwater为日饮水量,L·d−1;BW为平均体重,kg. 夏、冬两季Vwater分别取3.1 L·d−1和1.9 L·d−1(甘肃省);BW取61.8 kg(甘肃省).
把该县人群EDI值同美国卫生和公共服务部给出的PFOA、PFOS、PFHxS和PFNA人群每日可耐受摄入量参考值(分别为3、2、20 、3
ng⋅(kg⋅d)−1 2. 结果与讨论(Results and discussion)
2.1 窖水常规指标检测
冬季7个窖水样品的常规指标检测结果如表3所示,水温5.4—8.5℃,浊度1.8—29.5 NTU,pH 8.0—9.7,DO 4.9—8.8 mg·L−1,ORP 154.5—223.5 mV,TDS 59.6—110.0 mg·L−1,TOC 0.58—2.61 mg·L−1,TN 0.64—2.25 mg·L−1. 7个窖水样品pH均大于8,最高达到9.7. 不同窖水浊度差异较大,除了d、e、f 3个采样点窖水样品浊度低于《生活饮用水卫生标准》(GB5749-2022)中分散式供水所规定3 NTU,其它样品浊度均超出了标准中规定的限值.
表 3 冬季窖水样品常规指标检测结果Table 3. Test results of general parameters of cellar water samples in winter地点Position 温度/℃Temperature 浊度/NTUTurbidity pH 溶解氧/(mg·L−1)DO 氧化还原电位/mVORP 总溶解固体/(mg·L−1)TDS 总有机碳/(mg·L−1)TOC 总氮/(mg·L−1)TN a 8.5 11.7 9.7 6.9 154.5 61.4 0.69 0.99 b 7.8 29.5 8.0 7.7 207.0 92.4 2.61 2.25 c 7.9 4.2 8.8 6.2 223.5 67.7 1.02 0.78 d 6.1 1.8 9.0 4.9 207.7 59.6 0.58 1.00 e 5.8 3.2 8.6 5.6 196.3 62.4 0.62 1.02 f 6.2 2.3 8.4 5.2 213.3 65.6 0.73 1.23 g 5.4 6.1 8.6 8.8 185.4 110.0 1.62 0.64 h 7.2 5.3 8.3 5.9 218.6 68.0 0.98 0.68 | Show TableDownLoad:
CSV
夏季10个窖水样品的常规指标检测结果如表4所示,水温12.5—21.9℃,浊度4.7—58.5 NTU,pH 8.2—9.4,DO 2.9—7.6 mg·L−1,ORP 173.9—235.4 mV,TDS 46.4—238.0 mg·L−1,TOC 0.78—3.18 mg·L−1,TN 0.80—3.27 mg·L−1. 10个窖水样品pH均大于8,最高达到9.4.
表 4 夏季窖水样品常规指标检测结果Table 4. Test results of general parameters of cellar water samples in summer地点Position 温度/℃Temperatur 浊度/NTUTurbidity pH 溶解氧/(mg·L−1)DO 氧化还原电位/mVORP 总溶解固体/(mg·L−1)TDS 总有机碳/(mg·L−1)TOC 总氮/(mg·L−1)TN 1 18.8 30.6 8.8 5.9 193.2 51.3 1.91 2.07 2 21.3 58.5 9.4 6.3 211.7 223.0 3.18 3.27 3 12.5 4.7 9.0 7.6 176.5 62.9 0.78 1.38 4 17.0 18.8 8.5 6.0 204.0 60.0 1.39 0.98 5 21.9 14.1 8.5 4.8 188.2 68.7 1.19 0.80 6 20.2 11.8 8.4 6.5 226.7 61.6 1.18 1.97 7 19.3 15.1 8.6 5.1 189.7 51.7 1.34 0.99 8 21.6 19.6 8.5 4.3 235.4 238.0 1.56 2.45 9 19.5 8.8 8.3 2.9 228.0 46.4 1.22 2.05 10 18.2 19.0 8.2 5.6 173.9 53.7 1.51 1.68 | Show TableDownLoad:
CSV
夏季雨期采集的窖水样品TOC、TN、温度和浊度普遍高于冬季. 所有水窖样品浊度均超过GB5749-2022中分散式供水所规定限值,这是因为雨期水窖储水后没有经过自然沉淀,肉眼可见悬浮物较多. 同一采样点2(b)和8(g)样品浊度和TDS在各窖水样品中依然处于较高水平,实地调研发现2(b)水窖存在渗漏,8(g)水窖是集体水窖且位于道旁,两个水窖易受到周围环境的污染.
2.2 窖水中PAEs的浓度水平
对冬季窖水样品b、d、e、f和夏季10个窖水样品进行PAEs组成及浓度的检测分析,结果如图2(a)、(b)所示. 15种目标PAEs中共有13种被检出,冬季样品总浓度平均值为3.12 μg·L−1. 其中DBP在各窖水样品中含量最高,平均值达到1.06 μg·L−1,其次是DIBP和DEHP,浓度平均值分别为0.43 μg·L−1和0.33 μg·L−1. 值得注意的是,b水窖中的DINP浓度较其它三口水窖高出约8倍,达到了0.84 μg·L−1. 夏季窖水样品总浓度平均值为4.56 μg·L−1,DBP和DIBP是各窖水样品的主要组成部分,浓度平均值分别为1.3μg·L−1和1.2 μg·L−1,两者占PAEs总量的64.5%—92.7%,高于冬季窖水两种物质比例43.5%—52.1%. 同一采样点2(b)、6(d)和7(f),夏季样品中DIBP浓度均显著高于冬季,而夏季样品中的DMP和DEP则略少于冬季. DIBP、DMP和DEP的LogP分别为4.46、1.64和2.70,与DMP和DEP相比,DIBP表现出更大的疏水性. 冬季窖水在长期自然沉淀过程中,疏水性更强的DIBP更易于随着颗粒物沉淀,而亲水性较强的DMP和DEP则在窖水中稳定存在,甚至会因颗粒物的释放,导致窖水中的溶解性PAEs含量上升,这可能是夏季窖水样品中DIBP浓度较冬季窖水低,而DMP和DEP浓度低于冬季的原因.
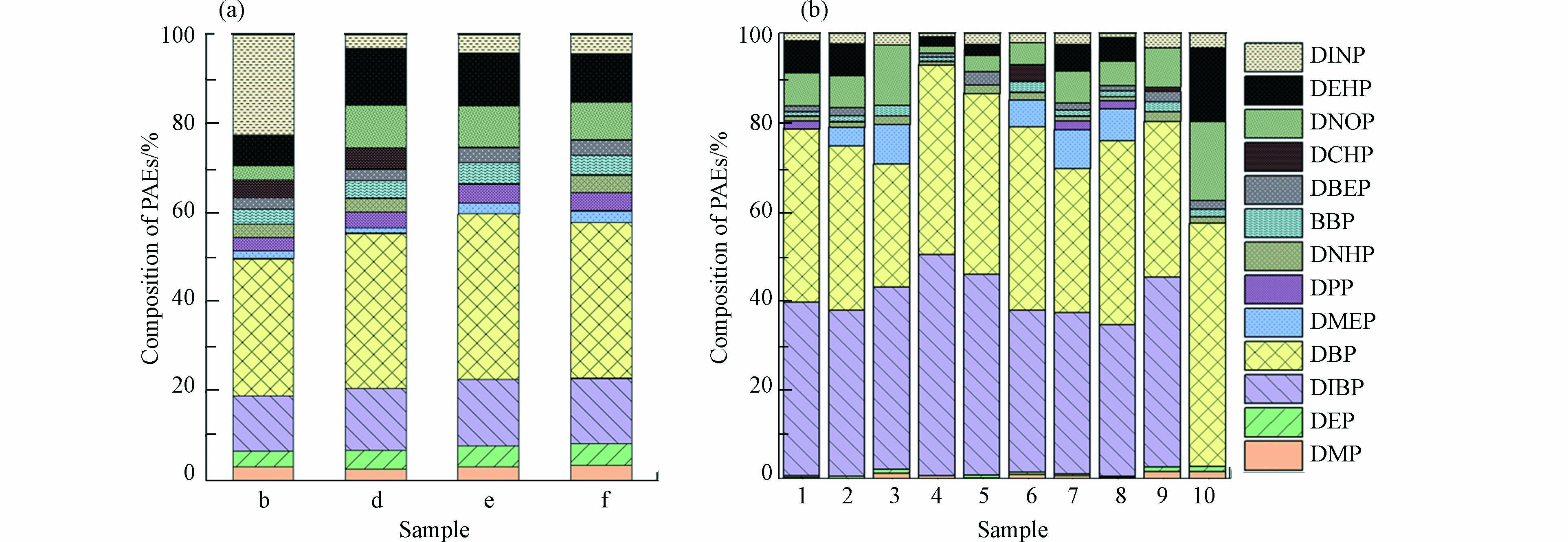 图 2 (a)冬季4个窖水样品和(b)夏季10个窖水样品PAEs组成Figure 2. PAEs composition of (a) four amples in winter and(b) ten amples in winter
图 2 (a)冬季4个窖水样品和(b)夏季10个窖水样品PAEs组成Figure 2. PAEs composition of (a) four amples in winter and(b) ten amples in winter窖水中PAEs的可能来源:一是农村常使用塑料薄膜来建造温室大棚。塑料膜在使用及降解过程中,其含有的PAEs,最终可以水为介质进入环境中. 二是农村卫生状况相对较差,破损塑料膜、垃圾袋及包装纸等随意丢弃后,其中的PAEs可能通过降雨或径流进入到集雨水窖之中,带来水质风险[19].
通过与文献报道的其他水源中PAEs含量对比发现,一般地表水中的PAEs浓度范围波动较大,如三峡库区中6种优先控制PAEs(DMP、DEP、DBP、BBP、DEHP和DNOP)的总浓度为0.42—0.77 μg·L−1[20],远小于窖水中这6种PAEs的总浓度0.85—4.56 μg·L−1,而黄河甘肃兰州段干湿季PAEs的平均总浓度分别为3.24 μg·L−1和2.30 μg·L−1,与窖水中PAEs总浓度平均值相近[21]. 此外,窖水中的PAEs浓度普遍高于地下水,如东莞地区地下水中6种PAEs(DMP、DEP、DNBP、BBP、DEHP和DNOP)总浓度平均值为0.93 μg·L−1[22],湖北江汉地下水中PAEs总浓度平均值为0.98 μg·L−1[23],均小于此次窖水中的含量3.12 μg·L−1(冬季)和4.56 μg·L−1(夏季).
2.3 窖水中PFCs的浓度水平
采用与PAEs测定相同的样品进行PFCs分析. 结果如图3和图4所示,所有窖水样品中,17种目标PFCs中共有14种被检出. 图3是冬季4个窖水样品PFCs组成情况,图4是夏季10个窖水样品PFCs组成情况,冬季PFCs总浓度范围为143.93—246.47 ng·L−1,夏季PFCs总浓度范围为275.90—405.51 ng·L−1,各窖水样品中的PFCs由PFCAs和PFSAs两部分组成. 其中PFCAs占窖水PFCs总量的80%以上,居于主体地位. PFCAs和PFSAs根据碳链长度可分为长链和短链,短链PFCAs占窖水PFCs总量的75%以上,全氟丁酸(PFBA)是短链PFCAs中含量最高的污染物,冬季窖水样品中平均浓度达到88.09 ng·L−1,而夏季窖水样品中平均浓度能达到214.98 ng·L−1.
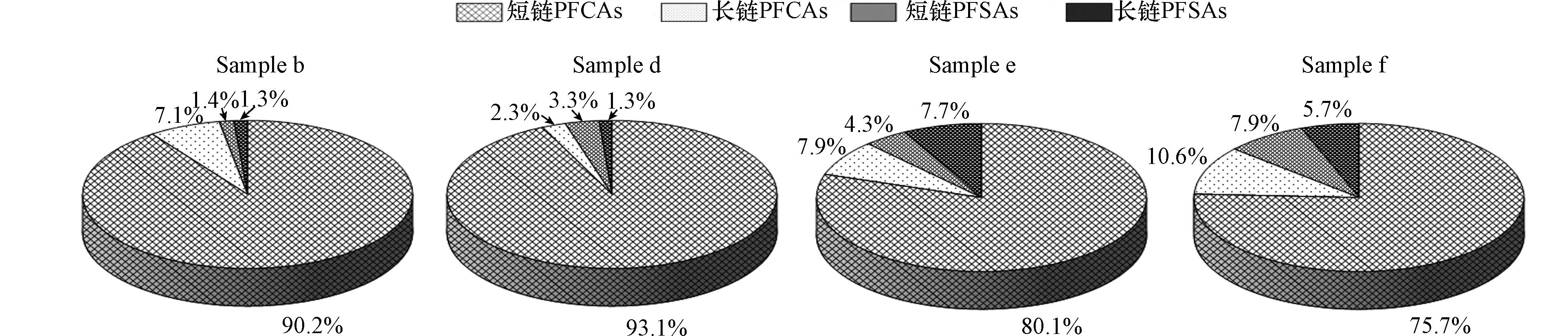 图 3 冬季4个窖水样品中的PFCs组成情况Figure 3. Composition of PFCs in four cellar water samples in winter
图 3 冬季4个窖水样品中的PFCs组成情况Figure 3. Composition of PFCs in four cellar water samples in winter 图 4 夏季10个窖水样品中的PFCs组成情况Figure 4. Composition of PFCs in ten cellar water samples in summer
图 4 夏季10个窖水样品中的PFCs组成情况Figure 4. Composition of PFCs in ten cellar water samples in summer同一采样点2(b)、6(d)和7(f),除PFPeA和PFHpA外,夏季雨期窖水PFCs浓度均高于冬季,尤其是PFOA和PFOS在所有PFCs中的浓度占比分别从6.7%和1.4%上升到了9.7%和15.1%. 这可能是由于夏季当地造纸、皮革和石油开采等氟化工产业扩大生产,提高产能,使得PFCs排放量上升,这些PFCs进入大气后随着降雨过程进入到水窖中,使得窖水PFCs浓度水平上升.
与地表水和地下水中的PFCs含量对比可知,相比于上海黄浦江和山东部分区域地表水中10种PFCs (PFBA、PFPeA、PFBS、PFHxA、PFHxS、PFHpA、PFOA、PFOS、PFNA、PFDA)总浓度(3.38—362.37 ng·L−1和35.71—1236.21 ng·L−1)[24-25],窖水10种PFCs总浓度121.82—395.16 ng·L−1处于中间水平,波动范围相对较小. 而地下水中的PFCs含量普遍较低,如天津市郊区和北京市部分地区地下水PFCs总浓度范围分别为0.32—8.30 ng·L−1和N.D.—165.80 ng·L−1[26-27],均低于窖水和地表水中的PFCs浓度. 可以发现,不同水环境中PFCs含量的差异与其所处区域周围工业活动以及水体自身特点密切相关. PFCs在工业发达地区水环境中检出率高,地表水作为工业废水处理后的受纳水体,易于受到污染. 而地下水由于经过渗流,水质一般较好. 水窖中的PFCs则可能来自于大气污染物湿沉降和集雨面径流,受间接污染.
2.4 窖水中新污染物的健康风险评估
根据美国环保署优先控制污染物名单和本次窖水检测出的污染物浓度水平,选择DMP、DEP、DIBP、DBP、DEHP和DNOP 等6种PAEs进行健康风险评价,除了对DEHP进行致癌风险评价外,其它5种PAEs均进行非致癌风险评价. 根据相关研究得到DMP、DEP、DIBP、DBP、DNOP等 5种PAEs的参考剂量RfD 分别为 0.1、0.8、0.098、0.1、0.02 mg·(kg·d)−1;DEHP的致癌斜率因子SF为0.014 (kg·d)·mg−1[28]. 将所有参数代入“1.5”中的公式(1)—(3),得到冬夏两季窖水中这6种PAEs的致癌和非致癌风险值. 非致癌风险值HI值小于1时,该污染物浓度不会存在非致癌风险,反之该污染物将存在非致癌健康风险. 致癌风险R值小于10−6 时,不会对人产生致癌风险;R为
10−6—10−4 4.75×10−6— 5.11×10−4 1.11×10−7—1.86×10−7 1.11×10−7—1.35×10−3 3.39×10−8—2.99×10−7 根据美国卫生和公共服务部提出的人群饮用水途径全氟化合物每日耐受量和两季窖水中全氟化合物的检出情况,选择PFOA、PFOS、PFNA和PFHxS等 4种PFCs进行EDI值的计算,来评估健康风险水平. 经计算,冬季PFOA的EDI平均值为0.35
ng⋅(kg⋅d)−1 ng⋅(kg⋅d)−1 ng⋅(kg⋅d)−1 ng⋅(kg⋅d)−1 ng⋅(kg⋅d)−1 ng⋅(kg⋅d)−1 ng⋅(kg⋅d)−1 ng⋅(kg⋅d)−1 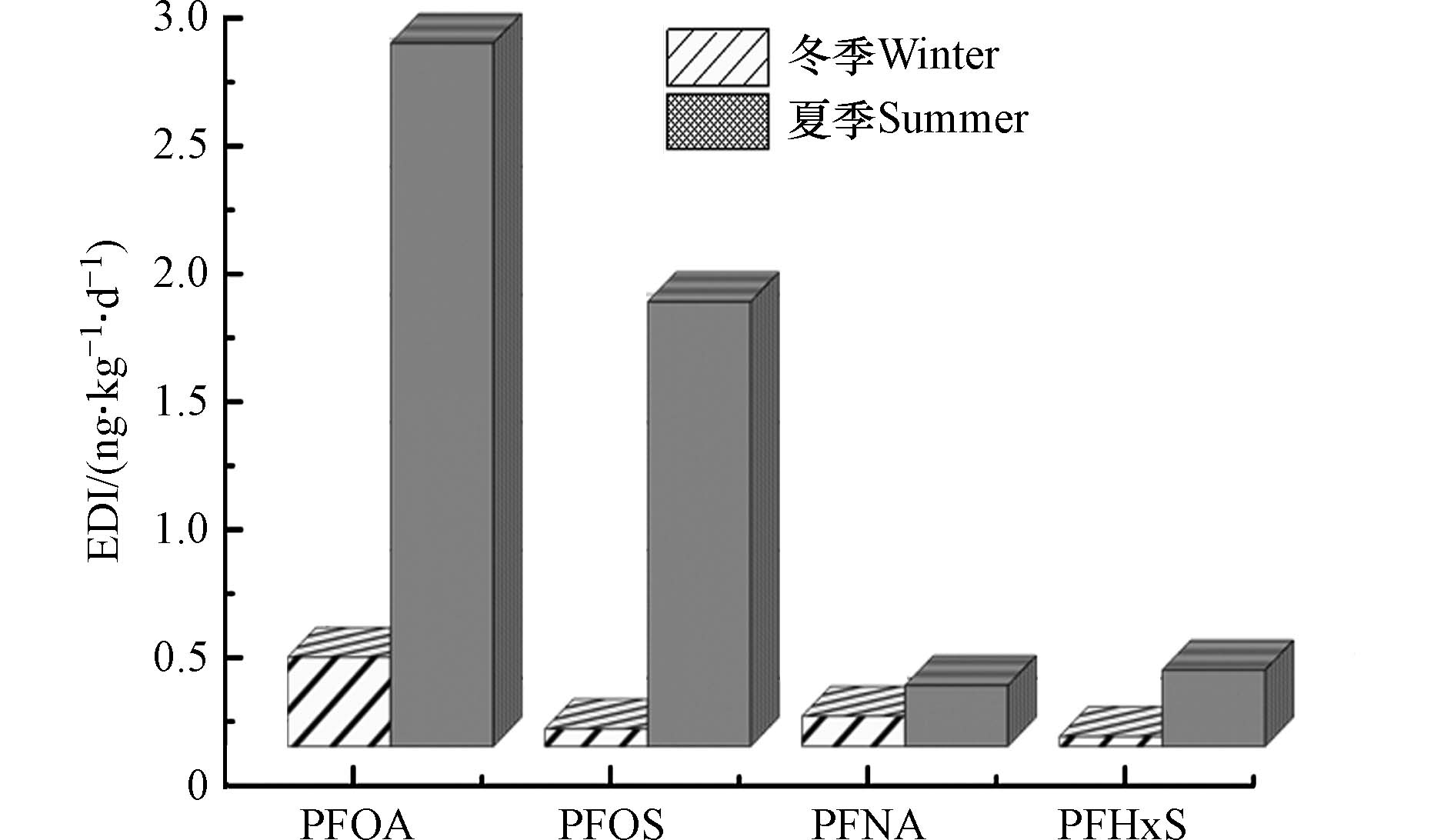 图 5 冬夏两季窖水样4种PFCs平均EDI值Figure 5. Average EDI of four PFCs of samples in winter and summer
图 5 冬夏两季窖水样4种PFCs平均EDI值Figure 5. Average EDI of four PFCs of samples in winter and summer3. 结论(Conclusion)
(1)甘肃该县冬、夏两季的窖水均呈现弱碱性(8.0≤pH≤9.7),浊度普遍偏高,且夏季雨期窖水浊度显著高于冬季.
(2)15种目标PAEs中共有13种在窖水中检出,冬季窖水中PAEs总浓度平均值为3.12 μg·L−1,夏季为4.56 μg·L−1. DBP和DIBP是各窖水样品的主要组成部分. 夏季雨期窖水中PAEs总浓度平均值明显高于冬季.
(3)17种目标PFCs中共有14种在窖水中检出,冬季PFCs总浓度范围为143.93—246.47 ng·L−1,夏季PFCs总浓度范围为275.90—405.51 ng·L−1. 窖水样品中的PFCs由PFCAs和PFSAs两部分组成,短链PFCAs中的全氟丁酸(PFBA)在PFCs中含量最高. 夏季雨期窖水PFCs浓度水平显著高于冬季,尤其是PFOA和PFOS在所有PFCs中的浓度占比显著上升.
(4)该县两季窖水中PAEs无致癌风险和非致癌风险;PFCs中的PFOA、PFOS、PFNA和PFHxS的平均EDI值均小于参考值. 此外,夏季窖水中PFOA和PFOS的EDI值大于冬季,且个别窖水EDI值超过参考值,应该予以重视.
-
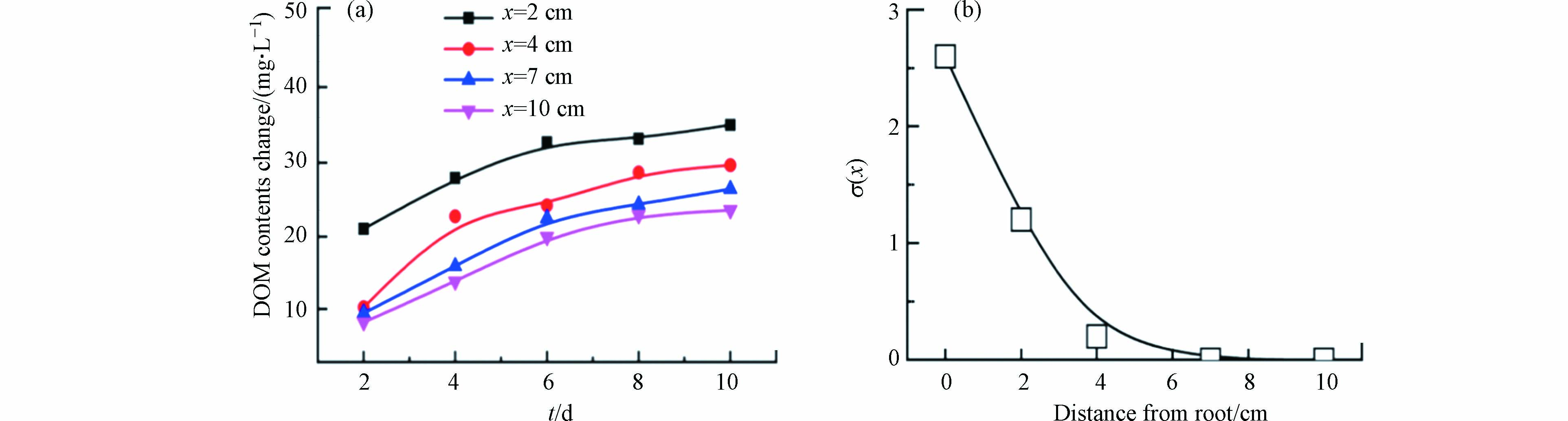
图 3 基质中DOM含量变化(a)和DOM的根部诱导生成率
σ(x) 与离假根距离的模型模拟(b)Figure 3. The change of DOM contents (a) in the substrate and the model simulation of the relationship between root induced generation rate of DOM
σ(x) and the distance to artificial root (b)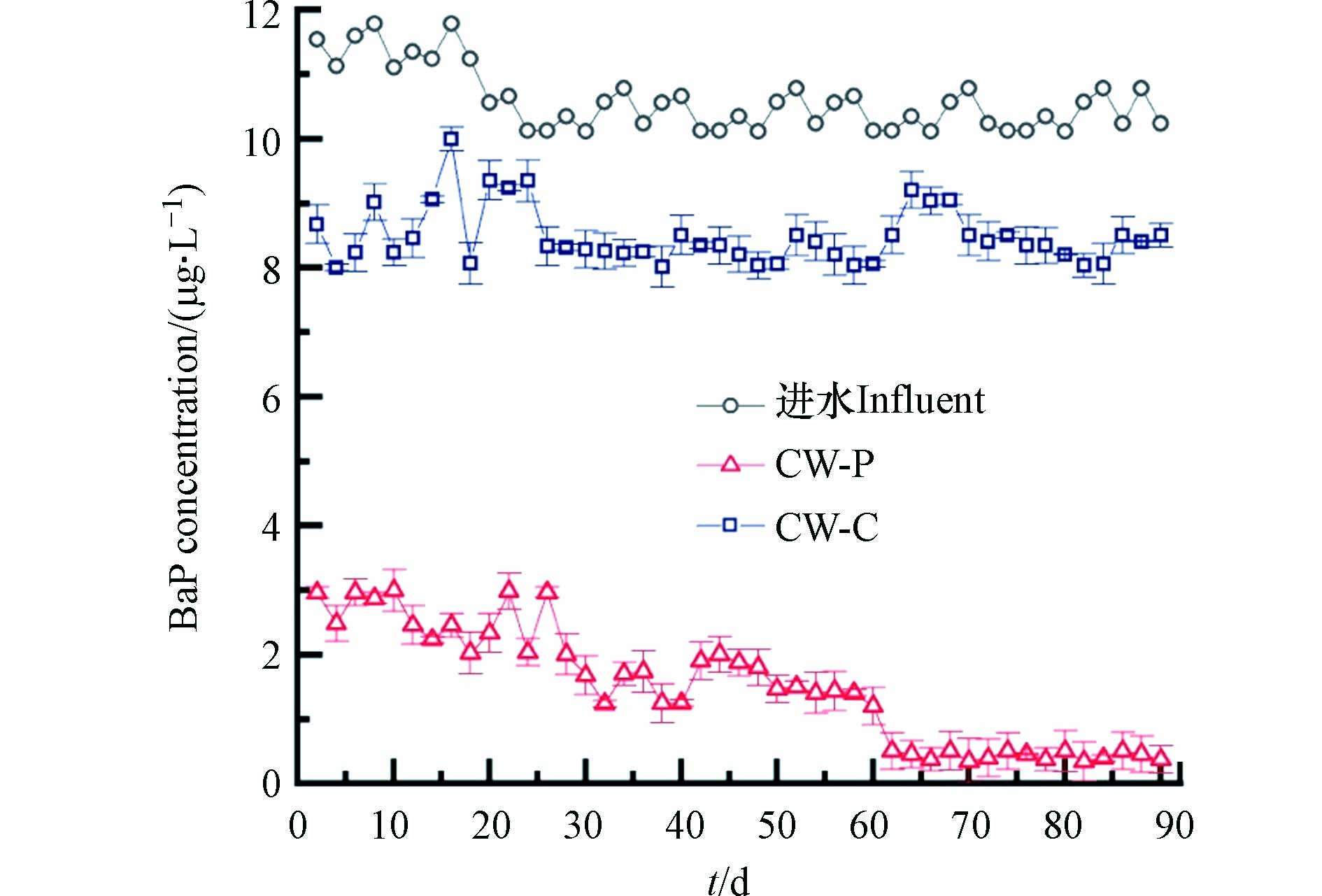
图 5 CW-P和CW-C废水中BaP浓度的变化
Figure 5. Variation of BaP concentrations with time in the effluents of CW-P and CW-C
表 1 菖蒲组织中BaP分配规律(*P≤0.05;**P≤0.01)
Table 1. Distribution of BaP in various tissues of Acorus calamus (*P≤0.05; **P≤0.01)
指标 Index 菖蒲 Acorus calamus 根系富集因子RCF 13.00** 茎叶富集因子SCF 3.30 迁移因子TF 3.60 RCF × 6.20** RCF × 14.60** SCF × 2.40
下载: 导出CSV
-
[1] 亓昕, 代嫣然, 王飞华, 等. 人工湿地去除特殊污染物的研究进展[J]. 环境科学与技术, 2017, 40(S1): 119-124. QI X, DAI Y R, WANG F H, et al. Research progress of constructed wetlands for treatment of particular pollutants[J]. Environmental Science & Technology, 2017, 40(Sup 1): 119-124 (in Chinese).
[2] 陈婧, 栾天罡, 罗丽娟. 烷基化多环芳烃的细菌降解研究进展 [J]. 环境化学, 2022, 9: 31-45. doi: 10.7524/j.issn.0254-6108.2020090801 CHEN J, LUAN T, LUO L. Research progress in bacterial degradation of alkylated polycyclic aromatic hydrocarbons [J]. Environmental Chemistry, 2022, 9: 31-45(in Chinese). doi: 10.7524/j.issn.0254-6108.2020090801
[3] MENG Y, LIU X H, LU S Y, et al. A review on occurrence and risk of polycyclic aromatic hydrocarbons (PAHs) in lakes of China [J]. Science of the Total Environment, 2019, 651: 2497-2506. doi: 10.1016/j.scitotenv.2018.10.162 [4] SU X, YUAN J, LU Z J, et al. An enlarging ecological risk: Review on co-occurrence and migration of microplastics and microplastic-carrying organic pollutants in natural and constructed wetlands [J]. Science of the Total Environment, 2022, 837: 155772. doi: 10.1016/j.scitotenv.2022.155772 [5] FOUNTOULAKIS M S, TERZAKIS S, KALOGERAKIS N, et al. Removal of polycyclic aromatic hydrocarbons and linear alkylbenzene sulfonates from domestic wastewater in pilot constructed wetlands and a gravel filter [J]. Ecological Engineering, 2009, 35(12): 1702-1709. doi: 10.1016/j.ecoleng.2009.06.011 [6] ZHAO C C, XU J T, SHANG D W, et al. Application of constructed wetlands in the PAH remediation of surface water: A review [J]. The Science of the Total Environment, 2021, 780: 146605. doi: 10.1016/j.scitotenv.2021.146605 [7] JIAO X C, XU F L, DAWSON R, et al. Adsorption and absorption of polycyclic aromatic hydrocarbons to rice roots [J]. Environmental Pollution, 2007, 148(1): 230-235. doi: 10.1016/j.envpol.2006.10.025 [8] ROPER J C, SARKAR J M, DEC J, et al. Enhanced enzymatic removal of chlorophenols in the presence of co-substrates [J]. Water Research, 1995, 29(12): 2720-2724. doi: 10.1016/0043-1354(95)00101-P [9] CHEN X H, HU Z, XIE H J, et al. Priming effects of root exudates on the source-sink stability of benzo[a]pyrene in wetlands: A microcosm experiment [J]. Journal of Hazardous Materials, 2022, 429: 128364. doi: 10.1016/j.jhazmat.2022.128364 [10] WANG Y D, OUYANG W, LIN C Y, et al. Higher fine particle fraction in sediment increased phosphorus flux to estuary in restored Yellow River Basin [J]. Environmental Science & Technology, 2021, 55(10): 6783-6790. [11] HAN) WENG Z, van ZWIETEN L, SINGH B P, et al. Biochar built soil carbon over a decade by stabilizing rhizodeposits [J]. Nature Climate Change, 2017, 7(5): 371-376. doi: 10.1038/nclimate3276 [12] GAO Y, WU S C, YU X Z, et al. Dissipation gradients of phenanthrene and pyrene in the rice rhizosphere [J]. Environmental Pollution, 2010, 158(8): 2596-2603. doi: 10.1016/j.envpol.2010.05.012 [13] CHENG Y, DING J, LIANG X Y, et al. Fractions transformation and dissipation mechanism of dechlorane plus in the rhizosphere of the soil-plant system [J]. Environmental Science & Technology, 2020, 54(11): 6610-6620. [14] KANG F X, CHEN D S, GAO Y Z, et al. Distribution of polycyclic aromatic hydrocarbons in subcellular root tissues of ryegrass (Lolium multiflorum Lam. ) [J]. BMC Plant Biology, 2010, 10: 210. doi: 10.1186/1471-2229-10-210 [15] SIVARAM A K, LOGESHWARAN P, SUBASHCHANDRABOSE S R, et al. Comparison of plants with C3 and C4 carbon fixation pathways for remediation of polycyclic aromatic hydrocarbon contaminated soils [J]. Scientific Reports, 2018, 8: 2100. doi: 10.1038/s41598-018-20317-0 [16] SOLÍS-DOMINGUEZ F A, WHITE S A, HUTTER T B, et al. Response of key soil parameters during compost-assisted phytostabilization in extremely acidic tailings: Effect of plant species [J]. Environmental Science & Technology, 2012, 46(2): 1019-1027. [17] TORN M S, TRUMBORE S E, CHADWICK O A, et al. Mineral control of soil organic carbon storage and turnover [J]. Nature, 1997, 389(6647): 170-173. doi: 10.1038/38260 [18] KEILUWEIT M, BOUGOURE J J, NICO P S, et al. Mineral protection of soil carbon counteracted by root exudates [J]. Nature Climate Change, 2015, 5(6): 588-595. doi: 10.1038/nclimate2580 [19] ZHALNINA K, LOUIE K B, HAO Z, et al. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly [J]. Nature Microbiology, 2018, 3(4): 470-480. doi: 10.1038/s41564-018-0129-3 期刊类型引用(0)
其他类型引用(3)
-

 点击查看大图
点击查看大图
计量
- 文章访问数: 4185
- HTML全文浏览数: 4185
- PDF下载数: 106
- 施引文献: 3




