


-
2021年中国社会化石燃料的使用比重高达89%,二氧化碳(CO2)排放量高达7500万吨,随着化石燃料的大肆消耗和CO2排放量的逐年攀升,人类面临着温室效应和能源危机的双重挑战[1]. 在“碳中和”的大背景下,如何将CO2收集与资源化利用成为关键问题. 常见的CO2利用手段包括地质利用(CO2驱油提高采收率)、化工利用(以CO2为原料生产化学品或燃料)和生物利用(利用微藻类植物进行CO2生物转化)[2-3].
生物电化学系统(Bioelectrochemical systems,BES)是近年来发展起来的,结合了化工利用和生物利用的一项CO2资源化技术,其中微生物电解池(Microbial electrolysis cells,MEC)可有效利用CO2转化为低碳燃料[1, 3-4]. 甲烷(CH4)作为最简单的有机物且含碳量最少的烃,是当今主要的燃料和化工原料. 相较于MEC制氢气(H2)所面临的H2燃点范围宽,常温常压难储存且价格昂贵(每kg 60—70元)等挑战,利用MEC将CO2转化为低碳燃料CH4,不仅极大地降低了CO2的排放并促进碳循环,且CH4作为一种清洁的低碳燃料,价格低廉(每kg 20—30元)且便于运输,可缓解紧张的能源需求[2, 4]. 与传统的厌氧消化产甲烷相比,MEC辅助CO2电产甲烷可将有机物氧化和产甲烷过程分开进行,减少废水对产甲烷菌的冲击,提高甲烷产量[2]. 其原理如图1所示,MEC阳极通过多种氧化反应(析氧反应或有机物的氧化分解)提供质子,质子通过质子交换膜到达阴极,电活性微生物在生物阴极上完成CO2电甲烷化,HCO3-/CO2常常可以被微生物利用产生甲烷[5-6].
近年来关于MEC的研究如雨后春笋破土而出(图2),研究人员多分布在不同国家,但关于MEC的研究方向大多集中在制H2;随着“双碳”目标的提出,MEC精炼废物质能源被开发,相应的电极材料、反应器构造类型、能耗等成为了研究者们关注的问题[2, 5, 7]. 因此,本文全面回顾了MEC电极材料、电压和电产甲烷菌等对CO2电甲烷化效能的影响,并重点讨论阴极电活性功能菌和胞外电子传递机制,以期为MEC在未来应用方面的技术挑战提供理论支撑.
-
MEC辅助CO2电产甲烷的效率受到多方因素的影响,为了获得更高的甲烷产率,需性能稳定的阴极催化剂、高效传递电子的电活性微生物和合适的外加电势等.
MEC阴极作为电活性微生物电子传递的主要介质和产甲烷菌附着的主要场所,是影响MEC性能的关键因素之一,MEC阴极材料一般具有良好的导电性、无微生物毒性、高比表面积、低过电位等[2-3]. 铂(Pt)是最早被用在MEC上的电极材料,其能量效率高达75%—80%,但由于Pt开采、提取过程中负面环境影响大,故亟需开发环境友好且效率与Pt相当的电极材料,碳基材料、金属基材料和复合材料近年来渐渐崭露头角[8-9]. 图3(a)总结了近5年MEC研究者利用不同电极材料所获得的最高甲烷产率和最大电流密度[2-4, 6-21](注:产甲烷率和电流密度均做归一化处理,气体体积按照标况下进行折算). 从数据统计看,最高甲烷产生速率为复合材料>金属基材料>碳基材料. 碳布、碳纸作为MEC生物阴极的基底可达到的最高甲烷产生速率仅为35—87 mL·L−1·d−1之间,低于碳毡和碳刷,可能是由于碳布这类平面电极比表面积低、微生物难以富集在材料内部所导致的,碳毡和碳刷则具有更大的比表面积,更有利于致密电活性生物膜的形成[2, 7]. 不锈钢和过渡金属(主要是镍(Ni))作为金属基材料则具有良好的导电性,其最高甲烷产生速率可达到135—350 mL·L−1·d−1,有研究证实不锈钢可引起电解液脱氢并释放H2,进而促进氢营养型产甲烷菌通过间接电子传递的方式产生CH4[9, 21]. 此外,Ni还是产甲烷菌的关键酶(甲基辅酶M)的组成金属元素,产甲烷菌相较于其他微生物更易占据Ni电极的阴极位点,进而提高甲烷产率[15]. Pt-碳毡和Pt-钛网等是常见的贵金属修饰复合阴极,但Pt修饰的贵金属电极也极易发生析氢反应产生H2,被氢营养型产甲烷菌利用,进而通过间接电子转移的方式刺激CO2快速向CH4转化[11-12, 16]. 电流密度可以侧面反映阴极单位表面积上单位时间内通过的电量,贵金属修饰的复合电极最大电流密度可达45—60 A·m−3,说明其导电性较好[2-4, 6-21]. 纳米管和纳米粒子等纳米材料通过电沉积、空气喷涂等方法修饰在碳基或金属基材料表面制备复合阴极材料,纳米修饰的复合材料通常具有良好的催化活性和稳定性,且多孔疏松的纳米形态有利于细胞的聚集和胞外聚合物的产生,其最高甲烷产生速率高达200—350 mL·L−1·d−1,纳米修饰的复合阴极材料也是近年来MEC阴极的研究热点[10, 14, 19],如磁铁矿/沸石纳米复合材料[22]、Ni/Co-NC纳米复合材料[23]、磁性GO/Fe3O4纳米复合材料[24]等,研究表明,纳米金属复合材料作为MEC阴极更有利于电活性微生物在其上形成厚且致密的生物膜,同时促进氢营养产甲烷通过直接电子转移的方式来来增强CO2到CH4的生物电化学还原作用[22-25].
电压调控对于MEC电产甲烷也尤为重要,图3(b)总结了近几年各个学者在不同电压下利用MEC产生的最高甲烷产生速率[5, 7, 9, 25-36]. −0.6—−1.0 V vs. AgCl是常见的阴极电压,其间的最高甲烷产生速率一般在15—100 mL·L−1·d−1之间[5, 7, 9, 25-36]. 阴极电位越负甲烷产量越高,可能是因为更负的电压提高了产电菌群的活性,并增强了其内部的电子传递效率,但是最佳的甲烷产量是在−0.9—−1.0 V vs. Ag/AgCl 的平衡电位下实现的,虽然研究者常通过施加更负电位的方法以克服CO2电甲烷生成的能垒,但研究表明电压高于−1.2 V时,产甲烷菌的生物膜会遭到破坏,生长活性和代谢速率会降低,所以MEC电压保持在一个合适的范围是非常重要的[5, 9, 28, 33].
除了电极材料和阴极电压外,MEC的结构也会间接影响微生物的电子传递性能及CH4产率[2, 10]. 图3(c)总结了近些年相关文献中,单室和双室MEC的最大产甲烷速率,单室MEC的最高甲烷产生速率一般在85—200 mL·L−1·d−1之间,而双室则在45—100 mL·L−1·d−1之间[3, 4, 7-8, 11-14]. 单室MEC由于无质子膜阻隔,阴阳两极间距较双室MEC更近,故内阻更小、物质间的传质阻力更低、电流密度更高,适合进行规模化CH4生产[13-14]. 但是单室MEC由于无质子膜阻隔,阴阳电解液相互接触,极易发生副反应,所以在单室MEC中如何实现目的产物CH4的高效定向转化是研究者们应该关注的重点问题[37].
此外,MEC中最大电流密度是CO2电甲烷化过程中相对电子传递效率的重要指标,与最高产甲烷速率之间也有一定关系,电流较高意味着更快的反应动力学,更低的生物膜电阻和电荷转移电阻,同时高电流响应意味着较好的生物膜活性和较低的电位损失[14, 16-17]. 从近5年的文献中可以发现,贵金属修饰的复合材料产生的最高电流密度最大(图3(a)),达到47—59 A·m−3之间,可能是贵金属修饰的复合电极更易生物膜的快速成型,从而产生更高的电流;较高的外加电压会产生较高的电流密度,MEC在−1.2 V vs. AgCl时的最高电流密度最大(图3(b)),达到7.5—11.4 A·m−3之间,可能是较高的电压输入给予MEC更高的电流流动;单室MEC的电流密度高于双室MEC(图3(c)),可能是单室MEC内阻小、传质阻力低[13-14].
-
MEC辅助CO2电产甲烷的运行关键在于阴极腔室内的电活性功能微生物,它们承担着电子转移等重要工作,被誉为MEC的“心脏”. 电活性功能微生物可借助外源电势差突破超电势与内电阻的限制,摄取来自电极表面的电子,还原为CO2为低价态的有机物(如CH4),以及将氧化态物质还原成还原态无机物[38]. 为了提高产MEC产甲烷的性能,了解电活性功能微生物的类型、群落组成和微生物之间的相互作用也至关重要. 能够还原CO2产甲烷的微生物是一种重要的电活性功能微菌,在分类学上,属广古菌门(Euryarchaeota),其包括5个目(Methanobacteriales、Methanococcales、Methanomicrobiales 、Methanopyrales和 Methanosarcinales)、10个科(Methanobacteriaceae、Methanocaldococcaceae、Methanococcaceae、Methanocorpusculaceae、Methanomicrobiaceae、 Methanopyraceae、Methanosaetaceae 、Methanosarcinaceae、Methanospirillaceae、Methanothermaceae)及31个属,这些微生物对氧气极为敏感,因此大多是都需要在严格缺氧的环境中培养[39]. CO2电产甲烷菌常利用的底物类型为甲基类、H2和乙酸,对应的产甲烷菌分别可以被命名为甲基营养型(方程式1)、氢营养型(方程式2)和乙酸营养型(方程式3)产甲烷菌[39].
产甲烷菌的主要生理特征(碳源、温度和pH范围)总结在表1中,大多数产甲烷菌是嗜温细胞,在pH值约为7时生长最佳[40]. 形态上,典型的产甲烷菌有甲烷杆菌属(Methanobacterium)、甲烷球菌属(Methanosarcina)和甲烷丝状菌属(Methanosaeta)(图4)[40-41]. 淡水沉积物、泥炭沼泽、稻田和污水消化池等为产甲烷菌最适宜生长的环境[42].
-
MEC中电子是如何从阴极电极表面转移用于CO2还原的过程受到学界的关注,一种解释是在MEC的阴极表面产生了H2,附着在阴极的电产甲烷菌以H2/CO2为底物合成CH4,还有一种解释是电产甲烷菌可以直接从阴极表面获得电子还原CO2产生CH4 [46-47]. 培养了一段时间后的MEC的阴极将会被一层厚厚的电活性生物膜覆盖,电活性生物膜的活性外层负责转移电子产生电流,死的内层将作为导电基质发挥作用[46, 49]. 这种电活性生物膜是MEC电产甲烷中微生物和电极之间电子转移过程的重要一环,可以使得电产甲烷菌之间通过有效的细胞“交流”促进细胞间的活动与代谢,利用阴极电极表面的电子产生CH4[50].
在MEC还原CO2产甲烷的体系中,电产甲烷微生物间通过种间电子传递的合作方式对底物及阴极表面的电子进行利用,形成复杂且高度组织化的多细胞和多物种结构,达到互营共生的效果. 一些胞外活性菌会先把大分子有机物分解为小分子有机物,电产甲烷微生物通过利用小分子产生甲烷[46, 49]. 在MEC生物阴极表面,根据电子传递路径的不同可以分为直接种间电子传递(direct interspecies electron transfer,DIET)和间接种间电子传递(mediated interspecies electron transfer,MIET)两种方式[50]. 在DIET中,微生物通过细胞表面的蛋白质(如c型细胞色素)或细胞附属物(如菌毛或纳米线)与不溶性电子受体(如电极)建立直接接触,在MIET中,微生物利用可溶性氧化还原活性化合物作为电子穿梭的媒介,以介导细胞表面暴露的导电蛋白和不溶性电子受体之间的电子转移[50, 51].
H2是最早被发现的能够进行间接种间电子传递的中间载体(图5a),且MIET过程中产甲烷菌的耗H2速率与NAD+/NADH、FAD/FADH2、Fd(ox)/Fd(red)和F420/F420-H2等多种内源性氧化还原介质有关[49]. 辅酶M和辅酶B形成混合二硫化物作为整个厌氧产甲烷呼吸链的电子受体,分子氢、还原型辅酶F420和还原型铁氧还蛋白作为电子供体;A1A0-ATP催化合成的驱动力型ATP合成酶、甲基转移酶和甲酰基甲烷呋喃脱氢酶是与产甲烷过程有关的能量转导酶. 其中,甲基转移酶是一种独特的、可逆的钠离子泵,它将甲基转移与Na+跨膜转运相结合,电子受体菌则利用电子供体菌提供的H2还原辅酶(F420)和铁氧还蛋白(Fd(ox)),进而将CO2还原成甲烷[52].
除H2外,甲酸也可作为电子载体介导MIET的发生,Rotaru等[53]在Pelobacter carbinolicus和Geobacter sulfurreducens共培养体系中发现P. carbinolicus和不能利用H2但能利用甲酸的G. sulfurreducens转基因菌株共培养时很容易生长,但若以乙醇作为电子供体、富马酸盐作为电子受体时,两菌没有互营共生且生长受到抑制,证明G. sulfurreducens的甲酸脱氢酶(fdnG)基因可有效弥补氢化酶的缺失,P. carbinolicus通过H2/甲酸盐的种间转移而不是DIET与G. sulfurreducens交换电子.
除H2和甲酸外,具有氧化还原性能的电子介质,如黄素、吩嗪和醌类等,也可介导电产甲烷微生物的MIET[54](图5b). Liu等[55]构建了一个含有S. oneidensis MR-1和可产生核黄素的枯草芽孢杆菌RH33联合体,发现RH33产生的高浓度核黄素可被S. oneidensis MR-1用于提高生物电. Engel等[56]联合培养G. sulfurreducens和S. oneidensis,发现混合培养优于单独培养,可能由于S. oneidensis代谢产生的氢化酶和黄素被G. sulfurreducens用于增强直接电子转移,促进该生物体能够在阳极表面形成更厚的生物膜,同时增加电流密度. 除核黄素之外,腐殖质也具有独特的电子转移能力,能在介导微生物种间电子转移方面发挥重要作用,G. metallireducens和S. alga可利用蒽醌-2,6-二磺酸盐作为电子受体进行厌氧呼吸[57]、维持细胞生长,研究显示,低剂量的蒽醌-2-磺酸盐(50 µmol·L−1)可得到最大的甲烷产量[58].
除了间接电子传递,近年来,研究者发现电活性微生物可通过菌体DIET传递电子,实现互营共生. Summers等[59]在G. metallireducens和G. sulfurreducens的共培养体系中首次证实了DIET的存在,微生物种间DIET机制通过导电菌毛[60]、导电材料[61]与功能蛋白复合物[62]3种方式实现.
导电菌毛是一类由电活性微生物合成、具有导电性的纤维状的蛋白质细丝,通过产甲烷微生物的导电菌毛,微生物胞内代谢产生的电子可以长距离输送到胞外受体或其他产甲烷微生物,实现了微生物-胞外环境的沟通交流[60](图5c). 微生物纳米导线最初通过导电探针原子力显微镜发现,G. sulfurreducens及S. oneidensis,蛋白PilA组成的Ⅳ型菌毛能够实现快速、长距离的电子传递,将电子从细胞表面转移到 Fe (Ⅲ) 表面. 近年来的研究表明,大部分的导电菌毛的主要成分为细胞色素OmcS、PilA蛋白起到调节omcS和其他多血红素细胞色素的分泌的作用[63]. 除了OmcS,常见的功能蛋白复合物还有血红素细胞色素c(MtrC和OmcA),可转运细胞间的蛋白质,实现细胞间的交流协作[64](图5d). 当G. sulfurreducens中血红素细胞色素c基因被敲除后,菌体生长受到抑制,这表明细胞色素所组成的跨膜电子通道主导了DIET过程[62]. 敲除MtrC和OmcA后的S. oneidensis MR-1 细胞,其在电极上的覆盖量和在电极上产生的电流比未敲除基因的细胞减少80%,此外,分离并纯化两种基因后发现,两种外膜细胞色素c可以结合金属氧化物如Fe2O3,并能将电子直接转移到Fe2O3的电极表面,Xiong等[65]还利用光学波导光模型和蛋白膜伏安法测得OmcA与Fe2O3的结合力为1.2—2.6 nmol·cm−2,总之,电活性菌外膜上的MtrC和OmcA可以相互接触通过其暴露在蛋白质表面的血红素将电子直接传导给胞外电子受体[60]. Zhang等在MEC电活性系统中检测到了产甲烷菌菌毛状的纳米线的存在,并认为其有电子转移的能力.
导电材料构建的“微生物-电极”的电子传递系统也是近年来DIET的研究热点,导电材料的加入可明显提升多种厌氧系统的效率并作为导体加强产甲烷微生物种间DIET[61,66-67](图5e). Liu等[61]通过在OmcS蛋白缺失的突变种G. sulfurreducens加入纳米磁铁矿使其弥补了细胞外电子交换中菌毛相关c型细胞色素的缺乏,恢复DIET的能力. Park等[66]将颗粒活性炭加入到厌氧消化的小瓶中使得总甲烷产量比对照组升高了75%,并通过宏基因组学证明了产甲烷总量与效率的提升是颗粒活性炭的加入改变了微生物群落结构并调整了DIET的相关基因的丰度两方面同时作用的结果. 此外,无导电菌毛或导电性能较差的菌株亦可以在导电材料表面与其他微生物建立联系,从而达到互营共生,Liu等[61]研究发现,磁铁矿纳米颗粒(20—50 nm)吸附在导电菌毛上,通过作为菌毛上OmcS的替代物,增强突变种Geobacter的DIET能力. 但有学者提出磁铁矿等导电材料只接受电子,却不会将电子传递给其他微生物[67],因此研究导电材料在电子传递中的作用时,还应该综合评估对产甲烷微生物的作用.
尽管,产甲烷微生物在碳氮磷的全球循环、环境污染物的修复以及各种生物能源策略中发挥着关键作用,已经有研究将电活性功能菌的电化学行为用于提高甲烷生产,减少启动时间,并提高厌氧消化系统的稳定性[68-69]. 在混合菌种共培养体系中平衡代谢过程的潜在驱动力可能与菌种之间的种内和种间的密切交流有关,然而电活性微生物之间传递偶联尚未得到广泛探索,因此了解并识别微生物通讯通信机制对于MEC电产甲烷的应用至关重要,并为工程系统的潜在应用开辟了无数可能性. 依靠微生物的本能来增强胞外电子传递可能是增强微生物电活性的一种有前途的方法,但仍需大量研究来解开电活性生物相互“通信”的过程.
-
近年来,基于电催化耦合的技术近年来越来越多见,学者将多个处理方法有机融合到一起,达到耦合多个生化过程到一个装置、多工艺协同作用产甲烷的效果,如MEC-AD、MEC-UASB、M-MEC-AD等. 图6总结了部分典型MEC耦合其他系统产甲烷的工艺.
虽然近年来MEC相关的研究势如破竹,但MEC的大规模生应用仍面临诸多问题. 首先是成本问题,MEC的运行需要传质效率高的阴极导电材料,如活性炭颗粒(每t 2000—5000 元)、碳布(每m3 100—250 元)、生物炭(每t 1000—2000 元)、不锈钢(每m3 15—30 元)、钛网(每m3 80—120 元)、铂片(每g 300—400 元)等;虽然相较于其他材料而言,碳基材料成本较低,相关研究也比较充分,但其电催化性能不如金属电极. 金属基材料导电性能优越,但其表面光滑,不利于微生物的附着且高昂的价格也限制了其商业化的应用[4, 10, 14, 18]. 故而用复合材料修饰电极表面是近年来的研究热点,贵金属或纳米修饰的阴极材料不仅导电性好,且表面积大、易于微生物附着,但其制备过程需要用到PTFE等粘合剂,极易造成催化材料的活性位点被覆盖等问题,为此开发高性能、稳定和低成本的阴极材料以降低过电位和整体内阻并优化负载方法是未来MEC阴极材料的大势所趋[10, 14],如可以根据产甲烷菌的生理特性开发金属-纳米-碳基复合材料,并观察电活性微生物的原位长势及富集情况,改善其催化活性和电子传递方式,以提高产甲烷效果.
再者,现阶段单室MEC和双室MEC规模化放大生产中比对研究较少. 虽然单室MEC结构简单无质子膜阻隔,但阴阳电解液相互接触,极易发生副反应或短路现象,而大规模应用时副反应的发生将更加具有不可控性;双室MEC虽然有质子膜将阴阳极分离,但是极易造成阴阳两极的浓差极化,且双室MEC的阴阳两极相距较远,物质间传质阻力较大[4, 6-9, 11-15, 26-34, 37]. 故未来应通过实时监控、定点取样,推进MEC规模化生产应用的研究.
此外,生物阴极虽然结构简单,但阴极与产甲烷菌之间的电子传递和相互作用基质尚不明确,如不同导电材料的投加对于产甲烷MEC性能的影响、参与直接电子传递产甲烷代谢途径的酶除了还原酶Fdox、F420等是否还有未被发现的酶、不同种类的产甲烷菌生态位之间是否存在协同或竞争的关系、执行电子传递的主要控制基因又是什么等. 故未来应采用更加先进的技术手段(宏基因、宏蛋白和多组学生物技术等),分析MEC产甲烷阴极微生物群落结构的动态变化规律,对产甲烷菌的特性进行定向调控,优化MEC性能.
目前,MEC产甲烷的研究主要以小瓶实验为主,中试规模运行效能仍然未知. 在不少MEC耦合工艺的中试试验中,均检测到在长期运行期间由于膜污染、膜形变、废水中过量的生物质等原因,系统性能下降的情况[70-71],如Wang等[32]构建的中式规模的MEC-UASB产甲烷系统,在较低的HRT情况(HRT=2.7 h)下,VFAs积累较快,甲烷产量下降显著. 故而未来的研究中应该着重于MEC工业化长期化运行期间可能遇到的问题如膜污染、膜形变、废水中过量的生物质等问题,放大MEC反应器将是其未来研究方向的主导趋势,以推进MEC工程化应用.
-
为了实现低碳社会和可再生能源利用的目标,许多国家正进行各种技术和产业革命,发展绿色能源和低碳产业.
(1)甲烷是比较理想的清洁能源,通过微生物电解池(MEC)辅助CO2电甲烷化可以强化CO2的资源化,MEC也可以和相关技术联用,实现减碳降碳的同时对污染物进行减量化与无害化,应用前景广阔.
(2)现阶段的MEC电甲烷化依然存在长期批次试验不足、单次循环时间短、甲烷产量及纯度不高、产甲烷菌生长缓慢等问题. 故而开发连续的、系统的CO2电甲烷化反应器,优化生物阴极材料,调节微生物最适宜的环境条件迫在眉睫.
(3)未来的MEC电甲烷化研究应该将理论研究和实际应用进行更深入的结合,开发出:新型CO2电甲烷化生物反应器;克服CO2电甲烷化实验周期短、不稳定的难题;优化反应的条件(电极物理化学性质、工作温度、水力停留时间和有机负载率等);解析MEC中电活性功能微生物菌群、能量代谢理论体系及产甲烷菌的时空演替、种间信息交流方式,通过调控产甲烷菌电子传递的主控基因和胞外电子界面传导路径,提高CO2燃料化动力源的活力. 通过构建稳定、高效的CO2电甲烷化生物反应器,助力CO2减排与碳中和技术,以期为电甲烷化由基础走向工程提供新思路.
微生物电解池催化CO2电转化为甲烷:影响因素、电子传递和展望
Microbial electrolytic cell catalyzed electroconversion of CO2 to CH4:Influencing factors, electronic transmission, outlook
-
摘要: 化石燃料作为能源供应的主要来源,燃烧导致大量CO2的释放和温室效应,CO2的捕获和再利用越来越受到人们的关注. 微生物电解池(MEC)作为一种新的CO2再利用技术,可通过将电活性微生物与电化学刺激相结合,将CO2通过生物电化学作用回收为低碳燃料(如CH4),从而实现CO2固定和能量回收. 尽管近年来MEC领域有较多研究,但仍然存在许多问题阻碍了该技术的规模化和产业化. 本文梳理了CO2电化学产甲烷的工作原理、性能影响的关键因素、生物阴极电活性功能微生物及其胞外电子传递机制、电催化耦合技术的最新研究进展,提出了MEC辅助CO2电甲烷化技术的未来研究需求和挑战.
-
关键词:
- 微生物电解池(MEC) /
- CO2电甲烷化 /
- 阴极材料 /
- 电活性功能菌 /
- 胞外电子传递机制
Abstract: Fossil fuels have been the main source of energy supply, and their combustion leads to the release of a large amount of CO2 and the greenhouse effect. The capture and reuse of CO2 have attracted more and more attention. Microbial electrolysis cell (MEC), a new CO2 reuse technology, can achieve CO2 fixation by combining electroactive microorganisms with electrochemical stimulation to recycle CO2 into low-carbon fuels (such as CH4) through bioelectrochemical action and energy recovery. Although there have many researches in the field of MEC in recent years, there are still many problems which hinder the scale and industrialization of this technology. This paper compares the working principle of CO2-electrochemical methanogenesis, key factors affecting performance, bio-cathode electroactive functional microorganisms and their extracellular electron transfer mechanism, and the latest research progress of electrocatalytic coupling technology. We also presents the future research needs and challenges of MEC CO2-electrochemical methanogenesis technology. -
全氟烷基酸(perfluoroalkyl acids, PFAAs)因其化学稳定性、热稳定性和表面活性良好而被广泛应用于工业生产和人类生活,其中全氟辛烷羧酸(perfluorooctanoic acid, PFOA)和全氟辛烷磺酸(perfluorooctane sulfonate, PFOS)是用途最广的两种化合物[1]。近年来,PFAAs污染受到了国内外学者的广泛关注,PFAAs在全球各区域多种环境介质中被普遍检出,尤其是在水环境中[2]。我国主要流域河流河表层水中检出的∑PFAAs高浓度分别为松花江流域8 ng·L−1、辽河流域128 ng·L−1、海河流域174 ng·L−1、黄河流域79 ng·L−1、淮河流域25 ng·L−1、太湖流域330 ng·L−1、长江流域362 ng·L−1、珠江流域62 ng·L−1[3-9],小清河流域报道∑PFAAs高浓度为325280 ng·L−1[10],在日本东京湾农村地区地下水中检出PFOA、PFOS和PFNA浓度分别为1800、990、620 ng·L−1[11]。研究表明PFAAs高值区域均存在工业污染源,且PFAAs能在环境中累积和远距离迁移,具有生物放大效应,潜在的生态风险和居民健康风险较高。Lu等[4]报道了我国辽河流域PFOA(0.38—74 ng·L−1)的潜在水生生物生态风险较高,并且备受关注的小清河流域下游水中PFOA的潜在生态风险更高[10]。Sun等[8]发现PFOA和PFOS对上海地区18—44岁成人的潜在健康风险较大。在我国东南主要流域有大型氟化工生产加工基地,加之环杭州湾是制造业的热点区域,PFAAs生产、消费量较大,但目前对该流域PFAAs研究不够完善,只有少数河流受到了关注[12-14],缺少对整个流域主要河流水体中PFAAs的研究和风险评价。
水环境中PFAAs的污染来源广,包括工业源、生活源、交通源和农业源等,PFAAs迁移途径有大气、水流、食物链等[15]。目前关于持久性有机污染物的来源解析方法有指纹图谱法、比值法、多元统计分析法、同位素分析法等[16],但污染物与受体环境复杂多样,单一源解析方法均存在一定的局限性,需采用多种方法组合进行全面分析。
对于PFAAs的风险评价,由于缺乏生态毒理学数据,大多数评价方法只关注了PFOA、PFOS的潜在生态风险和健康风险,存在较大的不确定性。德国环境署在HRIV(precautionary health related indication value)中制定了一项标准以弥补毒性数据的差异,不确定性分析结果表明该方法优于美国环保局开发的基于ADI(acceptable daily intake)和TDI(Tolerable daily intake)的评价方法[17-18]。饮水和饮食一直被认为是全氟烷基酸(PFAAs)的主要摄入途径之一[19],采用商值法能简单快速的判断是否潜在生态风险和健康风险,可为后续研究和风险调控提供可靠支撑。
本研究探究了我国东南主要河流表层水体中PFAAs的赋存特征和估算PFAAs的入海通量,采用组合方法分析了PFAAs污染来源,并分别评价了PFAAs对水生生物的生态风险和潜在的人体健康风险,研究结果有助于理解我国东南河流PFAAs的环境行为和潜在风险,为整个流域水资源保护、治理和合理利用提供科学支撑。
1. 材料与方法(Materials and methods)
1.1 研究区域概况
中国东南河流处在长江流域和珠江流域之间,如图1所示,采样河流均位于福建省和浙江省,且有潜在的工业排放源。浙江省陆域面积约10万km2,大陆海岸线约2200 km,人口密度约为550人·km−2,年平均降雨量1640.3 mm(2018年),地形自西南向东北呈阶梯状倾斜,西南以山地为主,中部以丘陵为主,东北部是低平的冲积平原,主要山脉呈西南-东北走向,自北而南分成3支。水系由北向南为苕溪、钱塘江、甬江、灵江、瓯江、飞云江、鳌江等。福建省陆域面积约12万km2,陆地海岸线3752 km, 年平均降雨量1566.0 mm(2018年),地势西北高东南低,闽西大山和闽中大山带斜贯全省,山带之间有互不贯通的河谷和盆地,东部沿海是丘陵、台地和滨海平原,主要水系由北向南有闽江、萩芦溪、九龙江、鳌江、晋江、木兰溪等。
 图 1 中国东南主要河流样点分布Figure 1. Distribution of sample points along major rivers in southeast China(R1—R13分别是苕溪、钱塘江、甬江、灵江、瓯江、飞云江、鳌江、闽江、萩芦溪、木兰溪、晋江、九龙江、诏安东溪;P1—P10是PFAAs潜在的生产和消费企业)(R1—R13 represent Tiao river, Qiantang river,Yong river, Ling river, Ou river, Feiyun river, Ao river, Min river, Qiulu river, Mulan river, Jin river, Jiulong river, Zhaoan river, respectively; P1—P10 represent PFAAs production and consumer plants)
图 1 中国东南主要河流样点分布Figure 1. Distribution of sample points along major rivers in southeast China(R1—R13分别是苕溪、钱塘江、甬江、灵江、瓯江、飞云江、鳌江、闽江、萩芦溪、木兰溪、晋江、九龙江、诏安东溪;P1—P10是PFAAs潜在的生产和消费企业)(R1—R13 represent Tiao river, Qiantang river,Yong river, Ling river, Ou river, Feiyun river, Ao river, Min river, Qiulu river, Mulan river, Jin river, Jiulong river, Zhaoan river, respectively; P1—P10 represent PFAAs production and consumer plants)1.2 样品采集
样品采集自13条入海河流和部分港口区域,除苕溪为2个采样点外,其它河流采集至少3个采样点,包括河口样点,以及上游20 km、40 km处的样点,样点分布如图1所示。此次样品采集于2018年8—9月完成,共采集样品52份,每个样点设置3个平行样,均使用经润洗的1 L聚丙烯塑料瓶收集表层水,运输途中避光低温(4 ℃)保存样品,分析前样品保存于实验室(−20℃)冰箱。
1.3 样品分析
水样采用固相萃取法进行前处理后用液相色谱串联质谱(Agilent 1290-6460)检测分析了常见的17种PFAAs,包括13种PFCAs(C4—C18)和4种PFSAs(C4—C10),质谱参数见表1所示。分析前所有样品冷冻保存于−20 ℃冰箱,因此待分析样品需要在室温(空调20 ℃)自然解冻并静置过夜,每个样品准确量取400mL上清液进行分析。样品前处理关键步骤如下[20]:每个分析样品添加5 ng PFAAs混合内标物,如表1所示;依次加入4 mL 0.1%氨水甲醇、4 mL甲醇和4 mL超纯水活化Oasis WAX cartridges柱子(Waters公司,美国);固相萃取(SPE)上清液;添加4 mL 25 mmol·L−1醋酸铵(pH = 4)洗涤固定PFAAs,冷冻干燥24 h,依次添加4 mL甲醇和0.1%氨水甲醇洗脱Oasis WAX小柱并将洗脱液转移至15 mL离心管;高纯氮浓缩洗脱液并定容至0.5 mL,用0.2 μm GHP针式过滤器(聚丙烯生物膜,Pall公司,美国)过滤并转移至进样瓶中待测。
表 1 17种全氟烷基酸的质谱参数和质量控制Table 1. Mass spectrometry parameters and quality control of 17 PFAAs化合物Compounds 英文名称English name 缩写Abbreviation 登记号CAS No 分子式Molecular formula 母离子/子离子MS/MS transition(m/z)* 回收率/%Recovery(mean ± SD) 标准曲线R2 Correlation coefficient 检出限/ (ng·L−1)LOD 定量限/ (ng·L−1)LOQ 全氟烷基羧酸 Perfluorocarboxylic acid PFCAs CnHF2n−1O2 全氟丁酸 Perfluorobutyric acid PFBA 375-22-4 C4HF7O2 213.0/169.1 110.6 ± 6.3 0.9986 0.250 1.250 全氟戊酸 Perfluoropentanoic acid PFPeA 2706−90-3 C5HF9O2 263.0/218.9 103.9 ± 9.4 0.9921 0.125 0.375 全氟己酸 Perfluorohexanoic acid PFHxA 307-24-4 C6HF11O2 313.0/269.0 110.9 ± 6.7 0.9957 0.050 0.175 全氟庚酸 Perfluoroheptanoic acid PFHpA 375-85-9 C7HF13O2 363.0/318.9 118.3 ± 8.6 0.9950 0.075 0.250 全氟辛酸 Perfluorooctanoic acid PFOA 335-67-1 C8HF15O2 413.0/368.9 105.7 ± 1.9 0.9940 0.025 0.125 全氟壬酸 Perfluorononanoic acid PFNA 375-95-1 C9HF17O2 463.0/419.0 112.9 ± 3 0.9975 0.025 0.125 全氟癸酸 Perfluorodecanoic acid PFDA 335-76-2 C10HF19O2 513.0/468.9 111.7 ± 10.7 0.9982 0.050 0.175 全氟十一烷酸 Perfluoroundecanoic acid PFUnDA 2058−94-8 C11HF21O2 563.0/519.0 104.6 ± 6 0.9989 0.100 0.250 全氟十二烷酸 Perfluorododecanoic acid PFDoDA 307-55-1 C12HF23O2 613.0/569.0 106.3 ± 9.2 0.9978 0.050 0.150 全氟十三酸 Perfluorotridecanoic acid PFTrDA 72629−94-8 C13HF25O2 662.9/619.0 94.8 ± 6.7 0.9974 0.075 0.225 全氟十四酸 Perfluorotetradecanoic acid PFTeDA 376-06-7 C14HF27O2 713.1/669.0 80.2 ± 10.9 0.9961 0.075 0.200 全氟十六烷酸 Perfluorohexadecanoic acid PFHxDA 67905−19-5 C16HF31O2 813.0/769.0 100.3 ± 27.8 0.9986 0.075 0.225 全氟十八烷酸 Perflfluorooctadecanoic acid PFODA 16517−11-6 C18HF35O2 913.0/869.0 101.8 ± 14.5 0.9976 0.075 0.250 全氟烷基磺酸 Perfluorinated sulfonic acid PFSAs CnHF2n+1O3S 全氟丁烷磺酸 Perfluorobutane sulfonate PFBS 375-73-5 C4HF9O3S 299.0/80.0 109.1 ± 6.2 0.9906 0.050 0.125 全氟己烷磺酸 Perfluorohexane sulfonate PFHxS 355-46-4 C6HF13O3S 399.0/80.0 104.9 ± 6.2 0.9920 0.050 0.100 全氟辛烷磺酸 Perfluorooctane sulfonate PFOS 1763−23-1 C8HF17O3S 498.9/80.0 105.2 ± 12.5 0.9965 0.050 0.125 全氟癸烷磺酸 Perfluorodecane sulfonate PFDS 335-77-3 C10F21O3S 599.0/79.9 101.6 ± 22.9 0.9927 0.125 0.300 内标化合物 Internal standards 13C4全氟丁酸 13C4 Perfluorobutanoic acid 13C4 PFBA 217.0/172.0 13C2全氟己酸 13C2 Perfluorohexanoic acid 13C2 PFHxA 315.0/270.0 13C4全氟辛酸 13C4 Perfluorooctanoic acid 13C4 PFOA 417.0/372.0 13C5全氟壬酸 13C5 Perfluorononanoic acid 13C5 PFNA 468.0/423.0 13C2全氟癸酸 13C2 Perfluorodecanoic acid 13C2 PFDA 515.0/470.0 13C2全氟十一烷酸 13C2Perfluoroundecanoic acid 13C2 PFUnDA 565.0/520.0 13C2全氟十二烷酸 13C2 Perfluorododecanoic acid 13C2 PFDoDA 615.0/570.0 18O2全氟己烷磺酸 18O2 Perfluorohexane sulfonate 18O2 PFHxS 403.0/103.0 13C4全氟辛酸 13C4 Perfluorooctane sulfonate 13C4 PFOS 503.0/99.0 注:登记号为 Chemical abstracts service registration number USA,缩写为 CAS No;*定性离子,仪器扫描捕捉目标离子参数. Note: The registration number is Chemical abstracts service registration number of USA, abbreviated as CAS No; * Qualitative ion, the instrument scans to capture the target ion parameters. | Show Table DownLoad:
CSV
DownLoad:
CSV
色谱条件:液相色谱串联质谱所用色谱柱为Agilent ZORBAX Eclipse Plus C18 (2.1 mm×100 mm, 3.5 μm),前接0.3 μm在线过滤器(Agilent 1290)。控制柱温为40 ℃,进样量为5 μL。用2 mmol·L−1乙酸铵(A)和乙腈(B)作为流动相,流速控制在0.3 mL·min−1,初始体积比例设置为80% A 和20% B,保持0.5 min,经16 min比例变为10% A 和90% B,保持4 min post time 后,回到初始状态。质谱条件:离子源为电喷雾离子源负离子监测模式(ESI-),雾化温度为350 ℃,雾化器压力为40 psi,辅助气(N2)流量为9 L·min−1,毛细管电压为3500 V。
质量控制(QA/QC)如下:PFAAs自然标和内标纯度均 > 98%,购买自加拿大威灵顿公司 (Wellington Laboratories, Guelph, ON, Canada);醋酸铵纯度 > 98%, 购自西格玛公司(Sigma-Aldrich Co, St. Louis, MO, USA);甲醇、乙腈均为色谱级,购自赛默飞公司(Thermofisher, MA, USA);超纯水由美国 Millipore公司的A10系统生产,电导率 < 18 μS·cm−1。
样品采集和分析过程中避免使用任何聚四氟乙烯(PTFE)或氟聚合物材料;PFAAs的标准曲线的浓度梯度是0.01、0.05、0.1、0.5、1、5、10、50、100、500 ng·L−1,均加入5 ng PFAAs混合内标物;用3∶1的信噪比确定了检测限(Limit of detection, LOD),定量限(Limitation of quantitation, LOQ)确定为产生10∶1信噪比的分析峰值。17种PFAAs 的LOD和LOQ结果以及回收率列于表1,PFAAs标线的R2 > 0.99,回收率范围是70%—130%,符合质控要求。
1.4 数据来源与统计方法
(1)PFAAs的入海通量估算
采用简单水力模型估算PFAAs的入海通量,计算公式如下:
stringUtils.convertMath(!{formula.content}) (1) stringUtils.convertMath(!{formula.content}) (2) 其中,fi是评价河流的入海通量,由于苕溪、灵江、萩芦溪和诏安东溪缺年径流量数据,未估算;C是各条河流采样点表层水体PFBA、PFPeA、PFHxA、PFHpA、PFOA、PFNA、PFBS、PFHxS、PFOS和∑PFAAs的检出浓度,单位为 ng·L−1;n是每条河流的样点数量;Qi是河流的年径流量,单位为m3·a−1;f是单位换算系数,f = 100;F是估算排放总量(单位为kg·a−1),分别对9条河流PFBA、PFPeA、PFHxA、PFHpA、PFOA、PFNA、PFBS、PFHxS、PFOS和∑PFAAs 的入海通量求和。
(2)PFAAs源解析
采用因子分析、相关性分析、主成分分析和典型PFAAs的浓度比值探究PFAAs的污染来源。因子分析解释各PFAAs的可能来源[21];相关性分析和主成分分析可以解释各河流PFAAs污染来源的相似性[22--23];PFOA/PFOS、PFHpA/PFOA、PFBA/PFOA和PFNA/PFOA的三维空间散点图中度浓度比值大小和各方向轴上散点的离散程一定程度上分别反映了工业污水排放系统、雨水冲刷与大气沉降、人畜代谢产物、全氟调聚醇(Fluorotelomeralcohols,FTOHs)及前体物降解对河流水体中PFAAs的污染来源的贡献[24]。
(3)PFAAs的生态风险
采用风险熵法分别评价了PFBA、PFPeA、PFHxA、PFOA、PFNA、PFDA、PFBS和PFOS对水生生物的生态风险,计算公式如下:
stringUtils.convertMath(!{formula.content}) (3) 式中,RI(Risk index)是生态风险指数,Ci是每条河流的表层水体中PFBA、PFPeA、PFHxA、PFOA、PFNA、PFDA、PFBS和PFOS的浓度(均值,ng·L−1),EQSi(Environmental quality standard for ecology safety)是基于生物实验数推导出的风险评估阈值,PFBA、PFPeA、PFHxA、PFOA、PFNA、PFDA、PFBS和PFOS的评价阈值分别是1400、600、200、20、100000、11000、600、50 ng·L−1[4]。
(4)PFAAs的健康风险
采用GEA(German environment agency)开发的方法评价河流水体中8种PFAAs通过饮水摄入的综合潜在健康风险,计算公式如下:
stringUtils.convertMath(!{formula.content}) (4) 其中,CI(Comprehensive health risk index)是评价每条河流8种PFAAs的综合健康风险指数;Ci是每条河流中PFOS、PFNA、PFBS、PFBA、PFOA、PFHxS、PFPeA和PFHxA的浓度(均值,ng·L−1);EQShi(Environmental quality standard for human health to drink-waters)是PFAAs的健康评估阈值。目前,德国饮用水规定的健康评估阈值为:PFOS 100 ng·L−1;PFNA 60 ng·L−1;PFBS 3000 ng·L−1和意大利饮用水规定的健康评估阈值为:PFBA 7000 ng·L−1;PFOA 100 ng·L−1;PFHxS 100 ng·L−1;PFPeA 3000 ng·L−1;PFHxA 6000 ng·L−1[17-18]。
2. 结果与讨论(Results and discussion)
2.1 表层水中PFAAs的赋存特征
东南主要河流表层水中17种PFAAs的浓度赋存水平,如表2所示,∑17PFAAs浓度范围是0.90—231.52 ng·L−1,其平均浓度为46.82 ng·L−1,各PFAAs检出率范围是1.8%—92.3%。数据分析表明∑13PFCAs的浓度范围是0.87—195.15 ng·L−1,平均浓度为35.40 ng·L−1;∑4 PFSAs的浓度范围是nd—108.53 ng·L−1,平均浓度为11.41 ng·L−1;其中两种备受关注的PFAAs——PFOA(C8)和PFOS(C8)的浓度范围分别为nd—147.89 ng·L−1,nd—62.52 ng·L−1。浙江省和福建省∑PFAAs的高值点分别出现在钱塘江流域、闽江流域。由图2可见,PFAAs的组成赋存特征表现为由北向南短链(C4—7)PFAAs比重变大,长链PFAAs(C8—18)占的比重逐渐变小的趋势,短链比重高值点为九龙江,长链比重高值点为钱塘江。研究发现PFAAs生产、消费相对集中的钱塘江流域(杭州、湖州和嘉兴)河段表层水体中∑PFAAs最高检出浓度均低于我国北方小清河流域报道的水体浓度 ∑PFAAs 325280 ng·L−1[10]和大凌河流域报道的水体浓度∑PFAAs 9540 ng·L−1[25],但高于美洲、欧洲和北极水环境中的赋存浓度[26-28]。
表 2 东南主要河流表层水中全氟烷基酸的赋存特征Table 2. Occurrence of PFAAs in surface waters of Southeast China河流Rivers 样点数量Points PFAAs/(ng·L−1) 参考文献References PFBA PFOA PFOS ∑PFAAs 苕溪 2 7.60(6.81—8.39) 23.01(18.91—27.11) 7.73(0.77—14.70) 83.57(41.43—125.71) 钱塘江 6 16.16(10.38—21.40) 124.77(59.92—147.89) 2.71(1.83—3.38) 184.91(107.19—231.52) 甬江 4 7.27(5.50—10.22) 46.962(34.44—6.63) 21.50(5.00—62.52) 105.73(63.77—188.81) 港口 5 1.86(0.82—3.44) 10.16(8.82—12.65) 0.62(nd—2.00) 18.88(14.17—21.97) 灵江 3 5.98(1.12—15.46) 12.79(6.63—24.40) 19.92(0.43—55.99) 51.46(12.67—124.79) 瓯江 3 10.28(8.17—13.59) 6.04 (1.95—8.63) 1.58(0.82—2.94) 21.72(14.07—28.51) 本文 飞云江 3 1.78(0.80—3.46) 1.58(nd—3.58) 1.89(nd—5.40) 8.96(3.55—19.55) 鳌江 5 1.26(0.19—2.78) 0.33(nd—0.89) 0.40(nd—1.10) 6.33(1.53—13.31) 闽江 3 23.55(13.17—30.60) 23.47(17.36—27.95) 1.07(nd—1.72) 71.36(43.65—90.31) 萩芦溪 3 nd nd nd 1.00(0.90—1.08) 木兰溪 3 0.64(nd—1.62) 0.69(nd—2.06) 0.06(nd—0.18) 3.56(1.02—7.85) 晋江 4 1.80(1.2—2.54) 2.65(1.88—4.46) 1.00(nd—1.49) 17.60(7.92—30.25) 九龙江 5 1.96(0.94—3.12) 1.15(0.52—2.25) 0.31(nd—1.53) 11.29(6.43—19.48) 诏安东溪 3 0.81(0.36—1.08) 0.44(0.24—0.58) nd 2.64(1.59—3.46) 钱塘江杭州段 0.59—538 nd—2.48 0.98—609 [31] 污水厂出水 nd—4.00 9.18—11.68 12.28—28.76 54.04—105.64 [32] 大凌河 nd—2430 nd—2280 0.16—483 1.77—9540 [25] 小清河 276240 4.66 325280 [10] 美国 安大略湖 nd—13.00 nd—4.95 nd—84.60 <97.40 [27] 波罗的海地区 nd—8.52 nd—88.30 [33] 大西洋 0.08—5.80 [26] 北极 0.04—0.25 [26] 注:nd represents no detection, 表示样品中未检出PFAA,下同. | Show TableDownLoad:
CSV
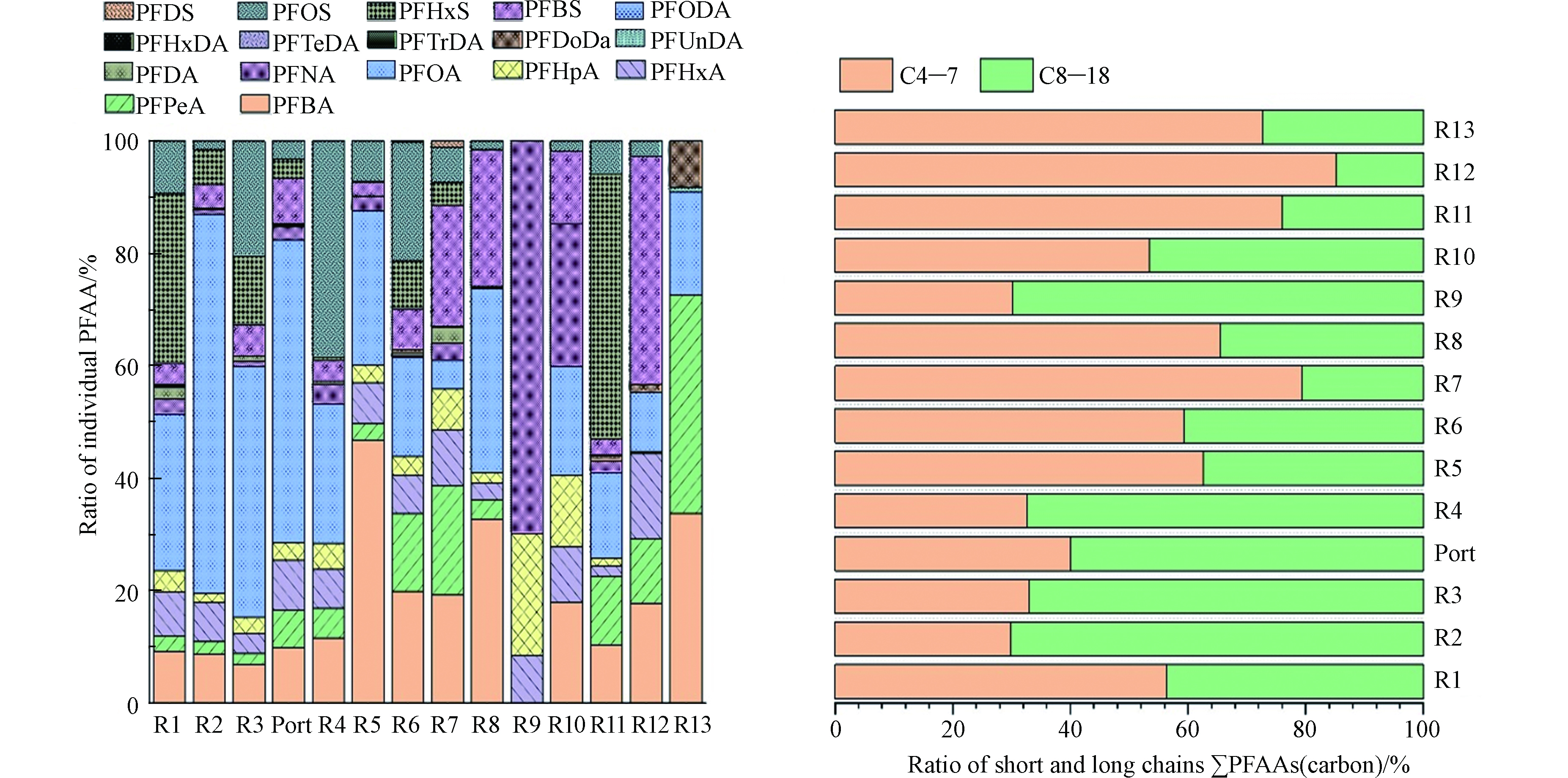 图 2 中国东南主要河流表层水体中17种PFAAs的组成特征Figure 2. Occurrence composition of 17 PFAAs in the surface waters of major rivers in the Southeast of China((a)PFAAs百分比水平、(b)基于碳链长度分类的百分比水平,C4—C7 短链,C8—C18长链)((a)Concentration ratio of individual PFAA(b)Concentration ratio of PFAAs base on carbon number,short chain: C4—C7, long chain: C8—C18)
图 2 中国东南主要河流表层水体中17种PFAAs的组成特征Figure 2. Occurrence composition of 17 PFAAs in the surface waters of major rivers in the Southeast of China((a)PFAAs百分比水平、(b)基于碳链长度分类的百分比水平,C4—C7 短链,C8—C18长链)((a)Concentration ratio of individual PFAA(b)Concentration ratio of PFAAs base on carbon number,short chain: C4—C7, long chain: C8—C18)造成表层水中PFAAs赋存差异的可能原因一是PFAAs排放受城市工业化程度影响,因为大部分河流自西向东流入海,河口区孕育的沿海港口城市的工业化程度均高于中国内陆城市,如东南主要河流的钱塘江、甬江和闽江以及北方河流海河、小清河、大凌河等流域均有工业布局,氟化工及相关行业的生产工艺和工艺废水的处理效率影响直接排放到水环境中的PFAAs的量[29-30];二是自然因素,雨水的冲刷和稀释作用影响PFAAs的迁移。东南主要河流中小河流众多,受季风气候、梅雨和台风影响,年均降雨量约1550 mm,瞬时降雨量大,泄洪快,可能将大气和地表赋存的PFAAs转移到河流、土壤有机质中[19],而北方河流汛期短,存在断流现象,河流径流量有较大差异从而影响PFAAs的赋存浓度。京杭运河(The Grand Canal)将环渤海区域与东南主要河流联通,然而张明等[31]发现∑PFAAs的高值点出现在江南运河的杭嘉湖平原,缺乏可靠证据说明跨流域的面源污染对东南主要河流的贡献大小。因此复杂的工业排放源和水文环境可能是造成表层水中PFAAs赋存在空间上存在差异的重要原因。
2.2 主要河流∑PFFAs的入海通量
东南9条主要河流PFAAs的入海通量估值结果如表3所示,该流域∑PFAAs的排放总量是7.12 t·a−1,其中PFOA排放入海量是3.71 t·a−1、PFBA 1.32 t·a−1、PFOS 0.2 t·a−1,钱塘江的排放入海量是4.04 t·a−1,闽江的排放入海量是2.14 t·a−1。河流年径流量范围是18.57×109—300.53×109 m3·a−1,径流量由高到低排序为闽江>钱塘江>瓯江>九龙江>飞云江>晋江>鳌江>甬江>木兰溪。钱塘江排放贡献最大是PFOA(38.3%,与区域∑PFAAs排放总量的比值,下同),高于闽江(10.0%),闽江河段排放贡献最大的是短链PFAAs(>19.5%)。我国东南9条主要河流∑PFAAs的入海通量远小于Zhou等[34]报道的黄渤海区域河流的入海通量,其中钱塘江流域PFOA的入海通量小于北方小清河(3.6 t·a−1),∑PFAAs排放值接近[21]。各河流表层水中PFAAs的入海通量取决于各PFAA及∑PFAAs浓度值、河流的年径流量大小,各PFAA在水相中的赋存浓度受污染源、水文特征、各PFAA的性质和水/沉积物分配系数(Koc)的影响[18]。污染源尤其是工业源的排放强度是影响各PFAA在水相赋存浓度的主要因素。PFBA、PFBS和PFHxs在水相中赋存浓度比较高,一方面可能是它们被用作长链PFAAs的理想替代物导致,另一方面是因为与长链PFAAs相比,短链PFAAs更容易留在水相[18]。通过河流持续的陆源输入,将对沿海生态系统产生不利影响,甚至使海产品爱好者潜在较高的人体健康风险[35]。河流和海洋是陆源污染的最终主要纳污受体[36],估算河流表层水中PFAAs的入海通量,有助于人们了解PFAAs经河流输入对海洋的贡献,为研究PFAAs水环境容量和今后制定、执行削减计划提供数据支持。研究厘清9条河流PFAAs入海量并与典型流域进行比较,有助于附近居民了解PFAAs的环境排放行为,并为深入研究PFAAs的环境行为提供参考依据。
表 3 中国东南主要河流中PFAAs的入海通量及与其他研究比较Table 3. Mass flux from rivers to sea of typical PFAAs along southeast China and comparison with other studies河流Rivers 采样时间Sampling time PFBA/(kg·a−1) PFPeA/(kg·a−1) PFHxA/(kg·a−1) PFHpA/(kg·a−1) PFOA/(kg·a−1) PFNA/(kg·a−1) PFBS/(kg·a−1) PFHxS/(kg·a−1) PFOS/(kg·a−1) ∑PFAAs/(kg·a−1) 径流量Runoff(×109)/(m3·a−1) 参考文献References 钱塘江 353.2 82.6 279.2 68.0 2726.2 37.7 167.3 255.3 59.1 4040.0 218.5 甬江 20.8 5.6 10.9 8.6 134.3 3.2 16.8 37.2 61.5 302.0 28.6 瓯江 199.4 6.6 31.3 13.3 117.3 10.5 10.2 0.8 30.7 421.3 194.0 飞云江 7.9 4.5 2.7 1.3 7.0 0.1 2.9 3.4 8.4 39.5 44.5 鳌江 2018年8月 6.9 5.0 3.8 2.3 2.0 1.3 7.2 1.1 2.5 33.3 36.9 本研究 闽江 707.7 51.2 64.2 40.8 705.2 4.6 524.8 3.6 32.3 2137.5 300.5 晋江 7.0 7.5 1.2 1.0 10.3 1.3 1.9 32.0 3.9 66.1 39.0 九龙江 12.8 6.8 10.9 0.3 7.5 0.0 29.1 0.0 2.0 69.3 65.2 木兰溪 1.2 0.0 0.7 0.8 1.3 1.7 0.9 0.0 0.1 6.6 18.57 东南主要河流 1316.9 169.6 404.9 136.5 3711.2 60.5 761.0 333.3 200.5 7115.7 945.9 钱塘江 2011年5月 849.0 1445.0 148.3 [12] 甬江 2011年5月 137.0 185.0 35.0 [12] 瓯江 2011年5月 13 24.0 195.5 [12] 飞云江 2011年5月 6.8 13.0 40.0 [12] 大凌河 2008年5月 145.0 19.6 [39] 大辽河 2008年5月 75.5 46.6 [39] 环渤海北部排放总量 2008年5月 216.0 122.0 [39] 小清河 2011年9月 3600.0 4000.0 19.0 [21] 渤海排放总量 2011年9月 246.0 127.5 207.6 154.7 3975.9 48.7 38.2 13.1 136.0 4962.0 [21] 海河 2016年6月 30.07 19.9 32.5 6.8 25.9 3.5 6.0 1.6 14.3 142.2 [34] 渤海、黄海排放总量 2016年6月 3329.3 1828.2 2868.4 2568.3 52952.3 1008.8 3878.2 272.9 2609.0 72170.3 [34] 德国萨勒河 2015年 164.0 [33] | Show TableDownLoad:
CSV
造成各流域入海通量存在差异的原因是多方面的,主要原因是工业排放。钱塘江上游存在PFOA排放点源[31],闽江上游可能存在PFBA、PFBS、PFOS排放点源[37],甬江附近有大型氟化工生产基地和众多乡镇企业,是潜在的工业排放源。因此这些河流的表层水中∑PFAAs浓度较高,但其入海通量不一定高。环渤海区小清河、大凌河流域附近也存在大型氟化工园区,可能是持续排放强度大于东南主要河流,使得水体中PFAAs赋存浓度较高[34]。然而与工业化较高的德国萨勒河流域、意大利亚高山湖泊流域和法国南部流域相比,东南主要河流表层水中∑PFAAs赋存浓度更高[28, 33, 38]。
2.3 主要河流中PFAAs的源解析
东南主要河流中PFAAs污染主要来源于工业源。工业排放的PFAAs有的是直接进入河流,有的通过大气干湿沉降或城市管网间接进入河流[11]。采用因子分析法即最大方差旋转提取主成分因子(Kaiser-Meyer-Olkin 值为0.615),如图3和表4所示,筛选出7个特征值 > 1的因子,可以解释水体中80.2%PFAAs的来源,符合主成分分析的要求。其中,因子1可以解释东南主要河流表层水体中29.0%的PFAAs来源,载荷较高的是PFODA、PFTrDA、PFUnDA和PFDoDA;因子2可以解释该流域表层水体中17.2%的PFAAs来源,载荷较高的是PFBA、PFBS、PFHxA和PFOA;因子3可以解释该流域表层水体中11.4%的PFAAs的来源,载荷较高的是PFNA和PFHpA。如图4(a),进一步应用主成分分析特征值较大的两个因子,在同一象限内的分值点越接近PFAAs污染来源越相似,即PC1可以解释钱塘江、闽江和甬江河段表层水中的PFAAs有相似的来源,PC2可以解释多数河流表层水中PFUnDA和PFDoDA来源于调聚法生产过程排放的FTOHs。PC1和PC2的累加贡献可以解释46.2%的变量,低于李法松等[40]分析的结果,说明两个研究区域PFAAs的环境排放和分配行为存在差异。
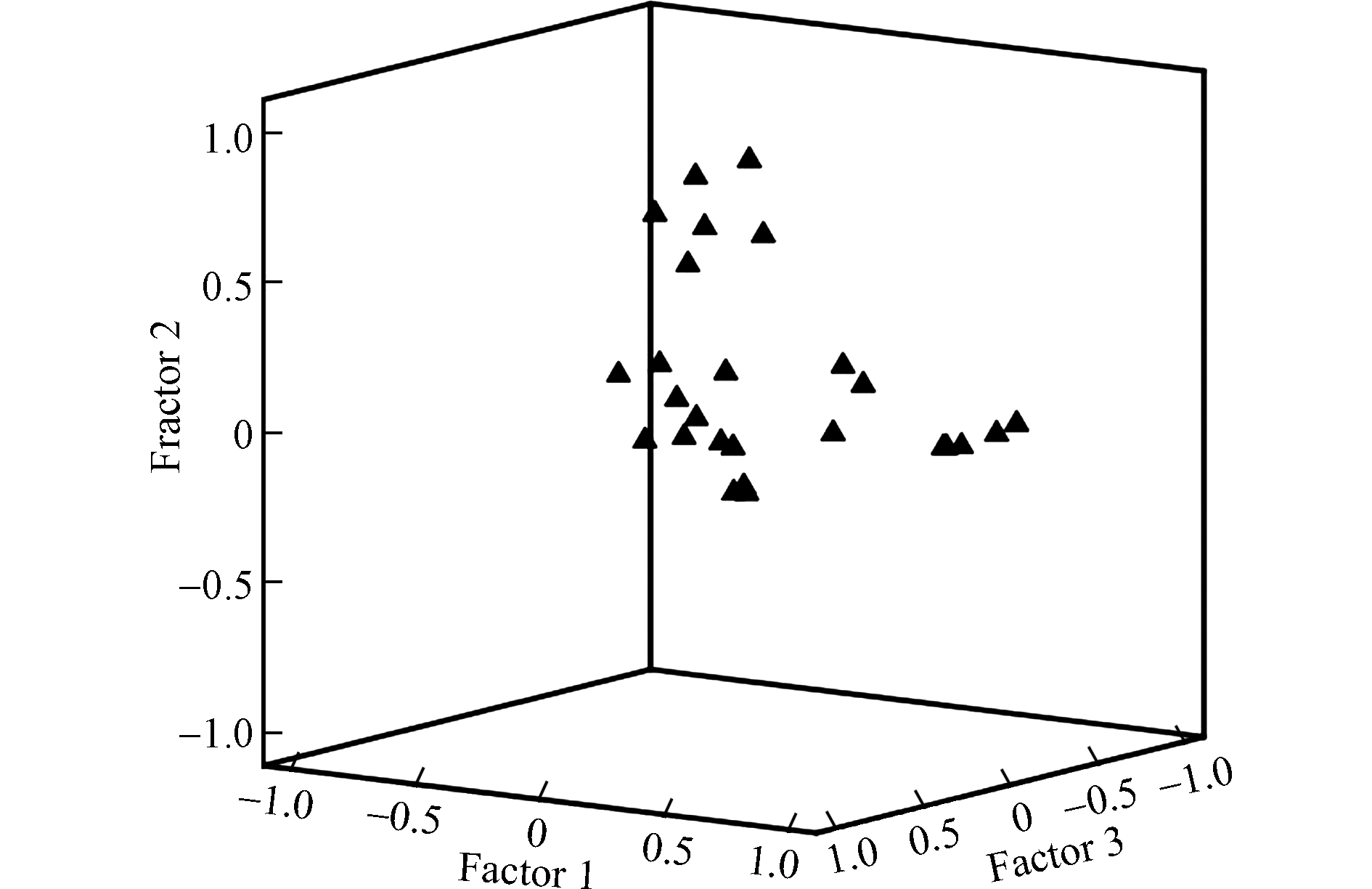 图 3 最大方差法旋转提取主成分因子(Kaiser-Meyer-Olkin 值为0.615)Figure 3. Maximum variance method rotation extraction for principle component factor(KMO value 0.615)表 4 17种全氟烷基酸的主成分因子分析Table 4. Principle component factor analysis of 17 PFAAs
图 3 最大方差法旋转提取主成分因子(Kaiser-Meyer-Olkin 值为0.615)Figure 3. Maximum variance method rotation extraction for principle component factor(KMO value 0.615)表 4 17种全氟烷基酸的主成分因子分析Table 4. Principle component factor analysis of 17 PFAAs变量Variance 因子载荷矩阵(Factor load matrix) 1 2 3 4 5 6 7 PFBA 0.022 0.845 0.249 −0.107 −0.145 −0.023 0.046 PFPeA 0.22 0.654 0.143 −0.125 0.421 0.065 0.052 PFHxA 0.166 0.712 0.402 0.105 0.415 0.036 −0.019 PFHpA 0.219 0.618 0.573 −0.305 0.235 0.139 0.032 PFOA −0.077 0.724 0.341 0.143 0.438 0.135 −0.038 PFNA 0.193 0.301 0.698 −0.212 0.389 0.003 0.207 PFDA 0.778 0.243 0.367 −0.088 0.096 0.122 −0.007 PFUnDA 0.899 0.023 −0.34 −0.045 0.113 −0.023 −0.028 PFDoDA 0.613 −0.082 −0.633 −0.046 0.196 −0.13 0.056 PFTrDA 0.962 0.013 0.066 0.06 0.049 −0.019 0.03 PFTeDA 0.961 0.023 0.151 0.111 0.057 0.001 0.032 PFHxDA −0.034 0.021 0.164 −0.064 0.112 −0.063 −0.906 PFODA 0.961 0.024 0.168 0.103 0.035 0.004 0.028 PFBS 0.023 0.855 −0.058 −0.091 −0.207 −0.05 −0.14 PFHxS 0.588 0.265 0.211 0.033 0.305 0.514 −0.06 PFOS 0.141 0.203 0.246 −0.516 0.075 0.624 0.135 PFDS −0.03 −0.079 0.028 −0.03 −0.083 0.896 0.004 纬度 0.084 0.266 0.779 0.05 0.173 0.131 −0.154 pH 0.209 0.063 0.806 0.196 0.117 −0.214 −0.062 特征值 7.54 4.48 2.98 1.80 1.58 1.40 1.08 贡献率/% 29.0 17.2 11.4 6.9 6.1 5.4 4.2 累积贡献率/% 29.0 46.2 57.7 64.6 70.7 76.1 80.2 | Show TableDownLoad:
CSV
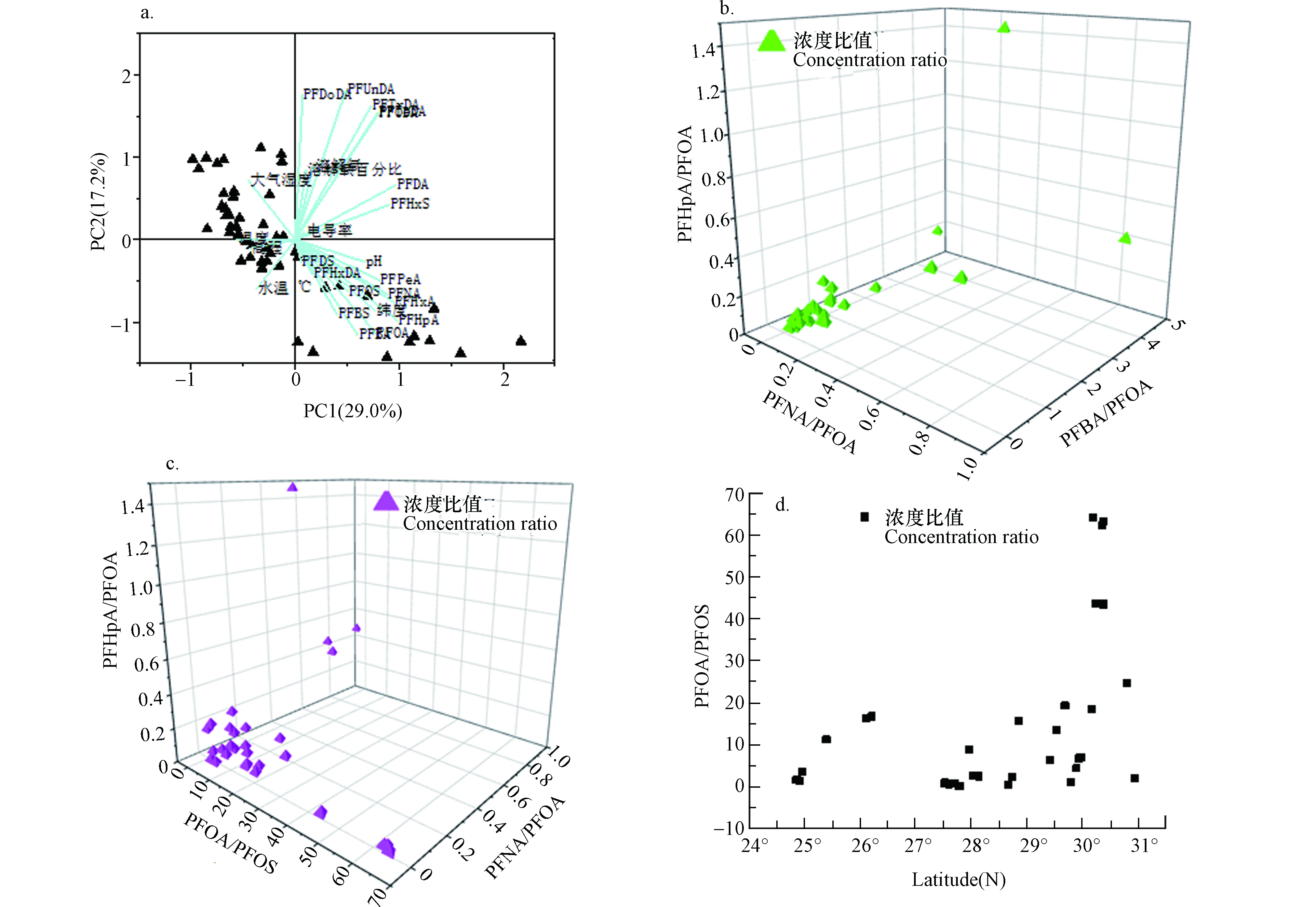 图 4 东南主要河流表层水体中PFAAs的源解析(注: (a)主成分分析;(b)和(c)特征PFAAs的浓度比值的3D散点图;(d)PFOA/PFOS的浓度比与纬度的关系Figure 4. Source of PFAAs in surface waters of Southeast China(Note:(a)Principle component analysis;(b)and(c)3D scatter plot of characteristic PFAAs concentration ratio (d)Relationship between PFOA/PFOS concentration ratio and latitude.
图 4 东南主要河流表层水体中PFAAs的源解析(注: (a)主成分分析;(b)和(c)特征PFAAs的浓度比值的3D散点图;(d)PFOA/PFOS的浓度比与纬度的关系Figure 4. Source of PFAAs in surface waters of Southeast China(Note:(a)Principle component analysis;(b)and(c)3D scatter plot of characteristic PFAAs concentration ratio (d)Relationship between PFOA/PFOS concentration ratio and latitude.Spearman相关性分析结果表明表层水中PFOA分别与PFBA(P < 0.01)、PFPeA(P < 0.01)、PFHxA(P < 0.01)和PFHpA(P < 0.01)呈显著正相关,可能是它们的主要污染来源相同或是有类似的环境行为;PFUnDA和PFDoDA均与PFOA(P < 0.01)和PFNA(P < 0.01)呈显著负相关,与长链PFAAs的降解行为有关[15],调聚法生产的全氟化合物在紫外光和高温条件下可能分解为C8结构;PFOS与PFCAs(C4—C10, P < 0.01)呈显著正相关,可能是PFOS生产过程中产生的副产物。
比值法结果说明PFOA和PFBA是东南主要河流表层水中主要的PFAAs污染物,可能来源于工农业排放。从图4分析得出工业污水排放对钱塘江的贡献最为显著,大气干湿沉降对鳌江、瓯江河段的贡献较大。由于PFBA水溶性良好,不易降解,迁移能力强,进入人体能很快被代谢排出,水环境中赋存的PFBA可能来源于长链PFAAs的降解,常被解读为人畜代谢产物或生活污水来源[24]。如图4(b)所示,大部分散点在PFBA/PFOA轴方向上,比值范围是0.1—4.2,均值为0.8,表明人畜排泄物中携带PFAAs对东南主要河流水体污染有较大的贡献,可能流域内有规模较大的畜牧农场、家禽养殖场分布[37]。如图4(c)所示,大部分点偏向于PFOA/PFOS轴方向上,比值范围是0.4—64.3,均值为15.0,说明工业污水排放对PFAAs进入水环境的贡献非常大。
表层水中PFAAs的可能污染来源是人类活动源,如工业源,交通源、生活源、农业源和消防等,其中氟化工园区排放是重要的工业源[11]。图4(d)表明高纬度地区PFOA/PFOS的值较大,是因为钱塘江上游兰江、衢江以及杭嘉湖平原均有氟化工生产基地,沿岸布局的制革、造纸、纺织印染、电镀等行业均使用PFOA和PFOS及其替代品作除渍和防水处理剂。分析发现PFOA的浓度高于其他PFAAs,也高于研究区的其他流域,说明该流域内有较强的排放点源,与张明等[6, 31]研究结果一致。
2.4 生态风险与健康风险评价
应用风险熵法分别评价东南主要河流表层水中赋存浓度较高的8种PFAAs对水生生物的生态风险,本研究采用了较为严格的评价阈值[4],如表5所示,结果表明PFBA、PFPeA、PFHxA、PFNA、PFDA和PFBS的风险熵 < 1,说明目前表层水中PFAAs的赋存浓度对该流域水生生物不构成生态风险。但钱塘江、闽江等部分河段PFOA和PFOS的风险熵 > 1,说明部分河段潜在生态风险,可能对水生生物产生不利影响。
表 5 东南主要河流表层水中 8 种PFAAs的生态风险评价Table 5. Estimated risk quotients of 8 PFAAs in surface watersof Southeast ChinaPFAAs PFBA PFPeA PFHxA PFOA PFNA PFDA PFBS PFOS 评价阈值 1400 ng·L−1 600 ng·L−1 200 ng·L−1 20 ng·L−1 100000 ng·L−1 11000 ng·L−1 600 ng·L−1 50 ng·L−1 苕溪 0.005 0.003 0.032 1.151 <<0.001 <<0.001 0.005 0.155 钱塘江 0.012 0.006 0.064 6.239 <<0.001 <<0.001 0.013 0.054 甬江 0.005 0.003 0.019 2.348 <<0.001 <<0.001 0.010 0.430 港口 0.001 0.002 0.008 0.508 <<0.001 <<0.001 0.003 0.012 灵江 0.004 0.004 0.018 0.640 <<0.001 <<0.001 0.003 0.398 瓯江 0.007 0.001 0.008 0.302 <<0.001 <<0.001 0.001 0.032 飞云江 0.001 0.002 0.003 0.079 <<0.001 <<0.001 0.001 0.038 鳌江 0.001 0.002 0.003 0.016 <<0.001 <<0.001 0.002 0.008 闽江 0.017 0.003 0.011 1.173 <<0.001 <<0.001 0.029 0.021 萩芦溪 <<0.001 <<0.001 <<0.001 <<0.001 <<0.001 <<0.001 <<0.001 <<0.001 木兰溪 <<0.001 <<0.001 <<0.001 <<0.001 <<0.001 <<0.001 0.001 0.001 晋江 0.001 0.003 0.002 0.132 <<0.001 <<0.001 0.001 0.020 九龙江 0.001 0.002 0.008 0.058 <<0.001 <<0.001 0.007 0.006 诏安东溪 0.001 0.001 <<0.001 0.022 <<0.001 <<0.001 <<0.001 <<0.001 流域均值 0.004 0.002 0.015 1.141 <<0.001 <<0.001 0.006 0.078 流域范围 0—0.0215 0—0.0107 0—0.1040 0—7.3943 <<0.001 <<0.001 0—0.039 0—1.251 统计 >1 0 0 0 14 0 0 0 2 | Show TableDownLoad:
CSV
采用商值法评价了8种赋存浓度较高的PFAAs的居民健康风险,如表6所示,各河流表层水中∑8PFAAs的风险熵范围是0.05—1.428,表明大部分评价河流不构成 ∑8PFAAs的人体健康风险;诏安东溪的CI小于其他河流,表明潜在排放源(P10)对其影响不大,PFAAs污染较轻(∑17PFAAs浓度范围为 1.59—3.46 ng·L−1),可能存在面源污染;钱塘江流域的部分河段水体的CI > 1,表明表层水中∑8PFAAs潜在人体健康风险,河水需经过处理达标才能用作饮用水。甬江的CI值为0.837,苕溪的CI值为0.599,为确保拥有充足的饮用水水源和饮用水安全,CI值较高的河流值得给予关注。目前缺乏相关毒理数据,大多数评价采用EDI(Estimated daily intake)和商值法组合评价PFOS和PFOA的风险大小。Chimeddulam等[8, 41]根据中国人群暴露手册分别评价了PFOA和PFOS对我国台湾和上海地区不同年龄人群的潜在饮水健康风险,然而对其余PFAAs关注较少,该方法目前不能全面评价多种PFAAs共同作用下的潜在风险,留有研究空白。
表 6 东南主要河流表层水中8种PFAAs的健康风险评价Table 6. Assessing the health risks of 8 PFAAs in surface waters of major rivers in southeast China河流Rivers 综合健康风险指数Comprehensive health risk index(CI) 标准偏差Standard deviation(SD) 变异系数Coefficient of variation(CV) 均值mean 最小值minimum 最大值maximum 苕溪 0.599 0.284 0.914 0.445 0.743 钱塘江 1.428 0.719 1.768 0.381 0.267 甬江 0.837 0.442 1.665 0.578 0.690 港口 0.123 0.108 0.165 0.023 0.191 灵江 0.364 0.093 0.128 0.440 1.207 瓯江 0.088 0.031 0.121 0.050 0.566 飞云江 0.044 0.006 0.113 0.060 1.380 鳌江 0.015 0.003 0.032 0.014 0.932 闽江 0.259 0.179 0.316 0.071 0.274 萩芦溪 0.012 0.011 0.012 0.000 0.026 木兰溪 0.023 0.013 0.043 0.017 0.750 晋江 0.125 0.024 0.263 0.107 0.855 九龙江 0.017 0.006 0.025 0.011 0.673 诏安东溪 0.005 0.003 0.006 0.002 0.415 流域 0.322 0.005 1.768 0.514 1.595 | Show TableDownLoad:
CSV
大量研究证实PFAAs存在于各个区域的水环境中,在藻类、底栖微生物、无脊椎动物和鱼类中均检出了较高的赋存浓度(PFOS, 1900 ng·g−1湿重,均值,下同);PFAAs具有生物毒性,表现为蓄积与隔代传递、营养级放大、干扰代谢、影响发育与生殖等[42]。Zhang等[43]发现成年斑马鱼暴露于PFNA 10、 100、 1000 μg·L−1浓度下72 h有明显的剂量效应,雄性性腺体指数和雌性产卵量显著降低,可能影响其繁殖发育。Mazzoni等[28]在意大利典型工业化区域的5个亚高山湖泊水体中发现各PFAAs的检出浓度接近检出限,但从当地居民消费的湖泊鱼类中检出的PFOS浓度是欧盟规定值(9.6 ng·g−1)的0.1—1.7倍,表明鱼类组织对PFOS有较强的富集能力。Simmonet等[38]发现在法国南部的淡水食物网中食用鱼类体内PFAAs的赋存比无脊椎甲壳类动物高,主要污染物为PFOS和PFDS。Chen等[19]报道了中国华东地区自来水中∑PFAAs的范围在9.29—266.68 ng·L−1,主要污染物为PFOA和PFBA。水体中PFAAs的浓度较高时潜在生态风险和饮水、饮食摄入人体健康风险,因此加强对东南主要河流水源地的保护以及对水质持续关注有重要意义。
3. 结论(Conclusions)
(1)东南主要河流表层水中∑PFAAs浓度范围是0.90—231.52 ng·L−1,平均浓度为46.82 ng·L−1,∑17PFAAs的入海通量约为7.12 t·a−1,表层水体中主要PFAAs污染物是PFOA和PFBA。
(2)PFAAs的排放源主要是工业污水排放,其中钱塘江流域污染较为严重。
(3)东南主要河流表层水中PFAAs的总体污染水平较低,但钱塘江、闽江、苕溪和甬江的部分河段对水生生物潜在生态风险。
(4)钱塘江流域部分河段潜在人体健康风险,后续工作将持续关注PFAAs风险熵较高河段的人体健康风险。
-
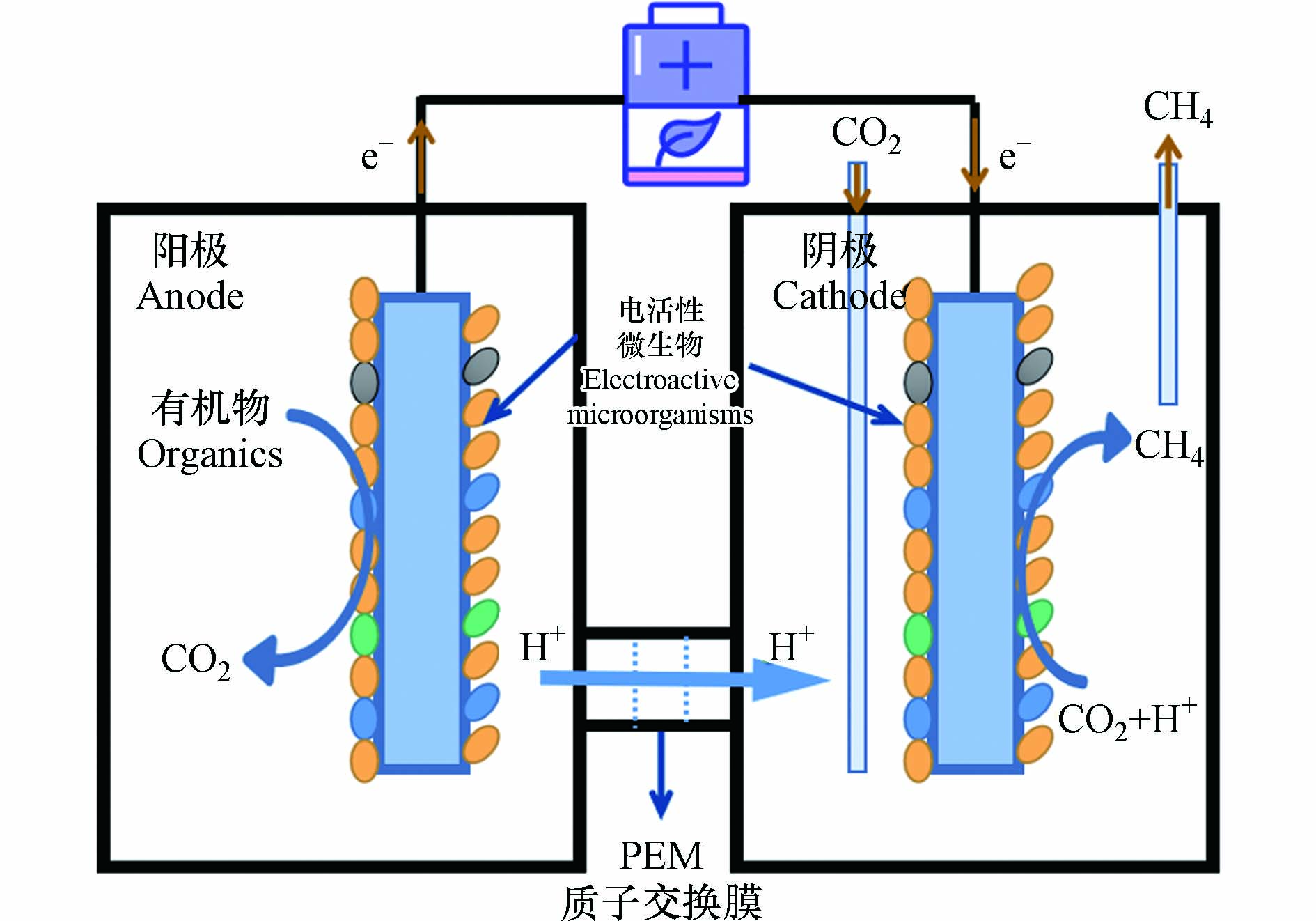
图 1 MEC CO2电产甲烷示意图
Figure 1. Schematic diagram of MEC CO2 electricity production of methane

图 2 (a)关于MEC的期刊论文数量,(b)关于MEC的文章在国家/地区的分布
Figure 2. (a) Number of journal articles on MEC, (b) distribution of articles on MEC by country

图 3 不同电极材料[2-4, 6-21](a)、不同电压[5, 7, 9, 25-36](b)和不同反应器构型[4, 6-9, 11-15, 26-34, 37](c)下最高甲烷产生速率和MEC最大电流密度
Figure 3. Maximum methane production rate and maximum MEC current density for different electrode materials [2-4, 6-21] (a), voltages [5, 7, 9, 25-36] (b) and reactor configurations [4, 6-9, 11-15, 26-34, 37] (c)
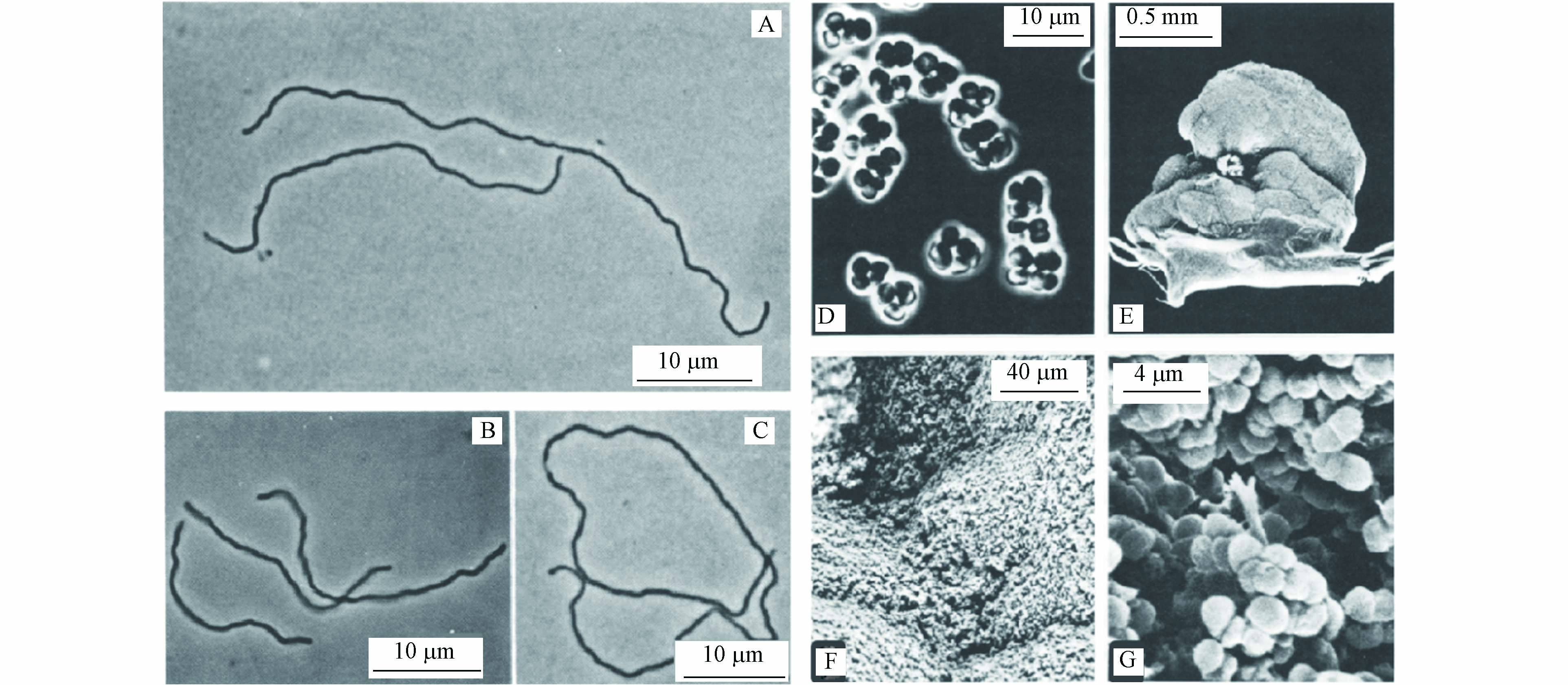
图 4 (A—C)65—70 ℃下生长的甲烷杆菌Methanobacterium thermoautotrophicus的SEM图,10 μm[40];(D—G)甲烷球菌Methanosarcina strain 227的SEM图[41],(D)10 μm,(E)500 μm,(F)40 μm,(G)4 μm
Figure 4. (A—C) SEM images of Methanobacterium thermoautotrophicus grown at 65—70 ℃, 10 μm[40]; (D—G) SEM images of Methanosarcina strain 227 [41], (D) 10 μm , (E) 500 μm, (F) 40 μm, and (G) 4 μm

图 5 细胞间的电子传递机制(a)通过可扩散分子(如H2和甲酸盐)[53],(b)通过电子穿梭(如黄素)[54],(c)通过导电菌毛[60],(d)通过细胞间的直接接触(如外膜c型细胞色素)[64],(e)通过导电材料(如活性炭、纳米磁铁矿)[61,66-67]
Figure 5. Electron transfer mechanism between cells (a) via diffusible molecules (such as H2 and formate) [53], (b) via electron shuttle (such as flavin) [54], (c) via conductive pili [60], (d) via intercellular direct contact (e.g. outer membrane c-type cytochromes) [64], (e) via conductive materials (e.g. activated carbon, nanomagnetite) [61,66-67]

图 6 近五年MEC耦合其他系统产甲烷示意图
Figure 6. Schematic diagram of methane production from other systems coupled with MEC in the last five years
表 1 产甲烷菌的主要生理特征
Table 1. Main physiological characteristics of the methanogenic bacteria
产甲烷菌Methanogenic bacteria 碳源Carbon sources 温度范围/ °C Temperature range pH 参考文献References Methanosarcinales 乙酸盐,H2 + CO2,CO, 甲醇, 甲胺,甲硫基丙酸甲酯,二甲硫 1.0—70 4.0—10.0 [43-45] Methanomicrobiales H2 + CO2,甲酸盐,乙醇,2-丙醇,2-丁醇,环戊醇 15—60 6.1—8.0 [42, 46] Methanobacteriales H2 + CO2,CO,甲酸盐,C1-甲基化合物 20—88 5.0—8.8 [44] Methanococcales H2 + CO2,甲酸盐 < 20—88 4.5—9.8 [44, 46] Methanopyrales H2 + CO2 84—110 5.5—7.0 [47-48] Methanocellales H2 + CO2,甲酸盐 25—40 6.5—7.8 [44]
下载: 导出CSV
-
[1] 谢典, 高亚静, 芦新波, 等. 能耗“双控”向碳排放“双控”转变的实施路径研究 [J]. 综合智慧能源, 2022, 44(7): 73-80. XIE D, GAO Y J, LU X B, et al. Research on the implementation path of the transition from dual control on energy consumption to dual control on carbon emission [J]. Integrated Intelligent Energy, 2022, 44(7): 73-80(in Chinese).
[2] 郑韶娟, 陆雪琴, 张衷译, 等. 微生物电解池: 生物电催化辅助CO2甲烷化技术 [J]. 环境化学, 2019, 38(7): 1666-1674. doi: 10.7524/j.issn.0254-6108.2018091502 ZHENG S J, LU X Q, ZHANG Z Y, et al. Microbial electrolysis cell (MEC): A new platform for CO2 bioelectromethanogenesis assisted by bioelectrocatalysis [J]. Environmental Chemistry, 2019, 38(7): 1666-1674(in Chinese). doi: 10.7524/j.issn.0254-6108.2018091502
[3] ZHEN G Y, ZHENG S J, HAN Y L, et al. Semi-continuous anolyte circulation to strengthen CO2 bioelectromethanosynthesis with complex organic matters as the e-/ H+ donor for simultaneous biowaste refinery [J]. Chemical Engineering Journal, 2022, 430: 133123. doi: 10.1016/j.cej.2021.133123 [4] JIANG Y, SU M, LI D P. Removal of sulfide and production of methane from carbon dioxide in microbial fuel cells-microbial electrolysis cell (MFCs-MEC) coupled system [J]. Applied Biochemistry and Biotechnology, 2014, 172(5): 2720-2731. doi: 10.1007/s12010-013-0718-9 [5] 邹亚娜, 臧越, 王恺元, 等. 生物电催化调控污泥-餐厨垃圾协同厌氧产酸研究 [J]. 环境化学, 2023, 42(1): 298-309. doi: 10.7524/j.issn.0254-6108.2021081202 ZOU Y N, ZANG Y, WANG K Y, et al. Regulated VFAs production from sewage sludge and food waste by insitu bioelectrocatalytic regulation [J]. Environmental Chemistry, 2023, 42(1): 298-309(in Chinese). doi: 10.7524/j.issn.0254-6108.2021081202
[6] WANG X T, ZHANG Y F, WANG B, et al. Enhancement of methane production from waste activated sludge using hybrid microbial electrolysis cells-anaerobic digestion (MEC-AD) process - A review [J]. Bioresource Technology, 2022, 346: 126641. doi: 10.1016/j.biortech.2021.126641 [7] ZHEN G Y, LU X Q, KOBAYASHI T, et al. Promoted electromethanosynthesis in a two-chamber microbial electrolysis cells (MECs) containing a hybrid biocathode covered with graphite felt (GF) [J]. Chemical Engineering Journal, 2016, 284: 1146-1155. doi: 10.1016/j.cej.2015.09.071 [8] DING A Q, YANG Y, SUN G D, et al. Impact of applied voltage on methane generation and microbial activities in an anaerobic microbial electrolysis cell (MEC) [J]. Chemical Engineering Journal, 2016, 283: 260-265. doi: 10.1016/j.cej.2015.07.054 [9] LIU D D, ZHENG T Y, BUISMAN C, et al. Heat-treated stainless steel felt as a new cathode material in a methane-producing bioelectrochemical system [J]. ACS Sustainable Chemistry & Engineering, 2017, 5(12): 11346-11353. [10] CHOI M J, YANG E, YU H W, et al. Transition metal/carbon nanoparticle composite catalysts as platinum substitutes for bioelectrochemical hydrogen production using microbial electrolysis cells [J]. International Journal of Hydrogen Energy, 2019, 44(4): 2258-2265. doi: 10.1016/j.ijhydene.2018.07.020 [11] JIN X D, ZHANG Y F, LI X H, et al. Microbial electrolytic capture, separation and regeneration of CO2 for biogas upgrading [J]. Environmental Science & Technology, 2017, 51(16): 9371-9378. [12] CAI W W, LIU W Z, ZHANG Z J, et al. mcrA sequencing reveals the role of basophilic methanogens in a cathodic methanogenic community [J]. Water Research, 2018, 136: 192-199. doi: 10.1016/j.watres.2018.02.062 [13] 程佳鑫, 李荣兴, 杨海涛, 等. 三维电催化氧化处理难生化降解有机废水研究进展 [J]. 环境化学, 2022, 41(1): 288-304. doi: 10.7524/j.issn.0254-6108.2020082804 CHENG J X, LI R X, YANG H T, et al. Review of three-dimensional electrodes for bio-refractory organic wastewater treatment [J]. Environmental Chemistry, 2022, 41(1): 288-304(in Chinese). doi: 10.7524/j.issn.0254-6108.2020082804
[14] KIM K R, KANG J, CHAE K J. Improvement in methanogenesis by incorporating transition metal nanoparticles and granular activated carbon composites in microbial electrolysis cells [J]. International Journal of Hydrogen Energy, 2017, 42(45): 27623-27629. doi: 10.1016/j.ijhydene.2017.06.142 [15] CHENG T F, LI H Y, XIA W, et al. Exploration into the nickel ‘microcosmos’ in prokaryotes [J]. Coordination Chemistry Reviews, 2016, 311: 24-37. doi: 10.1016/j.ccr.2015.12.007 [16] LI X, ZENG C P, LU Y B, et al. Development of methanogens within cathodic biofilm in the single-chamber microbial electrolysis cell [J]. Bioresource Technology, 2019, 274: 403-409. doi: 10.1016/j.biortech.2018.12.002 [17] ZHEN G Y, LU X Q, KOBAYASHI T, et al. Continuous micro-current stimulation to upgrade methanolic wastewater biodegradation and biomethane recovery in an upflow anaerobic sludge blanket (UASB) reactor [J]. Chemosphere, 2017, 180: 229-238. doi: 10.1016/j.chemosphere.2017.04.006 [18] ZHEN G Y, ZHENG S J, LU X Q, et al. A comprehensive comparison of five different carbon-based cathode materials in CO2 electromethanogenesis: Long-term performance, cell-electrode contact behaviors and extracellular electron transfer pathways [J]. Bioresource Technology, 2018, 266: 382-388. doi: 10.1016/j.biortech.2018.06.101 [19] KIM K Y, HABAS S E, SCHAIDLE J A, et al. Application of phase-pure nickel phosphide nanoparticles as cathode catalysts for hydrogen production in microbial electrolysis cells [J]. Bioresource Technology, 2019, 293: 122067. doi: 10.1016/j.biortech.2019.122067 [20] 唐韵. 阴极表面修饰和电压调控改善MEC阴极生物膜生长和产甲烷性能的研究[D]. 杭州: 浙江大学, 2017. TANG Y. The study of improving cathode biofilm growth and methane production in MEC by surface modification and voltage control[D]. Hangzhou: Zhejiang University, 2017(in Chinese).
[21] 薄涛, 翟洪艳, 季民. 不锈钢毡电极MEC甲烷原位纯化及原理 [J]. 环境科学学报, 2017, 37(11): 4057-4063. doi: 10.13671/j.hjkxxb.2017.0213 BO T, ZHAI H Y, JI M. Research on in suit methane purification in MEC with stainless steel felt as electrode and theory analysis [J]. Acta Scientiae Circumstantiae, 2017, 37(11): 4057-4063(in Chinese). doi: 10.13671/j.hjkxxb.2017.0213
[22] VU M, NOORI M, MIN B. Magnetite/zeolite nanocomposite-modified cathode for enhancing methane generation in microbial electrochemical systems [J]. Chemical Engineering Journal, 2020, 393: 124613. doi: 10.1016/j.cej.2020.124613 [23] ZHENG X M, LIN R J, XU J, et al. moEnhanced methane production by bimetallic metal-organic frameworks (MOFs) as cathode in an anaerobic digestion microbial electrolysis cell [J]. Chemical Engineering Journal, 2022, 440: 135799. doi: 10.1016/j.cej.2022.135799 [24] HE Y T, LI Q, LI J, et al. Magnetic assembling GO/Fe3O4/microbes as hybridized biofilms for enhanced methane production in microbial electrosynthesis [J]. Renewable Energy, 2022, 185: 862-870. doi: 10.1016/j.renene.2021.12.117 [25] BO T, ZHU X Y, ZHANG L X, et al. A new upgraded biogas production process: Coupling microbial electrolysis cell and anaerobic digestion in single-chamber, barrel-shape stainless steel reactor [J]. Electrochemistry Communications, 2014, 45: 67-70. doi: 10.1016/j.elecom.2014.05.026 [26] LIU W Z, CAI W W, GUO Z C, et al. Microbial electrolysis contribution to anaerobic digestion of waste activated sludge, leading to accelerated methane production [J]. Renewable Energy, 2016, 91: 334-339. doi: 10.1016/j.renene.2016.01.082 [27] WANG L, YANG C X, SANGEETHA T, et al. Methane production in a bioelectrochemistry integrated anaerobic reactor with layered nickel foam electrodes [J]. Bioresource Technology, 2020, 313: 123657. doi: 10.1016/j.biortech.2020.123657 [28] ZHANG Y, GONG L L, JIANG Q Q, et al. In-situ CO2 sequestration and nutrients removal in an anaerobic digestion-microbial electrolysis cell by silicates application: Effect of dosage and biogas circulation [J]. Chemical Engineering Journal, 2020, 399: 125680. doi: 10.1016/j.cej.2020.125680 [29] LEE M, REDDY C N, MIN B. In situ integration of microbial electrochemical systems into anaerobic digestion to improve methane fermentation at different substrate concentrations [J]. International Journal of Hydrogen Energy, 2019, 44(4): 2380-2389. doi: 10.1016/j.ijhydene.2018.08.051 [30] LIU D D, ROCA-PUIGROS M, GEPPERT F, et al. Granular carbon-based electrodes as cathodes in methane-producing bioelectrochemical systems [J]. Frontiers in Bioengineering and Biotechnology, 2018, 6: 78. doi: 10.3389/fbioe.2018.00078 [31] BRETSCHGER O, CARPENTER K, PHAN T, et al. Functional and taxonomic dynamics of an electricity-consuming methane-producing microbial community [J]. Bioresource Technology, 2015, 195: 254-264. doi: 10.1016/j.biortech.2015.06.129 [32] WANG D X, HAN Y X, HAN H J, et al. Enhanced treatment of Fischer-Tropsch wastewater using up-flow anaerobic sludge blanket system coupled with micro-electrolysis cell: A pilot scale study [J]. Bioresource Technology, 2017, 238: 333-342. doi: 10.1016/j.biortech.2017.04.056 [33] VU M T, NOORI M T, MIN B. Conductive magnetite nanoparticles trigger syntrophic methane production in single chamber microbial electrochemical systems [J]. Bioresource Technology, 2020, 296: 122265. doi: 10.1016/j.biortech.2019.122265 [34] SIEGERT M, YATES M D, CALL D F, et al. Comparison of nonprecious metal cathode materials for methane production by electromethanogenesis [J]. ACS Sustainable Chemistry & Engineering, 2014, 2(4): 910-917. [35] DOU Z O, DYKSTRA C M, PAVLOSTATHIS S G. Bioelectrochemically assisted anaerobic digestion system for biogas upgrading and enhanced methane production [J]. The Science of the Total Environment, 2018, 633: 1012-1021. doi: 10.1016/j.scitotenv.2018.03.255 [36] GOPAL J, HASAN N, MANIKANDAN M, et al. Bacterial toxicity/compatibility of platinum nanospheres, nanocuboids and nanoflowers [J]. Scientific Reports, 2013, 3: 1260. doi: 10.1038/srep01260 [37] 胡凯, 贾硕秋, 陈卫. 微生物电解池构型和电极材料研究综述 [J]. 能源环境保护, 2016, 30(5): 1-8,34. doi: 10.3969/j.issn.1006-8759.2016.05.001 HU K, JIA S Q, CHEN W. Review on configurations and electrode materials of microbial electrolysis cell [J]. Energy Environmental Protection, 2016, 30(5): 1-8,34(in Chinese). doi: 10.3969/j.issn.1006-8759.2016.05.001
[38] THRASH J C, van TRUMP J I, WEBER K A, et al. Electrochemical stimulation of microbial perchlorate reduction [J]. Environmental Science & Technology, 2007, 41(5): 1740-1746. [39] DASSARMA S, COKER J A, DASSARMA P. Archaea (overview)[M]//SCHAECHTER M. Encyclopedia of Microbiology (Third Edition). Oxford; Academic Press. 2009: 1-23. [40] ZEIKUS J G, WOLFE R S. Methanobacterium thermoautotrophicus sp. n., an anaerobic, autotrophic, extreme thermophile [J]. Journal of Bacteriology, 1972, 109(2): 707-715. doi: 10.1128/jb.109.2.707-713.1972 [41] MAH R A, SMITH M R, BARESI L. Studies on an acetate-fermenting strain of Methanosarcina [J]. Applied and Environmental Microbiology, 1978, 35(6): 1174-1184. doi: 10.1128/aem.35.6.1174-1184.1978 [42] GARCIA J-L, OLLIVIER B, WHITMAN W B. The order methanomicrobiales [J]. Prokaryotes, 2006, 3: 208-230. [43] ANGELIDAKI I, KARAKASHEV D, BATSTONE D J, et al. Chapter sixteen - Biomethanation and its potential[M]//ROSENZWEIG A C, RAGSDALE S W. Methods in Enzymology. Academic Press. 2011: 327-251. [44] LIU Y C, WHITMAN W B. Metabolic, phylogenetic, and ecological diversity of the methanogenic Archaea [J]. Annals of the New York Academy of Sciences, 2008, 1125: 171-189. doi: 10.1196/annals.1419.019 [45] DWORKIN M, FALKOW S, ROSENBERG E, et al. The Prokaryotes: Volume 3: Archaea. Bacteria: Firmicutes, Actinomycetes[M]. Springer, 2006. [46] THAUER R K, KASTER A K, SEEDORF H, et al. Methanogenic Archaea: Ecologically relevant differences in energy conservation [J]. Nature Reviews Microbiology, 2008, 6(8): 579-591. doi: 10.1038/nrmicro1931 [47] ANGELIDAKI I, KARAKASHEV D, BATSTONE D J, et al. Biomethanation and its potential [J]. Methods in Enzymology, 2011, 494: 327-351. [48] TIMMIS K N, MCGENITY T, VAN DER MEER J R, et al. Handbook of hydrocarbon and lipid microbiology[M]. Springer Berlin, 2010. [49] PAQUETE C M, ROSENBAUM M A, BAÑERAS L, et al. Let's chat: Communication between electroactive microorganisms [J]. Bioresource Technology, 2022, 347: 126705. doi: 10.1016/j.biortech.2022.126705 [50] COSTA N L, CLARKE T A, PHILIPP L A, et al. Electron transfer process in microbial electrochemical technologies: The role of cell-surface exposed conductive proteins [J]. Bioresource Technology, 2018, 255: 308-317. doi: 10.1016/j.biortech.2018.01.133 [51] BRUTINEL E D, GRALNICK J A. Shuttling happens: Soluble flavin mediators of extracellular electron transfer in Shewanella [J]. Applied Microbiology and Biotechnology, 2012, 93(1): 41-48. doi: 10.1007/s00253-011-3653-0 [52] STAMS A J M, PLUGGE C M. Electron transfer in syntrophic communities of anaerobic bacteria and Archaea [J]. Nature Reviews. Microbiology, 2009, 7(8): 568-577. doi: 10.1038/nrmicro2166 [53] ROTARU A E, SHRESTHA P M, LIU F H, et al. Inter species electron transfer via hydrogen and formate rather than direct electrical connections in cocultures of Pelobacter carbinolicus and Geobacter sulfurreducens [J]. Applied and Environmental Microbiology, 2012, 78(21): 7645-7651. doi: 10.1128/AEM.01946-12 [54] HUANG L Y, LIU X, YE Y, et al. Evidence for the coexistence of direct and riboflavin-mediated inter species electron transfer in Geobacter co-culture [J]. Environmental Microbiology, 2020, 22(1): 243-254. doi: 10.1111/1462-2920.14842 [55] LIU T, YU Y Y, CHEN T, et al. A synthetic microbial consortium of Shewanella and Bacillus for enhanced generation of bioelectricity [J]. Biotechnology and Bioengineering, 2017, 114(3): 526-532. doi: 10.1002/bit.26094 [56] ENGEL C, SCHATTENBERG F, DOHNT K, et al. Long-term behavior of defined mixed cultures of Geobacter sulfurreducens and Shewanella oneidensis in bioelectrochemical systems [J]. Frontiers in Bioengineering and Biotechnology, 2019, 7: 60. doi: 10.3389/fbioe.2019.00060 [57] 王弋博, 武春媛, 周顺桂. 腐殖质在Comamonas koreensis CY01介导的2, 4-二氯苯氧乙酸还原脱氯过程中的作用 [J]. 草业学报, 2011, 20(1): 248-252. doi: 10.11686/cyxb20110134 WANG Y B, WU C Y, ZHOU S G. Effect of humic substances on the reductive dechlorination of 2, 4-dichlorophenoxyacetic acid by Comamonas koreensis CY01 [J]. Acta Prataculturae Sinica, 2011, 20(1): 248-252(in Chinese). doi: 10.11686/cyxb20110134
[58] LOVLEY D R, FRAGA J L, COATES J D, et al. Humics as an electron donor for anaerobic respiration [J]. Environmental Microbiology, 1999, 1(1): 89-98. doi: 10.1046/j.1462-2920.1999.00009.x [59] SUMMERS Z M, FOGARTY H E, LEANG C, et al. Direct exchange of electrons within aggregates of an evolved syntrophic coculture of anaerobic bacteria [J]. Science, 2010, 330(6009): 1413-1415. doi: 10.1126/science.1196526 [60] REGUERA G, MCCARTHY K D, MEHTA T, et al. Extracellular electron transfer via microbial nanowires [J]. Nature, 2005, 435(7045): 1098-1101. doi: 10.1038/nature03661 [61] LIU F H, ROTARU A E, SHRESTHA P M, et al. Magnetite compensates for the lack of a pilin-associated c-type cytochrome in extracellular electron exchange [J]. Environmental Microbiology, 2015, 17(3): 648-655. doi: 10.1111/1462-2920.12485 [62] HA P T, LINDEMANN S R, SHI L, et al. Syntrophic anaerobic photosynthesis via direct inter species electron transfer [J]. Nature Communications, 2017, 8: 13924. doi: 10.1038/ncomms13924 [63] 刘星, 周顺桂. 微生物纳米导线的导电机制及功能 [J]. 微生物学报, 2020, 60(9): 2039-2061. doi: 10.13343/j.cnki.wsxb.20200177 LIU X, ZHOU S G. Electrical conductivity and application of microbial nanowires [J]. Acta Microbiologica Sinica, 2020, 60(9): 2039-2061(in Chinese). doi: 10.13343/j.cnki.wsxb.20200177
[64] MCGLYNN S E, CHADWICK G L, KEMPES C P, et al. Single cell activity reveals direct electron transfer in methanotrophic consortia [J]. Nature, 2015, 526(7574): 531-535. doi: 10.1038/nature15512 [65] XIONG Y, SHI L, CHEN B, et al. High-affinity binding and direct electron transfer to solid metals by the Shewanella oneidensis MR-1 outer membrane c-type cytochrome OmcA [J]. Journal of the American Chemical Society, 2006, 128(43): 13978-13979. doi: 10.1021/ja063526d [66] PARK J H, PARK J H, JE SEONG H, et al. Metagenomic insight into methanogenic reactors promoting direct inter species electron transfer via granular activated carbon [J]. Bioresource Technology, 2018, 259: 414-422. doi: 10.1016/j.biortech.2018.03.050 [67] KATO S, HASHIMOTO K, WATANABE K. Methanogenesis facilitated by electric syntrophy via (semi)conductive iron-oxide minerals [J]. Environmental Microbiology, 2012, 14(7): 1646-1654. doi: 10.1111/j.1462-2920.2011.02611.x [68] GIANG H N, ZHANG J, ZHU Z Y, et al. Single-chamber microbial electrochemical cell for CH4 production from CO2 utilizing a microbial consortium [J]. International Journal of Energy Research, 2018, 42(3): 1308-1315. doi: 10.1002/er.3931 [69] CHOI K S, KONDAVEETI S, MIN B. Bioelectrochemical methane (CH4) production in anaerobic digestion at different supplemental voltages[J]. Bioresource Technology, 2017, 245(Pt A): 826-832. [70] PARK J, LEE B, TIAN D, et al. Bioelectrochemical enhancement of methane production from highly concentrated food waste in a combined anaerobic digester and microbial electrolysis cell [J]. Bioresource Technology, 2018, 247: 226-233. doi: 10.1016/j.biortech.2017.09.021 [71] 蔡文忠, 张希晨, 周耀辉. MEC/AnMBR反应器组合处理生活污水 [J]. 南华大学学报(自然科学版), 2017, 31(2): 107-112. CAI W Z, ZHANG X C, ZHOU Y H. Combination of MAC/MBR reactor for domestic sewage treatment [J]. Journal of University of South China (Science and Technology), 2017, 31(2): 107-112(in Chinese).
[72] LIU H B, LV Y C, XU S Y, et al. Configuration and rapid start-up of a novel combined microbial electrolytic process treating fecal sewage [J]. The Science of the Total Environment, 2020, 705: 135986. doi: 10.1016/j.scitotenv.2019.135986 期刊类型引用(0)
其他类型引用(1)
-

 点击查看大图
点击查看大图
计量
- 文章访问数: 5824
- HTML全文浏览数: 5824
- PDF下载数: 135
- 施引文献: 1
