

-
有机化学品泄漏而造成的地下水和土壤污染一直是广受公众关注的问题,而随着我国双碳战略的发布,生物修复作为一种低成本、低碳绿色的污染场地修复技术逐渐受到科学界和工程界的青睐[1]。微生物在自然界中广泛存在,有的菌种可以利用有机污染物作为自身代谢的碳源和能源来降解污染物,例如使用Dehalococcoides菌可将PCE还原脱氯为乙烯[2],从土壤中分离出的芽孢杆菌Bacillus salmalaya 139SI能降解石油烃[3]。但是本地微生物的数量和活性取决于复杂地下环境,可能难以对污染物进行有效的降解[4]。此时,可以通过添加生物降解所需的营养物质来提高降解效率,即生物刺激,而生物强化方法是通过引入针对特定污染物的外源微生物来提高总体修复率。当从外部向场地添加微生物或营养物质时,场地中微生物和溶质传输效率是决定生物修复效果的关键所在。
现有污染场地生物降解数学模型主要关注有机污染物和附着在土壤上微生物之间的生物反应,并假定微生物固定不动且数量无限[5]。但是,污染含水层中大量微生物活跃在水相中并进行移动[6],尤其是当注入外源微生物时。因此,研究微生物在地下土壤环境中水相和固相中的运移对准确评估生物降解至关重要。微生物通常可以被视作一种生物胶体,同时具备胶体和生物性质[7],近十年来国内外学者通过实验[8-9]和现场试验[10-11]等方法研究了多孔介质中微生物迁移规律。同时,由于地下环境中微生物运移机制复杂,不同的微生物运移数学模型会针对具体研究问题进行假设和简化[12]。其中,细菌类型、温度、土层特性、pH值、含氧量、营养物质和有机物类型等重要因素都会影响微生物的运移。
污染地层内往往存在一些天然裂隙和人工裂隙[13],是微生物的优先运移通道,但也会使得微生物和溶质的运移变得更加复杂。目前,关于微生物在裂隙通道运移的研究较少,且主要集中在岩石裂隙的石油开采方面[14],部分适用于微生物采油的结论可能不适用于生物强化修复方法。目前尚未有文献研究生物修复技术体系下的微生物沿裂隙-基质运移。因此,本研究采用多场耦合仿真软件COMSOL建立了裂隙-基质介质中微生物和溶质反应性输运模型,揭示生物强化技术中微生物运移及降解机制。模型耦合了裂隙-基质介质内微生物的重要物理化学和生物现象,并考虑了注入微生物、电子受体和残留污染物之间的相互联系。最后,研究了微生物注入速度、微生物在基质内扩散系数和动力学吸附/解吸速率系数等对微生物修复效果的影响规律。
-
图1为裂隙-基质介质中微生物和溶质运移模型示意图。模型为二维轴对称,微生物和溶解氧通过裂隙左侧以固定流速注入,而污染物被固定在基质中,设裂隙宽度为2B,平行裂隙系统之间的距离为2H。此外,模型的其他基本假设还包括:1) 微生物和溶解氧均可扩散入基质中;2) 基质完全饱和、均质、各向同性,且分子扩散是基质的主要传输机制,对流忽略不计;3) 微生物分为游离相和吸附相,二者具有相似的表面特征和成长特征;4) 微生物仅将溶解氧和污染物作为生长消耗项,前者为微生物提供电子受体,后者提供电子供体;5) 不考虑基质中污染物和溶解氧的吸附[15]。
-
微生物自身特性 (如形状大小、生长状态、趋化性) 、多孔介质特性 (如土颗粒大小、含水率、渗透系数) 和环境因素 (如pH值、温度、离子浓度) 都与微生物运移关系密切,因此在进行微生物运移建模时需进行一定程度的简化和假设。微生物在多孔介质内的运移可以简化归为基本的对流扩散,吸附和自身的生物过程。其中裂隙中游离相微生物可表示为:
式中:Cmf是裂隙中游离相微生物的质量浓度 (kg·m−3) ;
$ {v}_{p} $ 是裂隙内流动速度 (m·s−1) ;$ {D}_{m} $ 是微生物扩散系数 (m2·s−1) ;$ {Q}_{\mathrm{m}\mathrm{f}}^{{\mathrm{s}}} $ 是裂隙中游离相微生物的动力学传质速率 (kg·m−3·s−1) ;$ {Q}_{\mathrm{m}\mathrm{f}}^{\mathrm{g}} $ 和$ {Q}_{\mathrm{m}\mathrm{f}}^{\mathrm{d}} $ 分别是裂隙中游离相微生物的生长和衰亡速率 (kg·m−3·s−1) 。当微生物在多孔介质中迁移时,会通过吸附/解吸过程在游离相和吸附相之间转换。静电吸引或排斥、范德华吸引力、疏水作用、偶极力和氢键是控制微生物在多孔介质界面上吸附的主要力量。前人一般将多孔介质中微生物的吸附过程考虑呈可逆或不可逆的平衡或动力学过程,本研究采用考虑吸附位点的一级动力反应式来表达:
式中:katt和kdtt分别为微生物吸附和解吸系数 (s−1) ;
$ {S}_{\mathrm{m}\mathrm{f}} $ 为裂隙中吸附相微生物的质量浓度 (kg·m−3) ;$ {\rho }_{\mathrm{m}} $ 为微生物质量密度 (kg·m−3) ;$ {\psi }_{\mathrm{p}\mathrm{s}\mathrm{i}} $ 为吸附点位系数,在裂隙中其值设为1。微生物对有机或无机化合物的利用可以通过多种方式表示,其中Monod动力学被广泛地用于描述微生物代谢过程中所需组分的利用速率。本文采用改进的双底物Monod方程来描述微生物的生长率,而微生物的衰亡过程则可视为一级动力学过程。因此,裂隙内游离相微生物的生长项和衰亡项如下:
式中:
$ {\mu }_{\mathrm{m}\mathrm{a}\mathrm{x}} $ 为微生物的最大比生长速率 (s−1) ;$ {C}_{\mathrm{o}\mathrm{f}} $ 和$ {C}_{\mathrm{c}\mathrm{f}} $ 分别为裂缝水中溶解的氧气和污染物浓度 (kg·m−3) ;$ {K}_{\mathrm{o}} $ 和$ {K}_{\mathrm{c}} $ 分别为溶解氧和污染物的半饱和常数 (kg·m−3) ,其值等于微生物比生长速率为最大值一半时营养液的浓度;$ {k}_{\mathrm{d}\mathrm{e}\mathrm{t}} $ 为微生物的衰亡系数 (s−1) 。将式(2)~(4)代入式(1)可得裂隙中游离相微生物的运移传输方程为:
吸附相微生物也同样经历生长和衰亡过程。若假设游离相微生物的生长和衰亡参数
$ {\mu }_{\mathrm{m}\mathrm{a}\mathrm{x}} $ 、$ {K}_{\mathrm{o}} $ 、$ {K}_{\mathrm{c}} $ 和$ {k}_{\mathrm{d}\mathrm{e}\mathrm{t}} $ 同时适用于吸附相[16],则裂隙中吸附相微生物的运移方程可表示为:式中:式(6)右边的四项分别代表微生物的吸附、解吸、生长和衰亡。
考虑分子扩散和吸附点位系数
$ {\psi }_{\mathrm{p}\mathrm{s}\mathrm{i}} $ 与吸附量相关的Lagrange函数,基质中游离相微生物的运移传输方程为:式中:
$ {C}_{\mathrm{m}\mathrm{m}} $ 为基质中游离相微生物的质量浓度 (kg·m−3) ;$ {C}_{\mathrm{o}\mathrm{m}} $ 和$ {C}_{\mathrm{c}\mathrm{m}} $ 分别为基质孔隙水中溶解氧和污染物的水相浓度 (kg·m−3);$ {S}_{\mathrm{m}\mathrm{m}} $ 为基质中吸附相微生物的质量浓度 (kg·m−3) ;$ {\sigma }_{\mathrm{m}\mathrm{a}\mathrm{x}} $ 为最大吸附系数。基质中吸附相微生物的运移传输方程可表示为:
-
本研究将氧看作溶解性组分,在裂隙-基质环境下运移时会被作为电子受体被微生物代谢消耗,溶解氧的消耗速率表示为:
式中:
$ {Y}_{\mathrm{o}} $ 为溶解氧产出系数,表示消耗单位溶解氧可生成的微生物量的比值。因此,裂缝中溶解氧的运移传输方程可表示为:
式中:
$ {D}_{\mathrm{o}} $ 为溶解氧的扩散系数 (m2·s−1) 。基质中溶解氧的运移传输方程可表示为:
-
本研究将污染物也假定为溶解性组分,但不考虑其在固相的吸附。因此,裂缝和基质中溶解态污染物的运移传输方程可表示为:
式中:
$ {Y}_{\mathrm{c}} $ 为污染物产出系数,表示消耗单位污染物可生成的微生物量的比值;$ {D}_{\mathrm{c}} $ 为污染物的扩散系数 (m2·s−3) 。 -
首先对前一节介绍的传输模型进行验证。由于目前尚无关于裂隙-基质介质中微生物和溶质混合反应迁移的实验数据,本研究选取2个室内微生物运移柱实验来验证本模型微生物运移的合理性。第一个实验[9],土柱长30 cm,入口处先注入微生物培养液1 h,然后注入去离子水进行驱替,流速为0.02 mm·s−1。图2(a)对比了本模型计算结果与该实验数据,可以看出两者的游离相微生物浓度规律较为一致。第二个实验[17],土柱内长17 cm,内径3.3 cm,入口处先注入20 mL大肠杆菌液,然后注入背景溶液进行驱替,流速为0.42 cm·min−1。图2(b)对比了本模型计算结果与该实验数据,可以看到两者的突破浓度曲线也较为一致。因此,通过数值模拟结果与两组实验数据的对比,很好的验证了本模型的合理性和正确性。
针对研究问题进行具体工况的计算。本研究数值模型由一组微生物、溶解氧和污染物传输的偏微分方程组成,采用COMSOL的自定义偏微分方程模块建立方程组并进行求解,以模拟生物强化技术中沿裂隙注入微生物和电子受体来修复污染土壤的过程。该二维轴对称模型包括两个区域:裂隙和基质,以及8种成分:裂缝中游离的微生物、裂缝表面吸附的微生物、基质孔隙游离的微生物、基质吸附的微生物、裂缝中的溶解氧、基质中的溶解氧、裂缝中的溶解态污染物、基质中的溶解态污染物。裂缝-基质界面的连续性是以通量和浓度连续为条件,并通过在每个时间步迭代解来获得的,采用隐式有限差分格式进行数值求解。由于裂缝的流动与管道流动非常相似,假设裂隙孔隙率为100%。表1列出了裂缝-基质中微生物修复的相关参数。为了便于计算,微生物、溶解氧和污染物的扩散系数视为相同,同时注入的微生物和溶解氧浓度设为1 mg·L−1,污染浓度也设为1 mg·L−1。
-
图3给出了沿裂隙持续注入 (40 d) 微生物和溶解氧后不同溶质浓度的时空分布。可以看到,在注入开始时 (1 d内) ,微生物主要分布在裂缝注入口附近。之后,微生物逐渐扩散到基质中,虽然注入口附近仍然浓度不变,但相邻基质内微生物的运移范围逐渐变大。20 d后微生物的扩散范围基本不变且浓度变化不大,表明由于微生物生长所需的底物 (污染物) 被消耗殆尽,在吸附、解吸附以及生长衰亡等多种物化生物作用下微生物的运移逐渐达到平衡状态。溶解氧随同微生物一起被注入污染场地,其分布云图如图4所示。与微生物相比,溶解氧的运移十分迅速,在注入的前10 d,溶解氧主要是沿着裂缝快速移动,之后逐渐向基质内部扩散;第40 d时,溶解氧已经几乎充满整个基质。由此可见,溶解氧和微生物的运移具有很大的区别,溶解氧的运移范围远远比微生物大,浓度也高的多,因此在本模拟工况下几乎可以忽略溶解氧运移的影响 (溶解氧不会成为微生物运移和降解活动的限制条件) 。
图5给出了场地污染物浓度空间分布随修复菌剂注入时间的变化。可以看出,首先是在裂缝入口处附近的污染物由于受到微生物降解被去除,随后周边的污染物由于浓度梯度向裂隙入口逐渐扩散移动并被微生物继续降解,导致生物修复实际影响范围随着时间的增加而渐渐扩大。当注入20 d后,整体修复率可以达到58.9%;注入40 d后,基质内污染物已经基本得到去除,修复率达到93.8%。
-
微生物在基质中运移主要呈现为受孔隙介质制约的布朗运动,微生物的扩散程度与基质孔隙度、孔隙大小和菌体大小密切相关,当基质孔隙度很大时,基质扩散系数也随之变高。为了便于研究介质孔隙特性对生物强化的影响,本研究假设微生物与溶质扩散系数相同,结果如图6所示。可以发现,基质介质扩散系数的增大会显著提高含裂隙污染地层中污染物的去除率。例如,当扩散系数为10−6 m2·s−1时,仅通过12 d就能达到100%去除率;而当扩散系数为10−8 m2·s−1时,40 d的修复率不足10%。由此可见,污染地层中基质扩散系数是影响生物强化修复效率的关键因素。
当多孔介质的孔隙明显大于细胞大小时,微生物滞留在介质的主要机制是吸附。SCHIJVEN等[21]发现在微生物迁移过程中平衡吸附可以忽略不计。因此,采用一阶吸附动力学模型来表征微生物的吸附特性,其中katt是表征吸附速率的常数。图7描述了吸附系数katt对污染修复效率的影响。结果表明,随着katt的增加,地层内污染物的修复率不断降低,当katt从1 d−1上升到5 d−1时,修复效率的降低最为明显,说明微生物的强吸附会延缓微生物在介质中的运移范围,从而减少微生物能够利用的底物,降低了修复效率。
-
在实际工程应用中,微生物菌液注入速度是一个非常很重要的设计参数。图8给出了不同注入速度条件下污染地层的修复效率。结果表明,修复率随着注入速度的加快而变高,随着注入速度由0.1 m·d−1增加到1 m·d−1 (注入浓度为1 mg·L−1),第40 d的修复率从90%上升到100%。但是,注入速度并不是越大越好,越大意味着更大的注入压力,即修复成本和技术难度的增加,并且可能会影响微生物的活性。值得注意的是当注入速度高于0.5 m·d−1 时,修复率随注入速度的增加而造成的增幅明显减小,因此对于本模拟工况来说,注入速度超过0.5 m·d−1之后再提高注入速度意义不大。对于其他某一特定工况,也可能存在一个临界速度,注入速度高于临界速度时注入速度对修复效率影响不显著,因此实际工程中注入时推荐选择临界速度作为注入速度。
-
1) 注入微生物后,微生物运移逐渐达到平衡并主要聚集在入口附近的基质中,而注入的溶解氧可快速沿裂隙均匀分布并逐渐向基质内扩散,基质内的污染物随着时间被逐渐去除,在污染物反向扩散的作用下生物修复的实际影响范围可能要比微生物的运移范围大。
2) 对于本研究情景中的裂隙污染地层生物强化修复,微生物基质扩散系数的变化对污染物生物修复效果有着显著影响,微生物吸附系数和注入速度的变化也对污染物生物修复效果有一定影响。提高强化生物修复效率的关键是提高微生物和污染物的接触程度。
含裂隙污染场地中微生物运移及降解修复规律
Study on microbial transport and biodegradation in fracture contaminated sites
-
摘要: 原位生物修复作为一种低成本、绿色的污染场地修复技术受到了极大的关注。大多数生物修复研究集中增强土著微生物自身的降解能力的生物刺激技术上,对引入外源微生物的生物强化技术研究有限。而污染场地内天然和人工裂隙等优势渗流会影响注入微生物的运移及其降解反应。通过提出一个数学模型来模拟裂隙含水层中微生物和相关溶质迁移以及污染物降解过程。该模型考虑了对流、扩散、吸附、生长、死亡和微生物代谢反应等过程,并耦合了微生物、电子受体和基质底物三者之间的相互联系,并与试验结果验证良好。模拟结果分析表明,保持微生物与污染物之间的充分接触并提供充足的养分是提高生物修复效率的关键。由于生物降解消耗加速了基质内污染物向裂缝周边区域的反向扩散,生物修复的实际影响范围比微生物运移范围要大。同时,裂隙外基质的扩散系数对生物修复效果的影响最大,其次是微生物吸附系数和注入速度。该数学模型是评估含裂隙地层生物强化策略有效性的重要工具,有助于设计和优化相应的生物修复方法。Abstract: In-situ bioremediation, as a low-cost and environmentally friendly technique for the remediation of contaminated sites, has garnered significant attention. Current research on bioremediation techniques largely focuses on biostimulation techniques that enhance the inherent degradation capabilities of indigenous microorganisms. Limited research exists on bioaugmentation techniques involving the introduction of exogenous microorganisms. While dominant seepage such as natural and artificial fissures within a contaminated site can affect the transport of injected microorganisms and their degradation reactions. A mathematical model was proposed to stimulate the migration of microorganisms, solutes, and contaminant degradation processes within fractured aquifers. The model accounts for processes including advection, diffusion, adsorption, growth, death, and microbial metabolic reactions. It further integrates the interrelationships among microorganisms, electron acceptors, and substrate, and in good agreement with experimental results. The results show that sustaining optimal contact between microorganisms and contaminants, while providing ample nutrients, constitutes a critical factor in enhancing bioremediation efficiency. Due to biodegradation accelerating the reverse diffusion of contaminants within the matrix to the region surrounding the fracture, the impact scope of bioremediation extending beyond microbial transport zones. Simultaneously, the diffusion coefficient of the extracellular matrix in fractures exhibits the most significant influence on bioremediation efficacy, followed by microbial adsorption coefficients and injection rates. The mathematical model presented herein stands as a vital tool for assessing the effectiveness of bioaugmentation strategies within fractured geological formations, thereby aiding the design and optimization of pertinent bioremediation approaches.
-
Key words:
- contaminant /
- microorganisms /
- fractures /
- transport /
- biodegradation /
- numerical simulation
-

-

图 2 数值模型与微生物运移实验突破曲线验证
Figure 2. Comparison with experimental data for bacterial breakthrough
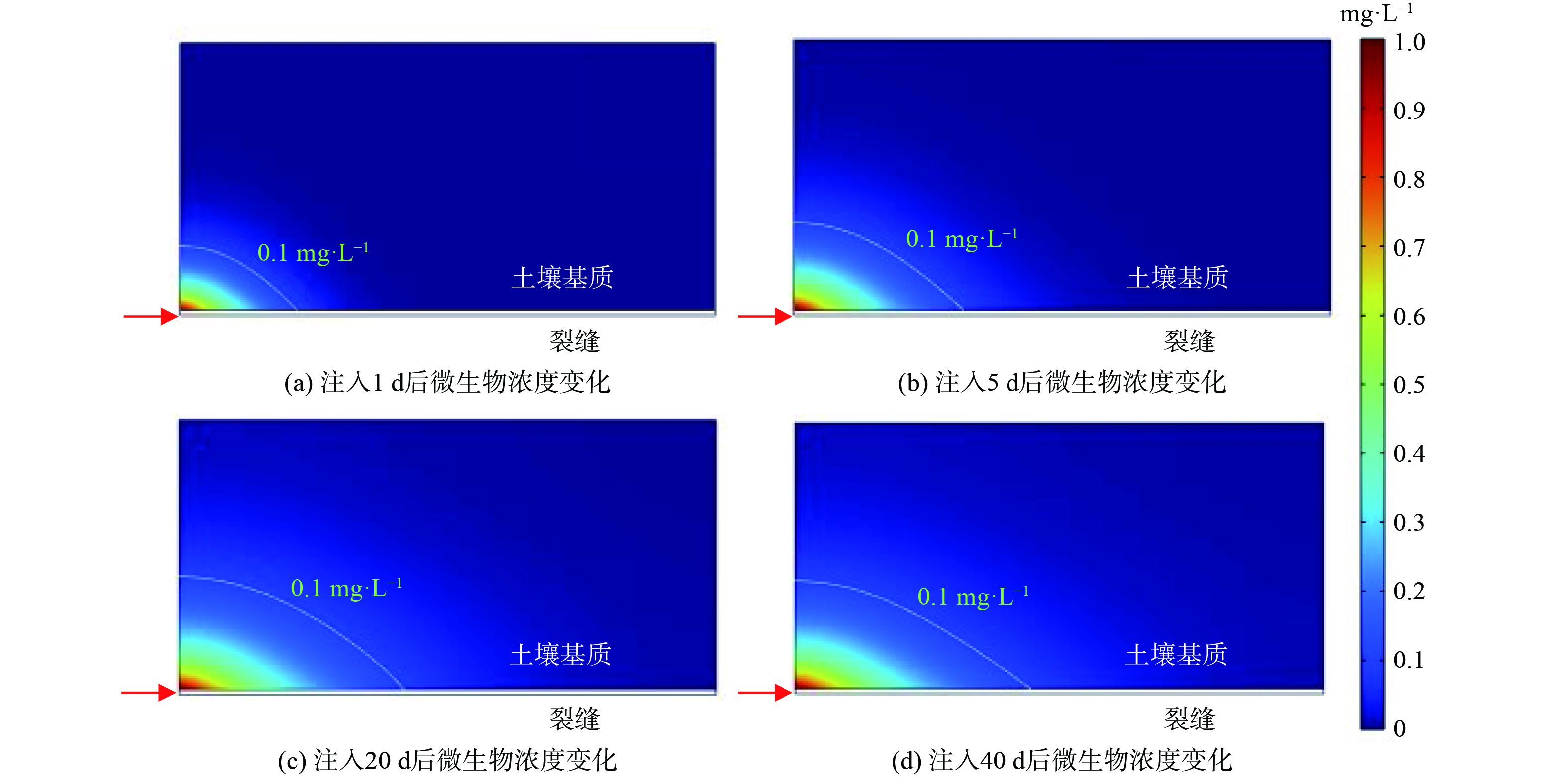
图 3 注入菌剂和溶解氧后微生物浓度时空分布变化
Figure 3. Changes in spatial and temporal distribution of microbial concentration after bacteria and dissolved oxygen injection
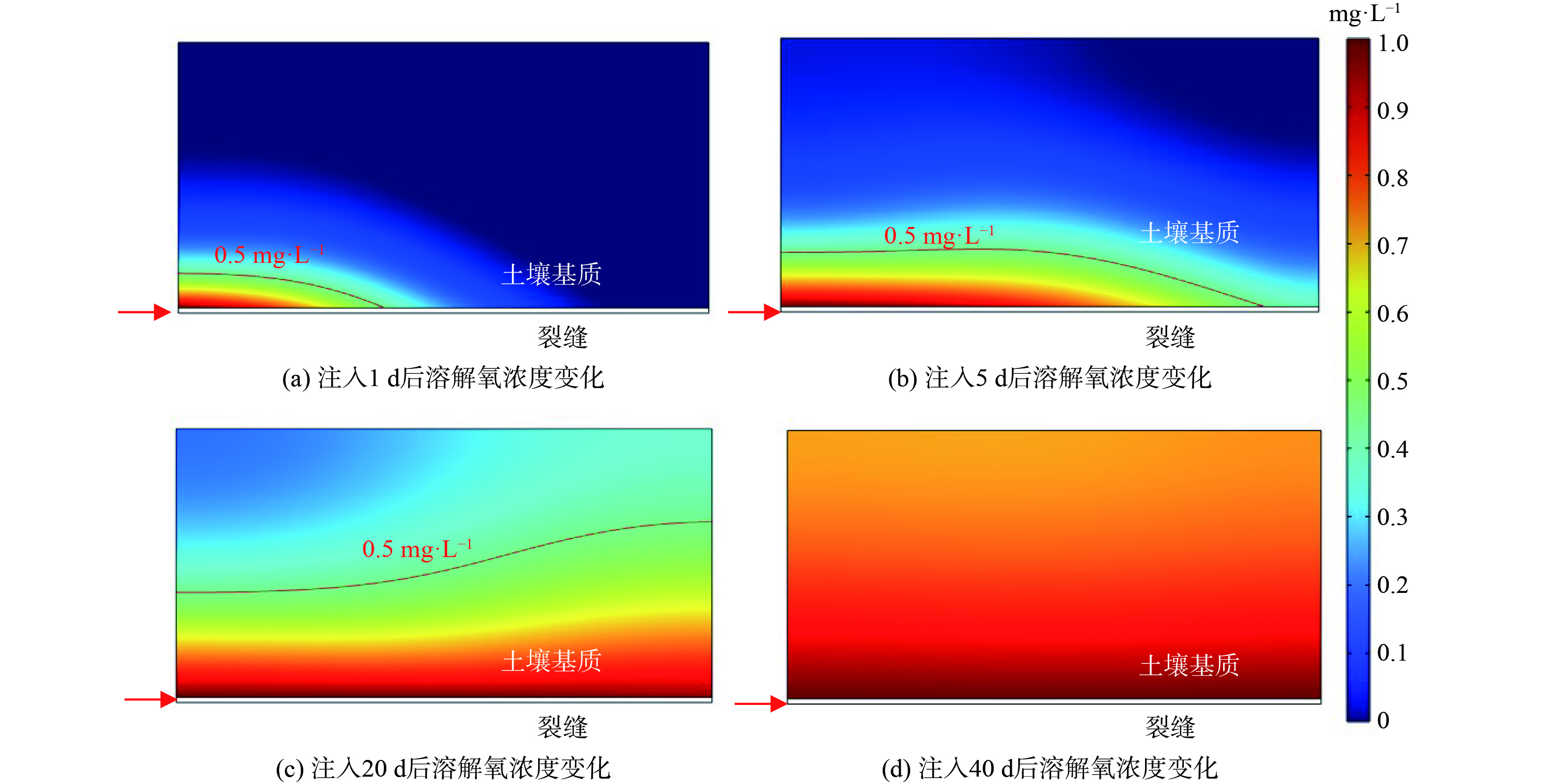
图 4 注入菌剂和溶解氧后溶解氧浓度时空分布变化
Figure 4. Changes in spatial and temporal distribution of dissolved oxygen concentration after bacteria and dissolved oxygen injection
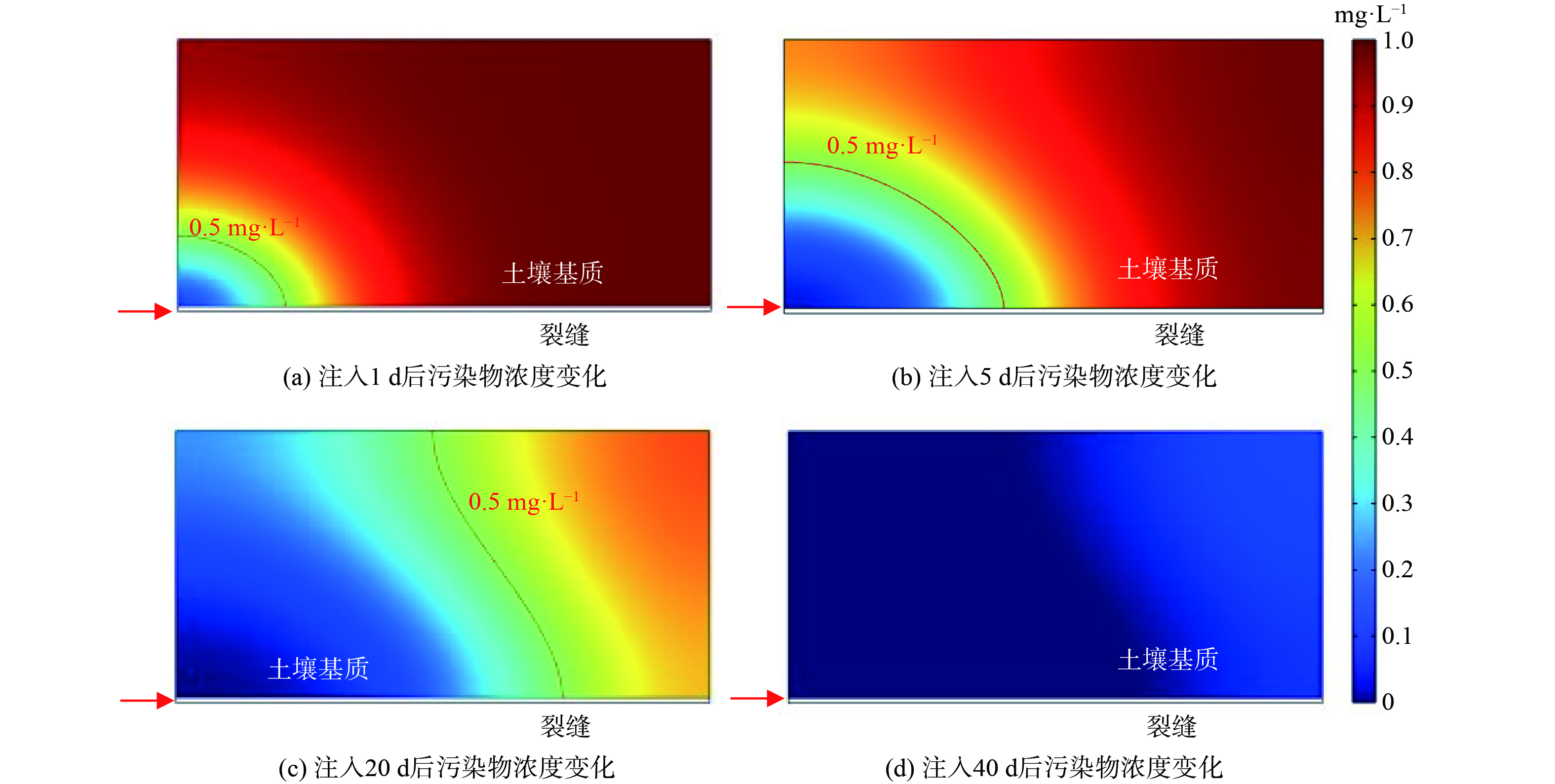
图 5 注入菌剂和溶解氧后污染物浓度时空分布变化
Figure 5. Changes in spatial and temporal distribution of contaminant concentrations after bacteria and dissolved oxygen injection
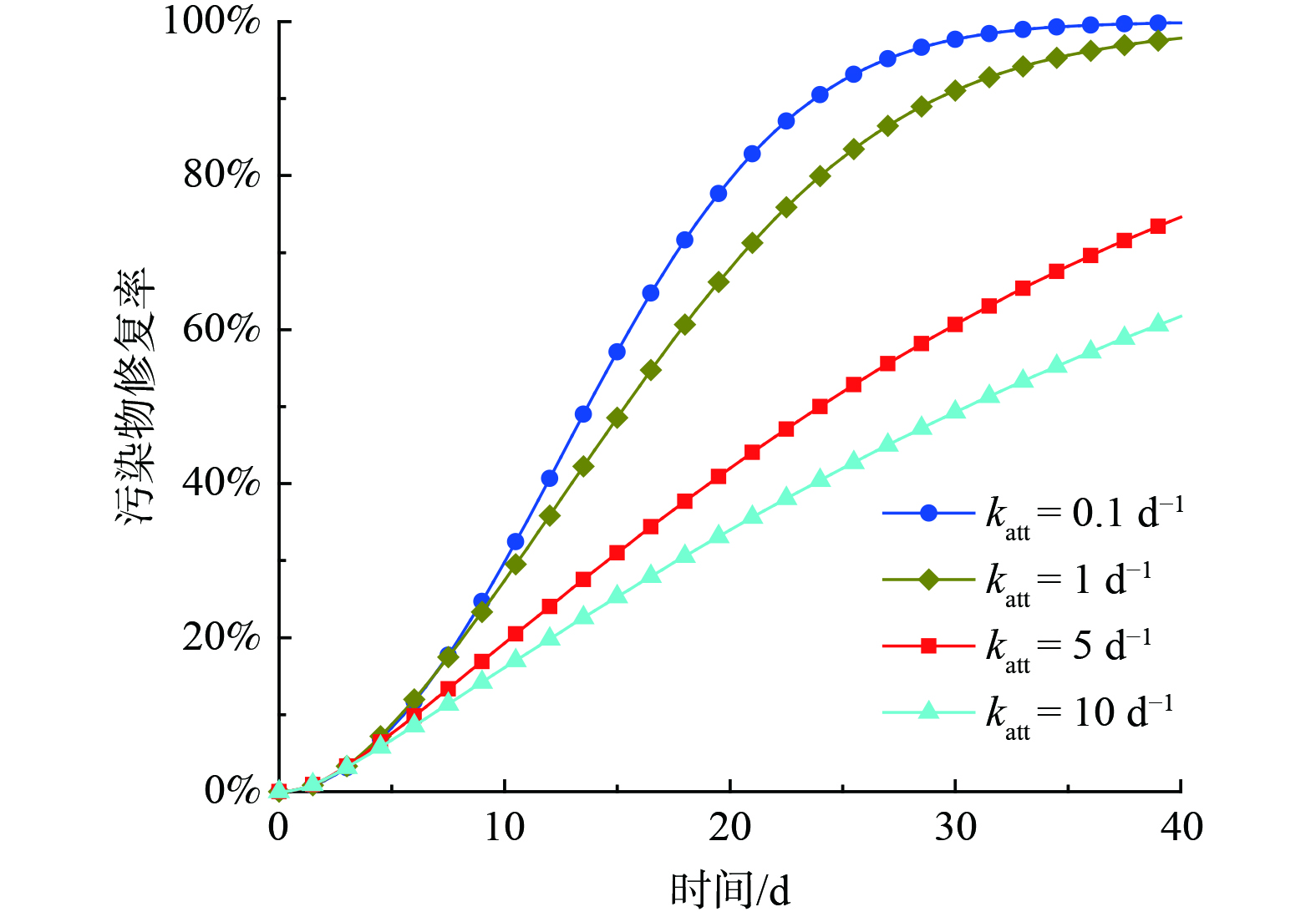
图 6 基质扩散系数对污染物修复效率的影响
Figure 6. Influence of diffusion coefficient on contaminant remediation efficiency
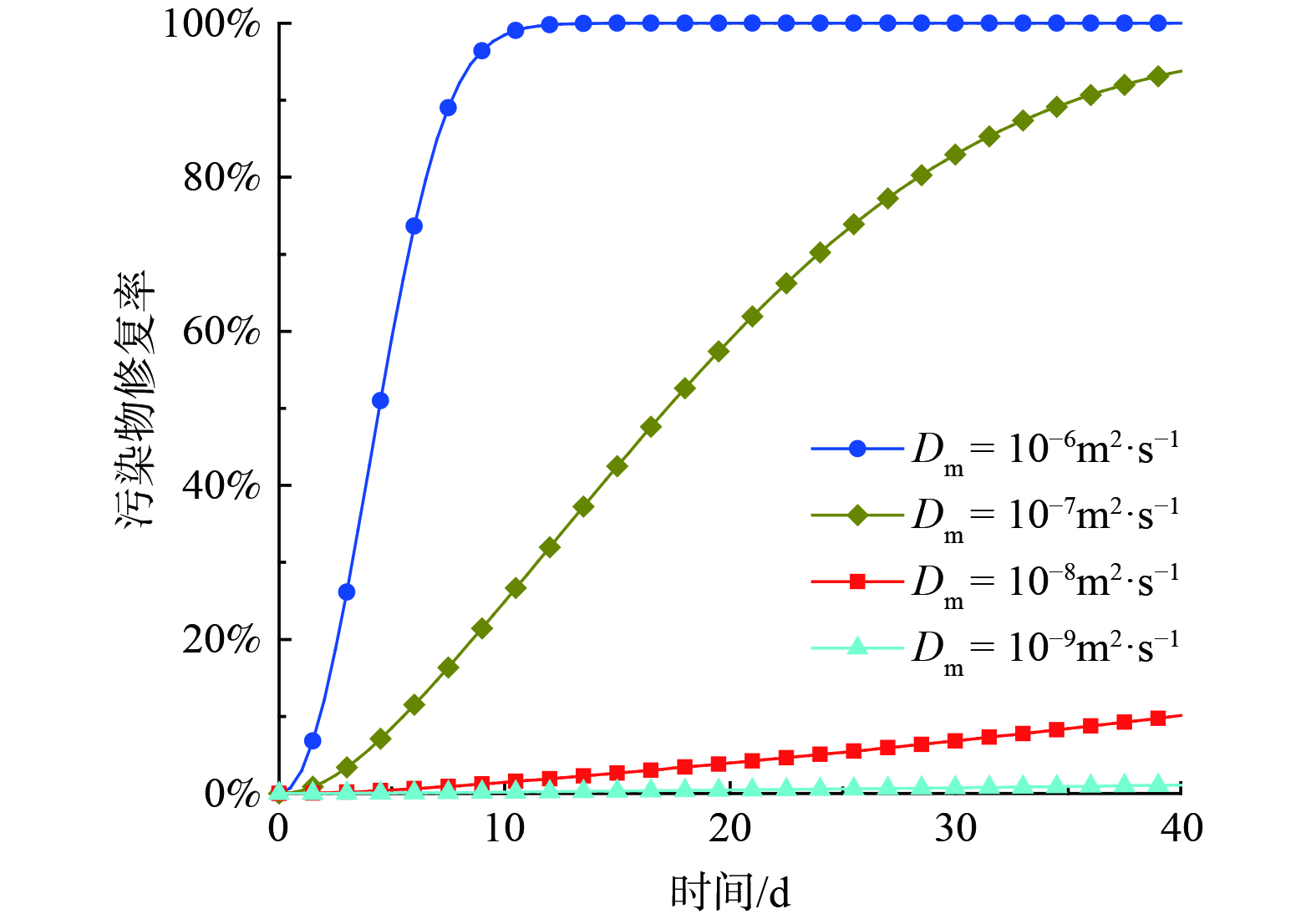
图 7 基质吸附系数对污染物修复效率的影响
Figure 7. Influence of adsorption coefficient on contaminant remediation efficiency
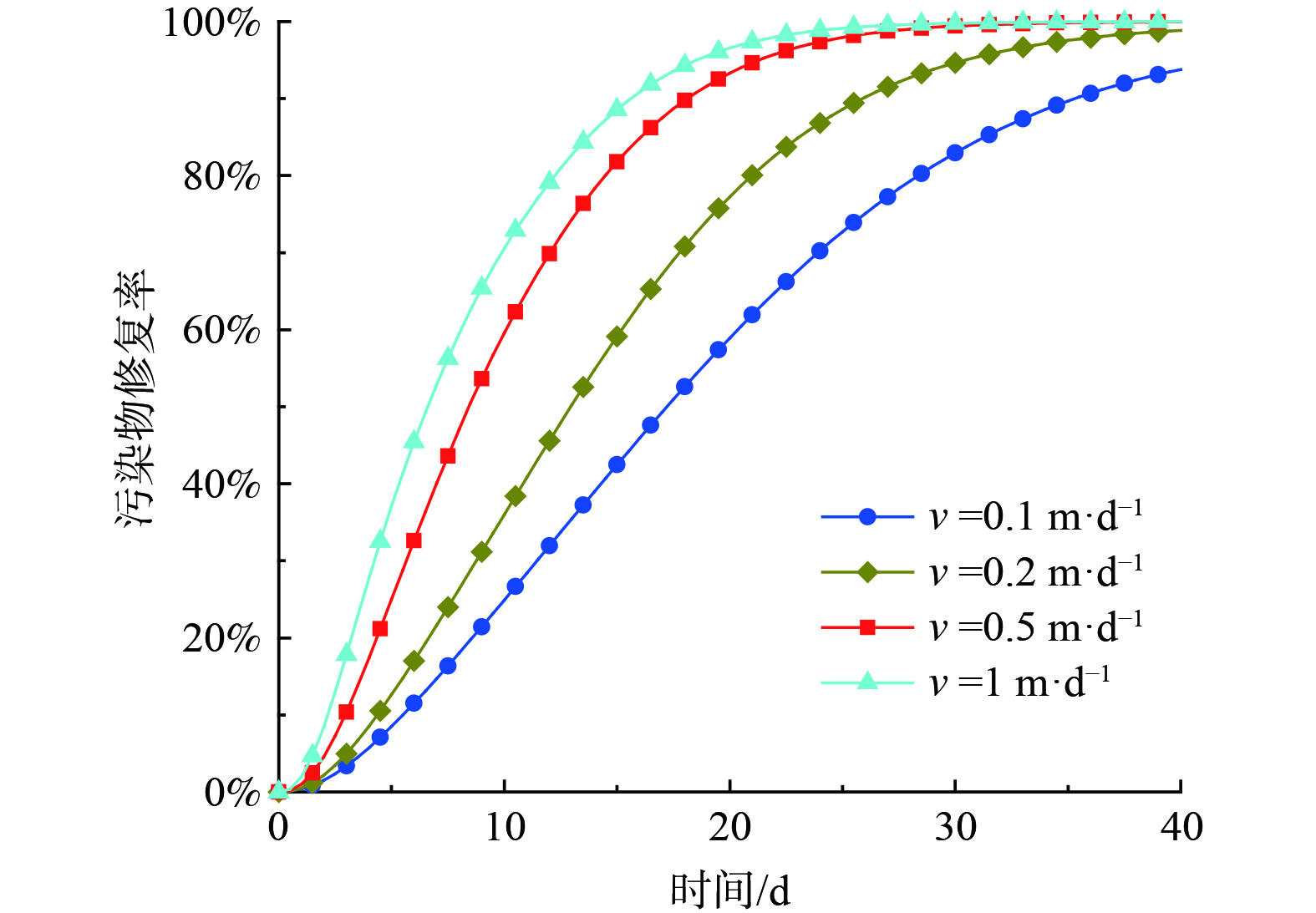
图 8 微生物菌液的注入速度对污染物修复效率的影响
Figure 8. Influence of the injection velocity of bacteria on contaminant remediation efficiency
表 1 数值模拟参数
Table 1. Parameters used for numerical simulation
参数 解释 2B = 2 cm 裂隙宽度 L = 1 m 裂隙长度 2H = 1 m 裂隙间距 θ = 0.25 基质孔隙率 Cm0 = Cn0 = Co0 = 1 mg·L−1 注入微生物、溶解氧和初始污染物浓度 Dm = Dn = Do = 10−6 m2·s−1 微生物、溶解氧和污染物扩散系数 ka = 1.73 d−1 微生物吸附系数[18] kr = 0.17 d−1 微生物解吸系数[18] μmax = 2 微生物最大生长系数[19] kd = 0.1 d−1 微生物衰亡系数[20] Kc = 2 mg·L−1 污染物半饱和系数[19] Yc = 0.40 污染物利用系数[19] Ko = 1 mg·L−1 溶解氧半饱和系数[19] Yo = 0.50 溶解氧利用系数[19] σmax = 0.02 微生物最大附着系数  下载: 导出CSV
下载: 导出CSV
-
[1] SALES I G, GOMES F C, PADILHA N M, et al. Soil bioremediation: Overview of technologies and trends[J]. Energies, 2020, 13(18): 4664. [2] TAKEUCHI M, KAWABE Y, WATANABE E, et al. Comparative study of microbial dechlorination of chlorinated ethenes in an aquifer and a clayey aquitard. Journal of contaminant hydrology[J]. 2011, 124(1-4): 14-24. [3] ISMAIL S, DADRASNIA A. Biotechnological potential of Bacillus salmalaya 139SI: a novel strain for remediating water polluted with crude oil waste[J]. PLoS ONE, 2015, 10(4): e0120931. [4] SCOW K M. Soil microbial communities and carbon flow in Agroecosystems[J]. Ecology in agriculture, 1997, 367. [5] MANOLI G, CHAMBON J C, BJERG P L, et al. A remediation performance model for enhanced metabolic reductive dechlorination of chloroethenes in fractured clay till[J]. Journal of contaminant hydrology, 2012, 131(1-4): 64-78. [6] SINGH R, OLSON M S. Application of bacterial swimming and chemotaxis for enhanced bioremediation[M]. Emerging Environmental Technologies, 2008, 149-172. [7] BAI H, CHEN J, HU Y, et al. Biocolloid transport and deposition in porous media: A review[J]. Korean Journal of Chemical Engineering, 2022, 39(1): 38-57. [8] 黄斯艺, 何江涛, 劳天颖, 等. 纳米乳化油修复硝酸盐污染地下水过程中的微生物特征模拟实验研究[J]. 环境科学学报, 2020, 40(4): 1242-1249. [9] TAN Y, GANNON J T, BAVEYE P, ALEXANDER M. Transport of bacteria in an aquifer sand: Experiments and model simulations[J]. Water Resources Research, 1994, 30(12): 3243-3252. [10] MCKAY L D, CHERRY J A, BALES R C, et al. A field example of bacteriophage as tracers of fracture flow[J]. Environmental Science & Technology, 1993, 27(6): 1075-1079. [11] FRAGKOU E, ANTONIOU E, DALIAKOPOULOS I, et al. In situ aerobic bioremediation of sediments polluted with petroleum hydrocarbons: a critical review[J]. Journal of Marine Science and Engineering, 2021, 9(9): 1003. [12] 刚洪泽, 刘金峰, 牟伯中. 多孔介质中微生物生长行为和传输过程的数学模型研究进展[J]. 化学与生物工程, 2009, 26(4): 1-6. doi: 10.3969/j.issn.1672-5425.2009.04.001 [13] CHEN H, FENG S J, ZHENG Q T, et al. Enhanced delivery of amendments in fractured clay sites based on multi-point injection: An analytical study[J]. Chemosphere, 2022, 297: 134086. [14] 毕永强, 俞理, 修建龙, 等. 采油微生物在多孔介质中的迁移滞留机制[J]. 石油学报, 2017, 38(1): 91. doi: 10.7623/syxb201701010 [15] GREER K D, MOLSON J W, BARKER J F, et al. High-pressure injection of dissolved oxygen for hydrocarbon remediation in a fractured dolostone aquifer[J]. Journal of Contaminant Hydrology, 2010, 118(1-2): 13-26. [16] CORAPCIOGLU M Y, HARIDAS A. Microbial transport in soils and groundwater: A numerical model[J]. Advances in Water Resources, 1985, 8(4): 188-200. [17] BAI H, COCHET N, DRELICH A, et al. Comparison of transport between two bacteria in saturated porous media with distinct pore size distribution[J]. RSC Advances, 2016, 6(18): 14602-14614. [18] HENDRY M J, LAWRENCE J R, MALOSZEWSKI P. Effects of velocity on the transport of two bacteria through saturated sand[J]. Groundwater, 1999, 37(1): 103-112. [19] WANG S, CORAPCIOGLU M Y. Simulation of bioaugmentation involving exogenous bacteria injection[J]. Water Resources Research, 2002, 38(12): 29-1-29-14. [20] PHANIKUMAR M S, HYNDMAN D W, WIGGERT D C, et al. Simulation of microbial transport and carbon tetrachloride biodegradation in intermittently-fed aquifer columns[J]. Water Resources Research, 2002, 38(4): 4-1-4-13. [21] SCHIJVEN J F, HASSANIZADEH S M. Removal of viruses by soil passage: Overview of modeling, processes, and parameters[J]. Critical Reviews in Environmental Science and Technology, 2000, 30(1): 49-127. -
 点击查看大图
点击查看大图

计量
- 文章访问数: 414
- HTML全文浏览数: 414
- PDF下载数: 42
- 施引文献: 0




























