



-
在长时间、远距离、多人驻足的外星球基地航天任务中,能够携带的资源十分有限,如何实现封闭环境内有限资源的再生和利用,是制约长期地外生存等载人航天任务成败的关键,建立受控生态生保系统 (controlled ecological life support system, CELSS) 是解决该问题的根本途径[1-2]。CELSS的作用是为人类在孤立空间中提供生存环境,其主要目的是再生大气、循环用水、提供食物和处理废物,实现密闭条件下的物质循环[3-4]。好氧堆肥技术具有反应条件温和、有害气体排放量小以及所得堆肥产物可直接利用于植物栽培等诸多优点,是未来处理CELSS内生物质固废的一种具有潜在优势的处理技术。然而,地球上常规使用的中大型反应器和垛式堆肥系统由于占地面积大,污染气体难以集中处理[5],并不适用于狭小的CELSS空间环境。因此,探究掌握微小型密闭好氧堆肥装置的运行条件对实现CELSS固废资源化十分重要。
影响好氧堆肥过程的因素有很多,如环境温度、通风条件、物料种类和反应器类型等。其中,通风条件是影响好氧堆肥的关键因子。聂二旗等[6]研究了不同通风量 (0.1、0.2、0.3 m3·(m3·min)−1) 对鸡粪堆肥效果的影响,结果表明通风量为0.2 m3·(m3·min)−1时氮素的损失最小,种子发芽指数最高。PENG等[7]采用了60 L密闭反应器研究了通风速率对厨余垃圾好氧堆肥的影响,发现通风量为0.2~0.3 L·kg−1·min−1,堆体保温效果最好,污染气体排放量较少,腐熟度最高。ZHANG等[8]利用100 L堆肥反应器对厨余垃圾进行好氧堆肥,结果表明,0.1 L·kg−1·min−1的通风速率有利于食物垃圾堆肥过程中的腐殖化和污染气体减量。然而,目前针对小于10 L容积的微小型反应器好氧堆肥相关报道不多,而且由于微小型堆肥自发产热少,载人航天CELSS中产生的生物质固废种类也有所不同,其部分堆肥参数例如通风条件等并不能直接采用以上研究结果,必须针对CELSS特殊物料和反应器类型进行调整。
基于此,项目以CELSS内主要种植作物小麦的秸秆和模拟粪便的混合物为处理对象,利用自研的微小型密闭好氧堆肥装置,重点考察了不同通风速率对堆肥化过程中O2消耗、主要气体 (CO2、CH4、N2O、NH3等) 的产生和物料特性的变化等方面的影响,以掌握适于未来星球基地长期载人航天任务应用的微小型密闭好氧堆肥装置的通风工艺,实现CELSS内生物质固废资源化处理。
-
本实验所用材料包括小麦秸秆和模拟粪便,基本性质如表1所示。模拟粪便根据美国宇航局 (NASA) 采用的配方[9]所调制,配方为 (质量比) 37.5%的麦麸粉、37.5%的酵母粉、20%的花生油、4%的氯化钾 (KCl,分析纯) 和1%的磷酸二氢钙 (Ca(H2PO4)2,分析纯) 。
-
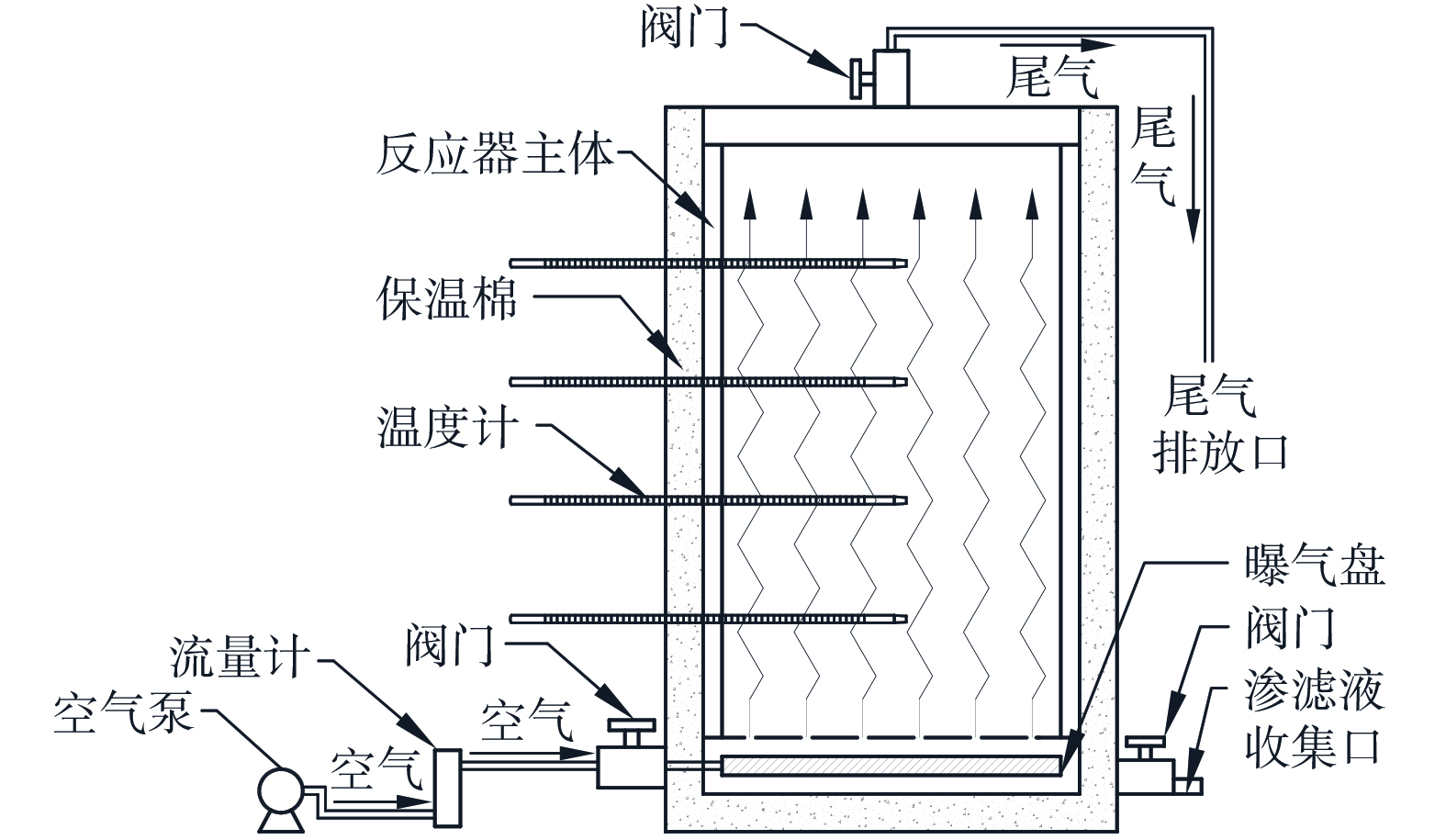
本实验共设置4个处理组,将其编号为V1、V2、V3和V4,分别代表0.025、0.050、0.075和0.100 m3·(m3·min)−14种通风速率。堆肥装置有效容积为8.0 L,其具体结构如图1所示。将小麦秸秆和模拟粪便按照初始C/N为25∶1进行充分混合,加入纯净水将含水率调至65%,实际含水率测得为66.5%,采用QM菌剂 (购自湖北启明生物工程有限公司,主要成分为芽孢杆菌、酵母菌、丝状真菌等多种有益微生物和胞外酶) 接种,剂量占总重的0.5%。分别向4个处理组堆肥装置内装入1.5 kg混合物料,按照0.025、0.050、0.075和0.100 m3·(m3·min)−1的通风速率,依次通入0.20、0.40、0.60和0.80 L·min−1的空气。
本实验共进行15 d,每天10:00、15:00和20:00测量堆体温度、尾气中O2和CO2体积分数,其中第2、4、7、10和15 d上午10:00用铝箔采气袋在尾气排放口采集3 L气体,并从堆体均匀取样进行含水率的测定,另外再取30 g鲜样保存至-80 ℃超低温冰箱。实验第5 d和第10 d对堆体进行人工翻堆。
-
1) 温度。采用水银温度计直接测量。
2) O2和CO2。采用复合型在线气体分析仪 (GT6005,北京国鼎环科科技有限公司) 直接测定。CO2超过6%的部分采用气相色谱仪 (Aglient7890A,美国安捷伦科技有限公司) 测定,载气为高纯氦气 (He,99.999%) ,分离柱型号为TDX-01,检测器为热导检测器TCD。O2消耗速率和CO2产生速率按照式(1)计算。
式中:v为O2消耗速率或CO2产生速率,L·min−1;V为通风速率,L·min−1;φ1为进气中O2或CO2体积分数;φ2为尾气中O2或CO2体积分数。
3) CH4和N2O。采用气相色谱仪 (Aglient7890A,美国安捷伦科技有限公司) 分析测定,载气为高纯氮气 (N2, 99.999%) 。其中CH4采用TRACE TR-Wax GC色谱柱和FID火焰离子化检测器。N2O采用TracePLOT TG-BOND Q+GC色谱柱和ECD电子俘获检测器。
4) 含水率。将样品置于105 ℃烘箱干燥8 h至恒重,减重质量占总质量的比值即为含水率。
5) pH、EC、GI。采用去离子水浸提,鲜样与去离子水按质量比1∶10混合,置于回旋振荡器上,以100 r·min−1的速率振荡120 min,然后过滤,收集过滤液,测试pH、EC和GI。pH采用pH计 (PHS-25,上海仪电科学仪器股份有限公司) 直接测定。EC采用电导率仪 (DDS-11A,上海仪电科学仪器股份有限公司) 直接测定。GI按照式(2)进行计算。
式中:GI为发芽指数;W1为堆肥浸提液培养的种子发芽率;L1为堆肥浸提液培养的种子平均根长,mm;W2为去离子水培养的种子发芽率;L2为去离子水培养的种子平均根长,mm。
6) DOC (可溶性有机碳) 、NH4+-N (铵态氮) 。DOC采用总碳分析仪 (TOC-VCPH,日本SHIMADZU公司) 测定,浸提液经过0.22 μm滤头过滤和rp预处理柱吸附后上离子色谱仪 (Aquion,美国赛默飞世尔科技公司) 测定NH4+。
-
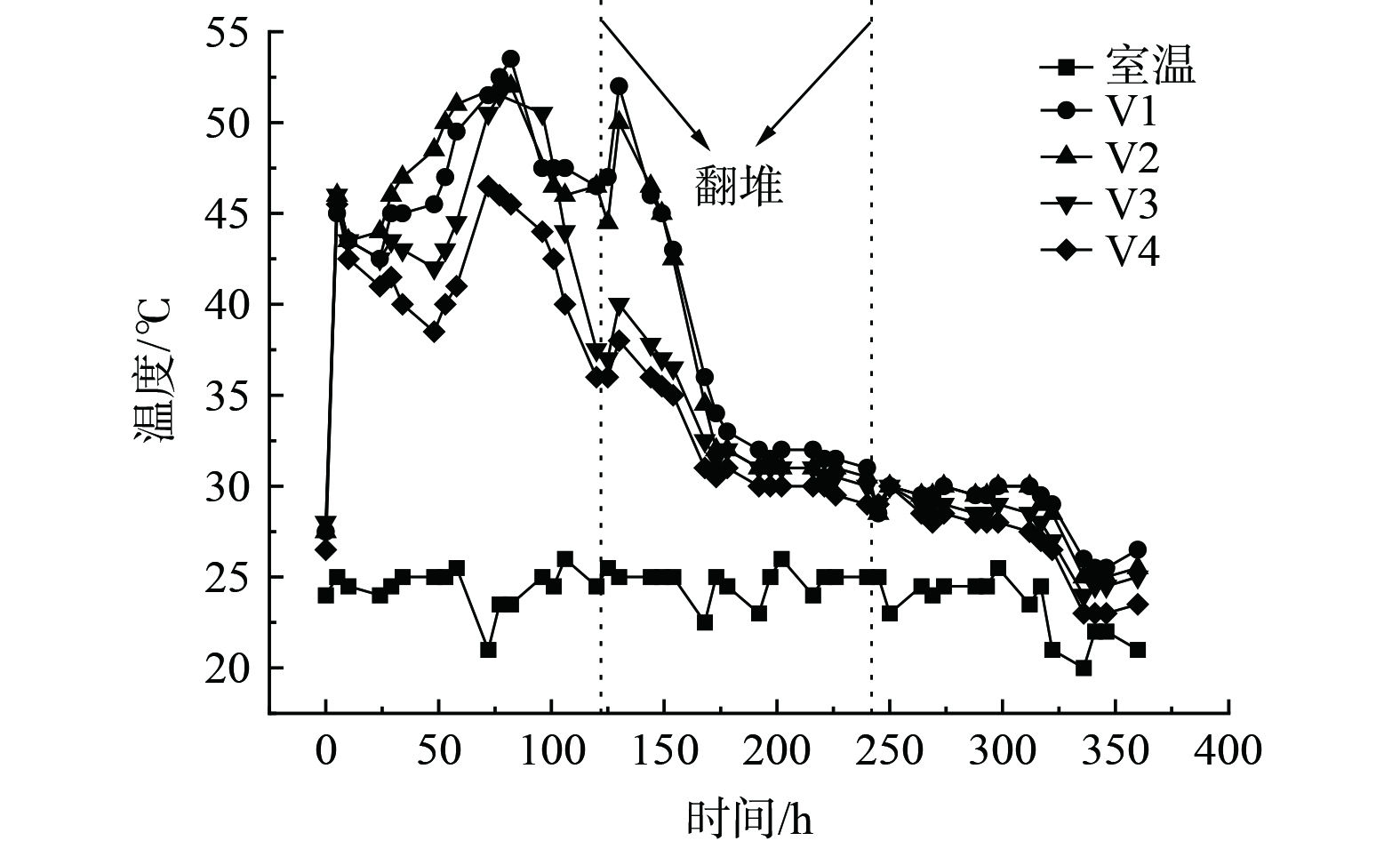
堆体温度的变化是微生物代谢产热的结果,同时也会反过来影响微生物的代谢活性[10]。堆体温度的变化趋势如图2所示。4个处理组的温度变化趋势基本一致,0~5 h升温迅速,在第5 h达到45 ℃以上高温后迅速下降,此阶段主要是由于酵母菌利用堆体中容易降解的有机物实现快速升温,然后被40 ℃以上高温灭活造成的[11]。随后4个堆体开始第2次升温,V1、V2、V3和V4处理组分别在第29、29、72和72 h再次进入高温 (大于45 ℃) ,高温期分别维持了125、125、43和24 h。温度在达到最高后便开始下降,观察堆体的形状和颜色,发现堆体内的菌丝生长十分茂盛,菌丝与物料结合十分紧密,水分分布不均,形成局部厌氧导致了温度的下降。在第5 d (第120 h) 时进行翻堆后,4个处理组温度均有不同程度上升,主要是因为翻堆改变了堆体的结构,通气效果得到了改善,微生物代谢频率得到加强[12]。在实验进行的第10 d (第240 h) 进行第2次翻堆,此次翻堆并未对温度造成较大影响,并逐渐趋于室温,主要是因为此时堆体内易降解物质基本降解完全,剩余物料难以被微生物进一步利用。
在第0~72 h内,堆体温度V2>V1>V3>V4,说明在堆肥升温期和高温前中期时,V1处理组供氧不足,抑制了有机物的降解过程,而V3和V4处理组通风速率又过大,导致热量散失过多,因此只有V2既能保证堆肥氧气的供应,又能减少热量的散失。在第72~360 h,堆体温度呈现V1>V2>V3>V4,说明在高温后期和降温腐熟期,V1处理组的通风速率基本满足了氧气供应,并且由于其通风速率小,热量散失相对较慢,因此温度比其它3组都要高。
实验过程中,各处理组高温期维持时间均相对较短,降温过程相对较快,主要是因为微小型堆体本身自发产热少、热量容易散失等。为达到良好的堆肥效果,使堆肥产品更好地运用到CELSS植物栽培中,可以从以下2方面加以改善:一是在堆肥高温后期和降温期可适当降低通风速率减少热量损失;二是提高实验的环境温度,减少堆体和环境的温度差。
-
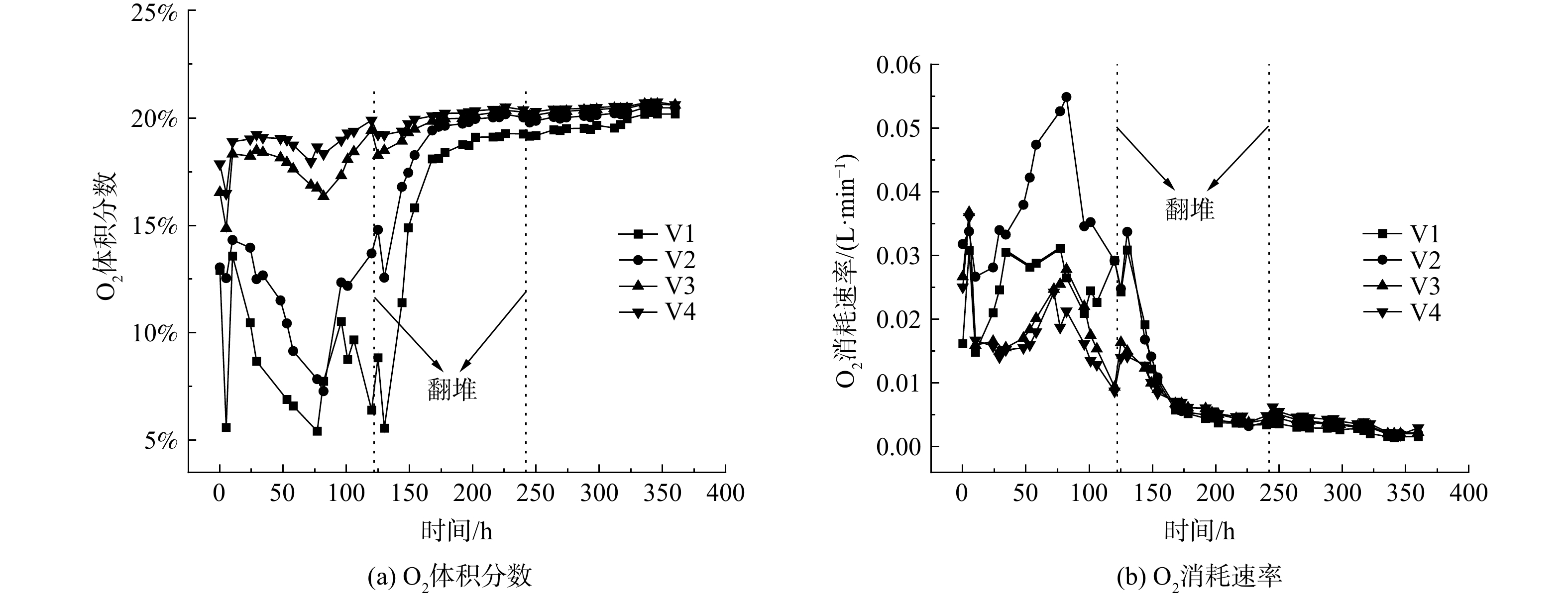
O2是影响堆肥过程的关键因素,充足的O2供应可以更好地满足微生物活性需求,加快堆体温度的上升,减少恶臭气体的排放,并缩短堆肥的周期[13]。堆肥尾气中的O2体积分数及消耗速率如图3所示,结合堆体温度变化趋势不难发现,O2体积分数变化大致同温度变化趋势相反,呈现显著的负相关关系 (r=−0.868,P<0.001) ,这与陈是吏等[14]的研究结果一致。第0~5 h升温期内O2消耗迅速,此阶段主要是酵母菌呼吸作用所致。在高温下,酵母菌活性受到抑制甚至死亡,因此4个处理组O2体积分数在第5 h后迅速回升。上升至第10 h后,4个处理组O2体积分数开始下降,分别于第77、82、82和72 h降至最低,为5.42%、7.28%、16.36%和17.97%。高温期后期,4个处理组O2体积分数均开始上升,这主要是由于高温前期有机质含量和微生物活性都比高温后期更加旺盛[15]。第5 d (第120 h) 翻堆后,O2体积分数开始下降,主要由于翻堆打破了局部厌氧状态,使微生物代谢活动增强。第10 d (第240 h) 再次对堆体进行翻堆,但O2体积分数变化情况并不明显,说明堆体内易降解物质基本降解完全。
从O2消耗速率图 (图3(b)) 可以看出,4个处理组O2消耗速率规律大致相同。但在高温期,V2处理组O2消耗速率显然比其它3组要大,其主要原因是V3和V4通风速率过高,带走了更多的水分和热量,而V1通风速率过小导致氧气供应不足,影响了微生物的代谢活动。降温期由于可利用的有机物变少,微生物代谢活动减弱,O2需求开始减少[16],导致4个处理组O2消耗速率的差异并不显著。对O2消耗速率图 (图3(b)) 4条曲线进行积分,得到4个处理组O2消耗总量分别为273.68、370.91、221.93和208.58 L (标准大气压) ,可见V2处理组氧气消耗总量远大于其它3组,好氧反应相对更加完全和彻底。
-
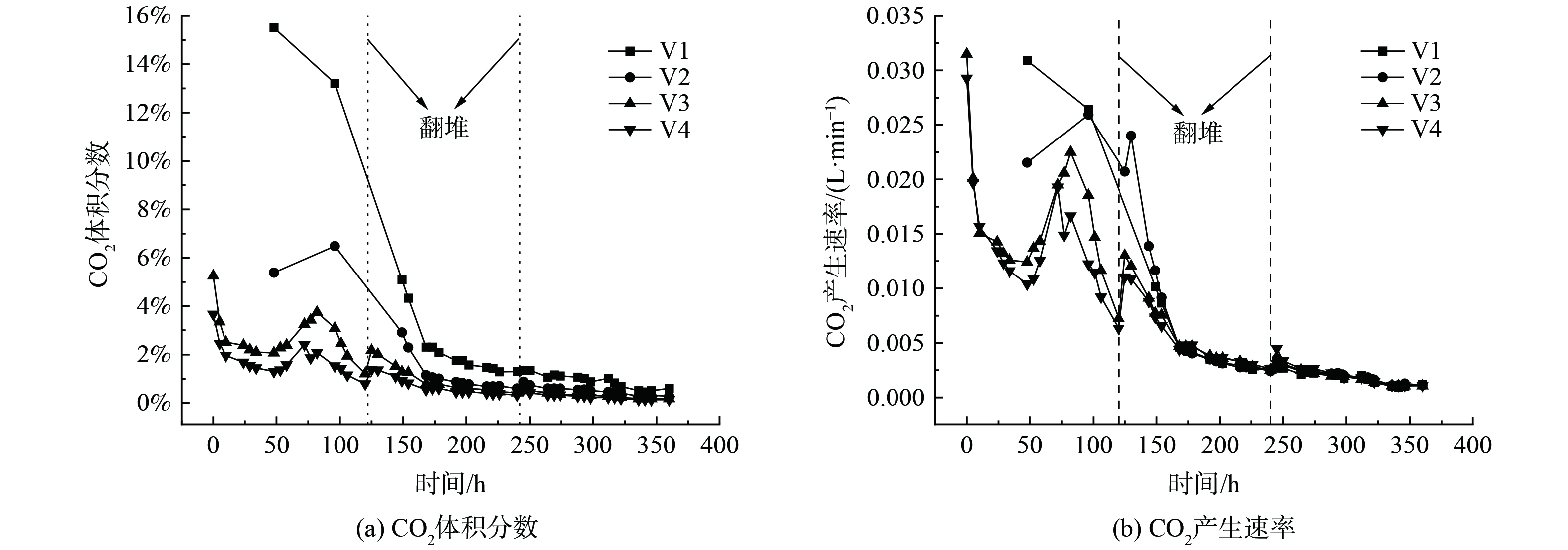
好氧堆肥中的CO2主要是微生物通过有氧呼吸分解有机物所产生,CO2产生速率可反映堆肥过程中微生物的活性和有机质矿化率[17]。堆体尾气中CO2体积分数和产生速率如图4所示,由于本实验采用的复合型气体分析仪的CO2量程仅为6%,因此对V1和V2处理组的第48 h和96 h气体样品进行气相色谱测定。CO2变化趋势大致同温度变化趋势相同,与O2变化趋势相反。V1处理组第48 h和96 h的CO2体积分数分别为15.51%和13.22%,产生速率分别为0.030 9和0.026 4 L·min−1。V2处理组第48 h和96 h的CO2体积分数分别为5.38%和6.48%,产生速率分别为0.021 5和0.025 9 L·min−1。V3和V4处理组在刚开始堆肥时CO2体积分数和产生速率就已经达到了峰值,分别为5.25%、0.031 5 L·min−1和3.66%、0.029 3 L·min−1。随后由于高温抑制了酵母菌的活性,导致CO2体积分数和产生速率大幅度下降。在第48 h后V3和V4处理组CO2体积分数增幅较大,此时的温度也在增加,主要是因为前期高温筛选出了QM菌剂中的优势菌种,利用了堆体内有机物进行繁殖扩增。在第48~120 h之间,微生物大量繁殖与物料结合紧密,通气受阻,CO2体积分数出现了快速的下降,直至第5 d (第120 h) 翻堆打破局部厌氧状态得到短暂回升。由于V1和V2处理组CO2变化趋势缺少部分数据,需要结合堆体温度和氧气变化趋势进行综合分析,但不难发现CO2产生总量V2>V1>V3>V4。
-
CH4是由产甲烷菌利用堆体内甲酸、甲醇和甲基胺类等简单有机物在厌氧条件下产生的温室气体[14],CH4的变化趋势可反映堆体的厌氧情况。堆肥尾气中CH4体积分数和产率如图5所示,不同处理组尾气中CH4体积分数和产生速率存在明显差异,但总体都呈现了先上升后下降的趋势,且主要产生于堆肥初期,这与之前学者的研究结果相似[18],主要是因为有机物降解造成大量氧气消耗,加剧了堆肥基质内的厌氧情况。后期由于易降解有机物的减少,对O2需求量减少,厌氧环境得到了改善,CH4体积分数开始下降。4个处理组CH4体积分数分别在第2、7、4和2 d达到最大值,为68.1、25.3、19.9和19.4 μL·L−1。其中以V1处理组尾气中CH4体积分数最高,其它3个处理组的差距并不显著,主要还是因为V1处理组通风速率太小,高温期O2供氧不足,局部厌氧更加显著。对CH4产生速率曲线进行积分,可以得到V1、V2、V3和V4处理组CH4排放总量分别为279.73、241.30、279.36和296.50 mg·kg−1,过大的通风速率并没有有效减少CH4的排放总量,这主要是受到了通风逸出的影响,较大的通风速率造成了更大的气压,更容易将污染气体及时从局部厌氧区扩散出来,而低通风速率处理组堆体含水率较大,导致内部孔隙度较低,且低通风速率气压过小,导致厌氧区产生的CH4难以逸散[7, 19]。
-
N2O是温室效应极强的一种气体,其产生机制十分复杂,目前普遍认为N2O的产生主要有铵态氮的硝化和硝态氮的反硝化2个途径[20-21]。堆肥尾气中N2O体积分数和产率如图6所示,各处理组的N2O主要排放在堆肥初期,这是由于硝态氮在堆肥初期高含水率、低孔隙度和易降解物质快速降解造成的厌氧环境中不完全反硝化所致[22]。但在第4 d高温期出现了快速下降,主要是因为高温和高氨氮环境不适宜硝化细菌的生长,抑制了反硝化所需的NOx−-N来源[23]。4个处理组分别于第2、2、2和4 d达到N2O最高浓度,为17.22、12.08、5.17和3.93 μL·L−1,此时产N2O率也达到了最大,分别为19.39、27.21、17.46和17.69 mg·kg−1·d−1。之所以V2处理组在高温期N2O产率最大,主要是因为该处理组产生了比其它组更多的NH4+-N,大量的NH4+-N不完全硝化导致了更多的N2O。而在降温期和腐熟期,4个处理组产N2O率随着通风速率的增大而增大,主要原因可能是充足的氧气产生了较多的NOx−-N,而NOx−-N是反硝化过程最重要的底物。对N2O产生速率曲线进行积分,得到4个处理组N2O排放总量分别为94.67、157.11、150.10和169.16 mg·kg−1。低通风速率V1处理组N2O的排放量显著小于其他3组,这主要是因为低通风速率处理组堆体含水率较高,堆体孔隙度相对较低,而较低的通风速率由于压力较小,难以将堆体内产生的N2O吹脱出来,高通风速率虽然O2含量很高,但并不能完全均匀地输送到各处,局部厌氧情况仍然存在,且由于高通风速率空气流速较快,N2O从堆体中逃逸相对容易[7, 24]。
-
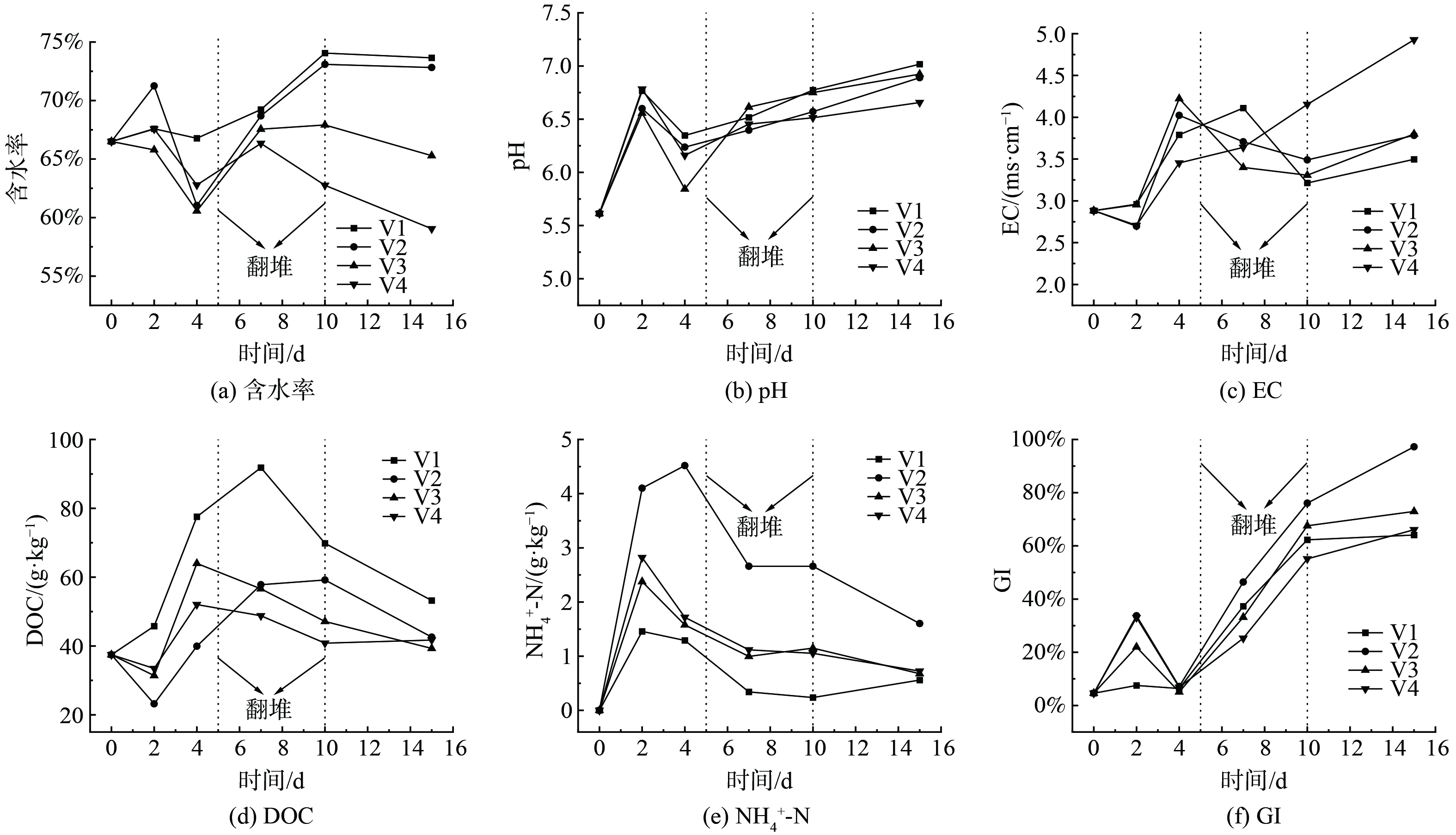
堆体含水率是影响堆肥质量重要指标之一,过高的含水率会降低堆体含氧量,造成厌氧,含水率过低则会影响微生物的繁殖[25]。堆体含水率变化趋势如图7(a)所示,4个堆体含水率在0-2 d有不同程度的上升,主要原因是易降解有机物分解产生了大量水分。但在第2~4 d,4个堆体的含水率均出现了不同程度的下降,因为此时的温度过高,水分蒸发速度过快,并且由于此时堆体存在局部厌氧致使微生物活性降低,难以有效降解有机物,导致分解产生的水分变少。但经过第1次翻堆后,打破了局部厌氧状态,微生物继续分解有机物,因此含水率又出现了短暂的回升。在第2次翻堆后,4个堆体含水率均有不同程度的下降,原因是容易降解的物质变少,微生物的代谢活动开始减慢,产生的水分少于通风带走的水分。与初始含水率相比,V4处理组和V3处理组损失的水分较多,但V1和V2含水率不减反增,主要是由于其通风速率小,带走的水分有限,并且由于本实验采用密闭反应器式堆肥,水蒸气易凝结在反应器上盖回滴至堆体。由此可见,V1和V2处理组通风速率可以有效保持堆体水分,一定程度上可以减少后续CELSS植物栽培所需水分的摄入。
堆体pH反映了微生物生存环境的酸碱情况,过高和过低的pH均会影响着微生物的活性。从pH变化趋势图 (图7(b)) 观察到,4个堆体的pH变化均出现升高、降低和再升高的趋势。第0~2 d堆体pH的升高,主要原因是在升温期大量的含氮物质降解,生成的氨类物质所致[26]。第2~4 d堆体pH出现了短暂的下降,主要是因为堆体内生长了大量微生物,与物料结合紧密,造成的局部厌氧产生了有机酸和CO2等酸性物质[27]。在第1次翻堆后,堆体内厌氧状态被破坏,此时堆体又源源不断产生氨气被留在堆体内部,因此pH逐渐开始上升。总体看来,4个处理组的pH差异并不显著,在堆肥结束时均处于6.5~7.0中性水平。
电导率EC是反映堆体可溶性盐含量的重要指标,过高的电导率会对微生物繁殖和植物的栽培造成毒害作用。从电导率变化趋势 (图7(c)) 可以看出,除了V4处理组,其它3组电导率均大体呈现了先升高后降低的趋势。堆肥升温期和高温期电导率的上升,主要原因是此时微生物代谢活动十分剧烈,大分子有机物的分解,产生了大量的矿物盐,使得电导率增加。而随后电导率出现了略微的降低,主要是腐殖化过程中微生物将盐分和有机酸转化为腐殖质引起的[28]。实验结束后,4个处理组的电导率分别为3.50、3.78、3.80和4.93 ms·cm−1,除V4处理组外,其它处理组均满足GARCíA等[29]对堆体EC<4.0 ms·cm−1的要求。
溶解性有机碳DOC是微生物分解大分子有机物的产物,同时又是微生物赖以生存的碳源和能源[30]。从堆体DOC变化趋势 (图7(d)) 观察到,除V1处理组是先升高后降低的趋势,其它3个处理组DOC均呈现先降低再升高后降低的趋势,主要得益于堆肥初期微生物利用了堆体自身含有大量的中小分子水溶性有机碳进行生命繁殖[31],而V1处理组由于O2供应量少,利用的溶解性有机碳较少,并且未对大分子有机物降解生产的DOC进行及时矿化,造成了DOC的积累。堆肥第2 d后,各组DOC含量增幅较大,主要是此时O2降至最低,限制了DOC的进一步分解。在堆肥中后期,堆体只剩下了难降解的有机物,剩下的DOC一部分被微生物微弱的代谢活动分解掉,另一部分被微生物重新合成芳构程度更大的大分子腐殖质,导致了DOC的不断减少[32]。在堆肥结束时,4个处理组的DOC都降至较低水平,分别为53.2、42.58、39.30和41.73 g·kg−1。在整个堆肥过程中,V1处理组DOC含量一直为4个处理组最高,说明V1处理组由于供氧不足,限制了其堆体内有机物的矿化。
铵态氮比较容易被植物所吸收并利用,属于速效性氮素,是好氧堆肥堆体内氮素主要存在形式之一。从堆体内NH4+-N变化趋势 (图7(e)) 观察到,4个处理组的NH4+-N均呈现先升高后降低的趋势。堆肥初期NH4+-N的增加,主要是大量的有机氮受到微生物氨化作用所导致的[7],并且由于本实验采用密闭反应器式堆肥的方法,大量水分凝结在反应器上盖并回滴于堆体表面,造成堆体表面含水率很高,大量氨气被冷凝水截留在堆体内。在降温期,4个处理组堆体内铵态氮均有不同程度降低,大量NH4+-N被硝化细菌反应生成NOx−-N,加之由于微生物活动减弱,大部分容易降解的有机物减少,氨化作用减弱,并且随着持续的高温和pH的升高,部分氮素以氨气的形式损失[33]。对比4个处理组的NH4+-N浓度,V2处理组的NH4+-N浓度远高于其它3个处理组,而V1处理组一直处于最低水平。4个处理组分别在第2、4、2和2 d达到了NH4+-N浓度最大值1.46、4.52、2.38和2.82 g·kg−1,以V2处理组浓度最高,主要原因是低通风速率氧气的供应不足,有机氮矿化受到了抑制,从而导致堆体内的NH4+-N浓度较低,而高通风速率会造成堆体内部更大的气压和更充足的氧气,促进了NH4+-N以NH3的形式逸散[34]和向NOx−-N的转化。总而言之,V2处理组由于提供了充足的O2和合适的内部气压,其NH4+-N浓度最高,保氮效果最佳。
种子发芽指数 (GI) 可以表征堆肥的腐熟程度,反映堆肥产物对植物的毒害作用,是评价堆肥产品质量最终且最具有说服力的指标[35]。从GI变化趋势图 (图7(f)) 观察到,4个处理组的GI均呈现先升高、后降低和再升高的变化趋势。各处理组在第4 d的GI都有不同程度的下降,主要是因为此时堆体内产生了有机酸、醛和氨气,对种子发芽造成了抑制作用[36]。在第5 d翻堆后,4个处理组的GI一路直升,直至第15 d堆肥结束时,达到最大值,分别为64.09%、97.26%、72.95%和66.07%,其中,只有V2 和V3处理组达到了生物质腐植酸有机肥料行业标准[37]中关于GI≥70%的要求。
-
1) 微小型堆体自发产热较少,热量容易散失,导致高温期相对较短,可在高温中后期适当降低通风速率,或提高环境温度,以减少堆体的热量损失。
2) 4个处理组尾气中CH4和N2O体积分数差异较大,CH4和N2O体积分数随着通风速率的增大而减小,但累积排放量差异不明显,且均处于较低水平,更高的通风速率并没有进一步减少CH4和N2O累积排放量。
3) 0.050 m3·(m3·min)−1的通风速率既能确保该微小型密闭好氧堆肥过程中微生物对O2的需求,又能减少堆体热量的损失,产生的污染气体量也相对较少,所得的堆肥产品质量最佳,可用于后续CELSS内的植物栽培。
通风对微小型密闭好氧堆肥过程的影响
Effects of ventilation on miniature and closed aerobic composting process
-
摘要: 为探究适于未来星球基地长期载人航天任务应用的微小型密闭好氧堆肥装置的运行条件,以小麦秸秆和模拟粪便的混合物为实验材料,开展了4种通风速率对堆肥过程的影响研究。结果表明,0.025、0.050、0.075和0.100 m3·(m3·min)−1处理组高温期分别维持了125、125、43和24 h,0.050 m3·(m3·min)−1的O2消耗速率和CO2产率为4个处理组最大。CH4和N2O主要产生于堆肥初期,累积排放量以0.100 m3·(m3·min)−1处理组的最高,分别为296.50和169.16 mg·kg−1。从堆体氨氮指标测试结果来看,0.050 m3·(m3·min)−1处理组含量最高,其保氮效果最好。4个处理组的GI分别为64.09%、97.26%、72.95%和66.07%。综合各种指标分析认为,适于未来星球基地任务应用的微小型密闭好氧堆肥装置通风速率可以设置为0.050 m3·(m3·min)−1,该通风速率既能确保堆肥过程中微生物对氧气的需求,又能减少热量的损失,产生的污染气体量相对较少,所得的堆肥产品质量最佳。Abstract: In this study, the operating conditions of miniature closed aerobic composting device suitable for long-term manned space mission in the future planet base were explored. The effects of four ventilation rates on composting process were investigated by taking the mixture of wheat straw and simulated human excrement as the experimental materials. The results showed that the high temperature periods of 0.025, 0.050, 0.075 and 0.100 m3·(m3·min)−1 were maintained for 125, 125, 43 and 24 hours, respectively. CH4 and N2O were mainly produced in the initial stage of composting, and the cumulative emissions of 0.100 m3·(m3·min)−1 treatment group were the highest, which were 296.50 and 169.16 mg·kg−1 respectively. The test results of ammonium nitrogen of the pile revealed that, the treatment group with 0.050 m3·(m3·min)−1 had the highest content and the best effect of nitrogen retention. The GI of the four groups were 64.09%, 97.26%, 72.95% and 66.07% respectively. Based on the comprehensive analysis of various indicators, it was concluded that the ventilation rate of the miniature closed aerobic composting device suitable for the future mission of Star Base should be set to 0.050 m3·(m3·min)−1. That ventilation rate can not only ensure the oxygen demand of microorganisms in the composting process, but also reduce the heat loss, resulting less pollution gas, and the best-quality composts.
-
Key words:
- manned space flight /
- aerobic composting /
- ventilation rate /
- wheat straw
-
随着我国经济和社会的快速发展,城镇化率和生活水平的提高,城市生活垃圾的产生量已突破2.5亿吨/年,其大量排放对自然环境构成了严重的威胁。生活垃圾焚烧处理法因其处置高效、减量显著、节省用地和消毒杀菌彻底等优点在国内外得到广泛应用[1]。
生活垃圾焚烧后将在烟气净化系统中产生飞灰,是重金属和二噁英重要的“汇”。飞灰由于富集大量有毒有害物质,被国家列为危险废物(类别HW18)。飞灰一般为浅灰色的粉末,结构比较复杂,大多以多晶聚合体结构和无定型态形式存在。不同来源的飞灰性质不同,颗粒大小也不均一,通常粒径<100 µm,且表面粗糙,具有较大的比表面积和较高的孔隙率。飞灰的化学成分包括钙、钾、钠、铁、硅、铝、硫、氯和氧等元素,主要化合物为氧化钙、氧化钾、三氧化硫、二氧化硅、氧化铝和氧化铁等。此外,飞灰含有高浓度的有害重金属,如汞、铅、镉、铜、铬和锌等,这些重金属主要以气溶胶小颗粒和富集于飞灰颗粒表面的形式存在;同时在飞灰中还含有少量的二噁英、呋喃等持久性有机污染物,因此飞灰具有很强的危害性,并且在环境中持续时间很长。在焚烧过程中多数重金属能够改变形态,更易发生迁移和转化,并且重金属由于不能被微生物分解,能在生物体内产生累积效应,因此这些污染物质若处置不当,则能使水体和土壤受到污染,进而危害到动植物,甚至通过食物链进入人体,危及人类的生命健康。
目前飞灰的主要处理技术包括安全填埋、分离萃取、固化稳定化和热处理等[2],其中固化稳定化技术应用最为广泛。固化是将飞灰、固化剂混合,在一定条件下反应生成固化体,从而达到固化重金属的目的;稳定化是通过物理化学反应使飞灰转化为毒性低、难迁移和不易溶解的稳定物质。常见的飞灰固化稳定化技术有水泥固化、药剂稳定化、水热稳定化和地聚合物固化等[3]。其中,地聚合物固化处置相较于其它处置方法,具有材料来源广,工艺简易可行,能源消耗低、低碳环保和初期投资及管理运营投资少等优势[4]。
1. 飞灰地聚合物固化稳定化技术
地聚合物(Geopolymer)是铝、硅质材料在高碱环境下发生反应制得的一种三维网状无机聚合物,由法国DAVIDOVITS JOSEPH实验室在20世纪70年代首次开创研究[5]。飞灰地聚合物固化稳定化是将飞灰与地聚合物基质,如偏高岭土、高炉矿渣、粉煤灰和固体废物等按一定顺序搅拌混合,加入碱激发剂后,混合物发生地聚合反应形成固化体,在预设的湿热条件下养护成型,从而将飞灰稳定在固化体内,其含有的重金属、二噁英等污染物经浸出测试达到国家相关标准,可根据需要进行后续处置和应用。
通过地聚合反应形成的固化体,具有良好的化学稳定性、力学特性、低缩性、材料稳定性和环境友好性,同时也具有资源化处置的潜力[6],因其固化体具有早强快硬、结构稳定和长期耐久等特点,成为固化处置飞灰的有效方法,从低碳环保、固化效果来看是最为有利的飞灰资源化利用方式之一。
在地聚合物固化体形成过程中,用于生成地聚合物的基质组分、碱激发剂、改性物质、搅混养护等材料和方法将对飞灰地聚合物固化体结构和性能产生直接的影响,其中基质材料配比、碱激发剂比例、添加物质类别,以及搅混顺序、养护条件等对飞灰的固化稳定化效果将起到关键作用。
2. 飞灰地聚合物固化反应材料与条件
2.1 实验方法
碱激发地聚合固化技术经过多年的研究探索,已经形成了一套成熟的实验方法。实验步骤一般为:按照飞灰固化需求,将一定比例的固化基质和飞灰干混,根据改性需要可以加入一定量的添加剂,然后加入制备好的碱激发剂和蒸馏水,混合搅拌制备具有一定流动性和粘性的浆体。将浆状物注入模具内,用振荡器振实,排出气泡,刮平覆膜,放入恒温恒湿的养护箱中在一定条件下养护,脱模后根据需要可继续养护一段时间。养护达到预设的时间后,取出模块,进行相应的指标检验、重金属浸出测试或其它性能测试。
2.2 基质组分与配比
国内外许多学者研究了使用粉煤灰、高岭土、偏高岭土和高炉矿渣等材料作为地聚合物固化飞灰的基质,并讨论了基质组分的配比对固化体性能的影响。
郭晓潞等[7]针对城市垃圾焚烧飞灰,应用高钙粉煤灰制备地聚合物进行固化,对高钙粉煤灰-城市垃圾焚烧飞灰复合地聚合物中飞灰的加入量开展了实验研究,结果表明高钙粉煤灰基地聚合物中飞灰含量达5%~15%时,其重金属浸出浓度均远低于标准最高限值,复合地聚合物中飞灰的最佳加入量宜控制在10%以内。谢吉星等[8]研究了将飞灰和高岭土作为主要基质,制备形成了飞灰地聚合物,当飞灰的加入量在70%时,其28 d抗压强度可达19.36 MPa。重金属浸出实验表明,养护28 d时重金属基本无溶出。袁正璞等[9]以偏高岭土和飞灰为原料制备了地聚合物,研究了大比例加入飞灰时,不同原料配比对地聚合物性能的影响,合成的地聚合物对重金属有明显固化效果。随着飞灰加入量从85%逐步调整到97%(偏高岭土比例逐渐减少),地聚合物的抗压强度整体呈依次降低的趋势。
地聚合物钠、硅、铝配比对反应生成的固化体性能也会产生重要的影响。王开等[10]探讨分析了硅、铝物质的质量最佳配比,以及硅、铝配比对飞灰中重金属固化效果和固化体力学性能的影响规律,提出了一种固化稳定化飞灰的新方法。当硅、铝物质的质量之比在2.5时,重金属的浸出浓度与固化体的抗压强度均趋于稳定,具有较好的固化效果。施惠生等[11]研究优化了PSS型地聚合物的配合比,发现当二氧化硅与三氧化二铝配比为4.0、氧化纳与三氧化二铝配比为1.1,水与氧化纳配比为6.8时,地聚合物固化效果最佳。金漫彤等[12]以抗压强度和重金属浸出质量浓度为测试指标,实验研究了地聚合物固化垃圾焚烧飞灰的工艺,探讨了地聚合物固化飞灰的合成配方。结果表明,氧化物中氧化纳与二氧化硅的质量配比为0.3、二氧化硅与三氧化二铝质量配比为4.7和加入飞灰占比为40%时,地聚合物固化体养护7 d后,抗压强度能达到31.25 MPa,固化体中铅、锌、铜和铬的浸出质量浓度分别达到0.1838、0.0231、0.0081和0.3679 mg/L。
2.3 碱激发剂
碱激发剂在地聚合物使用中应具备3个基本条件[13]:(1)可以不受空气影响,能够稳定存在;(2)可以参与反应,形成稳定的水化产物;(3)可以促使水化产物聚合,形成凝聚态网状结构。碱激发剂主要组分是苛性碱和碱金属硅酸盐。GALIANO et al[14]选择不同的碱激发剂,如Na2SiO3、K2SiO3和NaOH和KOH等,对飞灰地聚合固化结果进行了比较,研究表明:上述碱性激发剂对地聚合物固化垃圾飞灰中重金属有优良的效果。
多数研究采用氢氧化钠和水玻璃(主要成分为硅酸钠)作为地聚合化学反应的碱激发剂。DIAZ-LOYA et al[15]对燃煤飞灰地聚合固化处理垃圾飞灰开展了研究,加入硅酸钠和氢氧化钠激发剂后,重金属浸出浓度大幅降低,其中砷下降了48.44%,铬下降了97.06%。金漫彤等[12]以高岭土为基质材料对垃圾飞灰进行固化研究,加入水玻璃和氢氧化钠作为碱激发剂,实验发现4种重金属铅、铬、铜和锌的浸出质量百分比的下降比率分别达95%以上,固化体抗压强度最高可以达到44.25 MPa。LANCELLOTTI et al[16]应用高岭土采取地聚合方法固化20%的垃圾飞灰,利用氢氧化钠和水玻璃作为碱激发剂,对飞灰中重金属铬、镉、镍、铜和铅稳定固化后,其固化体浸出浓度均未超过检测标准。
另有一些研究直接使用氢氧化钠作为地聚合化学反应的碱激发剂。YE et al[17]对拜耳赤泥进行地聚合固化处理,应用氢氧化钠作为碱激发剂,经研究表明,浸出液中重金属锌、铅、铜和铬的浸出值均符合填埋标准。HANJITSUWAN et al[18]研究了在高钙垃圾飞灰形成地聚合物时加入碱激发剂氢氧化钠,研究其浓度对激发形成地聚合物的影响,结果表明随着氢氧化钠浓度升高,可以更多溶解垃圾飞灰颗粒,促使地聚合反应快速进行,能够逐步提升固化体抗压强度。
2.4 添加剂
在实际生产中有时为满足不同需求或解决现场应用中出现的各类问题,会在地聚合物固化时掺入不同的添加剂来改变地聚合物性能。金漫彤等[19]在地聚合物固化飞灰时分别掺入水泥、沸石和偏磷酸钠3种添加剂,考察掺入水泥、沸石和偏磷酸钠对飞灰基地聚合物固化体抗压强度和重金属浸出质量浓度的影响。研究表明,水泥可缓解固化体膨胀,沸石可减小固化体的总孔隙率,偏磷酸钠可作为缓凝剂延长浆砂的凝结时间。重金属能被很好地包容在固化体中,表现出良好的浸出安全性。刘道洁[20]以垃圾焚烧飞灰和二次铝灰为基质材料,加入碱激发剂反应生成地聚合物固化体,加入二氧化硅粉末后发现可以促进地聚合反应更好地进行。在二次铝灰中,由于二氧化硅和三氧化二铝摩尔比很小,因此添加适量的二氧化硅粉末有助于更快发生化学反应,取得最好的固化效果,当二氧化硅和三氧化二铝摩尔比为2.5左右时,浸出液中重金属达到浸出要求。
2.5 养护条件
飞灰地聚合物固化体制成后,一般都需要在一定条件下进行养护,以保证固化体内部化学反应充分完成,达到最佳的固化性能。董海丽[21]研究发现,固化体具有早期强度高的特点,固化体养护1 d后,抗压强度就能达到20 MPa以上。随着增加固化体养护天数,其抗压强度在7 d左右上升趋势有所放缓,由此表明固化体的抗压强度随着反应时间的增加而增加,直至趋于稳定。分析其原因,水玻璃对地聚合反应有双重作用,OH−促使原料玻璃态解聚,而含水硅胶又能与水反应,使水化生成物增多,所以早期强度发展快。随着固化时间不断延长,地聚合物中的氧化物摩尔比,如二氧化硅与三氧化二铝比值、氧化纳与二氧化硅比值和氧化纳与三氧化二铝比值均有所下降,并且结构当中反应水含量降低,这可能是因为铝氧四面体和硅氧四面体更多地参与了地聚合反应,使地聚合物抗压强度增加的结果。刘道洁等[22]研究了地聚合物固化体样品的养护条件对重金属浸出浓度的影响,养护箱参数为恒定条件(相对湿度80%,温度30~50 ℃)。当养护温度为50 ℃时,地聚合物中重金属浸出浓度满足浸出标准的要求。
3. 飞灰地聚合物固化稳定化机理
目前对于地聚合物固化稳定化飞灰的机理还没有系统的认识,对重金属的固化稳定化机理也存在分歧[23]。有学者[24-25]初步研究认为,重金属是通过化学键联和物理固封2种方式被固定在地聚合物中。而有些学者则认为化学结合没有在固化机理方面发挥重要的作用,重金属是被微囊密封在富硅铝的无定形结构中[26]。
硅、铝是固化体形成的必备核心元素,有研究[20]发现随着硅、铝摩尔比的增加,固化体内重金属的浸出浓度都明显降低,其原因主要是随着硅、铝摩尔比逐渐升高,地聚合物的聚合度逐渐增大,生成的地聚合物由单硅、铝地聚合物变成双硅、铝地聚合物,再变成三硅、铝地聚合物,更多的重金属离子与地聚合物官能团发生化学键键合,促使地聚合固化体的内部结构结合的更加紧密和稳定。化学药剂具备很多特殊的性能,其掺入可以实现固化体改性。郭晓潞等[27] 引入化学药剂对其制备的含重金属固废地聚合物进行改性,以粉煤灰为主要基质材料,分别加入冶金污泥和城市垃圾焚烧飞灰,探讨了地聚合的反应机理。研究发现加入化学药剂导致重金属的破坏作用被抑制,加速了地聚合反应进程,产物结构更匀质、致密,增强了化学药剂改性后的飞灰地聚合物固化能力。飞灰含量增加致使其它基质含量降低,硅、铝含量也相对较少,影响地聚合物的抗压强度。李春林[28]以偏高岭土为添加剂,研究飞灰含量对地聚合反应的影响。结果发现,随着飞灰含量的增加,固化体的抗压强度先升高后降低,当飞灰含量较高时,偏高岭土含量则相对较低,活性的硅、铝源难以足够释放出来,地聚合反应因此受到了一定的限制。碱激发剂在地聚合反应中的主要作用是使硅、铝质材料网络结构发生解体、缩聚,最终形成地聚合物结构[23]。ZHANG et al[29]研究了碱激发剂对地聚合反应和抗压强度的机理。结果表明,不断加入碱激发剂后,逐渐增多的激发剂使反应体系中活性硅、铝组分,以及地聚合物的硅氧四体含量逐渐增大,进而大大促进了地聚合反应的进行。但当继续增加碱激发剂的比例时,反应体系中过多的钠离子会减少铝氧四面体基团的绝对含量,从而降低产物的抗压强度[30]。养护条件为固化体形成的重要影响因素,特别是温度条件更为关键,温度升高会加快地聚合物化学反应的进程。彭晖等[31]研究了养护温度对固化效果的影响。结果发现,养护温度增加将会减小重金属离子的相对浸出比例,提升地聚合物固化稳定效果。地聚合反应过程中,不断升高养护温度,物质溶解和水解的过程在一定程度上得到加速,更多地聚合反应所需的前驱物逐步生成,从而使地聚合反应的进程加快进行。
普遍认为重金属地聚合物固化的机理有2种:化学结合和吸附作用。有研究[22,32]提出,飞灰基地聚合物与重金属离子可以通过化学键结合,从而实现对重金属更好地吸附和包裹。原因是重金属形态由游离的可交换态、结合态向铁锰氧化物结合态、有机物结合态以及残渣态转变,使固化体内部紧密结合在一起,结构更加稳定。静态吸附试验表明地聚合物由于具有三维笼状立体结构,具备强烈的吸附能力,能较好地吸附重金属,同时可以将重金属牢牢锁在其构建的牢笼状立体结构内,从而难以从固化体中溶出。
4. 飞灰地聚合物性能评价
目前,大多数研究对飞灰地聚合物固化稳定化效果开展了性能评价,实验研究均取得了较好的效果。董海丽[21]以抗酸碱性能、耐硫酸盐性能、抗海水侵蚀性能和重金属浸出浓度为测试指标,制备了偏高岭土基飞灰地聚合固化体,研究发现上述指标测试均表明具有良好的固化效果,结果显示地聚合物固化体耐受酸碱能力强,当碱溶液浓度较低时,表现出较好的稳定性能,当碱溶液浓度过高时会严重破坏地聚合物的结构;地聚合物固化体能较好抵抗硫酸盐侵蚀;固化体放在人工海水中浸泡,其外观基本不受影响;浸出液的pH值对重金属浸出浓度影响不大。文献[33]制备了飞灰基地聚合物固化体,分析了硫酸盐溶液对固化体的侵蚀影响。从重金属浸出质量浓度、抗压强度和质量等方面探讨硫酸盐侵蚀效果,研究表明地聚合物固化体具有良好的耐硫酸盐侵蚀性能,浸泡28 d后,重金属铬、铜、锌、镉、汞和铅的浸出质量浓度仅为122.00、15.16、13.28、0.04、0.32和4.51 mg/L,抗蚀系数均>0.95,抗压强度为41.17 MPa,质量损失率为−1.87%~0.61%之间。
5. 飞灰地聚合物固化体资源化应用
城市飞灰产生量目前仍在快速增长,对飞灰进行资源化处置已成为科研工作者的共同目标。随着人们对地聚合物的深入研究,利用地聚合物固化飞灰并进行资源化利用已成为重要的发展方向。张希等[34]研究认为,采用飞灰制备地聚合材料时利用率较高,成本及能耗较低,对重金属等有害物质固化效果明显。
綦懿等[3]综述了目前国内外固化稳定化安全处置飞灰的主要方式,分析了各个方法的优缺点,总结了飞灰在建筑材料领域中的研究现状,探讨了未来固化稳定化技术手段的关注点。飞灰地聚合物在建材行业将得到广泛应用,水泥混凝土中可以将地聚合物代替水泥,制备新型地聚合物基建筑材料。李春林[28]以城市生活垃圾焚烧飞灰为原材料,通过碱激发法制备出力学性能优异的地聚合物固化体。当固体质量和碱激发剂的比为1. 2,垃圾焚烧飞灰占比60%时,地聚合物具有最佳的抗压强度,可用作建筑材料。
飞灰基地聚合物固化体相对于水泥基固化体在力学性能、抗渗性和耐久性等方面都具有一定的优势,可以在垃圾填埋场作为垃圾覆盖材料或垫层材料,也可经进一步处理用作矿山填充材料。利用固化后的飞灰固化体掺入混凝土中用来制备半干混凝土,或与公路基料拌成混合料,用于铺筑公路路面基层[35]。飞灰基地聚合物通过模具还可浇筑成不同形状的固化体,用于沙漠地区的公路、铁路挡沙墙,沙区农户的挡沙柱和挡沙网,以及沙地道路路基材料等。
6. 研究展望
目前应用偏高岭土、矿渣和粉煤灰作为基质材料的地聚合物固化技术,是国内外飞灰地聚合物固化稳定化的主要研究方向,其原料研究范围较窄,需要进一步拓宽飞灰地聚合体系种类研究范围,广泛探寻硅源、铝源材料,为飞灰地聚合物的制备提供更多的基质来源材料。
飞灰地聚合物制备过程的反应机理尚未有系统认识,对二噁英等持久性有机污染物的固化机理研究也很少,地聚合物固化稳定化重金属的机理尚需进一步深入探究。在今后的固化机理研究中,应进一步明确飞灰地聚合物的反应机理与产物性能的关系,能更好为固化稳定化飞灰提供可靠的理论依据。
地聚合物固化体制备方式目前也比较单一,应在今后的研究中探索更多的固化体制备方法,建立一体化制备工艺,减少中间操作环节,并充分综合利用大宗固废,从而使实际应用更为简易,材料成本更为低廉,社会效益更为显著。由于飞灰属于国家规定的危险废物,对飞灰基地聚合物固化体的制备现多停留在实验室水平,走出填埋场进行资源化应用尚需核心技术攻关和国家政策允许,大规模的产业化仍需进一步探究。
-
表 1 小麦秸秆和模拟粪便的基本性质
Table 1. Basic properties of wheat straw and simulated human feces
物料名称 含水率/% pH 总碳/% 总氮/% 碳氮比/% 小麦秸秆 4.54±0.78 5.74±0.01 40.68±0.59 1.25±0.06 32.55±1.45 模拟粪便 3.82±0.13 5.76±0.03 49.80±0.25 4.53±0.01 11.01±0.04  下载: 导出CSV
下载: 导出CSV
-
[1] GUO S S, TANG Y K, ZHU J T, et al. Development of a CELSS experimental facility[J]. Advances in Space Research, 2008, 41(5): 725-729. doi: 10.1016/j.asr.2007.09.029 [2] ZHANG N, LI J L, LUO J, et al. Wheat cultivation and nutrient control for the 180-day CELSS integrated experiment[J]. Life Sciences in Space Research, 2020, 26: 46-54. doi: 10.1016/j.lssr.2020.04.001 [3] CIURANS C, BAZMOHAMMADI N, POUGHON L, et al. Hierarchically controlled ecological life support systems[J]. Computers & Chemical Engineering, 2022, 157: 107625. [4] 张秧, 艾为党, 冯海艳, 等. 小麦秸杆好氧堆肥过程中微生物多样性与优势菌群分析[J]. 农业工程学报, 2021, 37(11): 206-212. doi: 10.11975/j.issn.1002-6819.2021.10.025 [5] 张海霞, 李爱民, 鞠茂伟. 小型好氧堆肥设备处理有机垃圾[J]. 环境工程学报, 2013, 7(5): 1939-1944. [6] 聂二旗, 郑国砥, 高定, 等. 适量通风显著降低鸡粪好氧堆肥过程中氮素损失[J]. 植物营养与肥料学报, 2019, 25(10): 1773-1780. doi: 10.11674/zwyf.18432 [7] PENG L J, TANG R L, WANG G Y, et al. Effect of aeration rate, aeration pattern, and turning frequency on maturity and gaseous emissions during kitchen waste composting[J]. Environmental Technology & Innovation, 2023, 29: 102997. [8] ZHANG L X, GAO X Z, SHI T, et al. Regulating aeration intensity to simultaneously improve humification and mitigate gaseous emissions in food waste digestate composting: Performance and bacterial dynamics[J]. Science of the Total Environment, 2023, 889: 164239. doi: 10.1016/j.scitotenv.2023.164239 [9] WIGNARAJAH K, LITWILL E, FISHER J, et al. Simulated human feces for testing human waste processing technologies in space systems[J]. SAE Technical Paper Series, 2006, 115: 424-429. [10] 张相锋, 王洪涛, 周辉宇, 等. 花卉秸秆和牛粪联合堆肥的中试研究[J]. 环境科学学报, 2003(3): 360-364. [11] 陈维. 探究温度对酵母菌种群数量的影响[J]. 生物学通报, 2008, 43(12): 46-48. doi: 10.3969/j.issn.0006-3193.2008.06.019 [12] 陈文旭, 刘逸飞, 蒋思楠, 等. 微生物菌剂对厨余垃圾堆肥温室气体减排的影响[J]. 农业工程学报, 2022, 38(23): 181-187. doi: 10.11975/j.issn.1002-6819.2022.23.019 [13] 孔建松, 郑玉琪, 陈同斌. 好氧堆肥过程中的氧气变化及其监测[J]. 生态环境, 2003, 12(2): 237-239. [14] 陈是吏, 袁京, 李国学, 等. 过磷酸钙和双氰胺联用减少污泥堆肥温室气体及NH3排放[J]. 农业工程学报, 2017, 33(6): 199-206. doi: 10.11975/j.issn.1002-6819.2017.z1.030 [15] 徐智, 张琴, 张陇利, 等. 强制好氧堆肥不同阶段氧气浓度变化及其与腐熟指标关系[J]. 农业环境科学学报, 2009, 28(1): 189-193. doi: 10.3321/j.issn:1672-2043.2009.01.035 [16] 李翔. 电场促进好氧堆肥腐熟与减少N2O排放机理研究[D]. 福建农林大学, 2020. [17] CHEN H Y, AWASTHI S K, LIU T, et al. Effects of microbial culture and chicken manure biochar on compost maturity and greenhouse gas emissions during chicken manure composting[J]. Journal of Hazardous Materials, 2020, 389: 121908. doi: 10.1016/j.jhazmat.2019.121908 [18] ZHANG B X, XU Z C, JIANG T, et al. Gaseous emission and maturity in composting of livestock manure and tobacco wastes: Effects of aeration intensities and mitigation by physiochemical additives[J]. Environmental Technology & Innovation, 2020, 19. [19] 芶久兰, 罗文海, 袁京, 等. 通风速率对鸡粪-烟末联合堆肥腐熟度和污染气体排放的影响[J]. 西南农业学报, 2021, 34(4): 872-879. doi: 10.16213/j.cnki.scjas.2021.4.027 [20] SANTOS A, BUSTAMANTE M A, TORTOSA G, et al. Gaseous emissions and process development during composting of pig slurry: the influence of the proportion of cotton gin waste[J]. Journal of Cleaner Production, 2016, 112: 81-90. doi: 10.1016/j.jclepro.2015.08.084 [21] CAO Y B, WANG X, MISSELBROOK T, et al. Quantification of N and C cycling during aerobic composting, including automated direct measurement of N2, N2O, NO, NH3, CO2 and CH4 emissions[J]. Science of The Total Environment, 2023, 857(2): 159177. [22] HAN Z L, SUN D Z, WANG H, et al. Effects of ambient temperature and aeration frequency on emissions of ammonia and greenhouse gases from a sewage sludge aerobic composting plant[J]. Bioresource Technology, 2018, 270: 457-466. doi: 10.1016/j.biortech.2018.09.048 [23] 吴伟祥, 李丽劼, 吕豪豪, 等. 畜禽粪便好氧堆肥过程氧化亚氮排放机制[J]. 应用生态学报, 2012, 23(6): 1704-1712. doi: 10.13287/j.1001-9332.2012.0276 [24] JIANG T, LI G X, TANG Q, et al. Effects of aeration method and aeration rate on greenhouse gas emissions during composting of pig feces in pilot scale[J]. Journal of Environmental Science, 2015, 31: 124-132. doi: 10.1016/j.jes.2014.12.005 [25] 常远, 李若琪, 李珺, 等. 好氧堆肥腐殖酸形成机制及促腐调控技术概述[J]. 中国环境科学, 2023: 1-17. [26] 蔡涵冰, 冯雯雯, 董永华, 等. 畜禽粪便和桃树枝工业化堆肥过程中微生物群演替及其与环境因子的关系[J]. 环境科学, 2020, 41(2): 997-1004. doi: 10.13227/j.hjkx.201907153 [27] 李荣华, 张广杰, 秦睿, 等. 添加钝化剂对猪粪好氧堆肥过程中理化特性的影响[J]. 环境科学学报, 2012, 32(10): 2591-2599. doi: 10.13671/j.hjkxxb.2012.10.004 [28] WANG Y M, TANG Y, LI M Y, et al. Aeration rate improves the compost quality of food waste and promotes the decomposition of toxic materials in leachate by changing the bacterial community[J]. Bioresource Technology, 2021, 340: 125716. doi: 10.1016/j.biortech.2021.125716 [29] GARCíA C, HERNáNDEZ T, COSTA F. Study on water extract of sewage sludge composts[J]. Soil Science and Plant Nutrition, 1991, 37: 399-408. doi: 10.1080/00380768.1991.10415052 [30] 代静玉, 周江敏, 秦淑平. 几种有机物料分解过程中溶解性有机物质化学成分的变化[J]. 土壤通报, 2004(6): 724-727. [31] 李思雨. 市政污泥好氧堆肥氮素转化及氨气控制研究[D]. 河北工程大学, 2022. [32] 杨慧珍. 碳氮比对好氧堆肥过程中有机碳转化的影响机制研究[D]. 甘肃农业大学, 2022. [33] 刘宁, 周嘉良, 马双双, 等. 生物炭对鸡粪好氧堆肥主要氮素形态含量影响与保氮机制[J]. 农业机械学报, 2016, 47(12): 233-239. doi: 10.6041/j.issn.1000-1298.2016.08.030 [34] 秦维, 蒋心茹, 刘明刚, 等. 辅料配比及通风速率对污泥气流膜堆肥过程氮素迁移的影响[J]. 农业环境科学学报, 2022, 41(6): 1357-1367. [35] 杨新, 杨双全, 陈莉, 等. 以酒糟为基质的高温型生物有机肥复合发酵菌剂的制备[J]. 食品与发酵工业, 2019, 45(9): 242-249. doi: 10.13995/j.cnki.11-1802/ts.019887 [36] 宋宪强, 刘志聪, 杨宗谚. 固体废弃物高温好氧堆肥过程中重金属稳定化研究[J]. 城市地理, 2016(20): 67-67. [37] 中华人民共和国工业和信息化部. 生物质腐植酸有机肥料: HG/T 6082-2022[S]. 北京: 化学工业出版社, 2022. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 2686
- HTML全文浏览数: 2686
- PDF下载数: 58
- 施引文献: 0
