


-
景观水体尤其是城市景观水体大多为静态、流动性较差的封闭式缓流水体,具有水域面积小、水环境容量小、自净能力差、易受污染等特点,往往面临风险较高的水质恶化等问题[1-3]。底泥氮磷营养盐的释放是造成水体内源污染的重要因素,控制景观水体及沉积物内部污染物负荷已成为景观水体治理的关键。
原位处理技术和异位处理技术在景观水体底泥处理中运用广泛。物理方法比如底泥疏浚是最直接、有效的异位内源污染治理技术,可快速将受污染的沉积物从水体中清除,增加水体容量、改善水质情况;但其后续底泥处理量大、运输困难,工艺流程复杂且资源化利用有限,在时间空间上具有很大限制,成本高且不易操控[4],还存在破坏水底生态系统平衡,易造成二次污染等风险。投加化学试剂 (如铁铝盐、过氧化钙、锁磷剂等) 是常见的原位化学方法,能紧急修复污染水质,但是有增加水体毒性的可能性[5-7]。底泥覆盖是常见的原位物理处理技术,国外已有将覆盖技术成功应用于治理受污水体的案例[8-10],我国学者也开展相关研究。在近年研究中,天然矿物由于具有价格低廉、来源较广等优点,被广泛应用于实际污染底泥的治理中[11],如方解石、沸石、石灰石、白云石、伊利石、麦饭石等。这些材料应用于覆盖底泥中,对抑制氮、磷释放效果明显,清洁土壤比如红壤也由于良好的絮凝吸附性能、分布较广等优点被逐渐用于水体治理中[12]。学者普遍认为,底泥中存在着大量的可生物利用的营养物质, 这些营养物质可被生物通过生命代谢活动进行吸附、降解、转化,从而促进其生长[13-14] ,污染底泥资源化在水生态处理中有良好的运用前景。沉水植物可利用底泥中的N、P营养盐,还可通过影响微生物的代谢来影响营养物质的循环,在稳定生态系统和净化水体方面有特殊作用。
针对水体污染程度、地理位置等情况,选用处理效果好且经济效益高的底泥改造材料很有必要。紫色母岩是一种自然沉积岩,在西南地区广泛分布,矿物成分较砾石复杂,过去的研究大多集中在紫色母岩风化物理颗粒变化和破碎成土[15-16]、土壤营养肥力[17]等反面。近年来,对紫色母岩的研究逐渐深入到了其吸附性能方面。黄雪娇等[18]发现使用重庆地区常见的蓬莱镇组、沙溪庙组、飞仙关组和遂宁组紫色母岩作基质均能有效降低水体氮磷及有机物含量,其中蓬莱镇组效果最佳。石灰石(主要成分CaCO3)是人工湿地的常用的天然矿物基质,对水体污染物有很好的去除作用[19]。白云石(主要成分CaMg(CO3)2)与石灰石结构相似,被应用于水体治理中,是低成本的除磷材料[20]。
微生物是水生态环境中重要的组成部分,在氮、磷营养盐消减过程中发挥着重要作用[21-22],水环境也会影响微生物群落结构或丰度,了解微生物在水体氮磷变化中的分布特征、生态功能和作用等对水环境修复有重要意义。过去许多研究多集中在改造材料提高底泥吸附性能或者对沉水植物的生理生态影响上[23-25],而结合水环境物化性质变化与微生物群落结构变化的研究较少。为此,本研究选取蓬莱镇紫色母岩、石灰石、白云石作为底泥改造材料,通过阶段性的水质监测分析不同改造材料对水体氮磷去除效果及差异,结合水体元素质量浓度变化和微生物群落结构变化探讨差异的原因,以期评估出低耗高效的底泥改造材料,并在一定程度上丰富底泥改造的应用基础理论,同时为同类型污染水体底泥改造材料的选择提供参考。
-
供试底泥改造材料蓬莱镇组紫色母岩采自重庆市巴南区界石镇附近的倒置低山区,于通风干燥处风干后去除其中树枝、落叶等杂质,过60目筛备用;石灰石与白云石均购自于灵寿县北堤下村矿工厂,经干燥后磨碎,过60目筛备用。改造材料基本性状见表1。矮生苦草(Vallisneria natans (Lour.) Hara)为实验室养殖,挑选长势良好并相近的植株使用去离子水清洗表面杂质,每10株一组备用。底泥和试验用水取自西南大学崇德湖,供试底泥基本性质为全氮2.69 g·kg−1、全磷11.75 g·kg−1、pH6.9。
-
采用5 L斗抓式底泥采样器 (购于绍兴市上虞区亦晨仪器经营部) 于西南大学崇德湖采取底泥,静置至底泥与上层水体分离,倒去表面水体,过0.25 cm网筛去除底泥沉积物中的石块、腐叶等杂物。加入8 cm底泥至聚乙烯圆桶 (上口径∶下口径∶高=45.5 cm∶34.2 cm∶56.4 cm) 内,分别铺设2 cm石灰石 (T2) 、白云石 (T3) 与蓬莱镇紫色母岩 (T4) ,以不加任何底泥改造材料作为对照 (T1) ,加水略微淹没后静置24 h,待上覆水澄清后将10株长势相同的矮生苦草均匀种植于各桶内,加入200 L混合湖水至桶高40 cm左右。每组设置3个平行处理。
待沉水植物生长稳定后 (35 d) 进行换水,倒出桶内水体,再平稳加入实验水体进行水质监测。每一次换水后在每天上午9:00室外静水条件下采用虹吸法于水面下20 cm、30 cm处混合取样进行为期5 d的水质监测,共监测4次。再按公式 (1) 计算氮、磷的去除率。
式中:A0表示上覆水氮、磷初始质量浓度,mg·L−1,At表示氮、磷浓度测定值,E代表去除率。
在室外静水条件下连续监测 (5 d) ,通过对水质变化的分析确定分别在 0 h、12 h、36 h、60 h、84 h、108 h按照上述相同方法取样,水体初始 TN为(3.67±0.09) mg·L−1、初始TP为(0.46±0.02) mg·L−1、初始NH4 + -N为(0.65±0.03) mg·L−1、初始pH为 (7.74±0.03) 。分析认为36 h时的处理情况值得进一步研究,故对该时刻数据进行详细分析测定。采用虹吸法在沉水植物叶片周围1~2 mm处少量多次采集混合水样,进行元素质量浓度测定和细菌16S rRNA测定。
-
水质指标均按照地表水水质指标分析国家标准进行测定。总磷测定采用钼酸铵分光光度法(GB 11893-89), 总氮采用碱性过硫酸钾紫外分光光度法,氨氮使用靛酚蓝比色法,pH测定采用玻璃电极法(GB 6920-86)。
水体元素质量浓度使用电感耦合等离子体发射光谱仪(ICP- 3000)进行测定。取2 L水样采用0.22 µm孔径聚碳酸酯滤膜进行抽滤 (每次抽滤前装置用一级纯水反复清洗) ,使用第2次抽滤后的水体进行元素测定。
在抽滤完成后,将滤膜置于-80 ℃低温保存,之后进行微生物组总DNA提取,目标片段PCR扩增。使用DNA试剂盒(Omega Bio-tek, Norcross, GA, usa)从样本中提取基因组DNA。DNA提取液在1%琼脂糖凝胶上进行检测,并使用NanoDrop 2000 UV-vis分光光度计(Thermo Scientific, Wilmington, USA)测定DNA浓度和纯度。细菌基因使用引物对338F (5'ACTCCTACGGGAGGCAGCAG3')和806R (5'GGACTACHVGGGTWTCTAAT3')通过ABI GeneAmp®9700 PCR热循环器(ABI, CA, USA)进行扩增。16S rRNA高通量测序在上海美吉生物医药科技有限公司进行,数据分析基于该公司所提供的云服务进行 (www.majorbio.com) 。
-
Office excel 2010用于实验数据整理并制图;SPSS 25.0用于单因素方差分析(One-Way ANOVA)和最小显著性差异检验(LSD),用Origin2019b绘图。
-
在沉水植物生长稳定后的第10、20、35和 55天分别以5 d为1个周期,进行了4次氮磷去除率的监测。如图1和图2所示,由于测定时间不同,各个周期间不同处理的去除率均有一定波动。这可能是由于气候变化引起的,但整体来看各周期内不同处理间的氮、磷去除率趋势较一致,这说明系统效果比较稳定。各处理对氮、磷的去除率均有一定提高,除了底泥吸附的作用外,苦草可通过直接吸收、过滤吸附等作用去除水体的氮磷[26]。不同底泥改造处理间有差异,T2、T3、T4处理在监测周期内中的监测时间对TP 的去除率均明显高于T1。在第5天 时,除T1外,对TP的去除率均能达到80%以上,T4处理对TP的去除率最高。T1对TN的去除率在各个监测周期内也明显低于其余3组,T3对TN的去除效果最好。这是由于除了苦草的作用外,底泥覆盖能抑制底泥中氮、磷营养盐的释放,同时对上覆水中氮、磷悬浮物颗粒吸附作用较强[11]。
-
水体中的磷主要依靠植物直接吸收、植物体吸附过滤和物理化学沉淀等途径去除[27]。各系统对磷的去除在108 h内有相似的变化趋势(图3(a))。除T1外,各处理水体中的总磷均能降至0.1 mg·L−1以下,达到《地表水环境质量标准》 (GB3838-2002) Ⅳ类水标准。不同的底泥改造材料对磷的去除效果不同, T4在实验周期内对磷的去除效果最好,达到87.02%,与T1具有极显著差异 (P<0.01) ,T1为72.60%,T2、T3与T1有显著差异 (P<0.05) 。在36 h时, T2、T4与T1均具有显著差异 (P<0.05) 。
水体中氮的去除除了磷的去除途径外,还存在氨的挥发、微生物固氮、脱氮、氨化、硝化等生命活动等途径[28-29]。各系统总氮和氨氮质量浓度均在36 h时表现出显著差异(图3(b)和(c))。TN在84 h后趋于稳定。在108 h时,T4对总氮的去除效果最佳,达到了80.96%,T1为71.26%。在36 h时 T3对氮的去除效果最好,TN去除率达到71.49%,而T1仅有58.00%。在12~36 h,各系统对TN去除率与T1相比均有显著差异 (P<0.05) 。在60 h时,T3、T4与T1有显著差异 (P<0.05) 。在84 h后,各系统间均无显著差异。[NH4+-N]在36 h时骤降,之后在一定范围内波动,这是由于氨氮受环境影响较大,在pH>7.0时容易挥发[18],微生物硝化作用和沉水植物作用均会影响水体[NH4+-N]。T1、T2、T3、T4的[NH4+-N]分别为:(0.10~0.13) mg·L−1、(0.05~0.08) mg·L−1、(0.04~0.09) mg·L−1、(0.08~0.10) mg·L−1。在36 h时,T3对NH4+-N去除率最高,达到了92.81%。
已有研究表明蓬莱镇组紫色母岩作为底泥覆盖材料对氮、磷的吸附的效果优于石灰石。这是由于母岩具有团粒结构,表面蓬松,相较于矿物具有更多的吸附位点[30]。石灰石和白云石组成相似,但白云石对磷素的理论饱和吸附量比石灰石低[31-32],与本实验研究结果相似,而对氮的吸附效果白云石优于石灰石。推测除了吸附作用外,还与其他因素有关。
-
为进一步分析不同处理间氮磷去除率差异的原因,探究了上覆水体中的元素指标及其变化。表2表明,各组处理上覆水体中砷(As)、镉(Cd)、铜(Cu)、铁(Fe)、锰(Mn)、钼(Mo)元素的质量浓度无明显差异。不同处理间钙(Ca)、镁(Mg)的质量浓度具有显著差异 (P<0.05) ,T4中Ca的质量浓度显著高于T1,T1中的Mg的质量浓度显著低于其余各组。这与不同改造材料的元素浸出能力不同有关[33]。Ca、Mg不仅通过沉淀吸附促进N、P去除,也通过影响微生物群落强化生物脱氮除磷过程。这是由于Ca、Mg等活性物质可与水体中可溶性磷酸盐通过配位体交换作用发生吸附和沉淀作反应[32]。沉水植物也通过光合作用所形成的微环境促进钙、磷的共沉淀过程[34]。水体中的Mg2+能与铵根离子(NH4+)、磷酸根离子 (HPO42-) 反应生成难溶性的鸟粪石沉淀,从而消除水体中的氮、磷营养盐[35]。同时,作为除磷反应中酶促剂的重要组成成分参与整个生物除磷过程,有利于强化生物除磷过程[36]。
-
SMVS使上覆水悬浮态颗粒物中的微生物群落结构发生改变。对36 h不同SMVS系统中细菌群落组成进行分析,结果表明细菌种类共注释到9个门、12个纲、24个目、28个科、46个属和48个种。如图4(a)所示,在门的分类水平上,各处理细菌组成相似,但相对丰度存在差异。变形菌门 (Proteobacteria)、放线菌门 (Actinobacteriota)、蓝藻门 (Cyanobacteria)、拟杆菌门(Bacteroidota)是占比较大的菌门,相对丰度分别为34.7%~42.3%、25.0%~36.8%、9.5%~14.0%、9.2%~18.4%;异常球菌门(Deinococcota)、厚壁菌门(Firmicutes)、绿弯菌门(Chloroflexi)和疣微菌门(Verrucomicrobia)占比较低,相对丰度为0.20%~5.6%、1.4%~2.8%、0.55%~5.1%和0.65%~1.0%;浮霉菌门(Patescibacteria)占比小于1%。通过克氏秩和检验 (Kruskal-Wallis H test) 对不同处理下细菌比例是否发生显著变化进行分析,结果显示底泥改造后放线菌门、拟杆菌门、厚壁菌门、绿弯菌门相对丰度在各处理间存在显著差异 (P<0.05) ,不同底泥改造条件下上覆水微生物相对丰度在门水平上发生了变化 (图5(a)) 。放线菌门在T3中占比高于其余3组,T1中拟杆菌门占比低于其余各组,而厚壁菌门在T1中占比最高。放线菌门能吸收水体中的营养物质作为氮源,在降解氮素营养盐过程中发挥重要作用[37],结合使用白云石作为底泥改造材料对氮的去除效果最好,推测以白云石作为改造材料有利于提高放线菌门丰度,促进水体氮的去除。拟杆菌门是一类复杂碳水化合物多产降解者,能将纤维素转化成可溶性糖类,可参与到腐败植物组织分解的碳循环中[38]。大多数厚壁菌门是一类在污水厌氧处理中起到重要作用的微生物[39],当水体中厚壁菌门占比较大时,水体已经受到较为严重的污染,能说明在本实验条件下底泥改造后可缓解水体污染情况。
在属水平上,各处理优势菌属相同,但较优势菌属存在差异。叶绿菌目未定属 (norank_f_norank_o_Chloroplast) 、分枝杆菌属 (Mycobacterium)、根瘤菌科未定属 (norank_f_Rhizobiales_Incertae_Sedis) 、CL500-29_marine_group为优势菌属,分别占比9.0%~14.3%、6.9%~12.0%、5.0%~10.9%、4.7%~9.0%。 T1以不动杆菌属(Acinetobacter)、古字状菌属 (Runella) 和异常球菌属(Deinococcus)为较优势菌属,分别占比7.7%、6.0%、5.6%;T2以林土鼎湖杆菌属(Dinghuibacter)、微小杆菌属(Exiguobacterium)、hgcI_clade占比较大,分别占比8.2%、4.2%、4.1%;T3则以hgcI_clade、PeM15、林土鼎湖杆菌属(Dinghuibacter)为较优势菌种,分别占比4.3%、3.6%、3.0%;T4以hgcI_clade、林土鼎湖杆菌属(Dinghuibacter)、异常球菌属(Deinococcus)占比相对较大,分别为3.1%、2.3%、2.1%。不动杆菌属在T1占比较大,而在其余3组中占比均小于0.2%;红细菌属(Rhodobacter)在T2中占比相对最高,为4.4%,在其余3组中占比均小于1.2%。如图5(b)所示,在属水平上,叶绿菌目未定属、分枝杆菌属、不动杆菌属、hgcl_clade、Runlla在不同处理中的相对丰度发生了显著性变化。分枝杆菌属、norank_f_Rhizobiales_Incertae_Sedis在T3中相对丰度最高;norank_f_norank_o_Chloroplast在T2、T4中占比高于T1; hgcI_clade在T1中占比显著低于其余3组 (P<0.05) 。值得注意的是,不动杆菌属被认为是除磷能力较强的微生物,然而在T1中占比最高,显著高于其余3组 (P<0.05) 。GAO等[40]发现不动杆菌属广泛存在于污水排放的产生富营养化河段,结合本研究结果说明未进行底泥改造上覆水磷含量高更趋向于水体富营养化,适宜不动杆菌属生长。分枝杆菌属具有脱氮与分解有机物的功能[41-42],这也进一步解释了T3中氮素营养盐去除较快的原因。hgcI_clade对碳水化合物和富氮化合物有较强的吸收能力[43],通过参与上覆水体中的碳氮循环以达到对去除氮素营养盐效果。与T1相比,改造底泥有利于提高脱氮微生物占比。叶绿菌目未定属可利用光合作用吸收水体中的营养物质,促进上覆水磷的去除[44],故在T2、T4中磷含量较低。
-
微生物多样性指数是反映微生物群落结构特征的重要指标,SMVS影响了微生物多样性。在97%相似度水平下对细菌群落进行多样性分析,主要研究丰富度指数Chao1及多样性指数Shannon和Simpson。表3表明,进行底泥改造后上覆水细菌群落多样性明显升高,总体呈现群落多样性T4>T2>T3>T1。这可能与改造材料本身含有微生物有关,使用蓬莱镇紫色母岩作为改造材料明显提高了上覆水微生物群落多样性。
-
进行VIF方差膨胀因子分析排除共线性较强的环境因子,选择TP、TN、As、B、Cu、Mo、Mn、Ni、Ca、Mg、Simpson在门水平和属水平上进行冗余分析(RDA)。如图6(a)所示,第一、第二排序轴分别解释总变量的40.87%和16.93%,环境因子对不同处理中细菌门类分布的影响较大。T2、T4与 Ca相关性较高,T3与Mg相关性高。放线菌门与T3有明显的正相关性,且与TN呈明显负相关关系,放线菌门受Mg影响大。这说明使用白云石为底泥改造材料可能通过影响上覆水Mg含量进而促进放线菌门生长,从而提高TN去除率。Ca质量浓度与TP有明显的负相关关系。Simpson多样性指数与TN呈线性相关,微生物多样性越高,Simpson指数越低,TN越低。这说明提高微生物多样性有利于TN去除,与PRATT等[45] 的研究结果相反,这说明TN还受其他因素共同影响。如图6(b)所示,2个排序轴对细菌属类分布差异的解释度总共为47.97%。T1与T2、T3、T4的分布有明显差异,这说明底泥改造对上覆水体细菌群落结构影响较大。Ca、Mg对各处理均有影响,与TP、TN具有明显的负相关关系。
对比以往的研究,使用底泥改造材料协同沉水植物比使用单一底泥改造材料对氮、磷的去除率在短时间内有所提高[46]。这可能是由于沉水植物的增效作用。相关研究表明,在水体氮、磷去除中沉水植物通过吸附、改善生境提高水体微生物转化等而产生的净水增效作用贡献率明显大于对氮、磷的直接吸收。相反,在底泥-沉水植物-微生物构建的体系中,微生物在水体N、P的削减中起到了较大的净化作用[47]。本研究发现氮、磷去除效果的差异与底泥改造影响上覆水微生物群落有关。微生物与植物协同作用具有较强的净化能力,然而对于沉水植物的增效作用如何对微生物产生影响以及相互作用机制尚不清楚,有待进一步研究。
-
1) SMVS能提高水体氮磷营养盐的去除效率,在污染频率较高的水体的治理中具有很好的应用前景,研究结果可为景观水体短期水质危机提供解决思路。2) 使用蓬莱镇母岩作为底泥改造材料协同沉水植物对上覆水体的氮磷营养盐净化效果最佳,对磷素营养盐尤为明显,在景观水体底泥中作为改造材料稳定水体质量方面具有良好的应用前景。3) SMVS提高了上覆水Ca、Mg的质量浓度,驱动上覆水体微生物群落结构发生改变,强化了与脱氮除磷微生物的联系,促进水体氮磷的去除。
底泥-苦草系统对景观水体氮磷的去除效果及其中微生物群落的结构变化
Nitrogen and phosphorus removal effect of sediment-Vallisneria natans system on landscape water and changes of microbial community structure
-
摘要: 底泥-沉水植物-微生物系统在景观水体氮磷消减过程中有关键作用。为缓解底泥氮磷释放压力并探究底泥改造材料的选择与氮磷去除效果之间的联系,选用石灰石、白云石和蓬莱镇组紫色母岩作为改造材料,构建水-底泥-苦草系统,结合水体元素质量浓度和微生物群落结构进行分析。结果表明,底泥改造联合苦草 (sediment modification combined with Vallisneria natans,SMVS) 对上覆水体氮、磷的去除效果相较于未改造处理显著提高 (P<0.05) ,蓬莱镇紫色母岩组处理效果最好,对总磷、总氮的去除率分别达到87.06%、80.96%。上覆水悬浮态颗粒物中微生物群落结构发生改变,在门分类水平上, Actinobacteriota、Bacteroidota丰度在不同改造处理中存在显著差异 (P<0.05) ;在属分类水平上,norank_f_norank_o_Chloroplast、Mycobacterium为优势菌属,Mycobacterium在不同改造处理中存在显著差异 (P<0.05) 。冗余分析结果表明Ca、Mg是驱动上覆水氮磷营养盐降低的主要元素。蓬莱镇组紫色母岩作为底泥改造材料对氮、磷的去除效果优于石灰石和白云石,这与向上覆水中释放微量元素含量不同,进而改变了微生物群落结构有关。本研究结果可为景观水体污染底泥的治理提供参考。Abstract: The sediment-Submerged macrophytes-Microbial system plays a key role in the reduction of nitrogen and phosphorus in the nitrogen and phosphorus reduction process of in landscape water. In order to alleviate the nitrogen and phosphorus release pressure of sediment and explore the relationship between the selection of sediment transformation materials and the effect of nitrogen and phosphorus removal, limestone, dolomite and purple parent rock of Penglaizhen Formation were selected as the transformation materials to construct a water-sediment- Vallisneria natans system, which was analyzed by combining the mass concentration of water elements and microbial community structure. The results showed that the removal effect of sediment modification combined with Vallisneria natans (SMVS) was significantly higher than that of the overlying water body (P<0.05), and the treatment effect of Penglaizhen Formation was the best with the removal rates of TP and TN at 87.06% and 80.96%, respectively. The microbial community structure in the overlying aqueous suspended particulate matter had changed, and the abundance of Actinobacteriota and Bacteroidota was significantly different in different modification treatments at the taxonomic level (P<0.05). At the genus taxonomic level, there were significant differences between norank_f_norank_o_Chloroplast and Mycobacterium in different modification treatments (P<0.05). The results of redundancy analysis showed that Ca and Mg were the main elements driving the reduction of N and P of overlying. The removal effect of purple parent rock in Penglaizhen Formation as a sediment modification material was better than that of limestone and dolomite, which was related to the release of trace elements in upward overlying water and the change of microbial community structure. The research results can provide reference for the treatment of sediment polluted by landscape water.
-
印染行业是用水量和废水排放量均较大的行业之一。印染加工中预处理、染色、印花和整理等工序产生的废水混合而成印染废水,这个过程中残余的有机染料会随之进入到废水中[1]。有机染料种类繁多,按分子结构分类可分为偶氮染料、蒽醌染料、靛系染料、酞菁染料、硫化染料及硝基和亚硝基染料等[2]。因有机染料分子中含有芳香结构而具有化学稳定性强和生物可降解性差的特性,甚至一些有机染料具有生物毒性,导致传统的生物处理工艺难以将其脱色和去除[3]。印染废水生化出水中含有未降解的有机染料,使其色度难以达到生产回用要求或排放标准,已经成为印染废水处理的一大难题[4]。
针对上述问题,基于羟基自由基的高级氧化技术被认为是可以有效降解有机染料的处理方法并得到了广泛研究[5]。光催化氧化、Fenton氧化和电化学氧化等高级氧化技术被用于降解水中的有机染料,并显示出较好的处理效果[6-8]。然而对于印染废水而言,其生化出水具有水量大、污染物含量低和仅需脱色的特点,体系复杂及能耗高的高级氧化技术难以胜任。因此,印染废水的处理需要一种流程简单、脱色能力强且无二次污染的高级氧化技术。真空紫外高级氧化技术可满足上述全部要求并在近年来得到了广泛关注,逐渐成为研究热点。真空紫外是指波长在100~200 nm的紫外光,相对于其他波段的紫外光具有更高的能量。低压汞灯是典型的真空紫外光源之一,当灯管的材质为超纯石英时可同时辐射出波长为185 nm的真空紫外光和254 nm的UV-C波段紫外光,即为双波长紫外光源(UV185/UV254)。185 nm真空紫外光的光子能量达6.70 eV,可以将水分子直接光解生成∙OH、∙H和eaq−等高活性的氧化和还原物种[9]。UV185辐照的光子能量高于部分有机污染物分子化学键的键能,有机污染物可通过化学键直接断裂而被降解[10]。已有研究表明,双波长紫外可通过直接光氧化有效降解乐果[11]、1,4-二恶烷[12]和邻氯苯酚[13]等有机污染物。
针对不同有机染料在双波长紫外辐照体系中直接光氧化降解的对比研究,目前还鲜有报道。本研究以低压汞灯为双波长紫外光源,选取MB、RhB和AO7等典型有机染料为模型污染物,考察了UV185辐照强度、有机染料初始质量浓度和初始pH等因素对有机染料降解效能的影响,确定了主要的活性物种,并结合模型污染物光降解过程中紫外-可见光吸收光谱分析结果,推测出有机染料可能的降解机制。
1. 材料与方法
1.1 实验试剂和材料
实验中所用试剂纯度均为分析纯,亚甲基蓝(MB)、酸性橙7(AO7)、罗丹明B(RhB)、碘化钾(KI)和钼酸铵(H8MoN2O4)等购自成都科龙化工试剂厂,碳酸氢钠(NaHCO3)购自重庆博艺化学试剂有限公司,过氧化氢(H2O2)、氢氧化钠(NaOH)、盐酸(HCl)、对苯醌(p-BQ)和邻苯二甲酸氢钾(C8H5KO4)购自国药集团化学试剂有限公司。实验中所用的超纯水由Direct-Pure UP型(RephiLe)超纯水器制取。
1.2 实验装置及方法
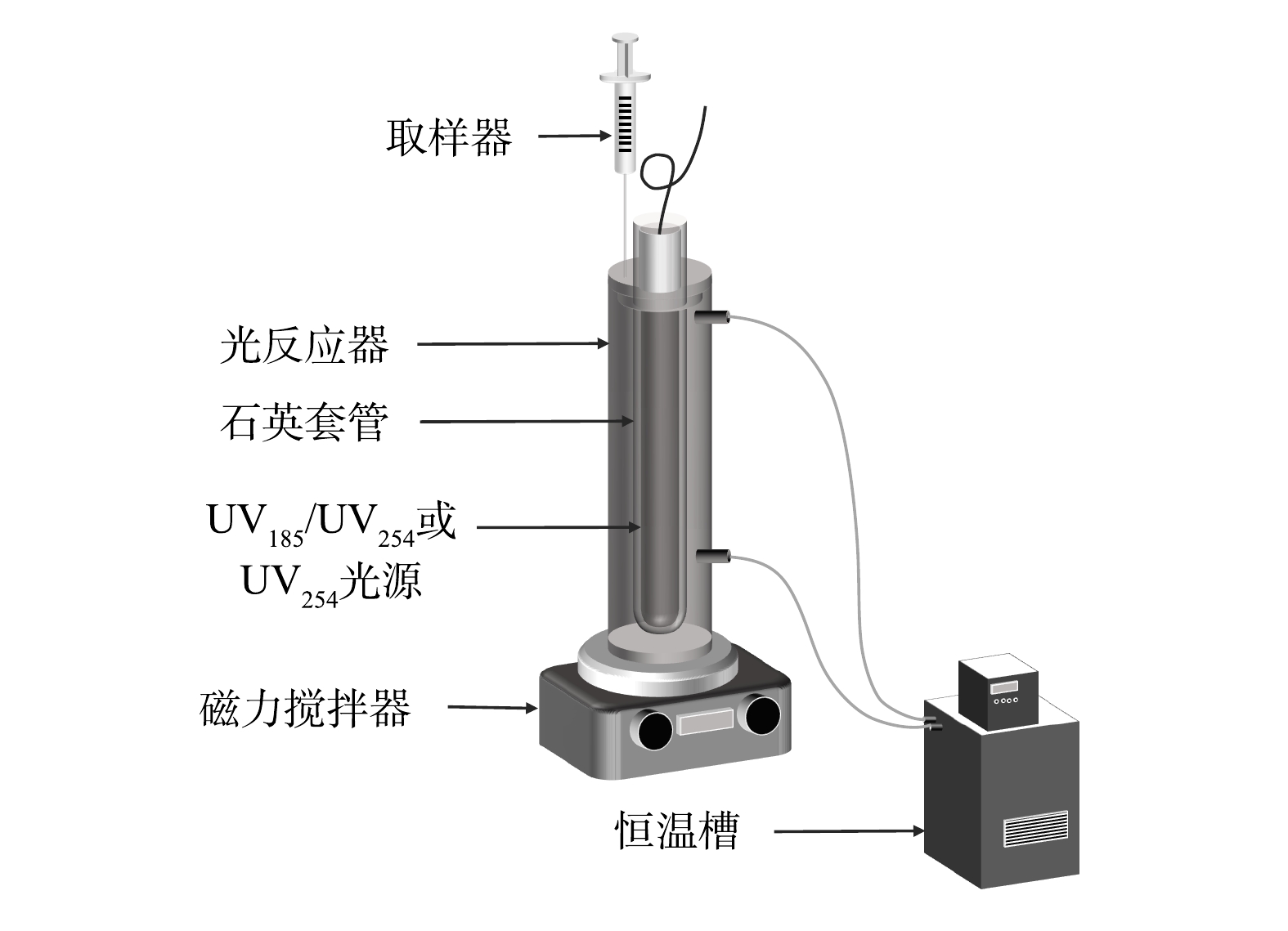
如图1所示,光反应器为一个自制的圆柱形玻璃管,有效容积为300 mL。光反应器外侧有水浴套,连接低温恒温槽(DC1006,宁波新芝生物科技股份有限公司)来保持反应体系温度稳定。实验光源为一个功率为8 W的双波长紫外灯(型号:10-08100,实验中紫外光源均来自北京航天宏达光电技术股份有限公司),可以同时发出UV185和UV254两种波长的紫外线,紫外灯置于石英套管中并位于光反应器中轴位置。实验中不同真空紫外强度分别由UV185辐照强度为10%、25%、50%和100%(以双波长紫外光源中UV185完全辐照强度计为100%,约1.3 mW·cm−2[14])的双波长紫外灯获得,采用仅辐照UV254(即UV185辐照强度为0%,型号:10-08100)的单波长紫外灯作为双波长紫外的对比光源。初始pH采用浓度为0.1 mol·L−1的盐酸和氢氧化钠调节。实验中将预定质量浓度的有机染料溶液加入反应器中,待溶液温度稳定到设定值时开启预热过的双波长紫外灯,依设定好的时间间隔取样并进行相应指标检测。
1.3 分析方法
有机染料的浓度采用紫外-可见光分光光度计(SPECORD® 200 PLUS,德国耶拿)进行测定,MB、AO7和RhB的特征吸收波长分别为664、485和554 nm,比色皿光程为1 cm。纯水体系中H2O2的浓度采用改进的KI法测定[15]:分别配置试剂A(0.1 mol·L−1邻苯二甲酸氢钾)和试剂B(0.4 mol·L−1碘化钾、0.06 mol·L−1氢氧化钠和0.1 mmol·L−1钼酸铵);测定过程中取水样3.0 mL,然后加入1.5 mL试剂A和1.5 mL试剂B,混合均匀后静置3 min,然后采用1 cm光程比色皿在波长352 nm处测定H2O2浓度。有机染料溶液的pH采用台式酸度计(FE28,瑞士梅特勒-托利多)测定。
2. 结果与讨论
2.1 UV185辐照强度的影响
以UV185/UV254为光源,在有机染料初始质量浓度为30 mg·L−1、反应温度为25 ℃和不调节原水pH的条件下,考察了不同UV185辐照强度对有机染料降解效能的影响,结果如图2(a)~(c)所示。UV254单波长紫外辐照条件下MB、AO7和RhB在60 min后去除率分别为15.3%、24.0%和51.1%。当有UV185存在时,3种有机染料的去除率明显增加。在UV185辐照强度为100%的条件下,水溶液中的MB、AO7和RhB在反应60 min后的去除率分别达到100.0%、99.0%和100.0%。3种有机染料的紫外光降解过程均符合伪一级反应动力学模型,动力学参数列于表1中。如图2(d)所示,UV185辐照强度从10%增加到100%时,MB、AO7和RhB的降解反应速率常数分别升高了0.13、0.08和0.36 min−1。3种有机染料的降解速率均随着UV185辐照强度增加而提高,表明UV185辐照是加速有机染料降解的关键因素。同时,可以看到UV185辐照对3种有机染料降解的加速程度不同,顺序为RhB>MB>AO7,UV185辐照对RhB光降解速率的影响最为显著。
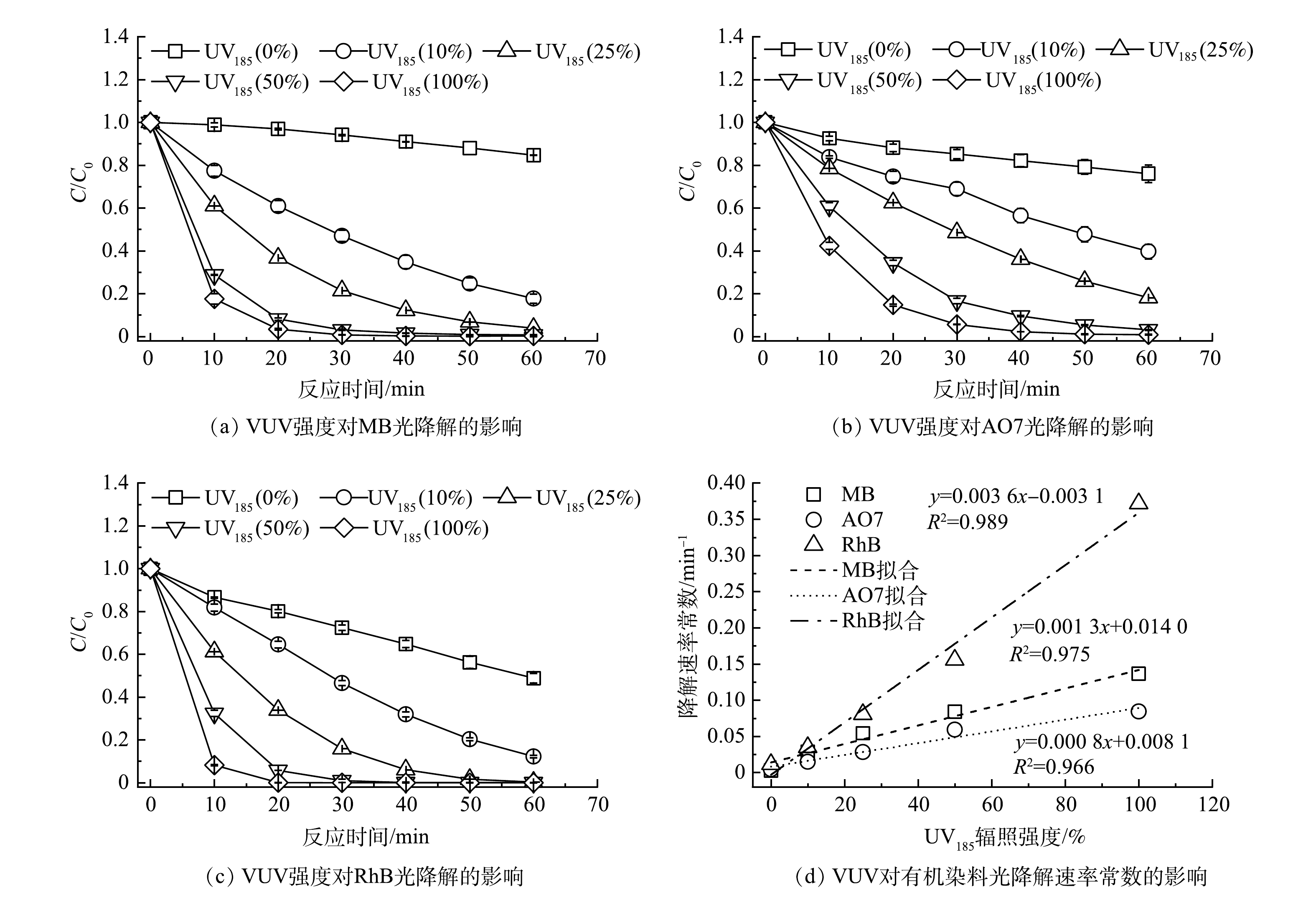 图 2 UV185辐照强度对双波长紫外光氧化降解有机染料效能的影响Figure 2. Effect of UV185 irradiation intensity on photodegradation of organic dyes by dual-wavelength ultraviolet表 1 3种有机染料在不同UV185辐照强度下光氧化降解过程反应动力学参数Table 1. Kinetic parameters of photooxidation degradation of three organic dyes under different UV185 irradiation intensity
图 2 UV185辐照强度对双波长紫外光氧化降解有机染料效能的影响Figure 2. Effect of UV185 irradiation intensity on photodegradation of organic dyes by dual-wavelength ultraviolet表 1 3种有机染料在不同UV185辐照强度下光氧化降解过程反应动力学参数Table 1. Kinetic parameters of photooxidation degradation of three organic dyes under different UV185 irradiation intensity有机染料 不同UV185辐照强度下光氧化降解反应动力学参数 0% 10% 25% 50% 100% k/min−1 R2 k/min−1 R2 k/min−1 R2 k/min−1 R2 k/min−1 R2 MB 0.003 0.970 3 0.029 0.993 8 0.054 0.998 9 0.084 0.942 2 0.136 0.964 9 AO7 0.004 0.975 5 0.015 0.985 7 0.028 0.990 6 0.059 0.998 0 0.084 0.978 9 RhB 0.012 0.991 2 0.035 0.970 3 0.081 0.960 1 0.156 0.985 8 0.372 0.930 7 | Show Table DownLoad:
CSV
DownLoad:
CSV
3种有机染料的降解均包含UV185和UV254 2种紫外光降解过程。实验中所选择的有机染料均具有光敏性,在UV254紫外光辐照下被激发为单重激发态,然后通过系间窜越转化为三重激发态[16-18]。当水中含有溶解氧时,处在激发态的有机染料分子将能量转移给基态的分子氧,生成具有氧化能力的单线态氧(1O2)并将有机染料降解(式(1))。
有机染料∗+O2λ=254nm→有机染料+1O2 (1) 水分子在UV185辐照时可以被直接光解生成·OH(式(2)和式(3))[19],随着辐照强度的增加,体系中·OH的生成量也随之增加。
H2O+UV185→⋅OH+H++e−aqΦ(e−aq)=0.045 (2) H2O+UV185→⋅OH+⋅HΦ(⋅H)=0.33 (3) 有研究表明,MB[20]、AO7[21]和RhB[22]等3种有机染料均可以被·OH攻击而氧化降解,因此,相较于UV254光解,UV185/UV254体系中产生的·OH可加速3种有机染料的降解过程。体系中UV185辐照强度的提高还可以增加水分子光解原生的还原物种·H/eaq−以及次生的活性氧物种HO2·/O2·−,因而可进一步提高有机染料的降解速率。另外,图2(d)表明UV185的辐照强度与有机染料降解速率常数的关系为线性正相关,说明UV185/UV254光降解有机染料过程中UV185光解水是限速步骤。
2.2 有机染料初始质量浓度的影响
以UV185辐照强度为100%的UV185/UV254为光源,在反应温度为25 ℃和原水pH不调节的条件下,考察了不同有机染料初始质量浓度对有机染料降解效能的影响,结果如图3(a)~(c)所示。当MB、AO7和RhB等3种有机染料初始质量浓度分别为10、30、50和70 mg·L−1时,降解速率随着初始质量浓度的升高而降低。当有机染料质量浓度为10 mg·L−1时,MB、AO7和RhB等3种有机染料分别经过7.5、17.5和10 min即可完全去除。而有机染料初始质量浓度增加到70 mg·L−1时,MB、AO7和RhB等3种有机染料在反应30 min后的去除率分别为81.9%、53.4%和90.3%。不同初始质量浓度条件下有机染料的降解过程均符合伪一级反应动力学模型,动力学参数列于表2中。不同有机染料初始质量浓度条件对MB、AO7和RhB等3种有机染料降解速率影响规律一致,降解速率均随着初始质量浓度的增加而降低,即较低的初始质量浓度有利于有机染料的光氧化降解。
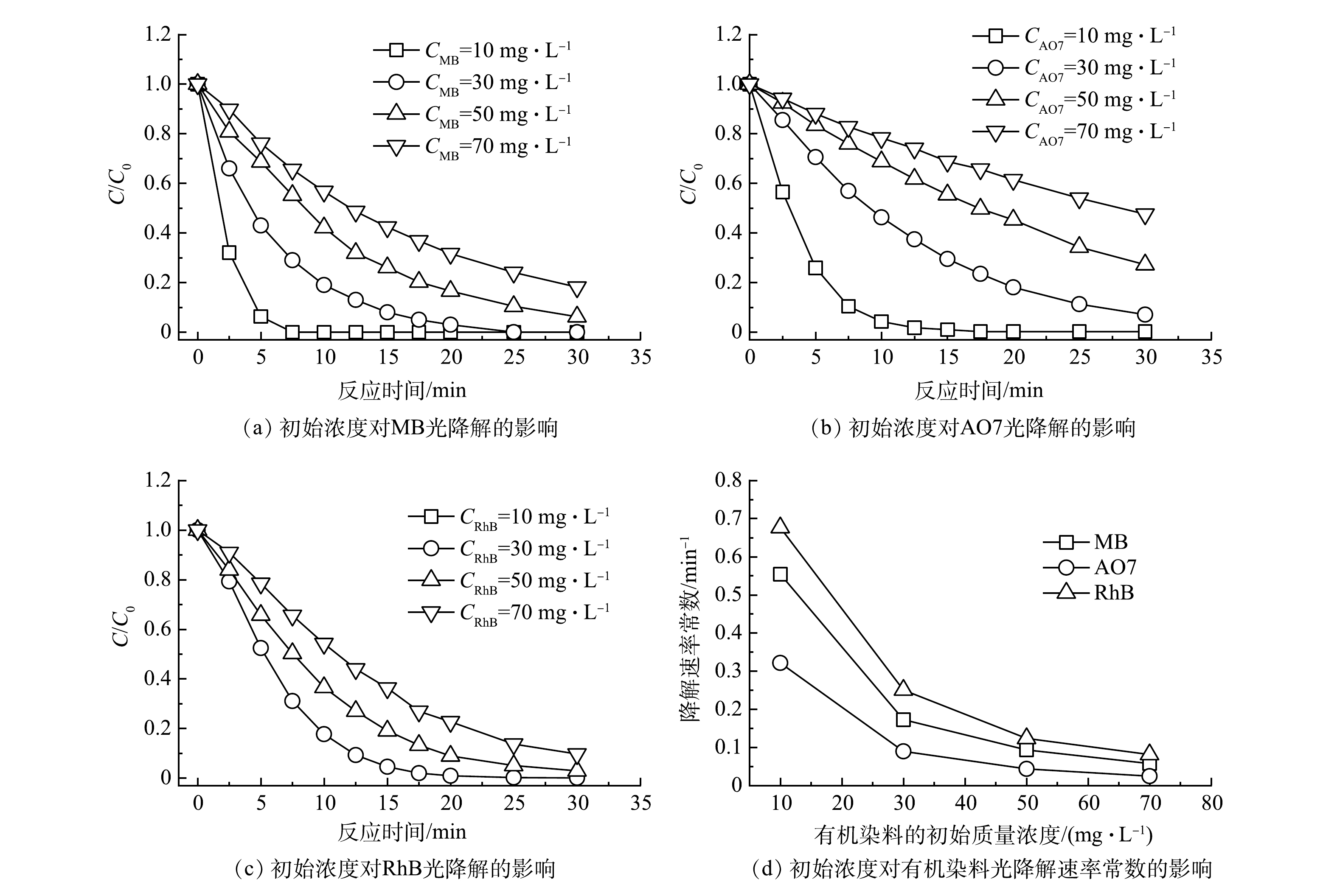 图 3 有机染料初始质量浓度对双波长紫外光氧化降解有机染料的影响Figure 3. Effect of initial concentration of organic dyes on photooxidative degradation表 2 3种有机染料在不同初始质量浓度下光氧化降解过程反应动力学参数Table 2. Kinetic parameters of photooxidation degradation of three organic dyes at different initial concentrations
图 3 有机染料初始质量浓度对双波长紫外光氧化降解有机染料的影响Figure 3. Effect of initial concentration of organic dyes on photooxidative degradation表 2 3种有机染料在不同初始质量浓度下光氧化降解过程反应动力学参数Table 2. Kinetic parameters of photooxidation degradation of three organic dyes at different initial concentrations有机染料 不同有机染料初始浓度下光氧化降解反应动力学参数 10 mg·L−1 30 mg·L−1 50 mg·L−1 70 mg·L−1 k/min−1 R2 k/min−1 R2 k/min−1 R2 k/min−1 R2 MB 0.554 0.978 8 0.173 0.998 1 0.093 0.998 3 0.058 0.999 6 AO7 0.321 0.995 3 0.090 0.997 3 0.043 0.995 4 0.025 0.999 5 RhB 0.676 0.945 7 0.251 0.985 4 0.123 0.995 1 0.082 0.991 6 | Show TableDownLoad:
CSV
初始质量浓度对有机染料光氧化降解效能的影响机制可以从以下3个方面来讨论。首先,较高浓度的有机染料分子间对活性物种产生更强的竞争反应。在相同光源条件下发射的光子总量是固定的,在无有机染料存在的情况下,通过光辐照产生的氧化活性物种数量也是固定的。由前节可知,UV185/UV254体系中UV185光解水反应是有机染料光降解的限速步骤,即活性物种生成量相对有机染料是缺乏的。当体系中有机染料存在时开始消耗氧化活性物种,其数量显著超过氧化活性物种时就开始出现对活性物种的竞争。较高浓度的有机染料降解也会导致更多中间产物的生成,加剧了对活性物种的竞争。其次,有机染料及其降解产物不仅对活性物种具有竞争作用,对紫外光同样具有类似竞争作用的屏蔽作用。以MB为例,在185 nm和254 nm处MB的摩尔吸光系数分别为7 229 L·(mol·cm)−1和5 562 L·(mol·cm)−1[16],对2种波长的紫外均具有强吸收。因此,随着MB初始浓度的升高,越来越多的紫外光会被面向光源的MB屏蔽,导致背向光源的MB和水分子光解速率降低。如图3(d)所示,有机染料的初始质量浓度由10 mg·L−1升高到70 mg·L−1,MB、AO7和RhB的降解反应速率常数分别下降了0.50、0.30和0.59 min−1,可见初始质量浓度对RhB的降解速率影响最大。这是由于直接光解作用对RhB的贡献较大,低初始质量浓度条件下引起的光屏蔽作用更低而有利于RhB的直接光降解。最后,真空紫外光在水中的辐照范围有限,有机染料分子向有效辐照区域的传质成为限制因素。UV254在水中辐照的距离相对较长,超过40 mm,然而UV185有效辐照距离仅为5.5 mm[23]。UV185辐照对有机染料的降解有着更重要的作用,但辐照距离的限制使得有效反应局限于光源表面。因此,在UV185/UV254体系中,有机染料分子向UV185辐照区域传输的过程成为限速步骤之一,在更高的有机染料初始浓度下表观降解速率降低。
2.3 初始pH的影响
初始pH是影响光降解反应最重要的参数之一,研究其对有机染料的降解效率的影响有重要意义。以UV185辐照强度为100%的UV185/UV254为光源,在有机染料初始质量浓度为30 mg·L−1和反应温度为25 ℃条件下,考察了不同初始pH对3种有机染料光氧化降解效能的影响(图4所示)。不同初始pH下MB、AO7和RhB等3种有机染料在光降解反应30 min后均可以基本去除,去除率为91.9%~100.0%,且初始pH对降解速率没有显著影响。这表明在本实验条件下UV185/UV254光氧化有机染料过程可以适应较宽的pH范围,有更强的水质适应性。
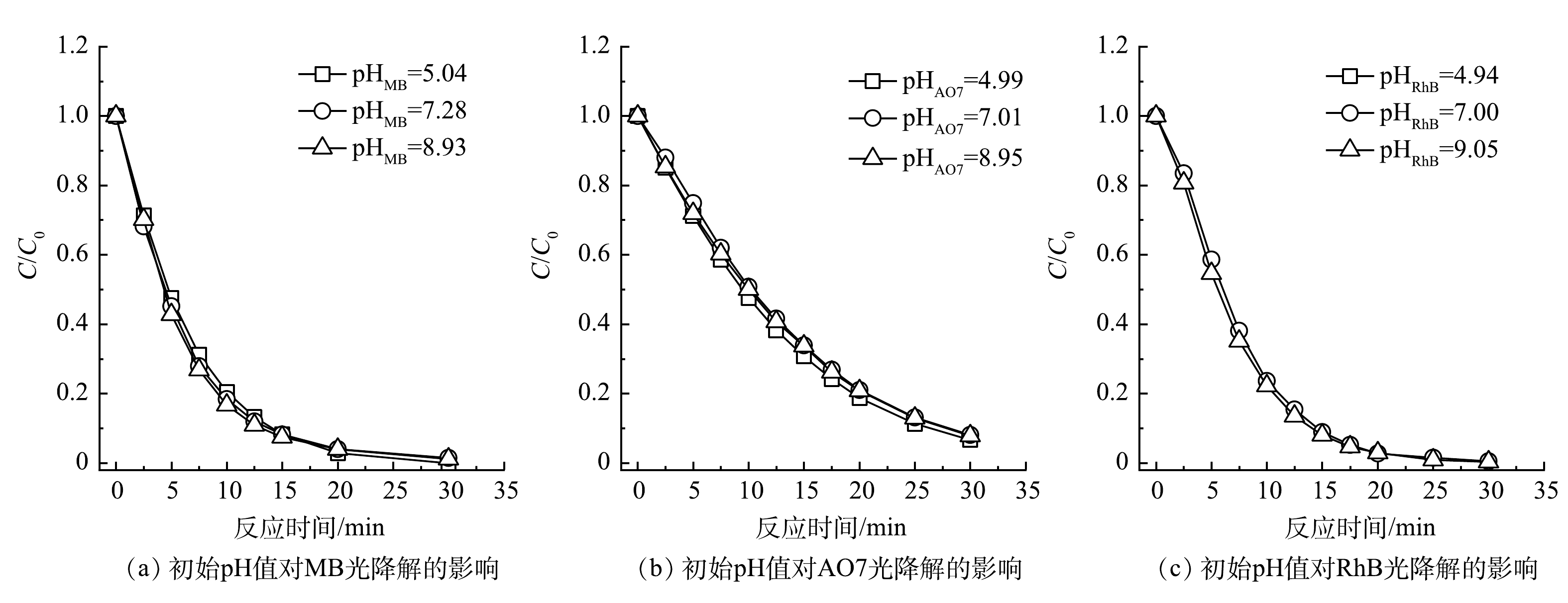 图 4 初始pH对对双波长紫外光氧化降解有机染料的影响Figure 4. Influence of initial pH on the photooxidative degradation of organic dyes
图 4 初始pH对对双波长紫外光氧化降解有机染料的影响Figure 4. Influence of initial pH on the photooxidative degradation of organic dyes在碱性条件下水中的OH−与·OH可以发生淬灭反应(式(4))[24],当水中存在大量OH−时,·OH被快速消耗导致其氧化攻击有机染料的概率减小。另外,·OH在不同pH下具有不同的氧化活性,在酸性和碱性条件下·OH的标准电位分别为2.8 V和1.55 V(vs NHE),这也可导致污染物降解速率有所不同[25]。GU等[26]认为,·OH是VUV光辐照降解三氯乙烷主要的氧化活性物种,并发现其降解速率受溶液的酸碱性影响显著,在pH为3的条件下最高,在pH为11的条件下最低。在本研究中,不同初始pH对有机染料的光氧化降解速率没有显著影响,表明除了·OH之外还存在其他活性物种或降解作用贡献于有机染料的降解。
⋅OH+OH−→⋅O−+H2Ok=1.3×1010L⋅(mol⋅s)−1 (4) 2.4 有机染料降解过程的UV-Vis吸收光谱分析
为了探明3种有机染料在双波长紫外光降解过程中的反应机制,分别对UV254和UV185/UV254光辐照条件下的MB、AO7和RhB溶液进行了UV-Vis吸收光谱分析。如图5(a)~(f)所示,MB、AO7和RhB分别在波长为664、485和554 nm处具有特征吸收峰。在UV254辐照条件下,3种有机染料的图谱没有显著变化,特征峰强度随光辐照时间的延长稍有降低,表明UV254单波长直接光解难以有效破坏有机染料分子结构(图5(a)~(c))。UV185/UV254光辐照时,3种有机染料的图谱均发生了较大变化且特征峰强度均有大幅降低,表明有机染料分子结构遭到破坏(图5(d)~(f))。对于MB的UV-Vis吸收光谱,600~700 nm的吸收峰可归因于包含共轭π体系的发色团,而292 nm处的吸收峰可归因于芳香结构[27]。在UV254单波长紫外辐照作用下,600~700 nm的吸收峰强度略有降低,这可能是由直接光解和光敏化产生的活性物种导致的,但这种作用对MB的降解贡献不大。而在UV185/UV254光辐照时,MB的UV-Vis吸收光谱两处的吸收峰强度均显著降低,表明发色基团和芳香环结构被UV185光辐照产生的活性物种攻击而破坏。AO7的特征吸收峰321 nm和242 nm分别对应分子结构中的萘基和苯基[28]。而485 nm处的吸收峰则对应腙形式的n→π*跃迁[29]。与MB类似,UV254单波长辐照对AO7的降解贡献较少,UV185/UV254光辐照时AO7的分子结构才能够被有效破坏。RhB的UV-Vis吸收光谱有2个主要的特征吸收峰:在554 nm处的特征吸收峰(C=N,C=O基团的跃迁)与染料发色有关;在259 nm处的特征吸收峰与RhB的芳香结构有关[30]。在UV254单波长辐照作用下,RhB在554 nm处的吸收峰强度有明显的降低,表明C=N和C=O基团在UV254单波长直接光解作用下即可破坏,并且对AO7降解的贡献不可忽略。在UV185/UV254光辐照时,RhB的吸收峰强度均显著降低直至完全消失。该区域吸收峰强度的大幅下降,说明RhB的发色基团和芳香结构被强烈破坏。值得注意的是,MB和RhB的特征峰随着光辐照时间的延长发生了蓝移。这是因为MB和RhB的分子结构中的二甲氨基和二乙胺基被破坏。二甲氨基和二乙胺基均为助色基团,当氮原子上的甲基或乙基被破坏时,其助色作用减弱,导致特征峰向短波长的方向移动[31-32]。特征峰的蓝移表明MB和RhB的降解过程发生脱甲基和脱乙基反应。
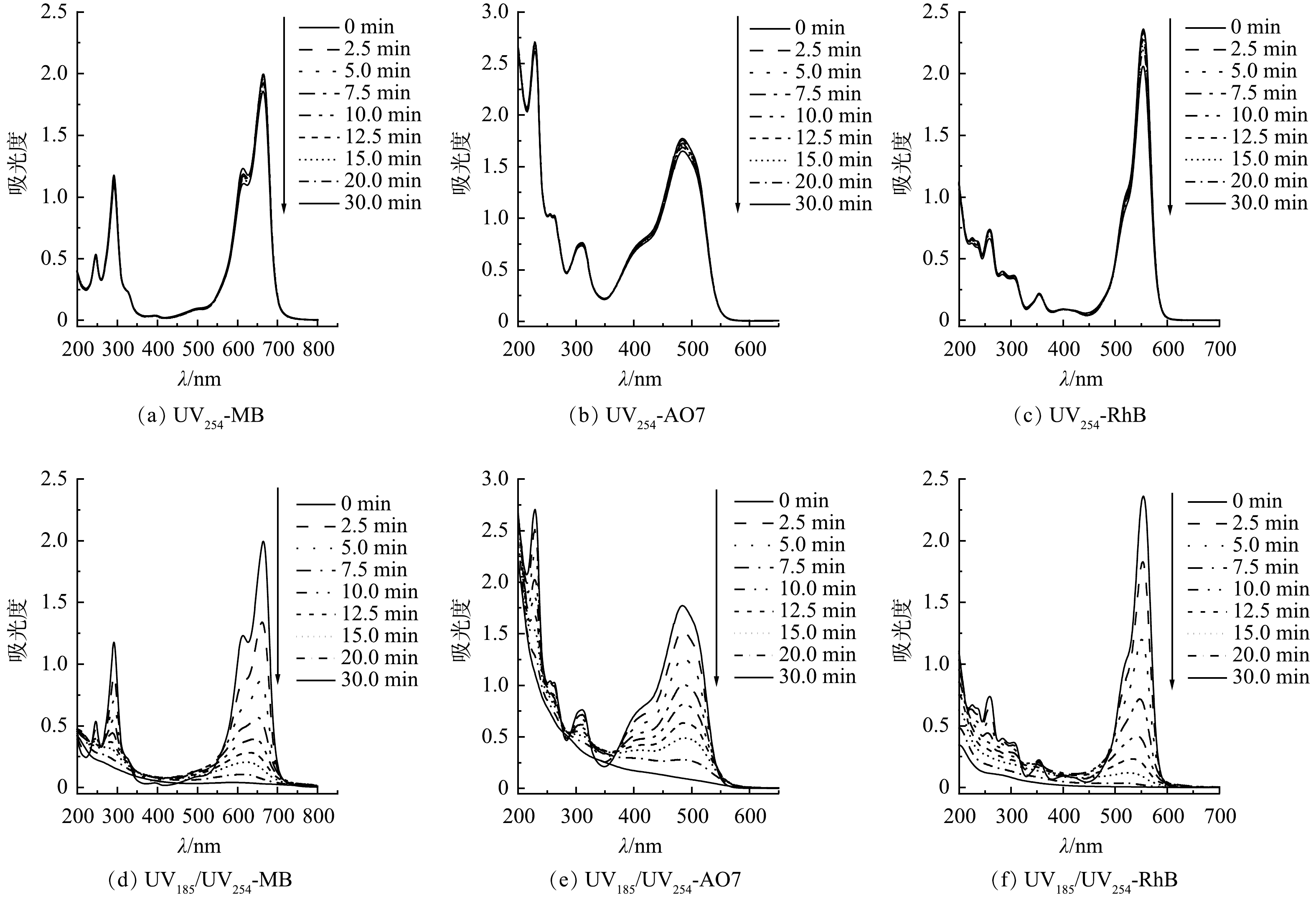 图 5 有机染料分别在UV254辐照和UV185/ UV254辐照下降解过程的UV-Vis吸收光谱Figure 5. UV-Vis absorption spectra of degraded organic dyes under UV254 irradiation or UV185/UV254 irradiation
图 5 有机染料分别在UV254辐照和UV185/ UV254辐照下降解过程的UV-Vis吸收光谱Figure 5. UV-Vis absorption spectra of degraded organic dyes under UV254 irradiation or UV185/UV254 irradiation2.5 双波长紫外光降解有机染料的机制
以UV185辐照强度为100%的UV185/UV254为光源,在纯水中生成的H2O2浓度的变化情况见图6。H2O2的质量浓度随光辐照时间的延长先升高后降低,在光辐照5 min时达到最高,为1.3 mg·L−1。CHEN等[33]指出,在双波长紫外辐照下纯水体系中主要通过的·OH复合生成H2O2,并且水中的溶解氧会提高水中H2O2的稳态浓度。MOUSSAVI等[34]研究表明:在连续流真空紫外光(UV185)反应器中检测到H2O2的质量浓度在4 min时达到约18 mg·L−1。水分子在UV185辐照下生成的·OH经反应后生成了H2O2(式(5)),另外一条H2O2的生成路径(式(7)~式(15))相对反应速率慢且路径更长,因此,式(5)为H2O2主要的生成路径。在UV254存在的条件下,H2O2又会分解产生·OH(式(6)),在反应初期式(6)、式(7)和式(8)等H2O2分解过程的速率低于H2O2生成速率,导致了H2O2浓度升高。随着反应的进行,水分子光解产生的eaq−增多,通过式(10)所示的反应可加速H2O2分解,因而导致光辐照5 min后H2O2的浓度开始降低。
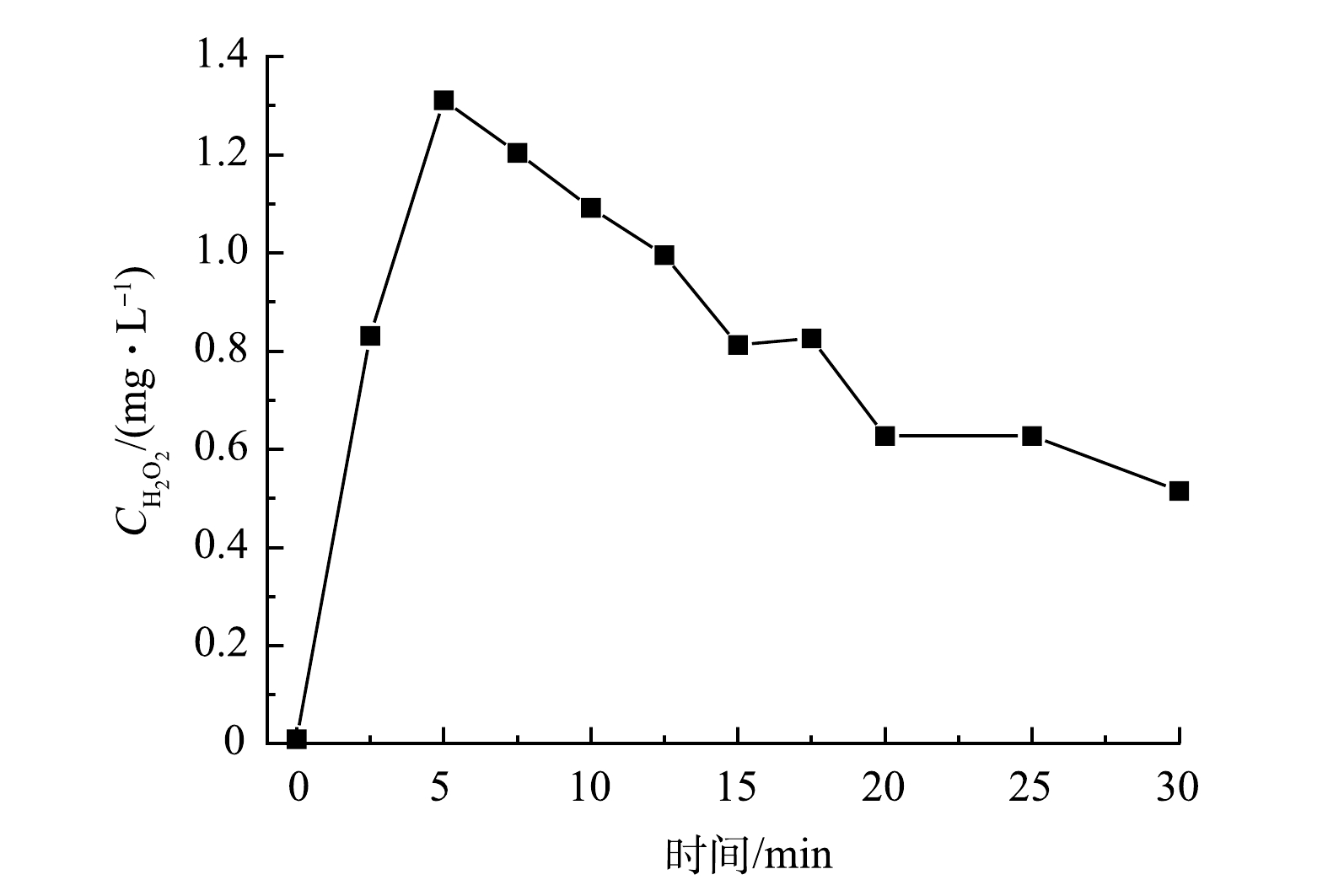 图 6 真空紫外光解水过程中H2O2生成量随光辐照时间的变化Figure 6. Variation of H2O2 production with time in vacuum UV photolysis
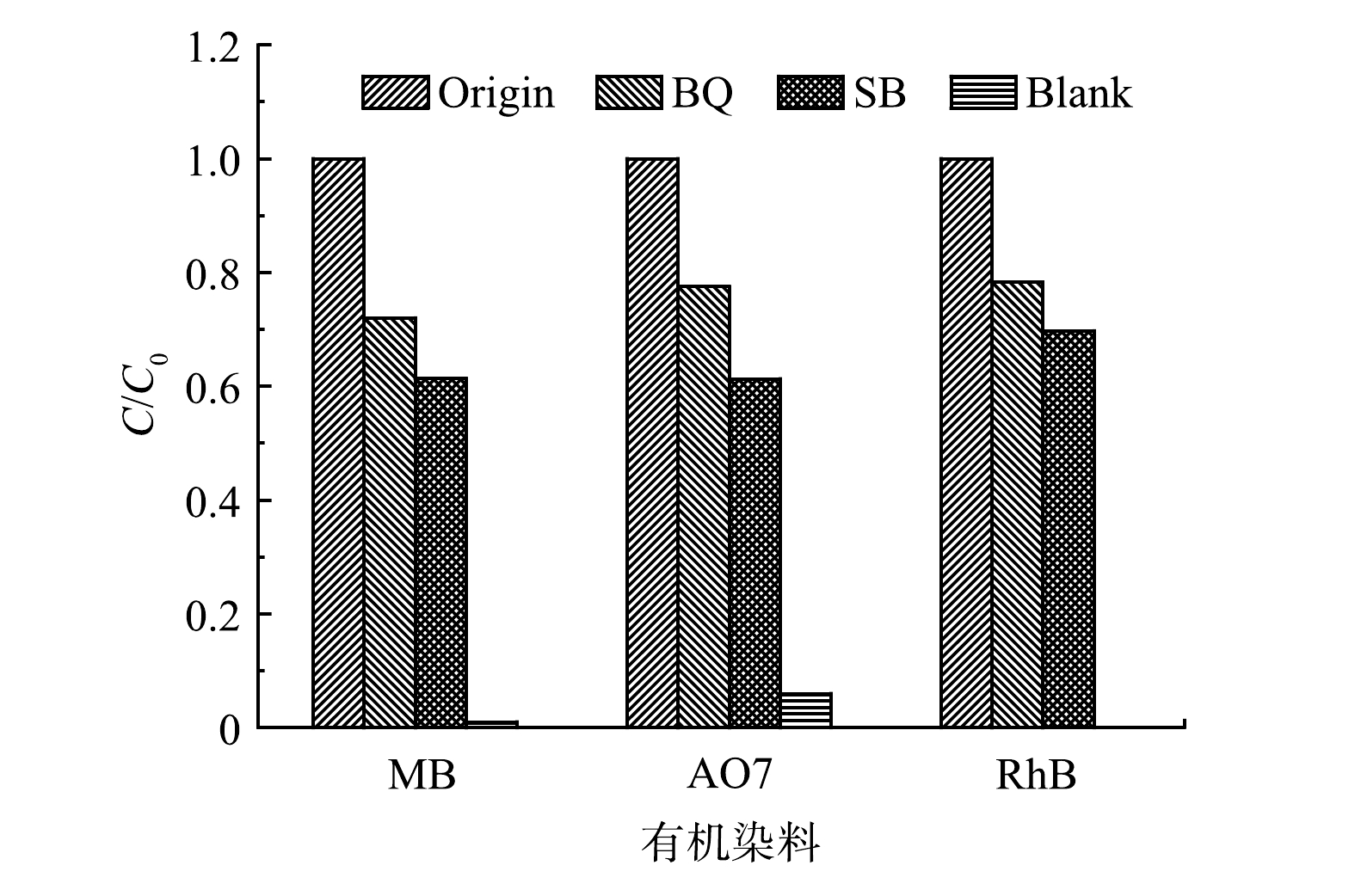
图 6 真空紫外光解水过程中H2O2生成量随光辐照时间的变化Figure 6. Variation of H2O2 production with time in vacuum UV photolysis溶解氧的存在及H2O2的生成使得UV185/UV254光降解体系中活性物种的转化更加复杂。为确定UV185/UV254光氧化降解有机染料的活性物种的种类,采用对苯醌(BQ)作为HO2·/O2·−和·OH的淬灭剂,采用碳酸氢钠(SB)作为·OH的淬灭剂。如图7所示,在加入SB后,MB、AO7和RhB的去除率均大幅降低至38.6%、38.7%和30.3%。这表明·OH在3种有机染料的降解过程中均有显著贡献。在UV185/UV254光氧化体系中加入BQ后,MB、AO7和RhB的去除率分别进一步降至27.9%、22.5%和21.7%,但相较于添加SB体系降低幅度变小。BQ对有机染料的降解抑制作用大于SB,表明除·OH外还存在HO2·/O2·−的作用,但·OH的作用更大。
·OH和HO2·的寿命分别为2 ns和0.38 s[16],虽然在反应体系中HO2·存在的时间更长,但其氧化能力相对·OH更弱。此外,·OH是UV185光解水产生的初级活性物种,而HO2·是经过一系列转化过程产生的次级活性物种,生成量相对·OH更少。由此可以推断,·OH为UV185/UV254光降解有机染料的主要活性物种。
2⋅OH↔H2O2k=5.5×109L⋅(mol⋅s)−1 (5) H2O2+UV254→2⋅OHΦ(H2O2)=0.5 (6) H2O2+⋅OH→HO2⋅+H2Ok=3.3×107L⋅(mol⋅s)−1 (7) H2O2+HO−↔HO−2+H2OpKa=11.6 (8) e−aq+O2→O⋅−2k=1.9×1010L⋅(mol⋅s)−1 (9) e−aq+H2O2→⋅OH+OH−k=1.1×1010L⋅(mol⋅s)−1 (10) O⋅−2+⋅H→HO−2k=2.7×107L⋅(mol⋅s)−1 (11) HO−2+⋅OH→O⋅−2+H2Ok=7.5×109L⋅(mol⋅s)−1 (12) HO−2+⋅OH→HO2⋅+OH−k=6.8×109L⋅(mol⋅s)−1 (13) HO2⋅+⋅OH→H2O+O2k=6.0×109L⋅(mol⋅s)−1 (14) ⋅H+O2→HO2⋅k=1.1×1010L⋅(mol⋅s)−1 (15) 2HO2⋅→H2O2+O2k=8.4×105L⋅(mol⋅s)−1 (16) 基于以上讨论,双波长紫外光降解水中有机染料的机制如图8所示。在UV185/UV254辐照下,有机染料可通过UV254的辐照作用转变为激发态,再与水中的溶解氧反应生成1O2,进而将部分有机染料氧化降解。水分子在UV185的辐照下生成初级活性物种·OH、eaq−和·H,通过他们之间以及水中溶解氧参与的反应,进一步生成HO2·和O2·−等次级活性物种。·OH通过复合反应生成H2O2,然后在UV254的光解作用下再次产生·OH。上述过程生成的·OH、HO2·和O2·−等活性物种通过氧化反应降解有机染料。
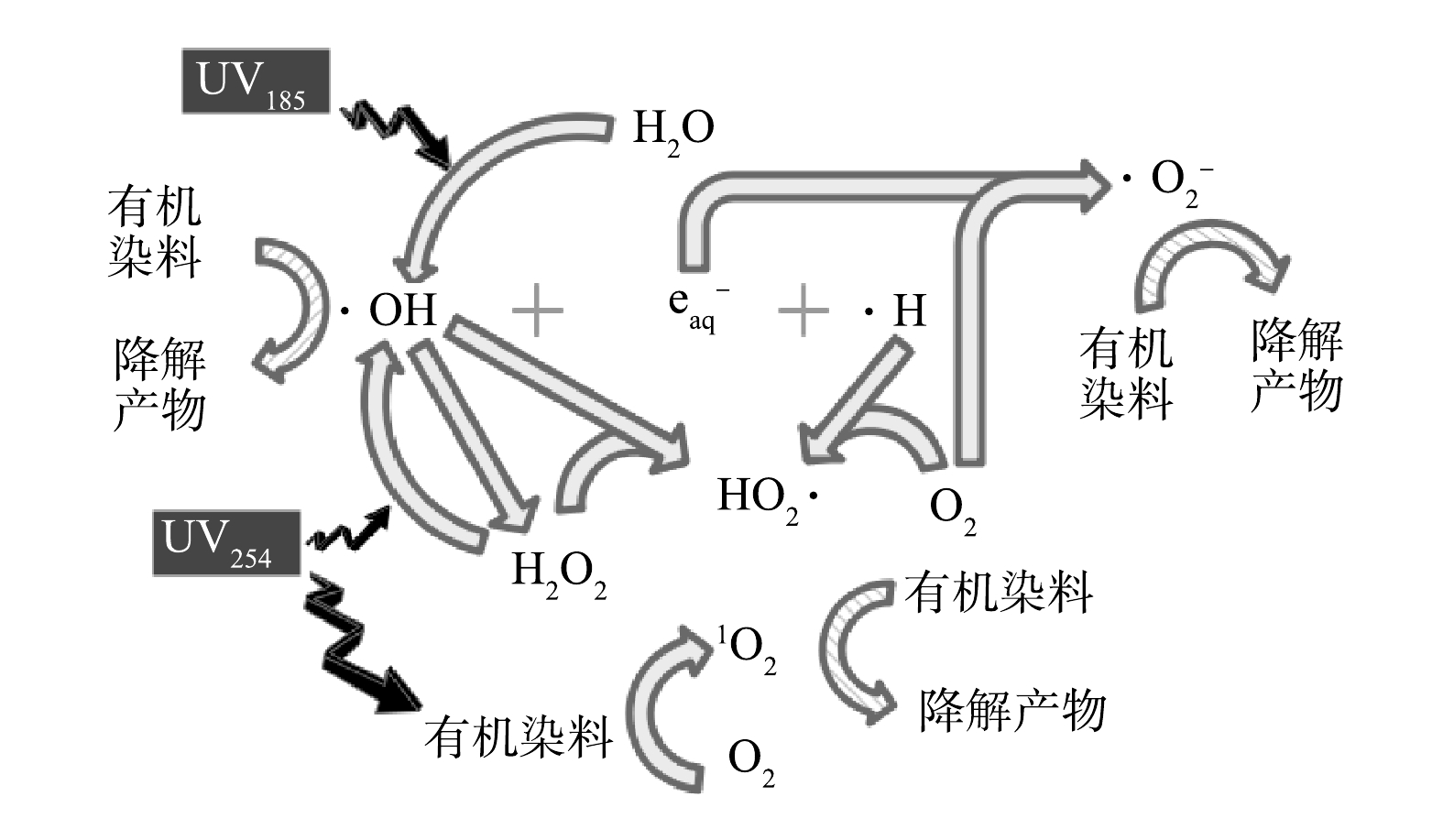 图 8 双波长紫外光氧化降解水中有机染料的机理假设Figure 8. Mechanism of oxidative degradation of organic dyes in water by dual-wavelength ultraviolet light
图 8 双波长紫外光氧化降解水中有机染料的机理假设Figure 8. Mechanism of oxidative degradation of organic dyes in water by dual-wavelength ultraviolet light3. 结论
1) MB、AO7和RhB等3种有机染料在双波长紫外(UV185/UV254)辐照下均可以有效降解,反应60 min后的去除率分别达到100.0%、99.0%和100.0%,并且降解过程均符合伪一级反应动力学模型。
2)高UV185辐照强度和低有机染料初始质量浓度可提高有机染料的降解速率,且有机染料降解动力学常数与UV185辐照强度的关系为线性正相关;初始pH在5~9内时,其数值变化对有机染料的降解速率没有显著影响。
3) UV254对有机染料的光解仅能少量破坏发色基团,UV185光降解有机染料除了可以大幅破坏其发色基团外,还可以深度破坏分子内的芳香结构。
4)有机染料的降解包含UV254直接光解及UV185光辐照产生活性物种间接光解2种机制,其中UV254直接光解对RhB整体降解的贡献大于MB和AO7,达到了51.1%,间接光解的主要活性物种为·OH。
-
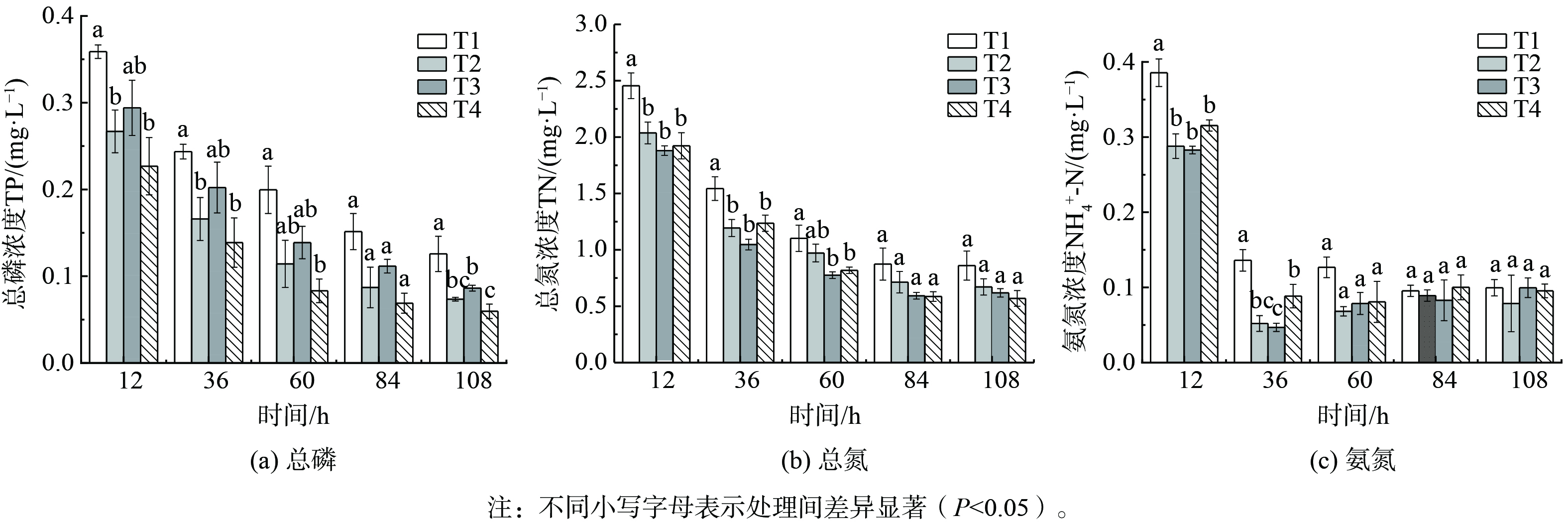
图 3 TP、 TN、 NH4+-N等指标的变化
Figure 3. Changes of the concentration of TP(a),TN(b) ,NH4+-N(c)

图 4 微生物群落在门水平和属水平上相对丰度
Figure 4. Relative abundance of microbial communities at the Phylum level and Genus level
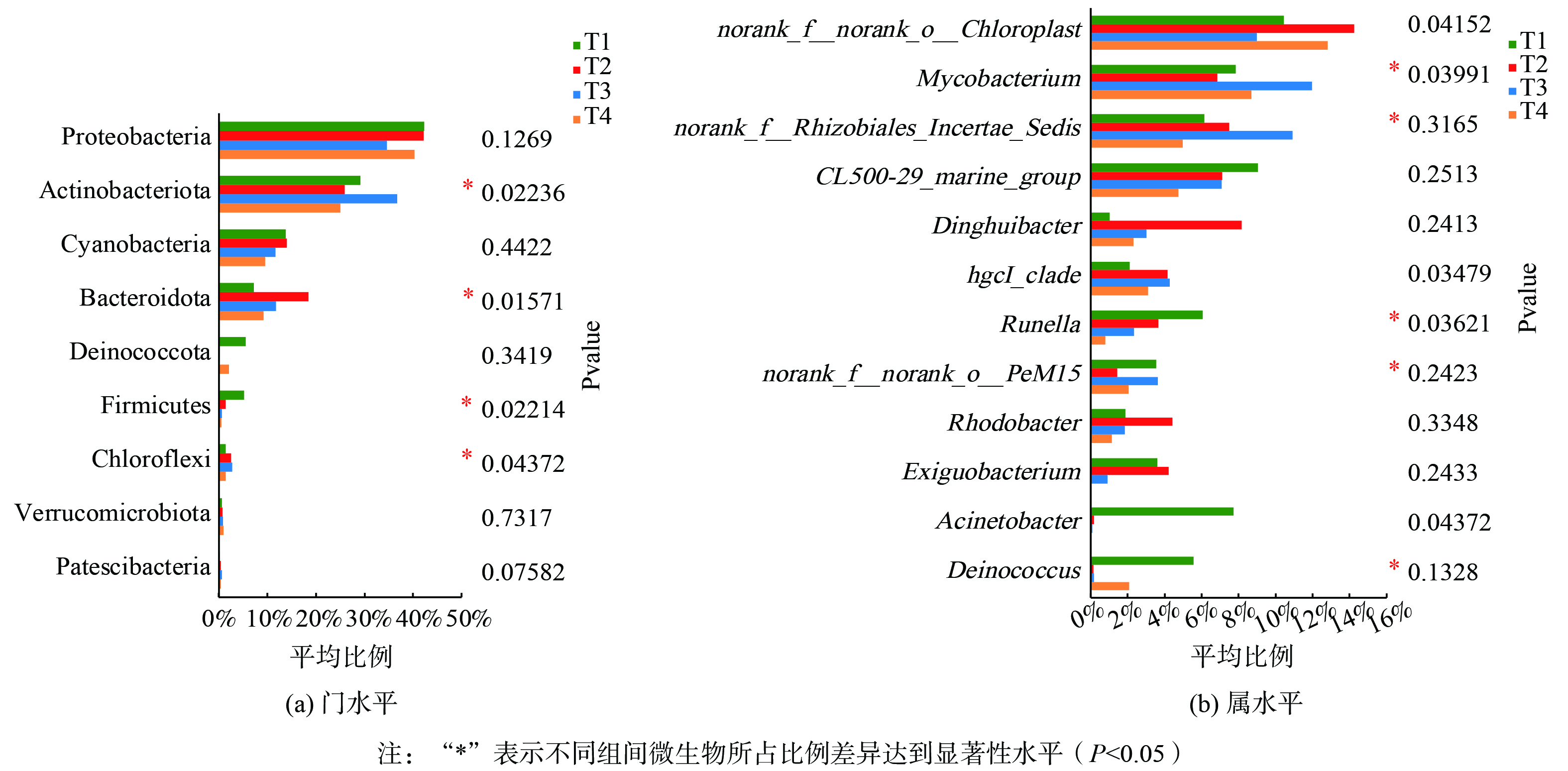
图 5 微生物相对丰度分别在门水平和属水平上的变化
Figure 5. Changes in relative microbial abundance at the Phylum level and Genus level
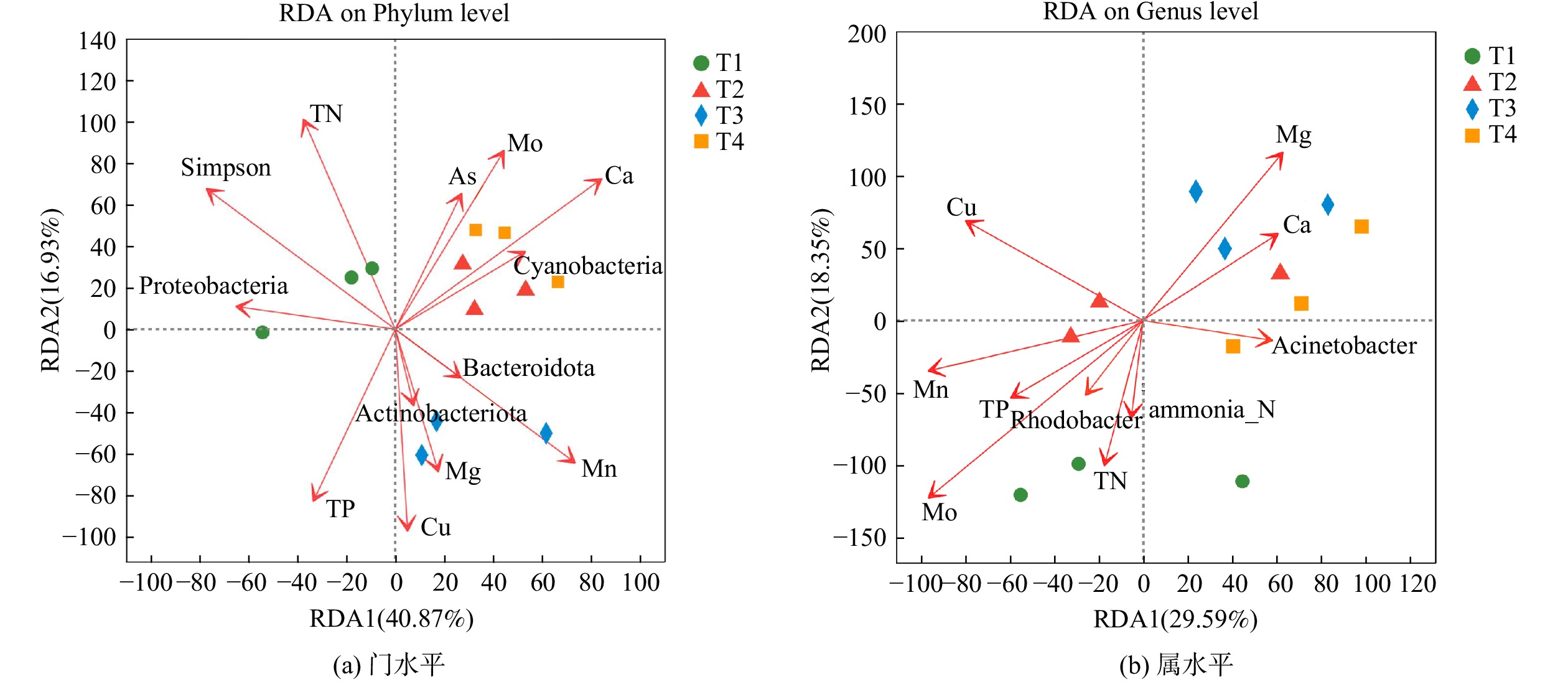
图 6 细菌群落与环境变量在门水平和属水平上的冗余分析
Figure 6. Redundancy analysis of bacterial communities and environmental variables on Phylum level and Genus level
表 1 改造材料基本性状
Table 1. Basic properties of sediment and modified materials
材料 TN/(g·kg−1) TP/(g·kg−1) 有机质/(g·kg−1) SiO2 /(g·kg−1) Al2O3 /(g·kg−1) Fe2O3 /(g·kg−1) CaO /(g·kg−1) MgO /(g·kg−1) pH 石灰石 0.12 2.55 0.57 40.2 9.7 2.1 524.2 9.6 8.3 白云石 0.10 2.07 0.85 6.4 2.7 1.9 575.5 470.3 8.6 蓬莱镇组母岩 0.58 9.43 11.36 653.6 138.1 20.1 27.3 5.8 8.0
下载: 导出CSV
表 2 上覆水体元素含量
Table 2. Elemental content of overlying water bodies
处理 As/(µg·L−1) Cd/(µg·L−1) Co/(µg·L−1) Cu/(µg·L−1) Fe/(µg·L−1) Hg/(µg·L−1) Mn/(µg·L−1) Ni/(µg·L−1) Mo/(µg·L−1) B/(µg·L−1) K/(mg·L−1) Ca/(mg·L−1) Mg/(mg·L−1) T1 15.53±10.25a 13.06±10.57a 8.33±3.03a 5.40±2.79a 2.40±1.90a 15.50±8.41a 3.17±1.08a 18.50±1.27ab 13.37±2.33a 89.40±17.29a 4.11±0.13a 22.95±0.02b 10.13±0.06b T2 12.47±6.52a 15.34±13.33a 0.63±0.63b 4.23±1.19a 2.50±0.50a — 3.50±1.71a 13.63±3.75ab 11.63±2.24a 92.97±4.66a 4.24±0.06a 23.57±0.27ab 10.89±0.26a T3 14.03±5.05a 7.62±7.62a 5.50±0.55ab 5.93±3.02a — — 4.00±1.97a 10.13±4.66b 8.33±2.20a 89.43±15.73a 4.13±0.10a 23.55±0.06ab 11.19±0.09a T4 15.87±6.29a — 6.47±2.47ab 3.80±3.31a 1.70±0.80a — 3.30±1.15a 23.17±1.24a 7.53±3.82a 80.83±5.07a 4.37±0.05a 24.28±0.45a 10.90±0.07a 注:同行不同小写字母表示处理间差异显著 (P<0.05) 。
下载: 导出CSV
表 3 细菌群落多样性指数 (OTU水平)
Table 3. Diversity index of bacterial communities(OTU level)
群落编号 Chao1 Shannon Simpson T1 460 3.83 0.085 6 T2 838 5.14 0.013 4 T3 898 5.01 0.017 8 T4 935 5.17 0.014 5
下载: 导出CSV
-
[1] HUANG S, GUO Q H. Research review on effects of urban landscape pattern changes on water environment[J]. Acta Ecologica Sinica, 2014, 34(12): 3142-3150. [2] 胡洪营, 孙迎雪, 陈卓, 等. 城市水环境治理面临的课题与长效治理模式[J]. 环境工程, 2019, 37(10): 6-15. [3] WANG W H, WANG Y, SUN L Q, et al. Research and application status of ecological floating bed in eutrophic landscape water restoration[J]. Science of the Total Environment, 2020, 704: 135434. doi: 10.1016/j.scitotenv.2019.135434 [4] 范成新, 钟继承, 张路, 等. 湖泊底泥环保疏浚决策研究进展与展望[J]. 湖泊科学, 2020, 32(5): 1254-1277. [5] 张玥, 徐栋, 张义, 等. 引水工程絮凝剂余铝对杭州西湖水体、底泥铝盐分布的影响[J]. 湖泊科学, 2017, 29(4): 796-803. [6] BANALA U K, DAS N P, INDRADYUMNA, et al. Microbial interactions with uranium: Towards an effective bioremediation approach[J]. Environmental Technology & Innovation, 2020: 101254. [7] ZHANG X M, ZHEN W, JENSEN H S. . The combined effects of macrophytes (Vallisneria denseserrulata) and a lanthanum-modified bentonite on water quality of shallow eutrophic lakes: A mesocosm study[J]. Environmental Pollution, 2021, 277. [8] LYONS T, ICKES J A, MAGAR V S, et al. Evaluation of contaminant resuspension potential during cap placement at two dissimilar sites[J]. Journal of Environmental Engineering, 2006, 132(4): 505-514. doi: 10.1061/(ASCE)0733-9372(2006)132:4(505) [9] SADEGHI S, HUA G H, MIN K, et al. Phosphorus and cyanobacteria precipitation and sediment capping in lake water using alum and natural minerals[J]. Journal of Environmental Engineering, 2020, 146(1): 04019095. doi: 10.1061/(ASCE)EE.1943-7870.0001621 [10] BONAGLIA S, RÄMÖ R, MARZOCCHI U, et al. Capping with activated carbon reduces nutrient fluxes, denitrification and meiofauna in contaminated sediments[J]. Water Research, 2019, 148: 515-525. doi: 10.1016/j.watres.2018.10.083 [11] 申粤, 聂煜东, 张贤明, 等. 底泥原位覆盖材料选择及应用研究进展[J]. 环境污染与防治, 2021, 43(7): 898-903. [12] 李雪菱, 张雯, 李知可, 等. 红壤原位覆盖对河流底泥氮污染物释放的抑制研究[J]. 环境污染与防治, 2018, 40(1): 28-32. [13] ZHANG Y, LABIANCA C, CHEN L, et al. Sustainable ex-situ remediation of contaminated sediment: A review[J/OL][J]. Environmental Pollution, 2021, 287: 117333. doi: 10.1016/j.envpol.2021.117333 [14] 郑杰. 城市湖泊清淤工程的探讨[J]. 中国农村水利水电, 2010(9): 43. [15] 冉卓灵, 钟守琴, 刘波, 等. 紫色页岩发育土壤的颗粒特性及其对抗剪强度的作用机制[J]. 土壤, 2019, 51(1): 127-134. [16] 赵吉霞, 邓利梅, 陆传豪, 等. 模拟酸雨淋溶对紫色母岩风化成土特征的影响研究[J]. 西南大学学报(自然科学版), 2021, 43(11): 151-161. [17] 唐将, 李勇, 邓富银, 等. 三峡库区土壤营养元素分布特征研究[J]. 土壤学报, 2005, 42(3): 473-478. [18] 黄雪娇, 刘晓晨, 李振轮, 等. 不同紫色母岩对景观水体氮磷及有机物去除的影响[J]. 环境科学, 2015, 36(5): 1639-1647. [19] 付新喜, 吴晓芙, 奚成业, 等. 农户型组合人工湿地系统生活污水处理效果分析[J]. 给水排水, 2017, 53(7): 25-30. [20] ŽIBIENÈ G, DAPKIENĖ M, KAZAKEVIČIENĖ J, et al. Phosphorus removal in a vertical flow constructed wetland using dolomite powder and chippings as filter media[J]. Journal of Water Security, 2015, 1: 46-52. doi: 10.15544/jws.2015.005 [21] ZHANG J B, WU P X, HAO B, et al. Heterotrophic nitrification and aerobic denitrification by the bacterium Pseudomonas stutzeri YZN-001[J]. Bioresource Technology, 2011, 102(21): 9866-9869. doi: 10.1016/j.biortech.2011.07.118 [22] LIU Z, IQBAL M, ZENG Z, et al. Comparative analysis of microbial community structure in the ponds with different aquaculture model and fish by high-throughput sequencing[J/OL][J]. Microbial Pathogenesis, 2020, 142: 104101. doi: 10.1016/j.micpath.2020.104101 [23] 陈重军, 潘钰伟, 谢嘉玮, 等. 河流污染底泥原位覆盖材料及其应用研究进展[J]. 环境工程技术学报, 2022, 12(1): 100-109. [24] 王磊, 胡效卿, 张卓伦, 等. 不同水深和基质下苦草(Vallisneria natans)的生理生态适应策略[J]. 生态学杂志, 2021, 40(8): 2421-2430. [25] 韩帆, 刘子森, 严攀, 等. 硅酸盐矿物麦饭石对沉水植物生理生态的影响[J]. 水生生物学报, 2019, 43(6): 1353-1361. [26] 张之浩, 吴晓芙, 李威. 沉水植物在富营养化水体原位生态修复中的功能[J/OL][J]. 中南林业科技大学学报, 2018, 38(3): 115-121. [27] SUNIL K R, LOREN G, GUNAWAN G, et al. A Comprehensive review of phosphorus removal technologies and processes[EB/OL][J]. Journal of Macromolecular Science, Part A, 2014, 51: 6,538-545. [28] SRIVASTAVA J K, CHANDRA H, KALRA S J S, et al. Plant–microbe interaction in aquatic system and their role in the management of water quality: A review[J]. Applied Water Science, 2017, 7(3): 1079-1090. doi: 10.1007/s13201-016-0415-2 [29] KUYPERS M M, HANNAH M K, BORAN K. The microbial nitrogen-cycling network.[J]. Nature Reviews. Microbiology, 2018, 16(5): 263-276. doi: 10.1038/nrmicro.2018.9 [30] 黄雪娇, 李振轮, 冯密, 等. 高效复合处理剂的微观结构及其去除景观水体氮磷的机制[J]. 土壤学报, 2020, 57(1): 100-107. [31] 赵桂瑜. 人工湿地除磷基质筛选及其吸附机理研究[D]. 上海: 同济大学, 2007. [32] 柯德峰. 人工湿地基质的筛选及其除磷机理研究[D]. 武汉: 武汉理工大学, 2016: 28-29. [33] 黄雪娇, 冯密, 李振轮, 等. 紫色母岩作基质或覆盖材料对景观水体氮磷去除效果研究[J]. 土壤学报, 2019, 56(2): 363-373. [34] CHRISTIANSEN N H, ANDERSEN F, JENSEN H S. Phosphate uptake kinetics for four species of submerged freshwater macrophytes measured by a 33P phosphate radioisotope technique[J]. Aquatic Botany, 2016, 128: 58-67. doi: 10.1016/j.aquabot.2015.10.002 [35] 刘小宁, 贾博宇, 申锋, 等. 金属元素改性生物质炭应用于磷酸盐吸附的研究进展[J]. 农业环境科学学报, 2018, 37(11): 2375-2386. [36] CHENG C, ZHANG J, XU J, et al. Enhanced removal of nutrients and diclofenac by birnessite sand vertical flow constructed wetlands[J]. Journal of Water Process Engineering, 2022, 46: 102656. doi: 10.1016/j.jwpe.2022.102656 [37] 甄茜, 蔡婕, 郭行, 等. 微藻在废水脱氮除磷中的应用[J]. 水处理技术, 2017, 43(8): 7-12. [38] 胡愈炘, 张静, 黄杰, 等. 长江流域河流和湖库的浮游细菌群落差异[J]. 环境科学, 2022, 43(3): 1414-1423. [39] 吴晓斐, 何源, 黄治平, 等. 不同处理梯度污水对细菌群落和酶活性的影响[J]. 农业环境科学学报, 2020, 39(9): 2026-2035. [40] GAO Y, WANG C C, ZHANG W G, et al. Vertical and horizontal assemblage patterns of bacterial communities in a eutrophic river receiving domestic wastewater in southeast China[J]. Environmental Pollution, 2017, 230: 469-478. doi: 10.1016/j.envpol.2017.06.081 [41] 张雪梅, 佘跃惠, 黄金凤, 等. 大庆油田聚合物驱后油藏微生物多样性研究[J]. 应用与环境生物学报, 2008(5): 668-672. [42] 李建柱, 侯杰, 张鹏飞, 等. 空心菜浮床对鱼塘水质和微生物多样性的影响[J]. 中国环境科学, 2016, 36(10): 3071-3080. [43] GHYLIN T W, GARCIA S L, MOYA F, et al. Comparative single-cell genomics reveals potential ecological niches for the freshwater ac I Actinobacteria lineage[J]. Isme Journal, 2014, 8(12): 2503-2516. doi: 10.1038/ismej.2014.135 [44] 程豹, 望雪, 徐雅倩, 等. 澜沧江流域浮游细菌群落结构特征及驱动因子分析[J]. 环境科学, 2018, 39(8): 3649-3659. [45] PRATT B, RIESEN R, JOHNSTON C G. PLFA analyses of microbial communities associated with PAH-contaminated riverbank sediment[J]. Microbial Ecology, 2012, 64(3). [46] HAN M F, WANG Y W, ZHANG Y H, et al. Efficiency and mechanism for the control of phosphorus release from sediment by the combined use of hydrous ferric oxide, calcite and zeolite as a geo-engineering tool[J]. Chemical Engineering Journal, 2022, 428: 131360. doi: 10.1016/j.cej.2021.131360 [47] 金树权, 周金波, 包薇红, 等. 5种沉水植物的氮、磷吸收和水质净化能力比较[J]. 环境科学, 2017, 38(1): 156-161. -

 点击查看大图
点击查看大图

计量
- 文章访问数: 3151
- HTML全文浏览数: 3151
- PDF下载数: 104
- 施引文献: 0
