



-
人粪污无害化处理对于疾病预防控制和农村人居环境整治具有重要意义。我国将推进人粪污无害化处理和资源化利用作为乡村振兴和美丽乡村战略的重要实施策略[1]。人粪污中含有大量有机物和营养元素[2],每人每年产生的粪污中约含5.7 kg氮、0.6 kg磷和1.2 kg钾[3],并且没有重金属污染的风险,可有效替代化肥[4]。我国自古一直有将人粪污堆沤还田农用的习惯,除了用作肥料,还可作为土壤改良剂,补充腐殖质[5]。但人粪污中含有大量病原体和大分子有机物质,为了避免粪口传播,在施入农田前必须充分腐熟以降低肠道疾病感染风险[6]。人粪污堆肥经过充分腐熟,可杀灭其中的病原菌以及寄生虫卵,形成含有高有机质以及氮磷钾等营养元素的农家肥,但若堆肥腐熟不充分,人粪污农用则会导致病原体的传播,造成二次污染。
不同地区不同的生活及如厕习惯,导致人粪污含水率差异较大[7-8],在堆肥过程中物料含水率会影响堆体的氧气运输量,还可以调节发酵温度、物料孔隙率以及微生物活性等[9],是影响堆肥进程及有机质降解率的主要因素[10]。一般认为,堆肥适宜的含水率是55%~65%[11],较低的含水率不利于微生物生长繁殖,易导致堆肥产品未腐熟等问题。较高的含水率会使物料产生团聚,不利于通风,产生局部厌氧,延长堆肥周期。不同物料好氧堆肥的最佳含水率也存在差异。ZAVALA等[12]认为,人粪污和木屑共堆肥含水率大于64%会产生厌氧降解。而PETRICA等[13]研究表明,禽粪和麦秸联合堆肥时的最佳初始含水率为69%。此外,GUO等[9]通过正交实验研究通风量、C/N和含水率对猪粪和玉米秸秆堆肥影响时发现,65%、70%和75%的初始含水率间无显著性差异。
好氧堆肥过程是在不同微生物参与的条件下完成的,含水率会影响微生物生理和代谢活动所需的溶解性营养物质的运输,进而影响微生物生命活动[9]。LI等[14]在研究不同表面活性剂对牛粪和甘蔗渣堆肥有机质降解和微生物群落功能特性影响时发现,含水率是影响堆肥初期细菌群落结构的主要环境因子,并且在整个堆肥过程中,其对细菌群落的影响仅次于纤维素。GUO等[15]以锯末和玉米芯为主要堆肥原料的研究也证实含水率是堆肥初期影响细菌群落的主要因素,他们还发现含水率与真菌群落的相关性大于其他环境因子。GE等[16]在研究初始含水率和pH对牛粪与麦秸共堆肥影响时发现,总氮、pH和含水率对堆体中真菌和细菌群落演替均有显著影响,但含水率对微生物群落差异的贡献度低于总氮和pH。因此,堆肥物料不同,含水率对堆肥过程中微生物群落的影响也不尽相同。
目前,初始含水率对好氧堆肥腐熟及微生物群落结构影响的研究多以畜禽粪污为原料,以人粪污为原料的研究鲜见报道。本研究借鉴畜禽粪污堆肥工艺研究结果,以人粪污为原料,选择含水率作为好氧堆肥关键影响因子,从堆肥物料的基本理化性质、养分质量分数、无害化效果3方面对堆肥产品进行评价,并进行微生物群落结构变化分析,探究初始含水率对人粪污好氧堆肥腐熟及微生物群落结构的影响,了解初始含水率对人粪污堆肥进程的影响,为优化人粪污好氧堆肥工艺参数提供参考,促进人粪污无害化处理和资源化利用。
-
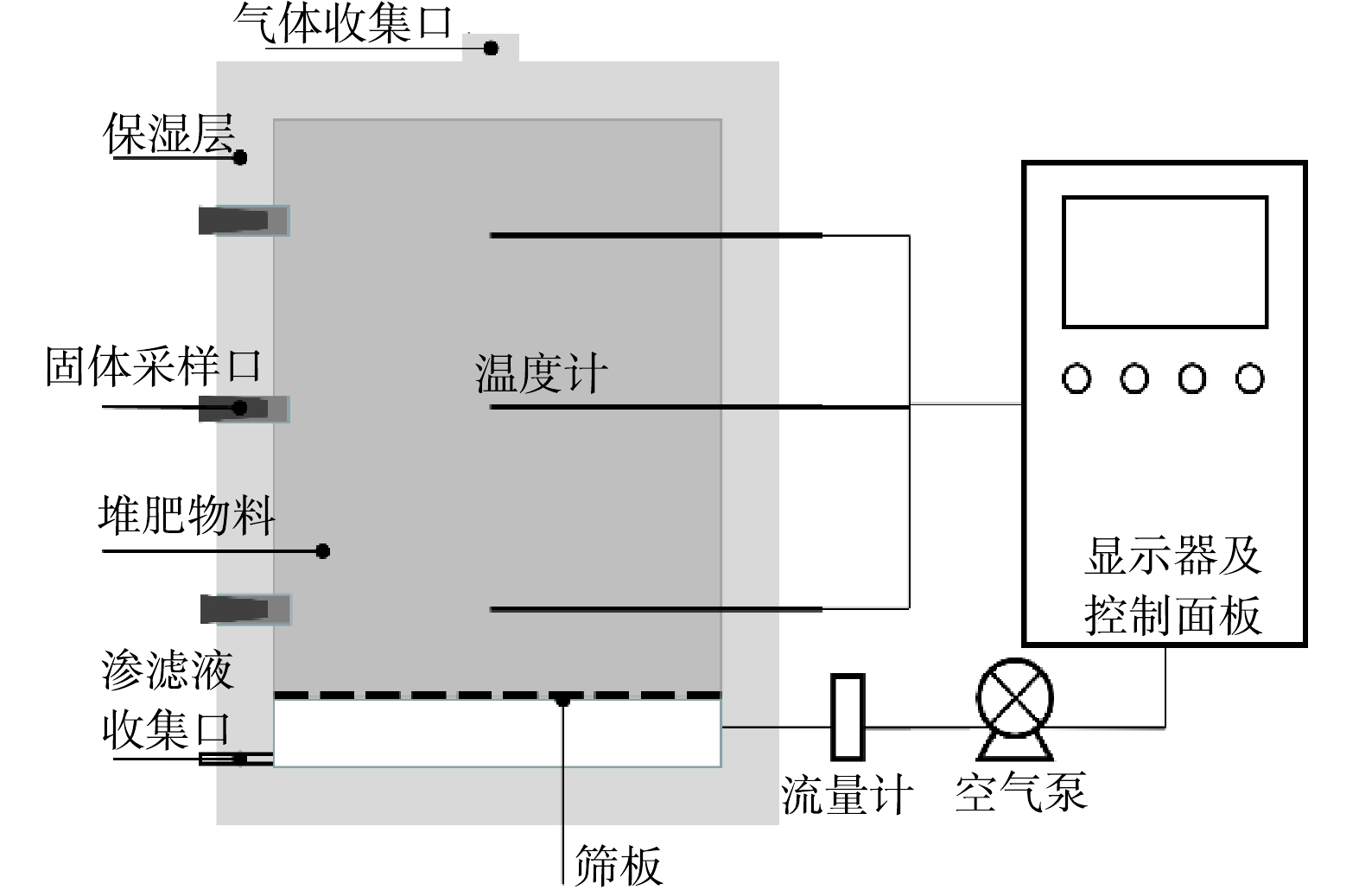
本研究使用的人粪污取自北京市顺义区某农村旱厕,调理剂为2 cm长的玉米秸秆。原料性质见表1。本研究中使用的堆肥发酵装置参考文献[17],该装置有效容积为60 L,由发酵罐罐体、通风控制系统、温度监测系统、空气泵等部分组成,可实时记录堆体温度,控制堆体通风量。罐体顶部设有1个通风口,侧面设有上、中、下3个取样口 (图1) 。
-
以人粪污为原料,选择初始含水率为关键影响因子,设置含水率 (MC,Moisture Content) 为55% (MC55) 、60% (MC60) 和65% (MC65) 3个处理组,开展实验。用玉米秸秆调节含水率与碳氮比 (C/N) ,实验设计如表2所示。曝气方式采用间隔曝气,隔45 min曝气5 min,曝气量为0.7 L·min−1·kg−1[16],实验周期为30 d。待堆体温度降至室温时进行翻堆。每2 d采集1次样本,每次在堆体的上、中、下均匀采样共500 g,并分为3部分,一部分储存在4 ℃冰箱用于测定MC、pH值、电导率 (EC,Electrical Conductivity) 、种子发芽指数 (GI,Germination Index) ;一部分晾干后用于测定总碳 (TC,Total Carbon) 、总氮 (TN,Total Nitrogen) 、总磷 (TP,Total phosphorus) 、总钾 (TK,Total potassium) ;另一部分储存在-80 ℃超低温冰箱用于粪大肠菌群数和微生物群落结构检测,取升温期、高温期和腐熟期-80 ℃保存样品进行微生物群落结构测定。
-
堆体温度由发酵罐自带的PT100温度探头测定,每天上午9点,下午3点进行记录。MC、pH值、EC、GI测定方法参考文献[18],TC、TN质量分数用元素分析仪 (Flash-2000,美国赛默飞世尔科技公司) 测定,TP、TK质量分数用电感耦合等离子体原子发射光谱仪 (Agilent 7800,美国安捷伦科技有限公司) 测定,粪大肠菌群数按照GB/T 19524.1-2004《肥料中粪大肠菌群的测定》[19]的方法测定。样品DNA提取及PCR扩增参照文献[20]的描述,委托第三方实验室在其Miseq平台进行16s rRNA和ITS深度测序。
-
1) 基本理化性质。温度是反应堆肥过程的一个直观参数。3个处理组温度变化如图2所示。各处理温度变化趋势相似,均出现2个高温期 (>50 ℃) ,但每个高温期最高温度与维持时间有明显不同。堆肥初期,3个处理组温度均迅速攀升,在第2 d达到第1个高温期,MC55、MC60和MC65处理组高温期分别持续1、2和5 d,最高温度分别为54.0、67.2和67.1 ℃。这表明,较高的含水率更加适合堆肥初期微生物的生长繁殖,有助于人粪便好氧堆肥启动。随后堆体温度开始下降,降至室温后进行了翻堆,MC55和MC60处理第8天翻堆,MC65处理第14 d翻堆。由于翻堆增加了物料与氧气的接触面积[10],进一步优化了微生物的生存环境[21],各处理翻堆后迎来第2个高温期,MC55、MC60和MC65处理组第2个高温期分别持续4、1和0 d ,最高温度分别为61.0、50.1和35.9 ℃。MC55处理的温度变化与HASHEMI等[7]的研究存在差异,这可能是由于堆体含水率较低,不适合微生物生长繁殖。由于前期大部分有机质已被降解,MC60和MC65处理第2个高温期持续时间均较短。堆肥后期,随着可降解有机物质的不断消耗,微生物活性逐渐减弱,堆体温度逐渐降低至室温。整体而言,各处理高温期均大于2 d,均已达到《粪便无害化卫生要求》 (GB 7959-2012) [22],MC65处理高温期持续时间最长,表明初始含水率65%更适合微生物的新陈代谢,有机质降解更充分[10]。
水分不仅参与微生物的生长繁殖,还影响着堆肥效果和理化性质的变化[23]。3个处理堆肥过程中MC变化如图3所示。3个处理组的MC均呈波动下降最终趋于稳定的趋势。3个处理在翻堆后MC均有所升高,由于在翻堆时观察到堆体表层有大约1 cm厚的物料明显比其他物料更加潮湿,因此推测MC的升高是翻堆造成的。MC65处理MC在前12 d下降幅度较大,这是因为该处理第1个高温期持续时间最长,蒸发带走了大量水分。由于MC55处理第2个高温期持续时间较长,其含水率在翻堆后才表现出明显下降。堆肥结束时,3个处理堆体MC均在45%左右,MC55、MC60和MC65处理组含水率较初始时分别下降了11.64%、12.9%和18.91%。
堆肥过程中微生物生长的最佳pH是6.7~9.0[24]。不同初始含水率人粪污好氧堆肥过程pH变化如图4所示。3个处理组pH值在升温期和高温期有明显不同,MC55处理pH值呈先下降后波动上升的趋势,而MC60和MC65处理pH值呈先升高后降低的趋势。产生这种差异的原因可能是堆肥初期MC55处理可被微生物利用的有机物质量分数高于其他处理,微生物分解有机物产生有机酸的速率高于含氮有机物分解产生铵态氮 (ammonium nitrogen,AN) 的速率,造成了有机酸积累[25],而MC60和MC65处理由于温度较高,加速了有机酸的分解和挥发,导致了pH的升高[26]。因为MC65处理高温期较长,NH3挥发较多,导致了MC65处理pH下降速率高于MC60处理[26]。堆肥后期,随着温度的下降,硝化作用增强,3个处理pH也出现不同程度的下降[16]。在堆肥反应结束时,3个处理的pH无显著性差异 (p>0.05)。
EC是反应堆体盐分的指标,与堆肥样品浸提液中离子总浓度呈正相关。不同初始含水率人粪污好氧堆肥过程EC变化如图5所示。MC55和MC60处理EC均呈先升高后降低,最终趋于稳定的趋势。由于微生物将有机物分解为小分子物质,堆体的离子浓度升高,造成堆肥初期EC值升高。随着NH4+以NH3形式挥发和腐殖质的形成,EC值逐渐降低[27]。由于MC65处理堆肥初期高温加速了NH3挥发[10],EC值呈现降低趋势,随着温度下降,NH3挥发减少,微生物的分解作用依然存在,EC值逐渐升高,并趋于稳定。堆肥结束时,3个处理组间EC值无显著差异 (p>0.05) ,MC55、MC60和MC65处理组的EC值分别为3.81、3.68和3.33 mS·cm−1,均低于4 mS·cm−1,作为肥料施入土壤不会对作物及土壤产生不利影响[28]。
2) 养分质量分数变化。不同初始含水率人粪污好氧堆肥过程TP、TK变化如图6所示。3个处理组TP、TK质量分数随着堆肥反应的进行均呈增长趋势。这是因为,堆肥过程是微生物不断分解有机物的过程,随着有机物的降解,物料会产生浓缩效应[29]。李英凯等[30]的研究也得到了同样的结论。堆肥结束时,MC55、MC60和MC65处理组TP质量分数较初始时分别增长了44.18%、55.71%和51.33%,TK质量分数分别增长了44.88%、74.87%和69.37%。堆肥结束时,3个处理组TP质量分数无显著性差异 (p>0.05) ,MC60处理TK质量分数显著高于其他2个处理组 (p<0.01) ,MC55和MC65处理组TK质量分数无显著性差异。
不同初始含水率人粪污好氧堆肥过程TC、TN变化如图7所示。由于前4 d微生物活性较高,有机质降解速度快,造成反应初期TC质量分数大幅下降。随着反应的进行,在堆肥后期,可被降解的有机物不断减少,微生物活性逐渐下降,TC质量分数下降幅度减缓,在腐熟期TC质量分数趋于稳定。堆肥结束时,MC55、MC60和MC65处理组TC质量分数较初始时分别下降了21.51%、18.42%和22.79%。这与袁京等[31]的研究结果一致。随着反应的进行,TN质量分数呈先上升后趋于稳定的趋势,反应结束时,3个处理组间TN质量分数无显著性差异 (p>0.05) 。在玉米秸秆堆肥[32]和蔬菜废弃物堆肥[29]的研究中都得到了这样的结果,这些研究认为这是物料浓缩效应造成的。另有研究结果表明,堆肥过程会造成氮损失,且初始含水率在60%~65%时,TN损失相对较高[31],而本研究中随着初始含水率的增加,TN增长量也在增加。产生这种异同情况的原因需在以后的研究中进一步分析。
3) 无害化效果。GI是反应堆肥腐熟度和作物毒性的指标[9]。不同初始含水率人粪污好氧堆肥过程GI变化如图8所示。初始时,3个处理组GI均低于30%,且初始MC越低,GI越低,表明未经发酵的人粪污具有较高的作物毒性。堆肥初期MC55处理GI缓慢上升, MC60和MC65处理GI升速较快。第4 d时MC60处理GI已经高于80%,达到无害化要求[10]。这表明,MC60更有助于堆肥对人粪污作物毒性的削减,进而提高堆肥腐熟效率[9]。在堆肥结束时,MC55、MC60和MC65处理组GI分别为115.52%、80.05%和92.14%,均达到《畜禽粪便堆肥技术规范》 (NY/T 3442-2019) [33]的要求。
粪大肠菌群是体现堆肥产物无害化程度的重要指标。3个处理组粪大肠菌群随时间变化趋势如图9所示。MC55处理粪大肠菌群呈先减少后增加又减少,从第18 d又大幅增长的趋势,堆肥结束时,粪大肠菌群数约为808 MPN·g−1;MC60处理粪大肠菌群呈先减少后波动增长,最后又减少的趋势,第30 d粪大肠菌群数降为93 MPN·g−1;MC65处理粪大肠菌群数呈先下降后增长,再下降再增长,最终趋于稳定的趋势,但最高未超过40 MPN·g−1。这表明,MC55和MC60处理降温期堆料适合粪大肠菌群生长,可能是由于粪大肠菌群是耐热菌,可在37~47 ℃的环境中生存[34]。在堆肥结束时,MC60和MC65处理均已达到无害化卫生要求[22],而MC55处理未达到无害化卫生要求,且大肠菌群数高于初始时。SADEGHI等[35]的研究也发现堆肥腐熟期粪大肠菌群再生的现象,他们认为这是高温期持续时间较短造成的。
-
1) 覆盖度和OUT聚类。本实验9个样本共检测出524 316个优质细菌序列,1085 401个优质真菌序列。经uparse聚类后,共产生1 357个细菌 operational taxonomic units (OTUs)和1 189个真菌OTUs,经过抽平处理剩余1 336个细菌OTUs和1 150个真菌OTUs。细菌和真菌分别观察到23和9门,46和35纲,130和92目,227和171科,456和274属。这表明,在人粪污与玉米秸秆堆肥过程中,细菌丰度明显高于真菌丰度。GUO等[15]的研究也得到了这样的结论,他们用桃木屑堆肥,共发现了1 538和383个细菌和真菌OTUs。
用稀释性曲线展现测序数据量合理性及各样本物种数量。如图10所示,各样本稀释性曲线都趋于平稳,表明更多的数据量只会产生少量的新OTUs,说明样本测序数据量合理。如图10 (a) 所示,MC60与MC65处理高温期细菌OTUs低于升温期和腐熟期。其中,MC60处理在腐熟期的细菌多样性高于升温期,而MC65处理腐熟期的细菌多样性低于升温期。这表明,在高温期细菌多样性减少,随着温度的下降,细菌多样性又有所增加,且MC60处理腐熟期样品细菌多样性高于升温期。MC55处理的OTUs数目随堆肥反应的进行呈下降趋势,表明随着反应的进行该处理细菌多样性逐渐降低。如图10 (b) 所示,随着堆肥的进行,MC55和MC65处理真菌OTUs数目逐渐减小,表明这2个处理真菌数量多样性随着反应的进行依次降低。MC60处理OTUs数目先增加后减少,表明在堆肥高温期其真菌多样性最高。WEI等[32]认为,堆肥过程OTUs的波动是堆体环境条件引起的。
2) 门和属水平上物种组成。堆体中优势细菌相对丰度如图11 (a)~图11 (b) 所示。从门水平上看 (图11 (a) ) ,几个样本细菌主要由厚壁菌门、放线菌门、变形菌门和拟杆菌门组成,这4个菌门是公认的堆肥过程的主要细菌菌门[36],它们都参与纤维素和木质素的降解[15]。各样本间各细菌菌门相对丰度存在较大差异。在升温期,变形菌门是MC55处理优势菌门;厚壁菌门是MC60和MC65处理优势菌门。到高温期,MC55处理优势菌门演替为厚壁菌门;MC60处理优势菌门演替为变形菌门;MC65处理优势菌门仍为厚壁菌门,其相对丰度有所增加。厚壁菌门演替为优势菌门是由于其可形成耐热的内生孢子[37],具有较强的热可塑性[38]。到腐熟期,MC55处理优势菌门仍为厚壁菌门;MC60和MC65处理优势菌门为变形菌门,同时,拟杆菌门相对丰度有所增加,MC65处理拟杆菌门相对丰度高于MC60处理。由于拟杆菌门是堆肥过程中木质纤维素的主要分解者[39],因此,较高的含水率加速了腐熟期木质纤维素的分解。在属水平上(图11 (b) ),3个处理不同时期样本间差异很大,这可能是由于堆肥初始时堆体间细菌相对丰度就存在很大差异[40]。在高温期,MC55处理优势菌属为乳酸菌 (Lactobacillus) 、葡萄球菌 (Staphylococcus) 和肠球菌 (Enterococcus) ,这3类菌属主要分解糖类并广泛存在于肠道中[41];MC60处理优势菌属由Carnobacterium演替为Psychrobacter,Psychrobacter能产生参与腐殖化过程的过氧化氢酶[42],表明MC60 处理在堆肥第2 d已经开始腐殖化进程;MC65处理优势菌属由Atopostipes演替为Bacillus,Bacillus可降解纤维素和有害物质[16]。在腐熟期,Saccharomonospora和Bacillus成为MC55处理的优势菌属,2者均可降低腐熟产物的作物毒性[16],这与MC55处理GI在腐熟期才达到无害化要求相吻合; MC60和MC65处理优势菌属均为uncultured和uncultured_bictureium,结合温度和无害化指标变化,推测uncultured和uncultured_bictureium可能是表征堆肥腐熟的菌属。由3个处理不同时期优势均属变化可以发现,高温期MC60和MC65优势菌属不同,并且分别以腐殖化和无害化进程为主,但在腐熟期2个处理微生物群落结构具有相似性。整个堆肥过程中,MC55处理优势菌属均不同于另外2个处理,且无害化进程主要发生在腐熟期。
真菌门和属水平相对丰度如图11 (c)~图11 (d) 所示,在门水平上 (图11 (c) ) ,不同处理不同时期的优势真菌门都是子囊菌门和担子菌门,这与GUO等[15]的研究结果一致。随着堆肥的进行,MC55和MC60处理子囊菌门相对丰度逐渐增加,MC65处理在高温期子囊菌门相对丰度也有所升高,这可能是因为子囊菌门和担子菌门都可以分泌多种纤维素和半纤维素降解酶[43-44],但Basidiomycota对高温敏感,而Ascomycota能很好的适应温度和营养胁迫[15]。担子菌门演替为MC65处理腐熟期优势菌门,这可能是因为腐熟期以木质素等难降解有机物的降解主要发生在腐熟期[45],担子菌门在木质素降解中起着关键作用[46]。在属水平上 (图11 (d) ) ,MC60处理各时期真菌相对丰度变化不大,其中曲霉 (Aspergillus) 相对丰度逐渐升高 (从37.63到42.67%) ,Thermomyces (从39.05%到32.98%) 相对丰度逐渐下降。Aspergillus和Thermomyces都能分解半纤维素[16],Aspergillus可以加速堆肥进程,缩短腐熟期[47]。HERNÁNDEZ-LARA等[40]也认为曲霉是腐熟期的优势真菌。在高温期,MC55和MC65处理优势菌属均为Meyerozyma,Meyerozyma的菌株能分泌多种大分子有机物降解酶[48],并且与富里酸的形成有关[49]。这表明,MC55和MC65处理在高温期有大量富里酸合成。在腐熟期,Melanocarpus成为MC55处理优势菌属;同时,在MC60和MC65处理中也观察到Melanocarpus相对丰度的增加。Melanocarpus可以产生纤维素酶和木聚糖酶,参与富里酸、胡敏素的降解和胡敏酸的形成[49]。因此,MC55处理在第30 d仍处于腐殖化进程。在腐熟期,MC65处理Coprinopsis相对丰度最高,车悦驰[50]在西藏地区污泥堆肥的研究中也发现Coprinopsis在堆肥腐熟期成为优势菌属,但其原因还需进一步研究。
3) 微生物丰富度和群落多样性。Chao1指数是菌种丰富度指数,Shannon指数是用来估算样品中微生物多样性指数之一。不同初始含水率人粪污好氧堆肥过程不同阶段细菌和真菌Chao1和Shannon指数如表3所示。MC55处理细菌和真菌的微生物丰富度与群落多样性都逐渐减少。MC60处理高温期细菌Chao1和Shannon指数低于另外2个时期。这表明,高温期细菌数量与种类都逐渐减少,随着温度的降低细菌数量逐渐恢复至初始水平,细菌群落多样性较堆肥初期还有所增加。随着反应的进行,MC60处理真菌丰富度逐渐减少,但真菌物种均匀度在高温期增加后又随着温度的降低而减小。MC65处理细菌丰富度和多样性变化与MC60处理相同,但真菌丰富度与均匀度都逐渐降低。GE等[16]在研究初始含水率和pH对牛粪堆肥过程微生物演替的影响时,也发现从高温期到腐熟期细菌多样性逐渐降低。总体来讲,MC65处理细菌丰富度高于其他处理,细菌多样性高于MC60处理,但升温期细菌多样性低于MC55处理。MC65处理真菌丰富度与MC60处理相当,低于MC55处理,真菌多样性低于MC60处理,高于MC55处理。这表明,MC65有助于提高堆肥过程中细菌丰富度与多样性,但MC60可提高真菌多样性。
4) 群落结构分析。主坐标分析(PCoA)可以了解不同处理间微生物群落差异性。PC1和PC2共解释了39.52%的细菌差异性和60.48%的真菌差异性 (图12) 。如图12 (a) 所示,3个处理不同时期样本细菌群落结构均有明显分离,表明堆肥时间对细菌分布有显著影响,DU 等[51]也得到了这样的结论。MC55和MC65处理在反应初期样本群落结构差异较小,与MC60处理在第二坐标轴上存在较大差异。MC60和MC65处理在高温期和腐熟期细菌群落结构具有相似性,而与MC55处理细菌群落有明显的分离。这表明,MC60和MC65处理细菌群落演替具有相似性,都与MC55处理细菌群落演替存在明显差异。如图12 (b) 所示,MC55和MC65处理不同时期样本真菌群落变化相似,升温期和高温期真菌群落结构在第一坐标轴上有明显的分离,腐熟期真菌群落与堆肥初期和高温期均有明显的分离。MC60处理不同时期样本间真菌群落差异性较小,且与MC55和MC65处理有明显的分离。这表明,MC55和MC65处理真菌群落随时间的变化具有相似性,但与MC60存在较大差异。
5) 冗余分析RDA。堆肥是微生物活动的过程,环境因子对微生物群落结构有很大影响。采用RDA分析方法,分析了环境因子 (MC、TC、TN、TK、TP、pH、EC、C/N、AN) 对人粪污堆肥系统中相对丰度前20的细菌属和真菌属的影响。如图13所示,RDA1和RDA2共解释了细菌群落结构和环境因子总变异的63.61%,RDA1和RDA2共解释了真菌群落结构和环境因子总变异的68.86%。RDA图中箭头的长度表示环境因子与样本分布的相关程度[40]。C/N是引起堆体中细菌群落变化的关键因素,C/N和MC是引起堆体中真菌群落变化的关键因素,此外,在引起细菌群落变化的环境因子中,C/N和MC具有很高的正相关性。这可能是因为,C、N是微生物代谢的主要营养物质[52],而MC是可溶性营养物质运输的载体[9]。随着堆肥的进行,MC对的微生物群落结构的影响逐渐减小。这表明,MC对微生物群落结构的影响主要发生在堆肥初期。这与LI等[14]的研究结果一致。
-
1) MC55、MC60和MC65处理,经30 d好氧堆肥,堆肥产品均可达到腐熟标准,堆肥产物没有作物毒性 (GI>80%) 。3个处理组堆体高温期 (>50 ℃) 持续天数为2~5 d,MC65处理高温持续时间最长。堆肥结束时,MC60和MC65处理堆肥产物均达到无害化卫生标准,MC55处理堆肥产物未达到无害化卫生标准。
2) 3个处理组细菌丰度均高于真菌。MC65处理有助于提高堆肥过程中细菌丰富度与多样性,但MC60处理可提高真菌多样性。MC65处理增加了腐熟期拟杆菌门、担子菌门和与腐熟相关的菌属的相对丰度,加速了木质纤维素的降解及堆肥腐熟进程。MC对微生物群落结构的影响主要发生在堆肥初期。
3) 综合考虑堆肥物料的基本理化性质、养分质量分数和无害化效果3方面因素,人粪污和玉米秸秆联合好氧堆肥的最佳初始含水率为65%。
初始含水率对人粪污好氧堆肥腐熟及微生物群落结构变化的影响
Effect of the initial moisture content on the maturity extent and the microbial community structure of the aerobic compost with human excrement
-
摘要: 为探究人粪污好氧堆肥最佳工艺参数,探究初始含水率对人粪污好氧堆肥腐熟及微生物群落结构变化的影响。以人粪污为原料,玉米秸秆为调理剂,设置初始含水率55% (MC55) 、60% (MC60) 和65% (MC65) 3个处理组,在60 L的密闭发酵罐中堆肥30 d。结果表明,在不同初始含水率条件下,堆肥产品均可达到腐熟标准,堆肥产物没有作物毒性 (GI>80%) 。MC60和MC65处理堆肥产物均已达到无害化卫生标准,MC55处理堆肥产物未达到无害化卫生标准,其粪大肠菌群数依然高于102 MPN·g−1。3组处理中,细菌丰度均高于真菌,MC65有助于提高堆肥过程中细菌丰富度与多样性,但MC60可提高真菌多样性。3组处理间主要细菌门和真菌门组成一致,MC65处理增加了腐熟期拟杆菌门和担子菌门的相对丰度。升温期和高温期3个处理优势细菌属具有明显差异;腐熟期MC60和MC65处理细菌属组成具有相似性,MC55和MC65处理真菌属随时间的变化具有相似性。MC65处理增加了高温期Bacillus和Meyerozyma的相对丰度,增加了腐熟期uncultured_bictureium和Coprinopsis的相对丰度。MC对真菌演替的影响大于细菌,随着堆肥的进行,MC对微生物群落结构的影响逐渐减小。MC65处理高温期持续时间最长,堆肥产物粪大肠菌群数最低,增加了腐熟期木质素降解菌的相对丰度,是人粪污好氧堆肥最佳初始含水率。本研究结果可为优化人粪污好氧堆肥工艺参数提供参考。Abstract: In order to study the optimal process parameter of the aerobic composting with the human excrement, the effect of the initial moisture content (MC) on the maturity extent of the aerobic composting and the microbial community structure of the human excrement were studied. Adopting the human excrement as the raw material and the corn straw as conditioner, three treatment groups were set up with initial moisture content of 55% (MC55), 60% (MC60) and 65% (MC65), respectively. The experiment was conducted in three 60 L closed fermenters with composting for 30 days. The results showed that the compost products could fulfill the maturity standard and had no crop toxicity (GI>80%) with different initial moisture contents. The compost products of MC60 and MC65 had satisfied the non-hazardous sanitary standard, while the compost products of MC55 could not, with the fecal coliforms which was higher than 102 MPN·g−1. The bacterial abundances were higher than those of fungi in all three treatments, which indicated that the treatment of MC65 could improve the richness and the diversity of the bacteria during the composting process, but MC60 could increase the diversity of the fungi. The compositions of the major bacterial phyla and fungal phyla were consistent among the three treatments. The treatment of MC65 increased the relative abundance of the Bacteroidetes and Basidiomycetes during the mature phase. The dominant bacterial generas among the three treatments were significantly different during the heating and the thermophilic phases. The bacterial genus was similar between MC60 and MC65 during the mature phase, and the variations of fungal genera during the compost period were similar for MC55 and MC65. The treatment of MC65 improved the relative abundances of Bacillus and Meyerozyma during the thermophilic phase, as well as uncultured_bictureium and Coprinopsis during the mature phase. The effect degree of MC on the fungal succession was greater than that of the bacteria, and the effect of MC on microbial community structure gradually decreased throughout the composting process. For compost products, MC65 has the longest thermophilic phase and the lowest number of fecal coliforms, and the relative abundance of lignin-degrading bacteria was increased in the mature phase, which illustrated that it was the best initial moisture content for the aerobic compost with human excrement. This research can provide the reference for optimizing the technological parameters for aerobic compost with human excrement and promote the resource utilization of human excrement.
-
Key words:
- moisture content /
- human excrement /
- aerobic composting /
- microbial community structure
-
目前,水污染仍是一个影响经济和生态系统的全球性问题,特别是重金属引发的水污染已成为备受关注的问题之一[1-3]。化学上将密度大于4.5 g·cm−3的金属称为重金属,如铬、铜、铅、锌、钴、镍等[4]。重金属离子(Cu(Ⅱ)、Pb(Ⅱ)、Cd(Ⅱ)、Zn(Ⅱ)等)具有毒害性、长期累积性和难降解性等特性,造成的水污染具有隐蔽性和不可逆性[5-6]。而砷(As)是一种类金属元素,在自然界中分布广泛。有色金属矿山的开采和冶炼、电镀等化工业的不合规排放以及过量使用化肥、农药是含砷废水的主要来源[7-8]。其中,冶炼厂和化工厂所产生的含砷废水主要含有As、Cu、Pb、Zn、Cd等重金属离子和氟、硫等元素。根据冶炼原料中砷含量的不同,废水中砷含量一般从几十毫克每升到几十克每升不等,其含量均高于《污水综合排放标准》(GB 18466-2005)中规定的0.5 mg·L−1标准限值,因此,需经过处理达标后才能排放[9]。
现阶段,用于处理重金属废水的方法主要有吸附法、化学沉淀法、离子交换法、膜滤法和溶剂萃取法等[10-13]。其中,吸附法具有操作简单、吸附高效和成本低等特点,被广泛应用于重金属废水治理[10, 14-16]。在吸附剂的选择上,使用易获取和可降解天然高分子材料也逐渐成为趋势。氧化石墨烯(graphene oxide, GO)作为一种新型纳米材料,因其比表面积较大,含氧官能团(如—OH、—C=O和—COOH)丰富和易表面修饰等优点,在处理重金属废水中备受关注[17-19]。刘伟等[18]通过制备磁改性GO,发现磁改性GO在pH=2时对5 mg·L−1的Cr(VI)去除率达82.9%。李仕友等[20]成功制备出GO/SiO2复合材料,但GO/SiO2在pH=2.5时对5 mg·L−1 Cd(Ⅱ)的去除率仅为45.2%。再者,有学者利用不同浓度NaOH对GO进行水热处理后,对重金属的吸附量比改性前提高了3倍,但改性后的GO亲水性较强,难以回收利用[21]。壳聚糖(chitosan, CS)是一种具有大量羟基和高活性氨基的天然可降解高分子材料,能通过螯合反应与重金属离子进行配位,也是具有潜力的新型吸附材料[22-23]。此外,CS作为脱乙酰化甲壳素产品,来源广泛,无毒、易降解和抗菌,使其在重金属废水处理中比其他材料更具优势[24-25],但CS在酸性条件下易流失。因此,将GO与CS组合后制备出的GO/CS复合材料不仅可以降低GO/CS的亲水性[26-28],也能进一步改善对水中重金属的去除效果。龚育等[29]研究发现,添加5% GO的CS对铌(Nb)的去除效果优于CS。罗肃霜等[30]制备的TiO2/CS微粒在紫外光催化下对As(Ⅲ)的最大吸附容量虽有4.97 mg·g−1,但在微粒中加入GO后,吸附容量达到12.43 mg·g−1。ZHANG等[31]通过制备EDTA-2Na改性氧化石墨烯壳聚糖(GEC)复合材料,发现对Cr(VI)的吸附容量大小依次为GEC>GOCS>CS。
此外,将GO或CS复合材料负载铁或铁的(氢)氧化物后,能进一步增强复合材料的抗酸性能和热稳定性[32-33]。PARASTAR等[34]将GO和CS先后加入到Fe2S3和H2O2混合溶液中,制备成GO/FeOOH/CS复合材料并用于去除水中的Pd(Ⅱ)和Cd(Ⅱ),发现在pH=6时复合材料对5 mg·L−1的Pd(Ⅱ)和Cd(Ⅱ)最高去除率分别为29.6%和43.07%。AHMAD等[35]利用共沉淀法制备壳聚糖-氧化铁(CS-Fe2O3)用于去除水中的Pb(Ⅱ)和Cd(Ⅱ),发现在pH=5时CS-Fe2O3对50 mg·L−1 Pb(Ⅱ)和Cd(Ⅱ)的去除率均在85%以上。SHERLALAC等[27]通过共沉淀法制备的磁性壳聚糖氧化石墨烯(CMGO)纳米复合材料在pH=7.3时对10 mg·L−1 As(Ⅲ)的去除率为61%。HOSSEINZADEH等[36]同样通过共沉淀法制备的磁性壳聚糖/氧化石墨烯(MCGONs)纳米复合材料在pH=8时对100 mg·L−1 Cu(Ⅱ)的去除率为38%。赵超然等[37]通过包埋法制备不同载铁量的GOCS球形材料用于吸附As(Ⅲ),发现As(Ⅲ)的吸附容量随pH增加呈下降趋势,在pH=3时吸附量达到3.53 mg·g−1。SHAN等[23]对比研究了浸渍蒸发法和共沉淀法制备FeOx修饰的氧化石墨烯壳聚糖复合材料对Cr(VI)的去除效果,发现浸渍蒸发法制备的复合材料具有更好的吸附性能。上述研究表明:铁改性GO或CS复合材料对水体中不同重金属离子均有去除效果,但复合材料中的载铁形态及其改性方法会明显影响去除效果。这将进一步影响其在不同水环境条件下对重金属离子(如As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ))的去除效果。然而,较少有研究对比讨论载铁形态或改性方法对复合材料在不同溶液pH中对重金属离子的去除影响。
因此,本研究基于常用的浸渍蒸发法和共沉淀法分别制备2种不同铁改性氧化石墨烯壳聚糖复合材料微球,设计并开展2种微球在不同水环境中(pH分别为3、7和11)对As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ)的吸附实验,对比分析去除效果。同时,结合扫描电镜(scanning electron microscope,SEM)、傅里叶红外光谱(fourier transform infrared, FTIR)、X射线衍射(X-ray diffraction, XRD)和比表面积(BET surface area)表征技术揭示吸附机制,为不同吸附剂应用于多种重金属废水的治理提供参考。
1. 材料与方法
1.1 试剂与材料
氧化石墨烯(GO, 95%)购于苏州碳丰石墨烯科技有限公司。壳聚糖(CS, 脱乙酰度>90%)购于上海蓝季科技发展有限公司。七水合砷酸钠(Na2HAsO4·7H2O, AR, 成都贝斯特)。二水合氯化铜(CuCl2·2H2O, AR),硝酸铅(Pb(NO3)2, AR),四水合硝酸镉(Cd(NO3)2·4H2O, AR),六水合三氯化铁(FeCl3·6H2O, AR),四水合氯化亚铁(FeCl2·4H2O, AR),盐酸(HCl, GR),氢氧化钠(NaOH, GR),冰醋酸(CH3COOH, GR),甲醇(CH3OH, AR),无水乙醇(C2H5OH, AR),硝酸(HNO3, AR)均采购于西陇化工股份有限公司。戊二醛(C5H8O2, 50 %),溴化钾(KBr, GR)购自于阿拉丁试剂(上海)有限公司。实验用水均为超纯水。
使用浸渍蒸发法制备Fe(Ⅱ)改性氧化石墨烯壳聚糖复合微球(Fe-GOCS)。首先将氧化石墨烯和壳聚糖粉末按照一定比例混合,加入配置好的200 mL 1.5%的醋酸溶液,超声搅拌直至二者混合均匀,得到GOCS混合液;将混合液滴入至2% NaOH溶液中,常温静置固化成球,用超纯水将洗涤液洗至接近中性,过滤;在过滤后的材料中加入200 mL 5%戊二醛-甲醛溶液后置于摇床振荡2~3 h,用50%乙醇溶液洗涤至无醛味后,过滤,放置45 ℃的鼓风干燥箱中烘干,得到GOCS复合微球;将微球放入0.1 mol的FeCl2·4H2O溶液中,放置于200 ℃的电热板上加热蒸干,冷却后洗涤至电导率低于25 μs·cm−1,烘干,得到Fe-GOCS复合微球。使用共沉淀法制备Fe(Ⅲ)改性氧化石墨烯壳聚糖复合微球(Fe@GOCS),在上述配置好的GOCS混合液中加入0.1 mol FeCl3·6H2O,继续超声搅拌直至固体完全溶解,得到Fe@GOCS混合溶液;将该溶液滴入至5% NaOH溶液中,静置24 h后,洗涤至中性,过滤,置于200 mL 5%戊二醛-甲醛溶液中摇床振荡2~3 h后,用50%乙醇溶液洗涤至无醛味,过滤,烘干,得到Fe@GOCS复合微球。
1.2 实验方法
1)实验设计。采用批实验对比研究在不同pH条件下 GOCS、Fe-GOCS和Fe@GOCS分别对As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ)的去除效果。分别称取50 mg GOCS、Fe-GOCS和Fe@GOCS置于若干100 mL离心管中,分别添加50 mL 25 mg·L−1 As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ)模拟液,再使用1 mol·L−1 HCl和1 mol·L−1 NaOH调节模拟液pH分别为3±0.1、7±0.1和11±0.1,然后将模拟液置于恒温水浴振荡器中,在298.15 K、170 r·min−1条件下,振荡48 h。使之充分反应后,取上清液过0.45 μm滤膜后测定重金属离子浓度。
在固液比为1.0 g·L−1、As(Ⅴ)初始质量浓度为25 mg·L−1、温度为298.15 K和pH分别为7和3的条件下,收集首次吸附As(Ⅴ)后的Fe-GOCS和Fe@GOCS,并用50 mL 1.5 mol·L−1 NaOH和超纯水洗脱,直到洗脱液电导率稳定,然后将这些复合材料干燥后再次用于As(Ⅴ)的吸附。解吸和吸附过程重复5次,每次吸附后收集上清液和脱吸液用于As(Ⅴ)浓度的测定,结果将用于分析多次解吸和吸附对As(Ⅴ)去除效果的影响,以及吸附剂的再生性能。
2)样品表征分析。吸附前后As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ)的浓度均利用电感耦合等离子体发射光谱仪(ICP-OES, Optima 7000 DV)测试,测试数据误差小于5%。对GOCS和吸附反应前后的Fe-GOCS和Fe@GOCS进行表征分析。其中,复合材料的官能团使用溴化钾压片法由傅里叶红外光谱仪(FTIR, iS 10, 美国赛默飞世尔公司)测试完成;样品结构测定采用X射线衍射仪(XRD,X'Pert3 Powder,荷兰帕纳科公司),测试扫描2θ步长0.026 26°,扫描速度为0.656 5(°)·s−1,扫描2θ为5°~90°;形貌微观特征由场发射电子扫描显微镜(SEM, JSM-7900F, 日本)测定;样品的比表面积和孔结构采用全自动比表面及孔隙度分析仪(BET, Micromeritics ASAP 2020, 美国)测试。
3)数据处理。GOCS、Fe-GOCS和Fe@GOCS对As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ)平衡吸附量(qe)和去除率(re)分别由式(1)和式(2)计算。
stringUtils.convertMath(!{formula.content}) (1) stringUtils.convertMath(!{formula.content}) (2) 式中:qe是吸附平衡时吸附剂对重金属离子的吸附量,mg·g−1;C0和Ce分别是重金属离子初始和吸附平衡时的质量浓度,mg·L−1;V是含重金属离子模拟液的体积,mL;m是吸附剂的投加量,mg;re是吸附平衡时重金属离子的去除率,%。
2. 结果与讨论
2.1 不同pH条件下重金属的去除效果
不同pH条件下,GOCS、Fe-GOCS和Fe@GOCS对As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ)的去除结果分别如表1和图1所示。可以发现:改性前后的复合材料对不同重金属的平衡吸附量(qe)和去除率(re)存在明显差异。其中,GOCS和Fe@GOCS对As(Ⅴ)的qe均随pH升高而逐渐降低,qe均在pH=3时最大,分别为7.72 mg·g−1和22.54 mg·g−1。在pH=11时GOCS对As(Ⅴ)的qe虽有0.12 mg·g−1,但Fe@GOCS却为0(图1(a)和图1(c))。而Fe-GOCS对As(Ⅴ)的qe随pH升高先显著增加后略微降低。当pH=3时,qe最小为7.02 mg·g−1;pH=7时,qe达到最大为14.54 mg·g−1(图1(b))。结合表1可知:酸性条件下,Fe@GOCS对As(Ⅴ)去除率显著高于GOCS和Fe-GOCS,三者对As(Ⅴ)的去除率分别为87.99%、30.20%和27.40%。同时,除As(Ⅴ)过程中Fe平衡质量浓度仅为0.04 mg·L−1,远低于《生活饮用水卫生标准》(GB 5749-2022)所规定的0.3 mg·L−1。而Fe-GOCS的Fe平衡质量浓度为0.4 mg·L−1,存在二次污染风险,说明酸性条件下Fe@GOCS比Fe-GOCS更稳定。中性和碱性条件下,Fe-GOCS对As(Ⅴ)的re则显著高于GOCS和Fe@GOCS,Fe平衡质量浓度分别为0和0.08 mg·L−1,均低于0.3 mg·L−1的限值。相比之下,Fe@GOCS更适用于酸性条件下对As(Ⅴ)的去除,而在中性和碱性条件下Fe-GOCS更适用。同时,在最佳适用pH条件下,不同铁改性后的GOCS对As(Ⅴ)的去除效果均优于GOCS。
表 1 不同pH条件下复合材料对重金属的去除结果Table 1. Removal results of heavy metals by composite materials under different pH conditions离子 pH GOCS Fe-GOCS Fe@GOCS qe/(mg·g−1) re/(%) qe/(mg·g−1) re/(%) Fe平衡质量浓度/(mg·L−1) qe/(mg·g−1) re/(%) Fe平衡质量浓度/(mg·L−1) As(Ⅴ) 3 7.72 30.20 7.02 27.40 0.40 22.54 87.99 0.04 7 2.01 7.86 14.54 56.74 0 10.38 40.53 0.04 11 0.12 0.47 9.45 36.87 0.08 — — 0.15 Cu(Ⅱ) 3 — — — — 0.26 — — 0.13 7 18.59 74.27 12.24 49.45 0 16.92 68.38 0 11 24.97 99.74 24.53 99.09 0 24.71 99.83 0 Pb(Ⅱ) 3 — — — — 0.15 — — 0.06 7 11.21 47.15 5.37 23.52 0 11.60 50.85 0 11 23.63 99.38 22.77 99.78 0 22.68 99.39 0 Cd(Ⅱ) 3 — — — — 0.13 — — 0.05 7 0.64 2.57 — — 0.05 — — 0 11 24.65 98.31 19.65 78.30 0 24.89 99.16 0 注: “-”表示未吸附。 | Show Table DownLoad:
CSV
DownLoad:
CSV
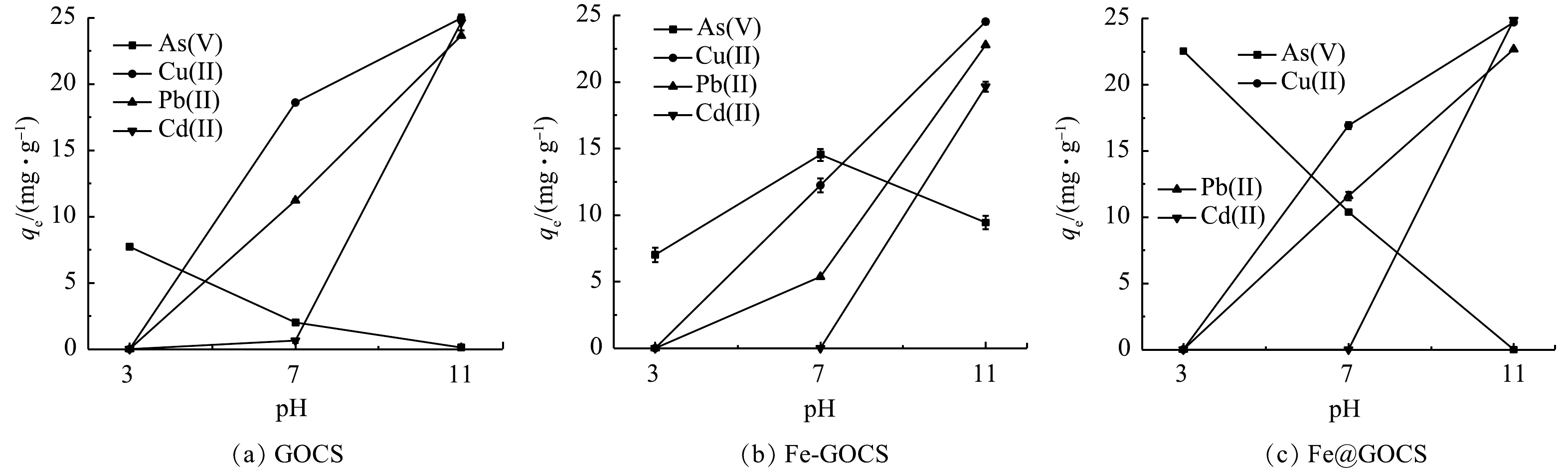 图 1 GOCS、Fe-GOCS和Fe@GOCS对As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ)的平衡吸附量随pH的变化Figure 1. Changes of equilibrium adsorption capacity of GOCS, Fe-GOCS and Fe@GOCS to As(Ⅴ), Cu(Ⅱ), Pb(Ⅱ) and Cd(Ⅱ) with pH
图 1 GOCS、Fe-GOCS和Fe@GOCS对As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ)的平衡吸附量随pH的变化Figure 1. Changes of equilibrium adsorption capacity of GOCS, Fe-GOCS and Fe@GOCS to As(Ⅴ), Cu(Ⅱ), Pb(Ⅱ) and Cd(Ⅱ) with pH由图1可知,3种材料对Cu(Ⅱ)和Pb(Ⅱ)的去除规律相似,qe均随着pH的增大呈上升趋势。其中,pH=3时,GOCS、Fe-GOCS和Fe@GOCS均未去除Cu(Ⅱ)和Pb(Ⅱ)。pH=7时,GOCS、Fe-GOCS和Fe@GOCS对Cu(Ⅱ)的qe分别为18.59、12.24和16.92 mg·g−1,对Pb(Ⅱ)的qe分别为11.21、5.37和11.60 mg·g−1。而pH=11时,3种材料对Cu(Ⅱ)和Pb(Ⅱ)的qe相近,均达到最大。其中,GOCS、Fe-GOCS和Fe@GOCS对Cu(Ⅱ)的qe分别为24.97、24.53和24.71 mg·g−1,对Pb(Ⅱ)的qe分别为23.63、22.77和22.68 mg·g−1。在中性和碱性条件下均未发现Fe溢出。由此可知,3种材料均不适用于去除酸性条件下的Cu(Ⅱ)和Pb(Ⅱ)。中性条件下,GOCS更适用去除Cu(Ⅱ),其次是Fe@GOCS;而对于去除Pb(Ⅱ)的结果则相反。碱性条件下考虑到Cu(Ⅱ)和Pb(Ⅱ)存在水解或共沉淀作用[38],3种材料均不适用于对Cu(Ⅱ)和Pb(Ⅱ)的去除。
从表1 Cd(Ⅱ)的去除结果中可知,3种材料在pH=3均未吸附Cd(Ⅱ)。pH=7时,仅GOCS具有微弱的去除效果,qe和re分别为0.64 mg·g−1和2.57%。当pH=11时,Fe@GOCS的去除效果均优于GOCS和Fe-GOCS,qe分别为24.89、24.65和19.65 mg·g−1。由此可知,GOCS更适用于中性条件下去除Cd(Ⅱ)。而在碱性条件下,Cd(Ⅱ)存在较强的水解或共沉淀作用[38],表明2种材料均不适用于去除Cd(Ⅱ)。
2.2 吸附介质前后变化特征
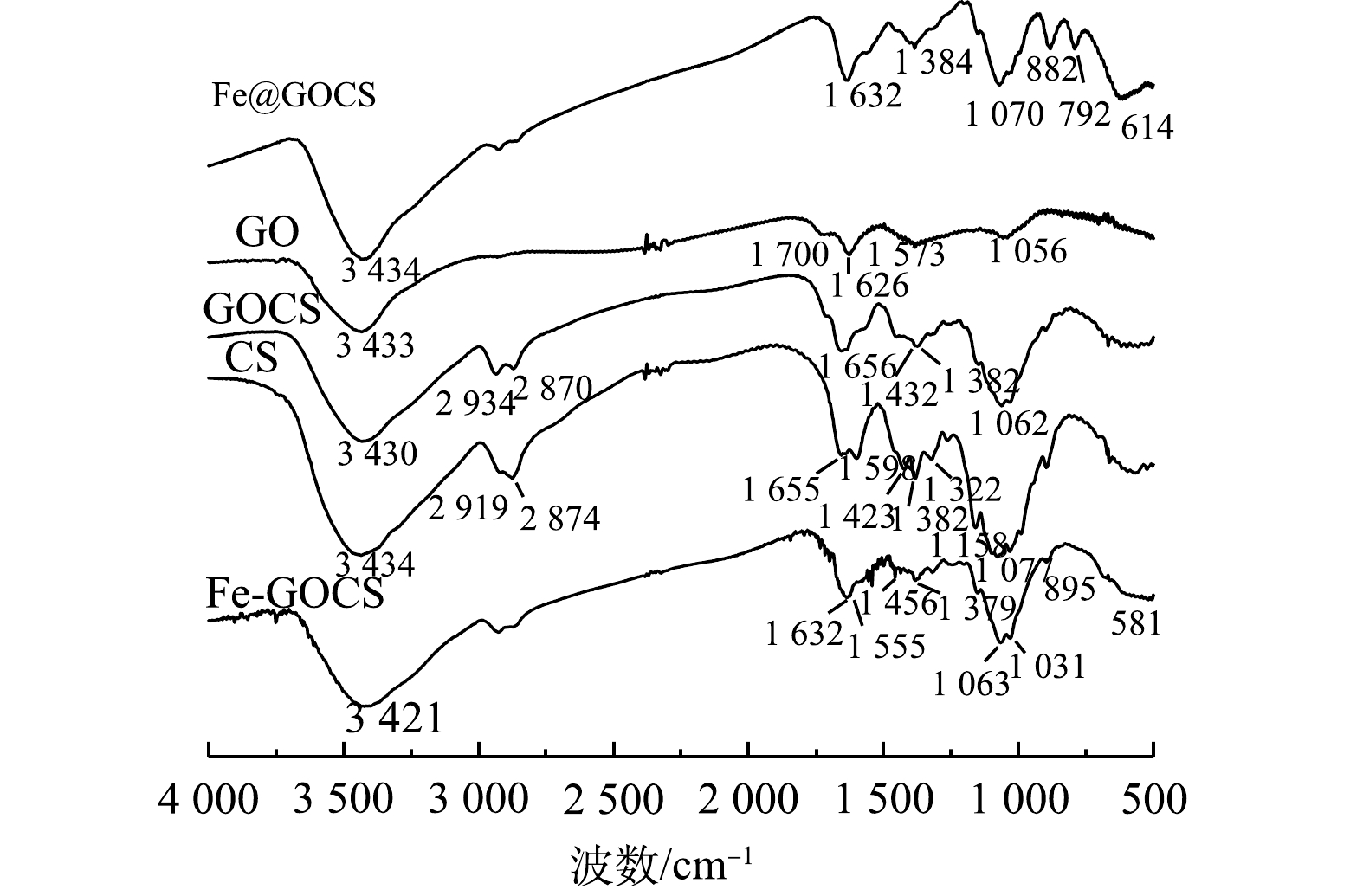
1) FTIR表征。吸附前复合材料的FTIR表征结果如图2所示。由图2可知,GO、CS和GOCS具有多种官能团。其中,GO图谱中1 700 cm−1附近是—COOH的C=O伸缩振动峰,1 626 cm−1附近是水分子中的O—H弯曲振动峰,1 573 cm−1附近是—NH2面内变形特征峰和1 056 cm−1附近的C—O—C弯曲振动峰。CS图谱中3 434 cm−1附近是N—H伸缩振动峰,2 919 cm−1和2 874 cm−1附近是CH3和CH2中C—H伸缩振动峰,1 655 cm−1和1 598 cm−1附近是酰胺I键C=O的伸缩振动峰,1 432~1 322 cm−1附近是CH3和CH2中C—H弯曲振动峰,1 158 cm−1附近是C—O—C弯曲振动峰以及1 077 cm−1的碳环弯曲振动峰[39]。将GO与CS混合后,图谱显示GOCS在3 430 cm−1附近是水分子中O—H和CS中N—H伸缩振动峰,在2 919 cm−1和2 870 cm−1附近保留了CS中的C—H伸缩振动峰,在1 062 cm−1附近形成较强的环氧基C—O—C的C—O弯曲振动峰。此外,GOCS在1 656 cm−1附近形成酰胺键—CONH—,该特征峰说明GO中的—COOH与CS中的—NH2组装成功[37]。在经过不同铁改性后,GOCS图谱出现不同特征。其中,Fe@GOCS在1 070 cm−1附近出现碳环弯曲振动峰,在1 384 cm−1附近出现C—OH弯曲振动峰。882 cm−1和792 cm−1附近出现α-FeO(OH)中O—H的面内和面外弯曲振动峰,是针铁矿的特征吸收峰,614 cm−1为Fe—O的特征峰。而Fe-GOCS在3 421 cm−1出现的宽峰是其表面—OH与水分子形成的氢键有关[40],1 063 cm−1和1 031 cm−1附近是C—O弯曲振动峰,1 555 cm−1附近为峰值增强的—NH2面内变形特征峰,在581 cm−1附近形成赤铁矿的Fe—O弯曲振动峰[41]。此外,Fe@GOCS和Fe-GOCS在1 632 cm−1附近均保留了GOCS的酰胺键—CONH—,这与赵超然等[37]和李仕友等[42]的研究结果一致。
Fe-GOCS和Fe@GOCS在最优条件下去除As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ)后的FTIR图谱如图3所示。由图3(a)可见,Fe-GOCS在去除As(Ⅴ)后,1 063 cm−1和1 031 cm−1附近的C—O弯曲振动峰增强,而581 cm−1处对应的Fe—O特征峰偏移至561 cm−1。这说明C—H、C—O和Fe—O均可能参与到As(Ⅴ)去除中。在去除Cu(Ⅱ)和Pb(Ⅱ)后,Fe-GOCS中的Fe—O分别偏移至548 cm−1和568 cm−1且峰强减弱,这说明Fe—O可能参与去除。Fe-GOCS-Cd图谱显示,Fe-GOCS在1 456~1 379 cm−1的C—H多组伸缩振动峰数量减少,仅在1 384 cm−1处均在C—H单峰。同时,在去除As(Ⅴ)、Cu(Ⅱ)和Pb(Ⅱ)后,该处的C—H峰值增强,说明C—H均可能参与到As(Ⅴ)、Cu(Ⅱ)和Pb(Ⅱ)去除。
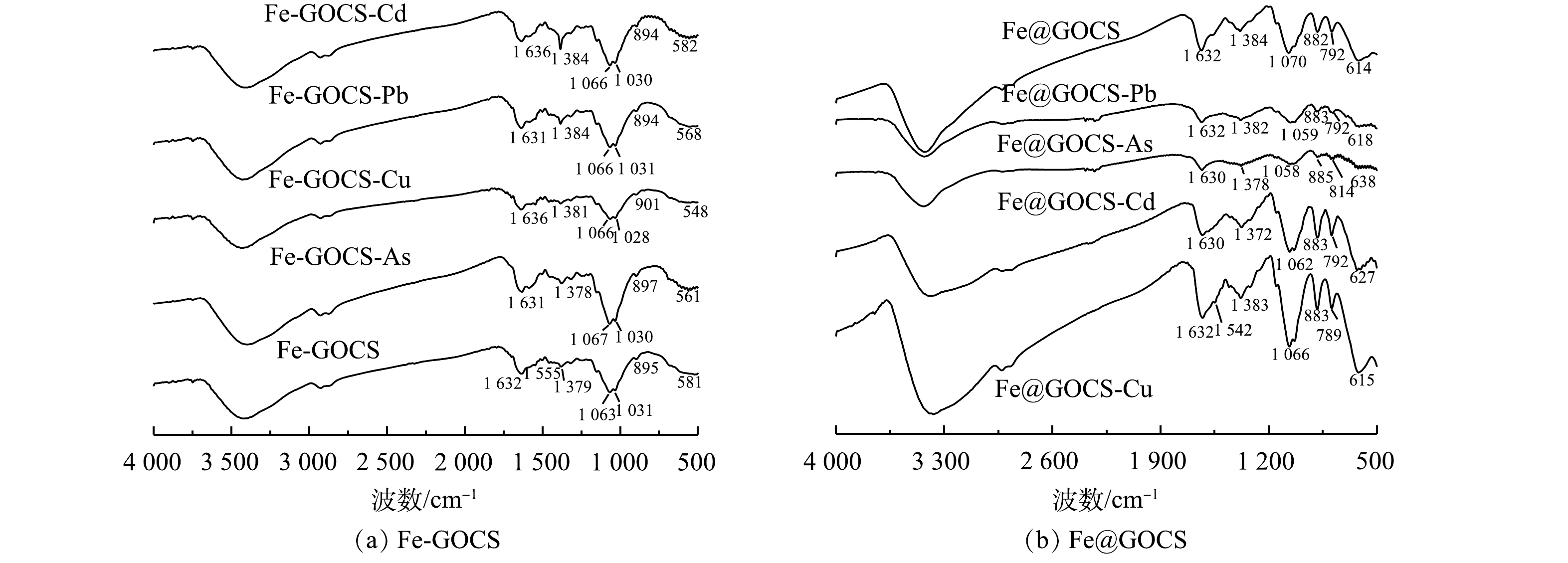 图 3 Fe-GOCS和Fe@GOCS去除重金属后的FTIR图谱Figure 3. FTIR spectra of Fe-GOCS and Fe@GOCS after removal of heavy metals
图 3 Fe-GOCS和Fe@GOCS去除重金属后的FTIR图谱Figure 3. FTIR spectra of Fe-GOCS and Fe@GOCS after removal of heavy metals与Fe-GOCS吸附前后表征结果类似,Fe@GOCS在去除重金属后,官能团也发生不同程度的变化(图3(b))。其中,在去除As(Ⅴ)和Pb(Ⅱ)后,614 cm−1附近对应的Fe—O特征峰偏移至638 cm−1。此外,多组官能团特征峰均出现不同程度减弱,包括882 cm−1和792 cm−1附近的α-FeO(OH)中O—H的面内和面外弯曲振动峰、1 070 cm−1附近的碳环弯曲振动峰、1 384 cm−1附近的C—OH弯曲振动峰以及1 632 cm−1的—CONH—。由此可知,Fe@GOCS去除As(Ⅴ)和Pb(Ⅱ)是多种官能团共同作用的结果。与去除As(Ⅴ)和Pb(Ⅱ)结果相反,Fe@GOCS在去除Cu(Ⅱ)后,多组官能团特征峰均显著增强。这可能是因为Fe@GOCS中的Fe为+3价,Fe—O体系中的氧带有负电荷,而Cu(Ⅱ)的电负性较其他3种重金属离子强[43],与Fe—O上的氧以及O—H、C—OH和—CONH—之间的相互作用较强,因而对分子筛的骨架振动改变较为明显。Fe@GOCS与Cd(Ⅱ)反应后,特征峰位置未出现明显变化,说明Fe@GOCS去除Cd(Ⅱ)后化学结构没有改变,表明Fe@GOCS的主要官能团并未与Cd(Ⅱ)发生显著作用。这一结果与范先媛等[44]使用孔径为4×10−10 m的分子筛去除Cu(Ⅱ)和Cd(Ⅱ)的结果相似。
2) XRD表征。由图4可以看出,GO和CS分别在2θ为12°和42°附近以及20°附近有明显衍射峰,为无定型结构,GOCS仅在2θ=20°附近存在衍射峰,且峰型变窄,峰面积减小。这说明GO和CS的成功组装[32]。Fe-GOCS在2θ为33°和35°附近出现新的衍射峰。但35°附近的衍射峰峰值较弱,这可能受到制备材料中一些杂质的影响。与标准PDF卡片(ICDD PDF 72-0469)进行对照比较,新出现的衍射峰为赤铁矿,以α-Fe2O3矿物形态存在,同时也意味着FeCl2成功将GOCS改性[37]。而Fe@GOCS在2θ为17°、21°、33°、34°、36°以及53°附近均出现新的衍射峰,对比标准PDF卡片(ICDD PDF 81-0462)后确定为针铁矿,以α-FeO(OH)矿物形态存在[45]。
Fe-GOCS和Fe@GOCS在最优条件下去除As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ)后的XRD结果分别如图5(a)和图5(b)所示。由图5可知,反应前后Fe-GOCS和Fe@GOCS的衍射峰位置未发生明显改变且未出现新衍射峰。这表明Fe-GOCS和Fe@GOCS稳定性良好,在去除重金属过程中并没有生成新的物相。此外,由图5(a)可知,Fe-GOCS位于2θ=33°和35°的衍射峰出现不同程度的增强。这说明Fe-GOCS上的α-Fe2O3参与As(Ⅴ)、Cu(Ⅱ)和Pb(Ⅱ)的去除。而对于Cd(Ⅱ),α-Fe2O3衍射峰并未出现明显变化,说明α-Fe2O3并未参与去除Cd(Ⅱ)。这些结果与2.1.1 FTIR分析的结果一致,也再次表明碱性条件下Cd(Ⅱ)的去除主要与离子水解或共沉淀作用有关[44, 46]。由图5(b)中也可以看出,Fe@GOCS衍射峰强度出现不同程度的变化。其中Fe@GOCS在吸附As(Ⅴ)和Pb(Ⅱ)后,位于2θ=17°、21°、36°和53°处的α-FeO(OH)衍射峰峰值明显增强。这可能受到了吸附剂中其他官能团的影响,如O—H、C—OH和—CONH—等。在吸附Cu(Ⅱ)后,α-FeO(OH)衍射峰峰值明显降低,说明α-FeO(OH)参与去除Cu(Ⅱ)。与Fe-GOCS去除Cd(Ⅱ)后的结果类似,Fe@GOCS-Cd的衍射峰并未出现明显变化,表明Fe@GOCS中的α-FeO(OH)没有参与反应。
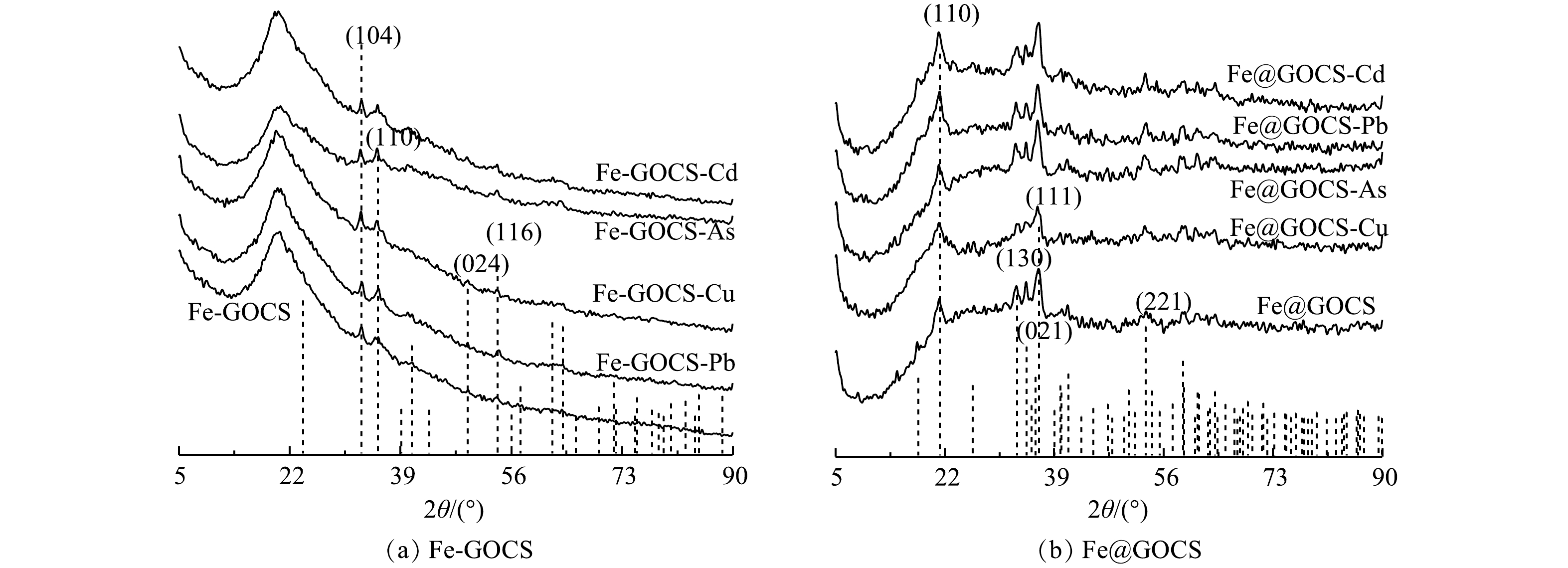 图 5 Fe-GOCS和Fe@GOCS去除重金属后的XRD图谱Figure 5. XRD patterns of Fe-GOCS and Fe@GOCS after removal of heavy metals
图 5 Fe-GOCS和Fe@GOCS去除重金属后的XRD图谱Figure 5. XRD patterns of Fe-GOCS and Fe@GOCS after removal of heavy metals3) SEM表征。GOCS、Fe-GOCS和Fe@GOCS的SEM表征结果如图6所示。可以看出,改性前GOCS表面光滑且富有光泽(图6(a)),堆叠着许多片状结构(图6(b))。经FeCl2浸渍改性后,Fe-GOCS表面变得黯淡且有裂隙出现(图6(c)),同时GOCS的片状层结构被排列紧密的球状物质所代替且有聚集成斑块状物质的现象(图6(d)),结合第2.1.2节XRD物相分析确定该球状物质为赤铁矿;经FeCl3改性后,Fe@GOCS表面出现大量且层次分明的褶皱(图6(e))。同时,褶皱之间充填有针刺状或块状物质(图6(f)),结合第2.1.2节的XRD物相分析确定该针刺状物质为针铁矿。由此可知,GOCS经不同载铁方式后微观表面形貌发生显著变化,同时证实GOCS均被成功改性。这一结果与FTIR和XRD的分析一致。
Fe-GOCS和Fe@GOCS分别吸附As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ)后的微观形貌如图7所示。Fe-GOCS吸附As(Ⅴ)后表面较未吸附之前球状物质虽没有明显变化,但成块面积明显减少(图7(a));吸附Cu(Ⅱ)后,表面的球状物质明显减少且分布不均,暴露出大量针刺状物质(图7(b));吸附Pb(Ⅱ)后的Fe-GOCS表面球状物质形貌发生改变,斑块状物质面积增加,少许的针刺状物质掺杂其中(图7(c));Fe-GOCS在与Cd(Ⅱ)反应后斑块状物质面积与未反应前并未发生明显变化,仅斑块状物质较为分散(图7(d))。这表明Fe-GOCS虽未能够去除Cd(Ⅱ),但Cd(Ⅱ)可以修饰材料表面形貌。Fe@GOCS在去除As(Ⅴ)后,原先的褶皱变得光滑,但有明显萎缩迹象且变得杂乱无序(图7(e));吸附Cu(Ⅱ)后的Fe@GOCS表面褶皱消失,变得粗糙且凹凸不平(图7(f));在去除Pb(Ⅱ)后,褶皱间的针刺状或块状物质明显减少,褶皱表面出现大小不等的凹陷(图7(g));与Cd(Ⅱ)反应后的Fe@GOCS表面褶皱几乎消失殆尽,仅一些针刺状物质残留在表面(图7(h))。
 图 7 Fe-GOCS和Fe@GOCS分别去除As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ)后的SEM图Figure 7. SEM images of Fe-GOCS and Fe@GOCS after removing As(Ⅴ), Cu(Ⅱ) , Pb(Ⅱ) and Cd(Ⅱ), respectively
图 7 Fe-GOCS和Fe@GOCS分别去除As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ)后的SEM图Figure 7. SEM images of Fe-GOCS and Fe@GOCS after removing As(Ⅴ), Cu(Ⅱ) , Pb(Ⅱ) and Cd(Ⅱ), respectively4) BET表征。GOCS、Fe-GOCS和Fe@GOCS的BET比表面积和孔结构参数列于表2。由表2可见,GOCS的比表面积和孔体积分别为4.22 m2·g−1和2.25×10−4 cm3·g−1。经FeCl3改性后,Fe@GOCS比表面积增至42.27 m2·g−1,孔体积增至6.58×10−4 cm3·g−1,这使得酸性条件(pH=3)下Fe@GOCS对As(Ⅴ)的吸附量比GOCS高出14.82 mg·g−1。同时,结合SEM表征结果,Fe@GOCS表面出现大量且层次分明的褶皱有利于进一步提高吸附量。经FeCl2改性后,相较于GOCS,Fe-GOCS的比表面积降低2.12 m2·g−1,孔体积降低0.38×10−4 cm3·g−1,这使得Fe-GOCS在酸性条件下对As(Ⅴ)的吸附量减少0.70 mg·g−1。但在中性和碱性条件下,3种材料对于As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ)的去除效果也存在差异,这说明除材料本身的空间结构外,材料表面的电荷、载铁形态以及离子形态也会影响去除效果[37, 47-48]。
表 2 GOCS和Fe-GOCS和Fe@GOCS的比表面积和孔结构参数Table 2. Specific surface area and pore structure parameters of GOCS, Fe-GOCS and Fe@GOCS吸附材料 比表面积/(m2·g−1) 孔体积/(cm3·g−1) GOCS 4.22 2.25×10−4 Fe-GOCS 2.10 1.87×10−4 Fe@GOCS 42.27 6.58×10−4 | Show TableDownLoad:
CSV
2.3 再生实验
As(Ⅴ)分别在Fe-GOCS和Fe@GOCS上重复解吸和吸附5次后,平衡吸附容量(qe)的变化如图8所示。Fe-GOCS在对As(Ⅴ)的初始qe虽有13.19 mg·g−1,但在循环吸附1次后,Fe-GOCS并未展现出良好的吸附效果。而Fe@GOCS对As(Ⅴ)的qe随着解吸和吸附循环次数的增加先上升后略有降低,这可能与材料表面上铁的溶解损失有关。在第5次解吸后,Fe@GOCS对As(Ⅴ)的qe仍高达22.02 mg·g−1,仅比初始qe减少0.31 mg·g−1,说明Fe@GOCS再生性良好,可重复用于酸性含砷废水中。同时在整个实验过程中,虽然Fe-GOCS和Fe@GOCS中的部分Fe溶解在水溶液中,但Fe平衡质量浓度分别为0~0.22 mg·L−1和0~0.04 mg·L−1,仍未超过《生活饮用水卫生标准》(GB 5749-2022)铁的0.3 mg·L−1的标准限值。这不仅进一步说明本研究制备的Fe@GOCS比Fe-GOCS具有更加稳定且良好的环保再生能力,也使Fe@GOCS可进一步用于吸附处理As(Ⅴ)脱吸液(C0=0.95~17.16 mg·L−1),降低脱吸液对环境的二次污染风险。
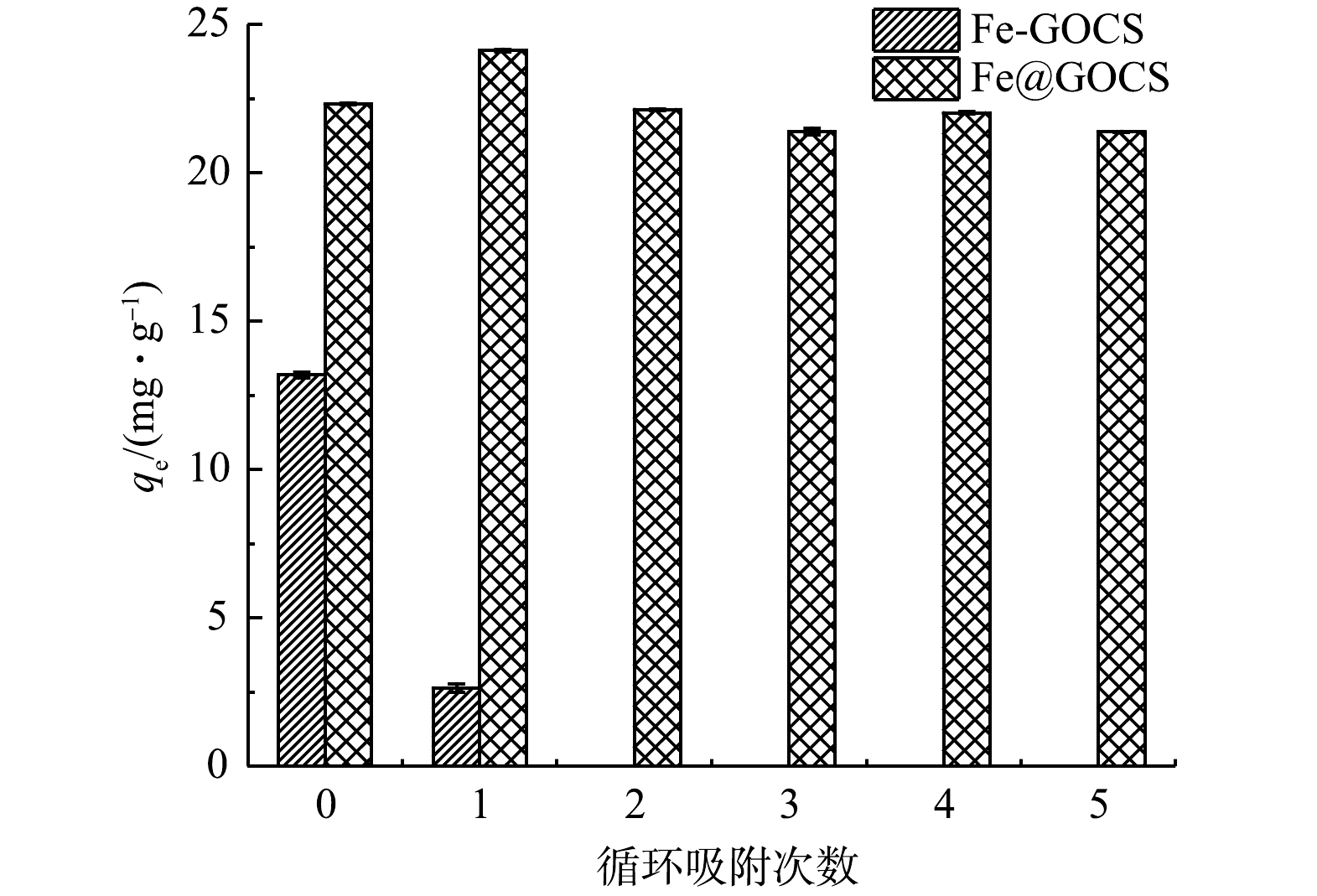 图 8 5次重复解吸-吸附过程中As(Ⅴ)平衡吸附容量(qe)的变化Figure 8. Variation of equilibrium adsorption capacity to As(Ⅴ) during five repeated desorption-adsorption processes
图 8 5次重复解吸-吸附过程中As(Ⅴ)平衡吸附容量(qe)的变化Figure 8. Variation of equilibrium adsorption capacity to As(Ⅴ) during five repeated desorption-adsorption processes2.4 性能评价
将所制备的Fe-GOCS和Fe@GOCS与目前所制备的复合材料对As(Ⅴ)、Cu(Ⅱ)和Pb(Ⅱ)去除效果进行对比,其结果如表3所示。一方面,在pH=3时Fe@GOCS对As(Ⅴ)的re为87.99%,均高于AMD污泥复合材料[49]和含铁锰污泥壳聚糖-海藻酸吸附剂[50]等。GO和CS来源广泛和价格低廉[17, 24-25],同时Fe@GOCS制备方法简便且具有良好的环保再生能力,这也为Fe@GOCS在As(Ⅴ)去除提供广阔的应用前景。此外,Fe-GOCS和Fe@GOCS分别在pH=7和3时对25 mg·L−1 As(Ⅴ)吸附达到平衡后,溶液中分别仍有11.08 mg·L−1和3.08 mg·L−1的As(Ⅴ),超出《污水综合排放标准》(GB 18466-2005)中砷的标准限定值0.5 mg·L−1。因此,可使用Fe@GOCS对酸化后仍超标的As(Ⅴ)废液进一步吸附处理。另一方面,在pH=7时,Fe-GOCS和Fe@GOCS对Cu(Ⅱ)的re分别为49.45%和68.38%,均高于磁性GOCS复合材料和纯CS,说明Fe-GOCS和Fe@GOCS对Cu(Ⅱ)去除效果良好。而对于Pb(Ⅱ)和Cd(Ⅱ)的去除,Fe-GOCS和Fe@GOCS却没有表现出良好的去除效果。
表 3 材料去除性能对比Table 3. Comparison of material removal performance吸附剂 离子 pH 初始质量浓度/(mg·L−1) 固液比/(g·L−1) qe/(mg·g−1) re/% 参考文献 Fe-GOCS As(Ⅴ) 7 25 1.0 14.54 56.74 本研究 Fe@GOCS As(Ⅴ) 3 22.54 87.99 纳米磁性壳聚糖/氧化石墨烯 Cu(Ⅱ) 8 100 0.2 200 38 [36] AMD-污泥 As(Ⅴ) 3 40 0.5 30 85 [49] 含铁锰污泥壳聚糖-海藻酸 As(Ⅴ) 3 10 1.0 8.5 85 [50] 钙氧化物纳米颗粒 Pb(Ⅱ) 7 60 7.5 7.85 97.07 [51] 功能化硅胶 Pb(Ⅱ) 6 50 10 4.94 98.78 [52] | Show TableDownLoad:
CSV
总的来说,Fe@GOCS能够兼顾多种重金属离子高效率去除且不会因材料中Fe溢出造成环境二次污染问题,是一种环保型可再生吸附剂。对于Fe-GOCS,能在不同的溶液环境有效去除多种重金属离子,但应进一步提升去除效果。
2.5 去除机理
结合pH、FTIR、XRD和BET分析结果,推测了Fe-GOCS和Fe@GOCS对As(Ⅴ)、Cu(Ⅱ)、Pb(Ⅱ)和Cd(Ⅱ)的去除机理。Fe-GOCS和Fe@GOCS的零点电荷pHpzc分别是7.5[47]和8.2[37]。当溶液环境为弱酸性(pH=3)时,As(Ⅴ)主要以HAsO42-存在[48]。Fe-GOCS中的α-Fe2O3在水溶液中可形成羟基化表面(Fe—OH+2、Fe—OH、FeO−)[53],使材料表面基团质子化,在静电吸引作用下去除HAsO42-形态的As(Ⅴ)。而Fe@GOCS中的α-FeO(OH)本身具有大量的-OH,在pH=3时,Fe—OH质子化形成Fe—OH+2,在静电吸引作用下去除As(Ⅴ)。具体的去除机理可解释为分式(3)和式(4)及总反应(式(5))。
stringUtils.convertMath(!{formula.content}) (3) stringUtils.convertMath(!{formula.content}) (4) stringUtils.convertMath(!{formula.content}) (5) 当溶液环境为中性时,Cu(Ⅱ)和Pb(Ⅱ)均会发生不同程度的水解。其中,Cu(Ⅱ)的水解常数pKCu(pK实测值和pK计算值分别为6.5和7.0)小于Pb(Ⅱ)的水解常数(pK实测值和pK计算值分别为7.2和11.7)[38],说明Cu(Ⅱ)的水解程度大于Pb(Ⅱ),Cu(Ⅱ)的σ杂化轨道接受氧的非键电子对的能力更强,形成Cu-OH配键的键能更大[54],这也证明在中性条件下,2种材料均对Cu(Ⅱ)的去除效果最好。除Cu(Ⅱ)和Pb(Ⅱ)自身水解外,结合FTIR和XRD分析结果,在其他官能团(Fe-GOCS中的C—H和Fe@GOCS中的C—OH、O—H及—CONH—)提供的键能帮助下,Fe-GOCS和Fe@GOCS中的Fe—O分别通过与Cu(Ⅱ)和Pb(Ⅱ)形成Fe—O—Cu和Fe—O—Pb表面络合物实现对其的去除。而Cd(Ⅱ)(pK实测值和pK计算值分别为9.7和9.5[38])主要通过自身水解被去除。
3. 结论
1) Fe-GOCS对As(Ⅴ)的去除率在pH=7时达到最大,为56.74%。而Fe@GOCS对于As(Ⅴ)的去除率在pH=3时达到最大,为87.99%。酸性条件下,应优先考虑Fe@GOCS。
2)溶液pH的改变对Cu(Ⅱ)和Pb(Ⅱ)的去除效果影响较大。Fe-GOCS、Fe@GOCS对Cu(Ⅱ)和Pb(Ⅱ)的去除率在pH=11时均达到最大。除GOCS外,在弱酸性~中性条件下,对于Cu(Ⅱ)和Pb(Ⅱ)的去除应优先考虑Fe@GOCS。
3) Fe-GOCS和Fe@GOCS对Cd(Ⅱ)的吸附在酸性和中性时效果较差。碱性条件下2种复合材料对Cd(Ⅱ)的re均达到最大,但这与复合材料中丰富的官能团无关,主要与Cd(Ⅱ)自身水解有关。
4)再生吸附循环5次后,Fe@GOCS对As(Ⅴ)的qe仍高达22.02 mg·g−1,仅低于初始qe的0.31 mg·g−1,说明说明Fe@GOCS再生性良好,可重复用于酸性含砷废水中,而Fe-GOCS的再生性能较差。
5) Fe-GOCS中的C—H、Fe—O和C—O等官能团,Fe@GOCS中的Fe—O、O—H、C—OH和—CONH—等官能团在As(Ⅴ)、Cu(Ⅱ)和Cd(Ⅱ)去除过程中发挥着主要作用;参与吸附的主要物质分别为Fe-GOCS中的赤铁矿(α-Fe2O3),Fe@GOCS中的针铁矿[α-FeO(OH)]。
-
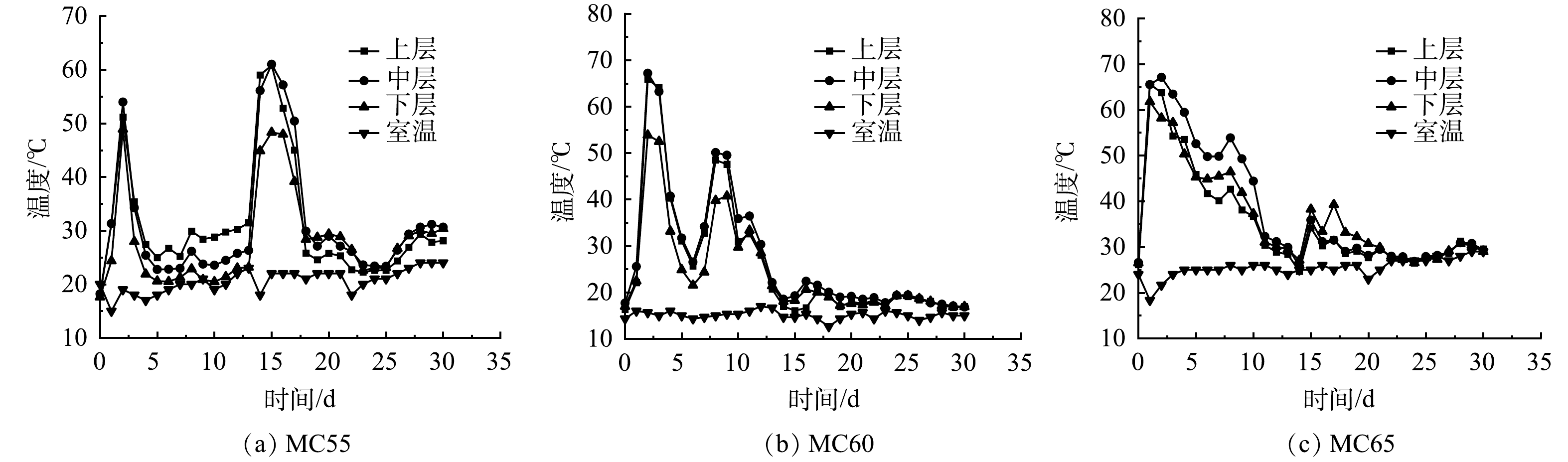
图 2 不同初始含水率对人粪污好氧堆肥过程温度变化的影响
Figure 2. Changes of temperature during human excrement composting with different initial moisture content
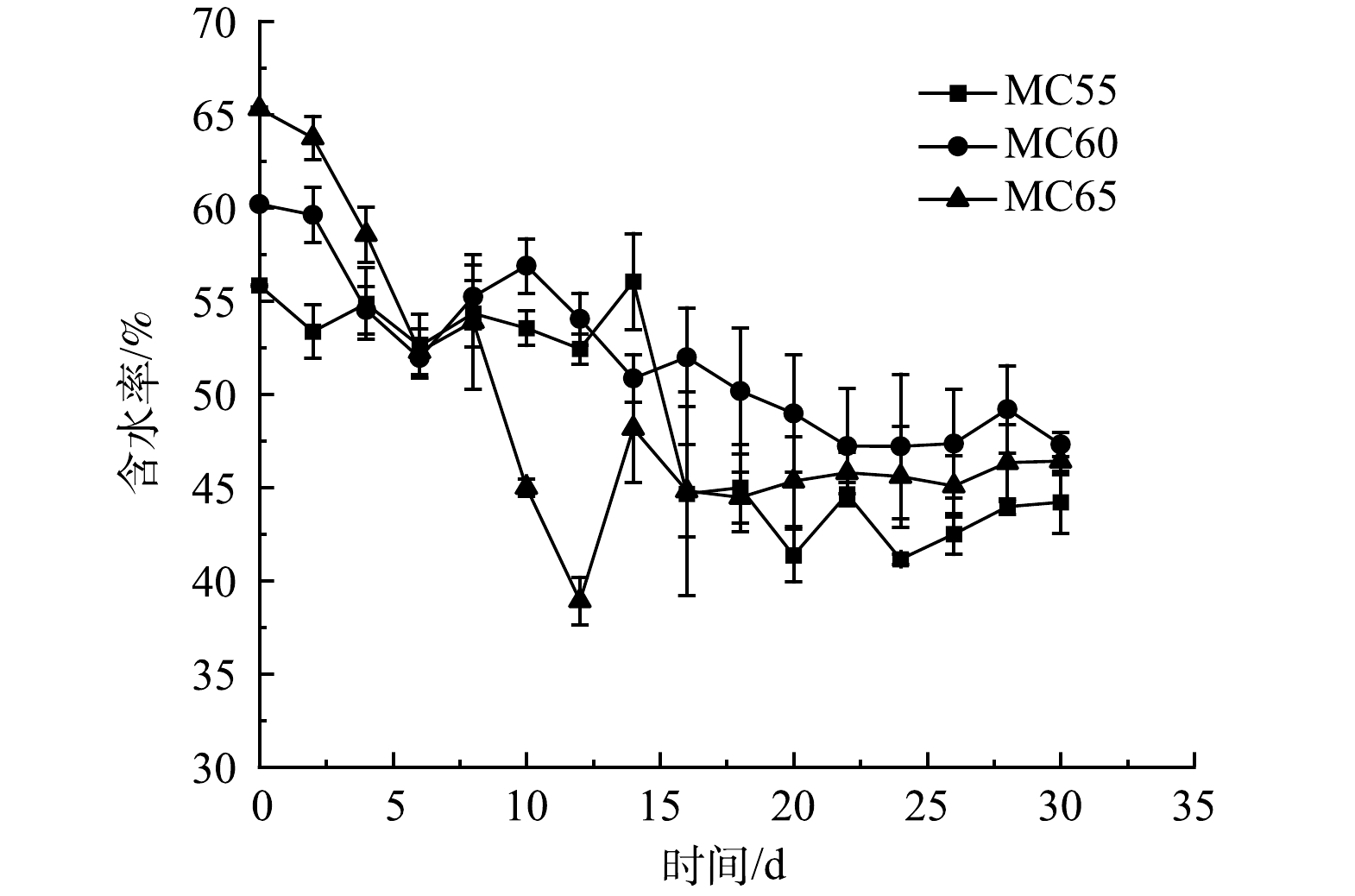
图 3 不同初始含水率对人粪污好氧堆肥过程含水率变化的影响
Figure 3. Changes of moisture during human excrement composting with different initial moisture content
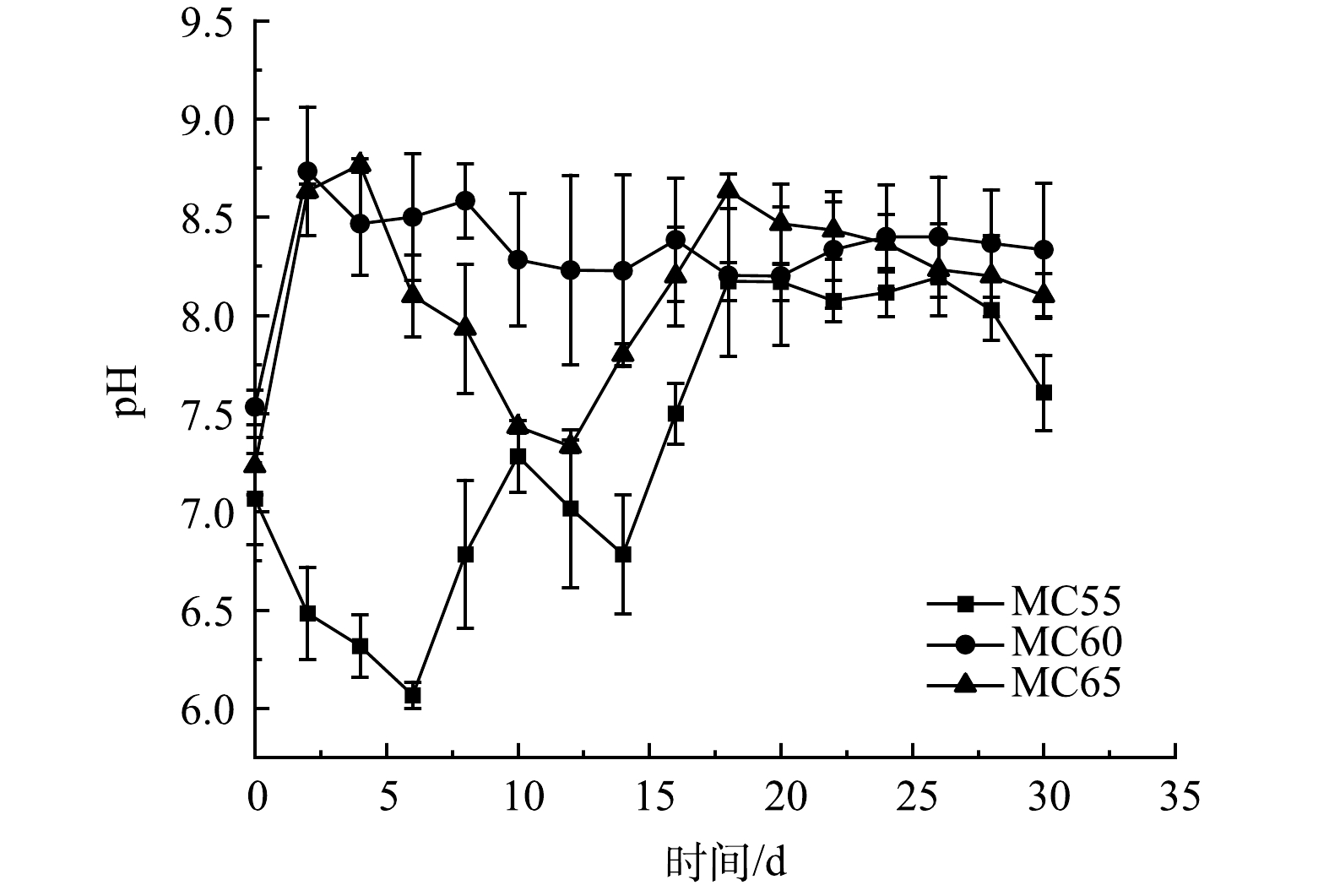
图 4 不同初始含水率对人粪污好氧堆肥过程pH变化的影响
Figure 4. Changes of pH during human excrement composting with different initial moisture content
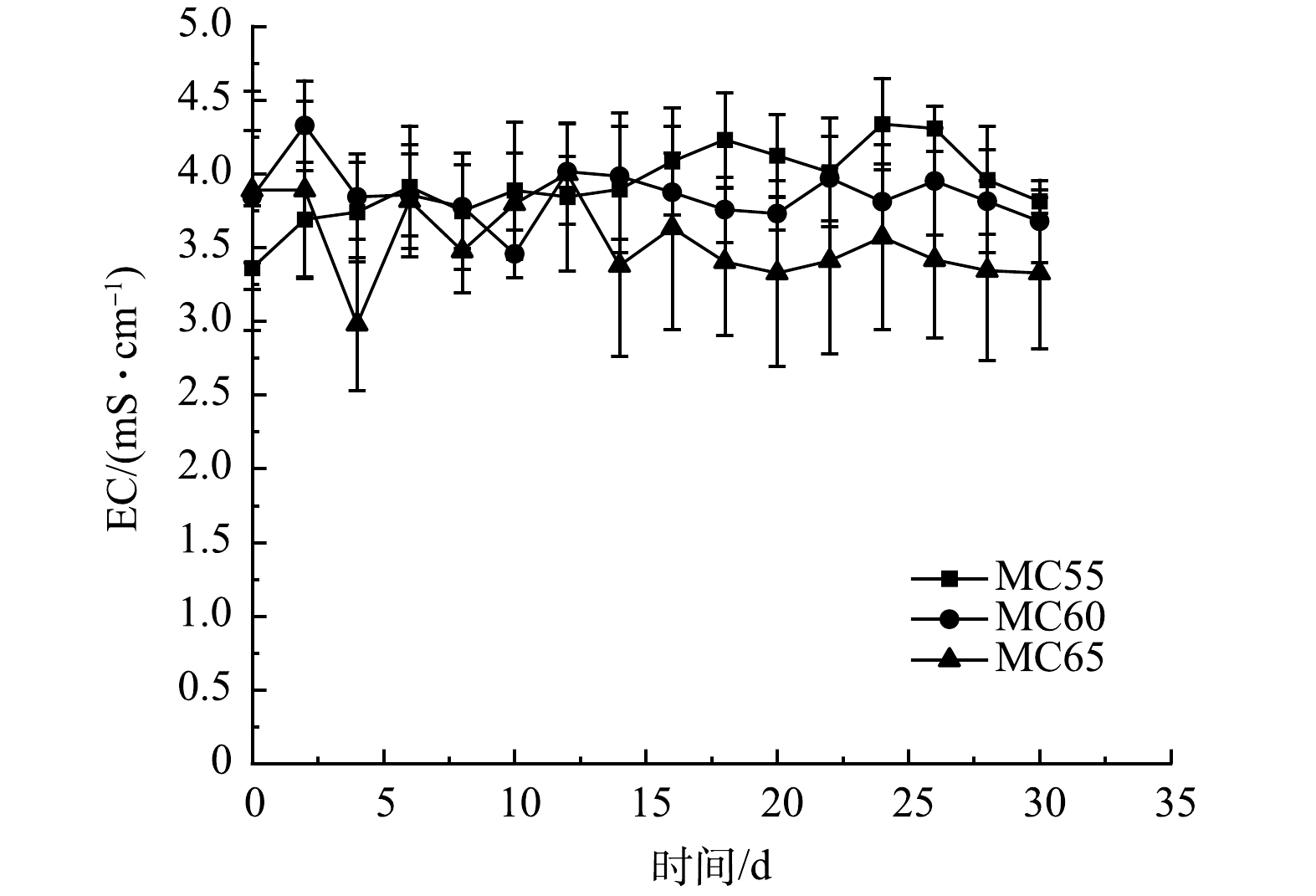
图 5 不同初始含水率对人粪污好氧堆肥过程EC变化的影响
Figure 5. Changes of EC during human excrement composting with different initial moisture content
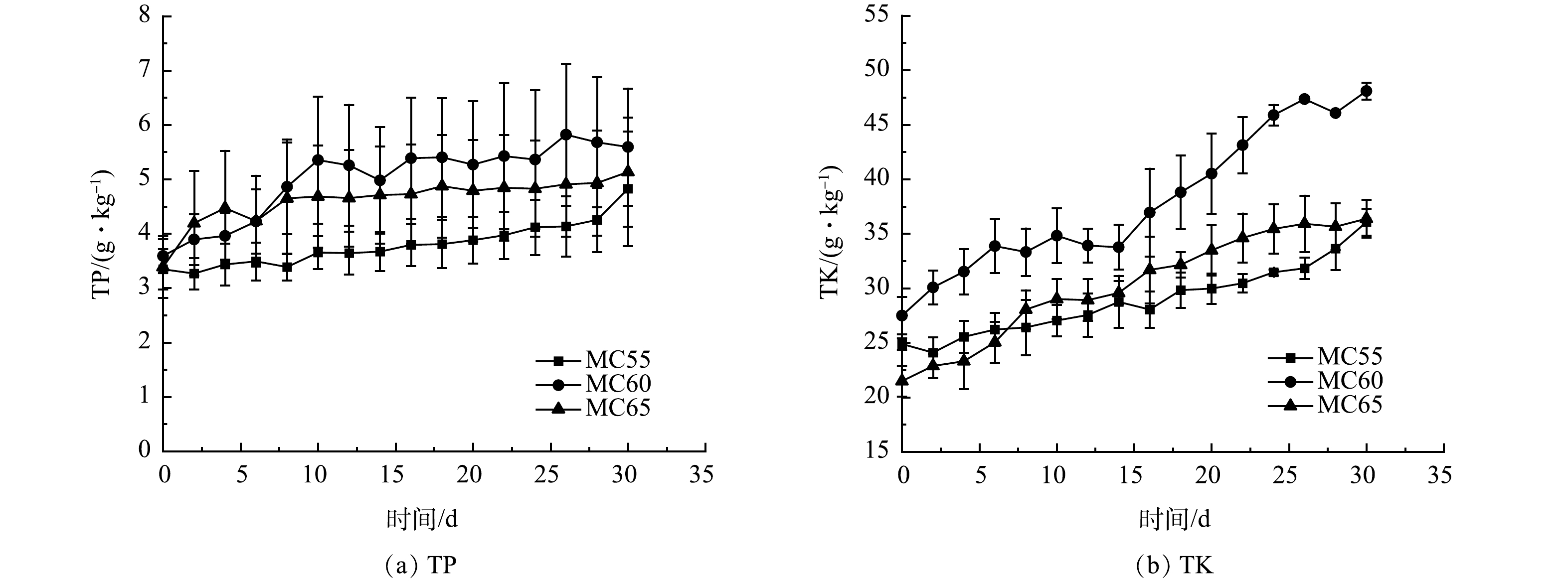
图 6 不同初始含水率对人粪污好氧堆肥过程TP、TK变化的影响
Figure 6. Changes of TP and TK during human excrement composting with different initial moisture content
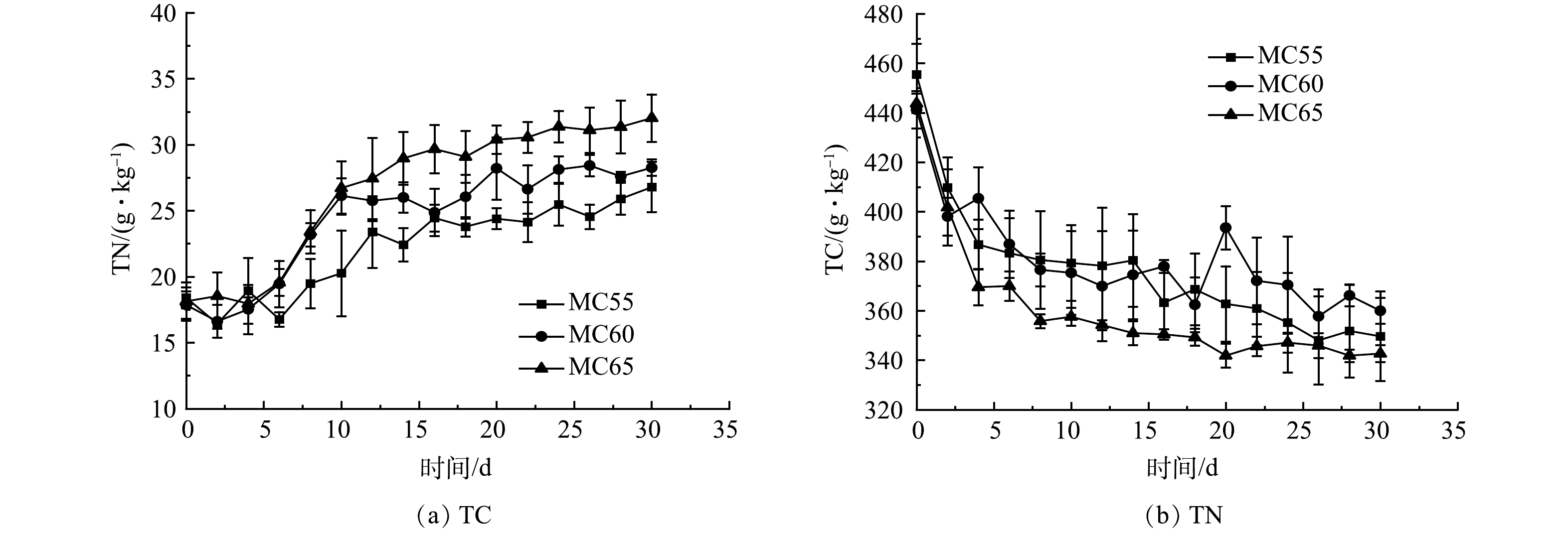
图 7 不同初始含水率对人粪污好氧堆肥过程TC、TN变化的影响
Figure 7. Changes of TC and TN during human excrement composting with different initial moisture content
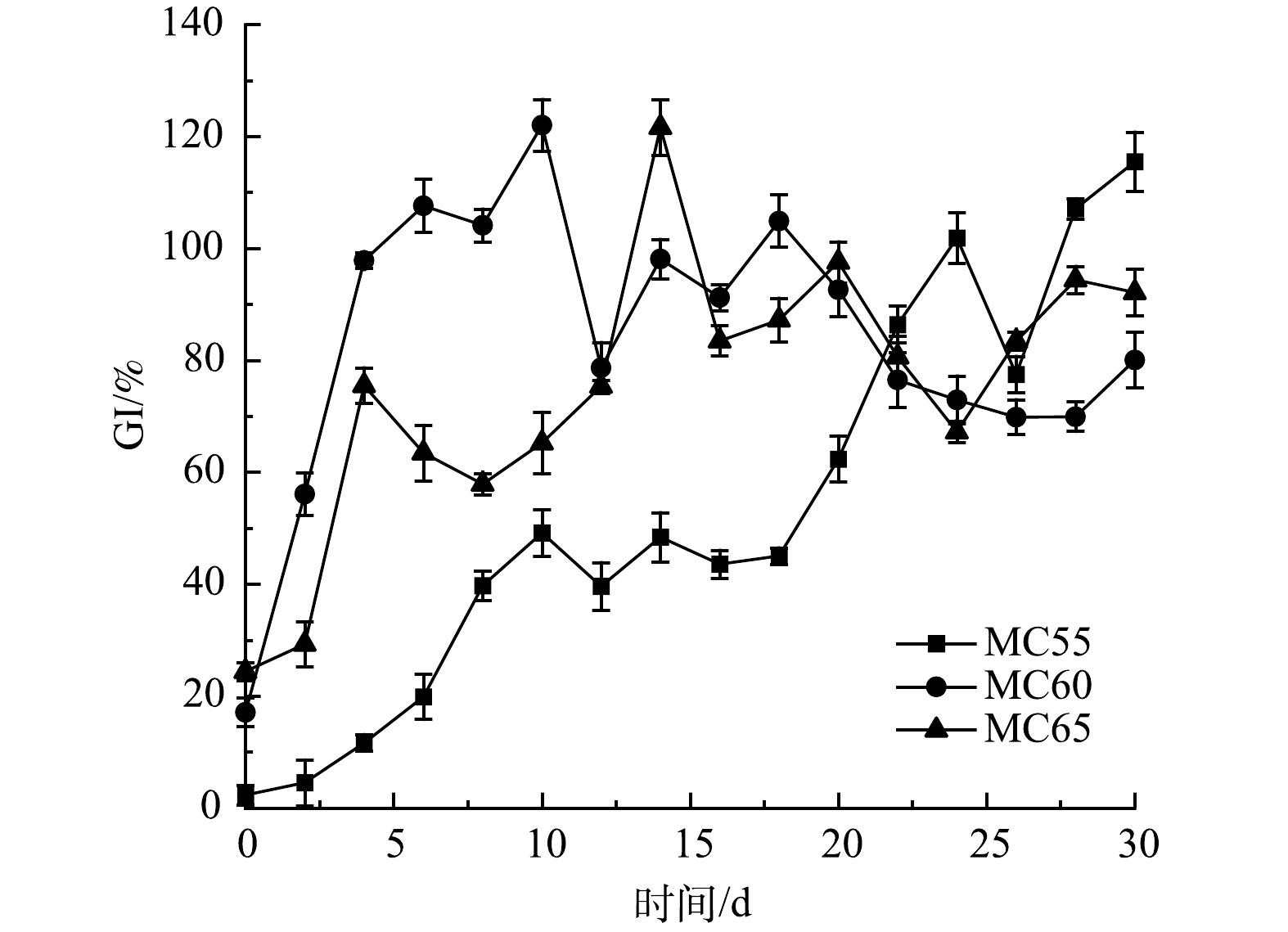
图 8 不同初始含水率对人粪污好氧堆肥过程GI变化的影响
Figure 8. Changes of GI during human excrement composting with different initial moisture content
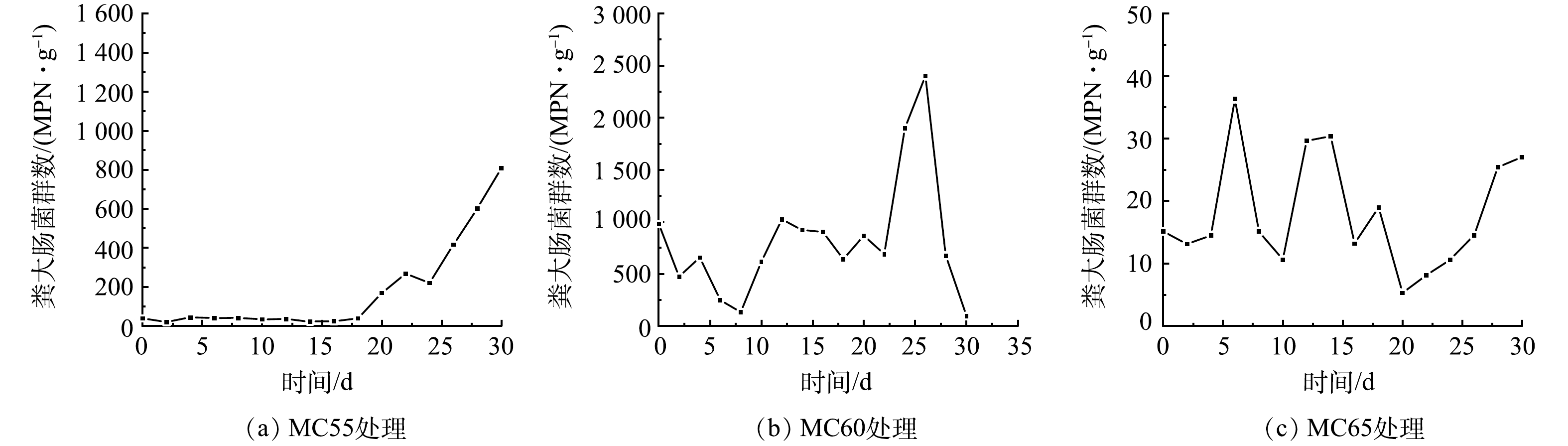
图 9 不同初始含水率对人粪污好氧堆肥过程粪大肠菌群数变化的影响
Figure 9. Changes of the number of fecal coliform during human excrement composting with different initial moisture content

图 10 不同初始含水率人粪污好氧堆肥过程不同阶段稀释性曲线
Figure 10. Rarefaction curve at the different phases of composting process with different initial moisture content
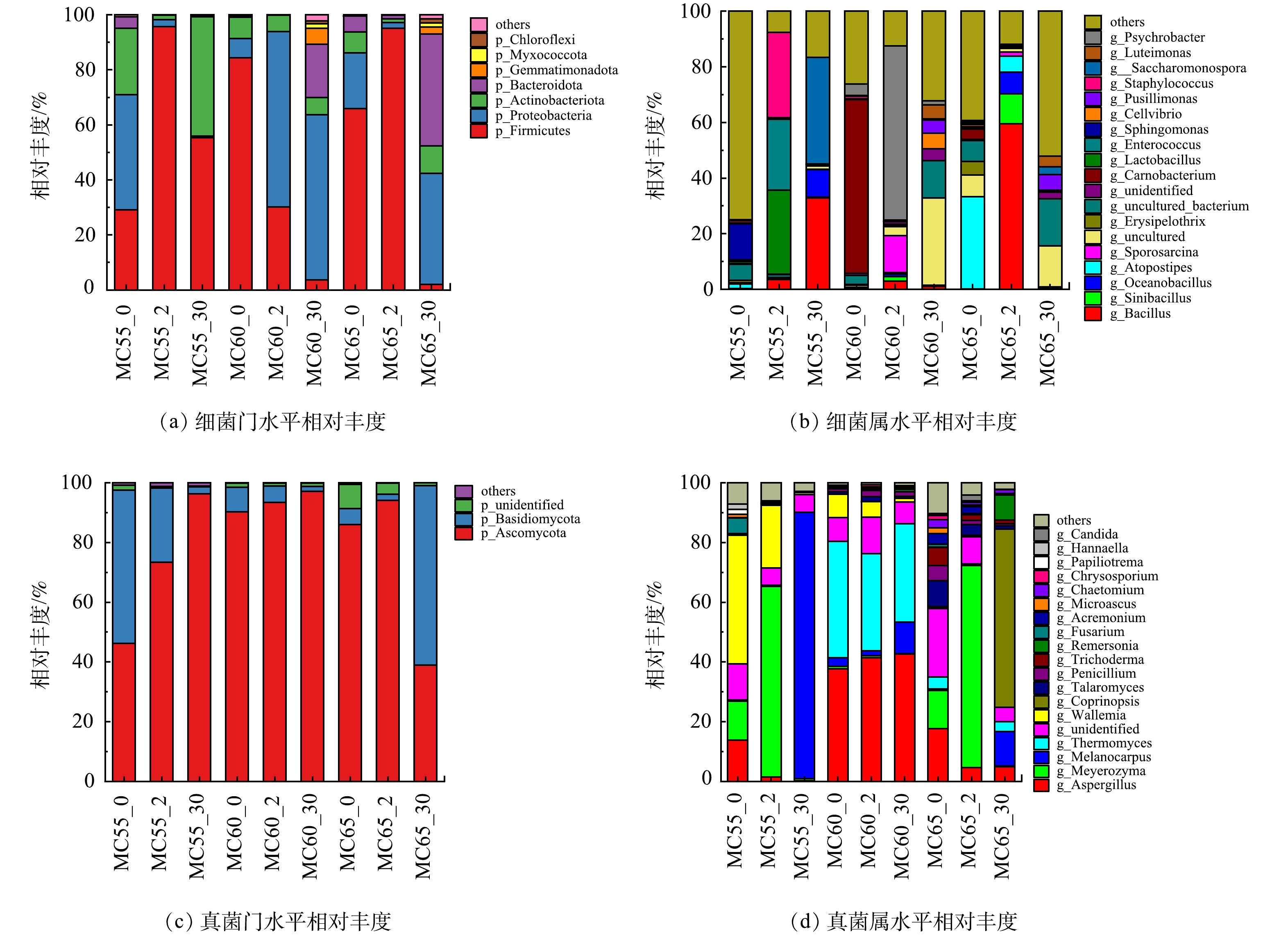
图 11 不同初始含水率人粪污好氧堆肥过程不同阶段细菌和真菌门、属水平相对丰度
Figure 11. Relative abundances of phylum and genus of bacteria and fungi at different phases of human excrement composting with different initial moisture content
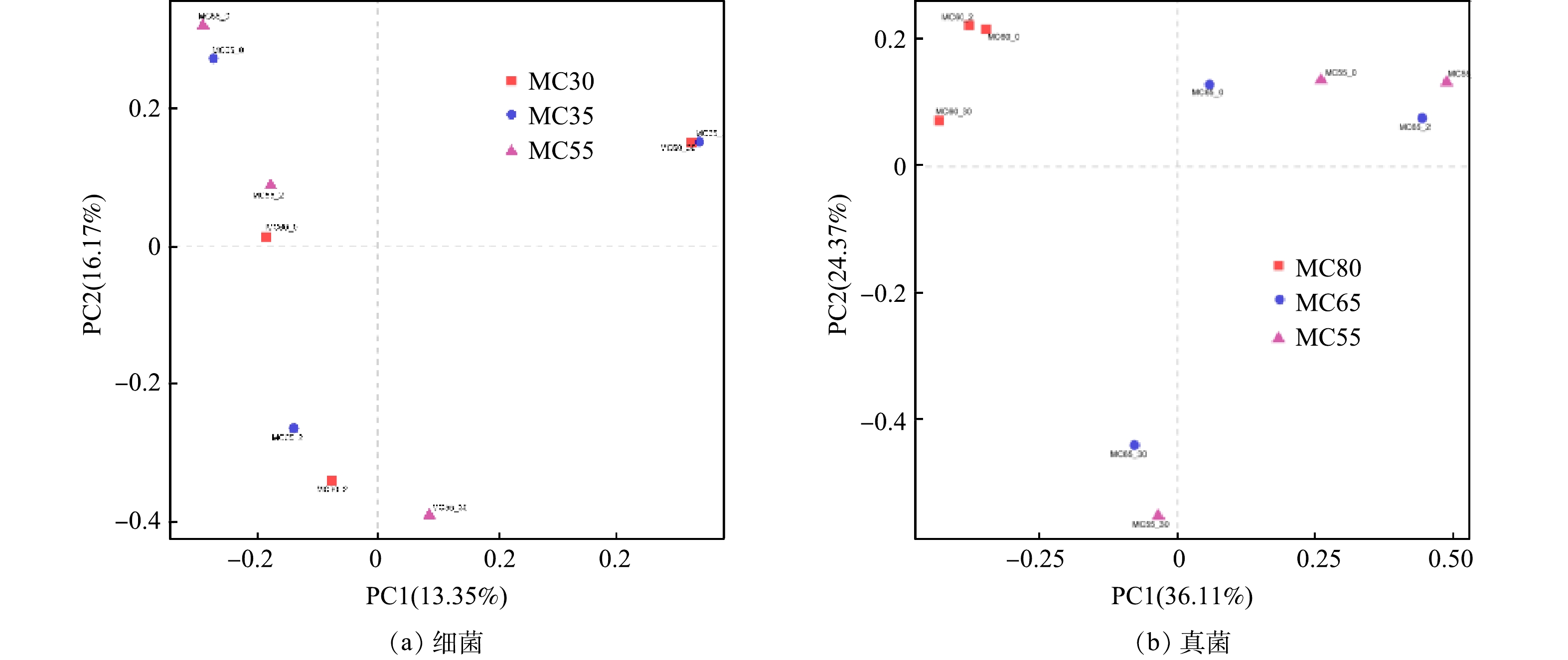
图 12 不同初始含水率人粪污好氧堆肥过程不同阶段细菌和真菌主坐标分析PCoA
Figure 12. Principal coordinate analysis (PCoA) of bacteria and fungi at different phases of human excrement composting with different initial moisture content
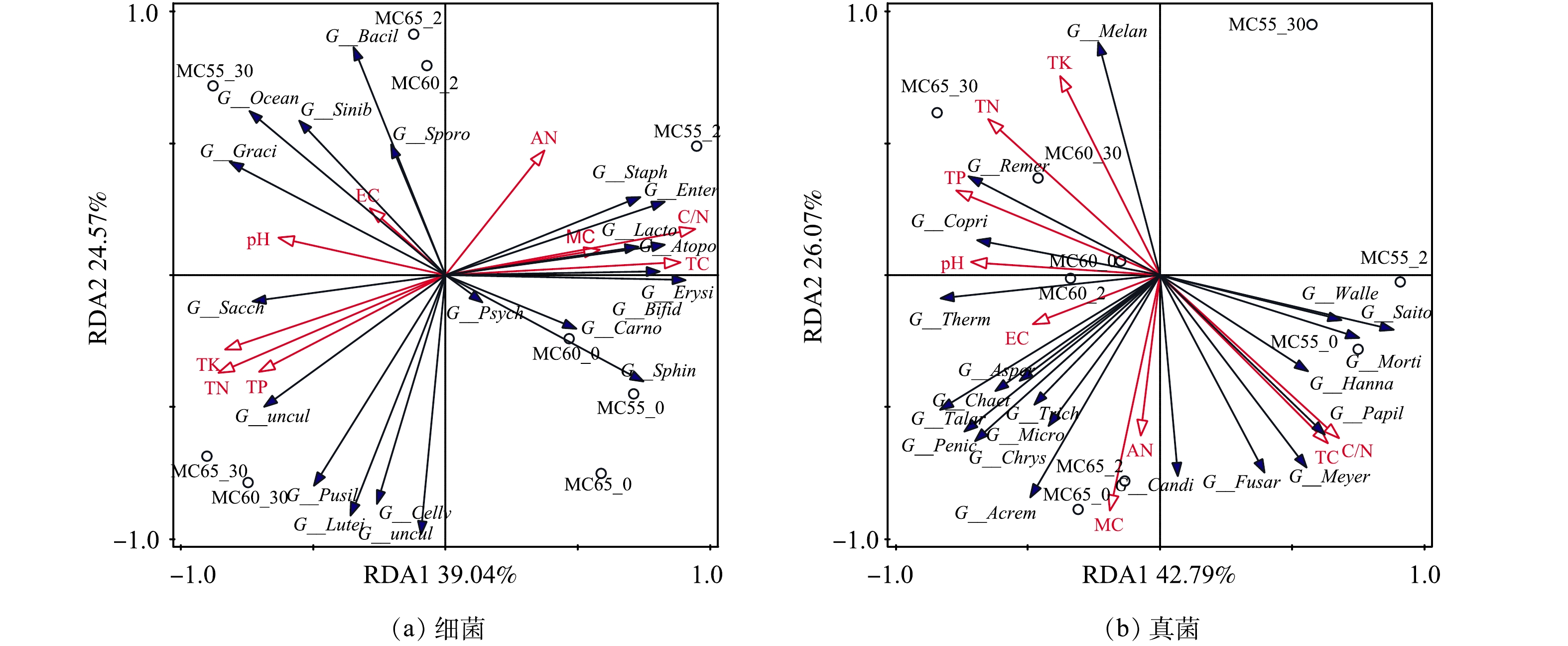
图 13 环境因子与细菌、真菌间的冗余分析 (RDA)
Figure 13. Redundancy Analysis (RDA) between Environmental Factors and Bacteria and Fungi
表 1 原料性质
Table 1. Characteristics of raw materials
原料 含水率/% pH TC/% TN/% C/N 沙门氏菌 蛔虫卵死亡率/% 人粪污 89.69 7.39 43.69 3.48 12.55 未检出 100 玉米秸秆 5.47 7.1 45.02 1.53 29.42 - - 注:“-”代表未检测。
下载: 导出CSV
表 2 不同处理组实验设计方案
Table 2. Experimental design schemes for different treatments
编号 人粪污/kg 玉米秸秆/kg MC/% MC55 14 10 55 MC60 14 7.5 60 MC65 14 5.8 65
下载: 导出CSV
表 3 不同初始含水率人粪污好氧堆肥过程不同阶段细菌和真菌Chao1和Shannon指数
Table 3. Bacteria and fungi Chao1 and Shannonindex at different phases of human excrement composting with different initial moisture content
样品编号 细菌 真菌 Chao1 Shannon Chao1 Shannon MC55-0 676.51 6.71 698.1 4.78 MC55-2 399.49 3.39 566.77 2.67 MC55-30 264.36 3.39 513.8 1.06 MC60-0 637.00 3.51 382.69 3.89 MC60-2 450.14 3.02 371.06 4.26 MC60-30 621.63 5.88 274.28 4.04 MC65-0 769.51 6.16 453.28 6.08 MC65-2 508.12 4.36 365.94 2.99 MC65-30 602.15 6.13 272.94 2.78
下载: 导出CSV
-
[1] 王仁, 宏曹昆. 《乡村振兴战略规划(2018—2022年)》专栏[EB/OL]. [2021-11-22]. http://politics.people.com.cn/n1/2018/0927/c1001-30315372.html. [2] YADAV K D, TARE V, AHAMMED M M. Vermicomposting of source-separated human faeces for nutrient recycling[J]. Waste Management, 2010, 30(1): 50-56. doi: 10.1016/j.wasman.2009.09.034 [3] CHENG S K, LI Z F, UDDIN S M N, et al. Toilet revolution in China[J]. Journal of Environmental Management, 2018, 216: 347-356. [4] SHRESTHA D, SRIVASTAVA A, SHAKYA S M, et al. Use of compost supplemented human urine in sweet pepper (Capsicum annuum L. ) production[J]. Scientia Horticulturae, 2013, 153: 8-12. doi: 10.1016/j.scienta.2013.01.022 [5] COFIE O, KONE D, ROTHENBERGER S, et al. Co-composting of faecal sludge and organic solid waste for agriculture: Process dynamics[J]. Water Research, 2009, 43(18): 4665-4675. doi: 10.1016/j.watres.2009.07.021 [6] VINNERåS B. Comparison of composting, storage and urea treatment for sanitising of faecal matter and manure[J]. Bioresource Technology, 2007, 98(17): 3317-3321. doi: 10.1016/j.biortech.2006.07.011 [7] HASHEMI S, BOUDAGHPOUR S, HAN M. Evaluation of different natural additives effects on the composting process of source separated feces in resource-oriented sanitation systems[J]. Ecotoxicology and Environmental Safety, 2019, 185: 109667. doi: 10.1016/j.ecoenv.2019.109667 [8] LU J W, ZHANG J R, ZHU Z B, et al. Simultaneous production of biocrude oil and recovery of nutrients and metals from human feces via hydrothermal liquefaction[J]. Energy Conversion and Management, 2017, 134: 340-346. doi: 10.1016/j.enconman.2016.12.052 [9] GUO R, LI G X, JIANG T, et al. Effect of aeration rate, C/N ratio and moisture content on the stability and maturity of compost[J]. Bioresource Technology, 2012, 112: 171-178. doi: 10.1016/j.biortech.2012.02.099 [10] 李丹阳, 马若男, 亓传仁, 等. 含水率对羊粪堆肥腐熟度及污染气体排放的影响[J]. 农业工程学报, 2020, 36(20): 254-262. [11] 李季, 彭生平. 堆肥工程实用手册[J]. 第二版. 化学工业出版社, 2011: 30-31. [12] ZAVALA M A L, FUNAMIZU N. Effect of Moisture Content on the Composting Process In a Biotoilet System[J]. Compost Science & Utilization, 2005, 13(3): 208-216. [13] PETRIC I, ŠESTAN A, ŠESTAN I. Influence of initial moisture content on the composting of poultry manure with wheat straw[J]. Biosystems Engineering, 2009, 104(1): 125-134. doi: 10.1016/j.biosystemseng.2009.06.007 [14] LI G, ZHU Q H, NIU Q Q, et al. The degradation of organic matter coupled with the functional characteristics of microbial community during composting with different surfactants[J]. Bioresource Technology, 2021, 321: 124446. doi: 10.1016/j.biortech.2020.124446 [15] GUO Y X, CHEN Q J, QIN Y, et al. Succession of the microbial communities and function prediction during short-term peach sawdust-based composting[J]. Bioresource Technology, 2021, 332: 125079. doi: 10.1016/j.biortech.2021.125079 [16] GE M S, SHEN Y J, DING J T, et al. New insight into the impact of moisture content and pH on dissolved organic matter and microbial dynamics during cattle manure composting[J]. Bioresource Technology, 2022, 344: 126236. doi: 10.1016/j.biortech.2021.126236 [17] SHEN Y J, ZHAO L X, MENG H B, et al. Effect of aeration rate, moisture content and composting period on availability of copper and lead during pig manure composting[J]. Waste Management & Research, 2016, 34(6): 578-583. [18] GE M S, ZHOU H B, SHEN Y J, et al. Effect of aeration rates on enzymatic activity and bacterial community succession during cattle manure composting[J]. Bioresource Technology, 2020, 304: 122928. doi: 10.1016/j.biortech.2020.122928 [19] 中华人民共和国国家质量监督检验检疫总局, 中国国家标准话管理委员会. 肥料中粪大肠菌群的测定: GB/T 19524.1-2004 [S]. 北京: 中国标准出版社, 2004. [20] GAO Y, LI H Y, YANG B, et al. The preliminary evaluation of differential characteristics and factor evaluation of the microbial structure of rural household toilet excrement in China[J]. Environmental Science and Pollution Research, 2021, 28(32): 43842-43852. doi: 10.1007/s11356-021-13779-9 [21] LIU H T, GUO H N, GUO X X, et al. Probing changes in humus chemical characteristics in response to biochar addition and varying bulking agents during composting: A holistic multi-evidence-based approach[J]. Journal of Environmental Management, 2021, 300: 113736. doi: 10.1016/j.jenvman.2021.113736 [22] 中华人民国和国卫生部, 中国国家标准化管理委员会. 粪便无害化卫生要求: GB 7959-2012 [S]. 北京: 中国标准出版社, 2013. [23] SHEN D S, YANG Y Q, HUANG H L, et al. Water state changes during the composting of kitchen waste[J]. Waste Management, 2015, 38: 381-387. doi: 10.1016/j.wasman.2015.01.011 [24] BERNAL M P, ALBURQUERQUE J A, MORAL R. Composting of animal manures and chemical criteria for compost maturity assessment. A review[J]. Bioresource Technology, 2009, 100(22): 5444-5453. doi: 10.1016/j.biortech.2008.11.027 [25] KONG W L, SUN B, ZHANG J Y, et al. Metagenomic analysis revealed the succession of microbiota and metabolic function in corncob composting for preparation of cultivation medium for Pleurotus ostreatus[J]. Bioresource Technology, 2020, 306: 123156. doi: 10.1016/j.biortech.2020.123156 [26] 李相儒. 农村易腐垃圾生物干化与腐熟工艺初探[D]. 杭州: 浙江大学, 2019. [27] 臧冰, 李恕艳, 李国学. 风干预处理对堆肥腐熟度及臭气排放量的影响[J]. 农业工程学报, 2016, 32(S2): 247-253. [28] GARCÍA C, HERNÁNDEZ T, COSTA F. Study on water extract of sewage sludge composts[J]. Soil Science and Plant Nutrition, 2012, 37(3): 399-408. [29] 刘文杰, 王黎明, 沈玉君, 等. 碳氮比对蔬菜废弃物好氧发酵腐熟度及臭气排放的影响[J]. 环境工程, 2020, 38(6): 233-239. doi: 10.13205/j.hjgc.202006038 [30] 李英凯, 李佳丽, 孙溪悦, 等. 添加牛粪和园林废弃物对污泥蚯蚓堆肥的影响[J]. 环境工程学报, 2020, 14(1): 197-208. doi: 10.12030/j.cjee.201903086 [31] 袁京, 刘燕, 唐若兰, 等. 畜禽粪便堆肥过程中碳氮损失及温室气体排放综述[J]. 农业环境科学学报, 2021, 40(11): 2428-2438. doi: 10.11654/jaes.2021-0986 [32] WEI H W, WANG L H, HASSAN M, et al. Succession of the functional microbial communities and the metabolic functions in maize straw composting process[J]. Bioresource Technology, 2018, 256: 333-341. doi: 10.1016/j.biortech.2018.02.050 [33] 中华人民共和国农业农村部. 畜禽粪便堆肥技术规范: NY/T 3442-2019 [S]. 北京: 中国农业出版社, 2019. [34] SHAHEDUZZAMAN M, RAHMAN M S, NUR I T. Influence of temperature on the growth of fecal coliform[J]. Stamford Journal of Microbiology, 2016, 6(1): 463-477. [35] SADEGHI S, NIKAEEN M, MOHAMMADI F, et al. Microbial characteristics of municipal solid waste compost: Occupational and public health risks from surface applied compost[J]. Waste Management, 2022, 144: 98-105. doi: 10.1016/j.wasman.2022.03.012 [36] CHANG H Q, ZHU X H, WU J, et al. Dynamics of microbial diversity during the composting of agricultural straw[J]. Journal of Integrative Agriculture, 2021, 20(5): 1121-1136. doi: 10.1016/S2095-3119(20)63341-X [37] ZHOU G X, XU X F, QIU X W, et al. Biochar influences the succession of microbial communities and the metabolic functions during rice straw composting with pig manure[J]. Bioresource Technology, 2019, 272: 10-18. doi: 10.1016/j.biortech.2018.09.135 [38] MORENO J, LÓPEZ-GONZÁLEZ J A, ARCOS-NIEVAS M A, et al. Revisiting the succession of microbial populations throughout composting: A matter of thermotolerance[J]. Science of The Total Environment, 2021, 773: 145587. doi: 10.1016/j.scitotenv.2021.145587 [39] WANG S P, WANG L, SUN Z Y, et al. Biochar addition reduces nitrogen loss and accelerates composting process by affecting the core microbial community during distilled grain waste composting[J]. Bioresource Technology, 2021, 337: 125492. doi: 10.1016/j.biortech.2021.125492 [40] HERNÁNDEZ-LARA A, ROS M, CUARTERO J, et al. Bacterial and fungal community dynamics during different stages of agro-industrial waste composting and its relationship with compost suppressiveness[J]. Science of The Total Environment, 2022, 805: 150330. doi: 10.1016/j.scitotenv.2021.150330 [41] HUANG R J, WU F, ZHOU Q, et al. Lactobacillus and intestinal diseases: Mechanisms of action and clinical applications[J]. Microbiological Research, 2022, 260: 127019. doi: 10.1016/j.micres.2022.127019 [42] GARCÍA-LÓPEZ M, SANTOS J A, OTERO A, et al. Psychrobacter//BATT C A, TORTORELLO M L[J]. Encyclopedia of Food Microbiology (Second Edition). Oxford:Academic Press, 2014: 261-268. [43] MENG Q X, YANG W, MEN M Q, et al. Microbial Community Succession and Response to Environmental Variables During Cow Manure and Corn Straw Composting[J]. Frontiers in microbiology, 2019, 10: 529. doi: 10.3389/fmicb.2019.00529 [44] LE G O, V B, H B, et al. The microbial signature of aerosols produced during the thermophilic phase of composting[J]. Journal of applied microbiology, 2010, 108(1): 325-340. doi: 10.1111/j.1365-2672.2009.04427.x [45] 马闯, 扈斌, 刘福勇, 等. 有机废弃物好氧堆肥过程中微生物及酶活性变化状况综述[J]. 环境工程, 2019, 37(9): 159-164. [46] LIU T, KUMAR AWASTHI M, JIAO M N, et al. Changes of fungal diversity in fine coal gasification slag amendment pig manure composting[J]. Bioresource Technology, 2021, 325: 124703. doi: 10.1016/j.biortech.2021.124703 [47] AWASTHI S K, DUAN Y M, LIU T, et al. Sequential presence of heavy metal resistant fungal communities influenced by biochar amendment in the poultry manure composting process[J]. Journal of Cleaner Production, 2021, 291: 125947. doi: 10.1016/j.jclepro.2021.125947 [48] VALSALAN R, MATHEW D. Draft genome of Meyerozyma guilliermondii strain vka1: a yeast strain with composting potential[J]. Journal, genetic engineering & biotechnology, 2020, 18(1): 54. [49] ZHANG W M, YU C X, WANG X J, et al. Additives improved saprotrophic fungi for formation of humic acids in chicken manure and corn stover mix composting[J]. Bioresource Technology, 2022, 346: 126626. doi: 10.1016/j.biortech.2021.126626 [50] 车悦驰. 西藏高原污泥好氧堆肥资源化特性与潜力研究[D]. 拉萨: 西藏大学, 2020. [51] DU J J, ZHANG Y Y, QU M X, et al. Effects of biochar on the microbial activity and community structure during sewage sludge composting[J]. Bioresource Technology, 2019, 272: 171-179. doi: 10.1016/j.biortech.2018.10.020 [52] QIAO C, PENTON C R, LIU C, et al. Patterns of fungal community succession triggered by C/N ratios during composting[J]. Journal of Hazardous Materials, 2021, 401: 123344. doi: 10.1016/j.jhazmat.2020.123344 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 6830
- HTML全文浏览数: 6830
- PDF下载数: 65
- 施引文献: 0
