

-
目前全球能源约90%来源于化石燃料[1]。据《BP世界能源展望(2019年版)》预测,2040年天然气需求量为4.62×109 t油当量,能源占比达26%,成为世界第2大能源[2]。截至2016年底,我国已建成沼气工程11.3万座(不含户用),其中大型沼气工程6 737座,超大型34座,沼气工程规模居世界首位[3]。然而,由于存在沼气净化提纯工艺复杂、成本高等问题,产生的沼气难以有效利用,造成能源浪费。H2S是沼气中的主要杂质之一,具有无色、易燃、高毒性的特点,会腐蚀管道,造成二次污染[4]。《GB/T 51063-2014大中型沼气工程技术规范》对沼气中H2S的浓度做出严格限定,要求民用集中供气H2S不超过20 mg·m−3,沼气发电H2S不超过200 mg·m−3。因此,沼气脱硫是沼气应用中亟待解决的问题。
传统的沼气脱硫方法主要有物理-化学法、生物法及电化学法等。物理-化学法脱硫迅速、周期短,但易造成二次污染。生物法脱硫反应条件温和,但处理周期长,运行维护较为复杂。电化学法脱硫适应性强、环境兼容性好,但其能耗较大,维护成本偏高[5]。利用生物电化学法脱硫是将电化学技术与微生物作用相耦合[6],在阳极发生硫氧化反应,产生的电子和质子流向阴极参与还原反应,该工艺具有反应条件温和、脱硫效率高、能耗低等优势,已成为研究热点。李莉等[7]采用微生物燃料电池(microbial fuel cell,MFC)研究了初始pH对S2−和偶氮染料的去除效果,最终得到最大S2−去除率为98.4%。GUO等[8]使用MFC开展了同步去除有机物、S2−和铵的研究,S2−去除率达到100%。CAI等[9]研究了MFC不同进水S/N比对S2−和硝酸盐去除的影响,最终S2−去除率均高于99.70%。NI等[10]使用双室MEC对比分析了阳极有无微生物对废水中S2−脱除的影响,发现生物阳极产生的最大电流密度是非生物阳极的17倍。目前,学者们主要围绕微生物电化学法在S2−脱除的可行性上开展了初步探索,然而在该工艺条件下S2−转化规律和电活性微生物脱硫机理研究方面还鲜有报道。
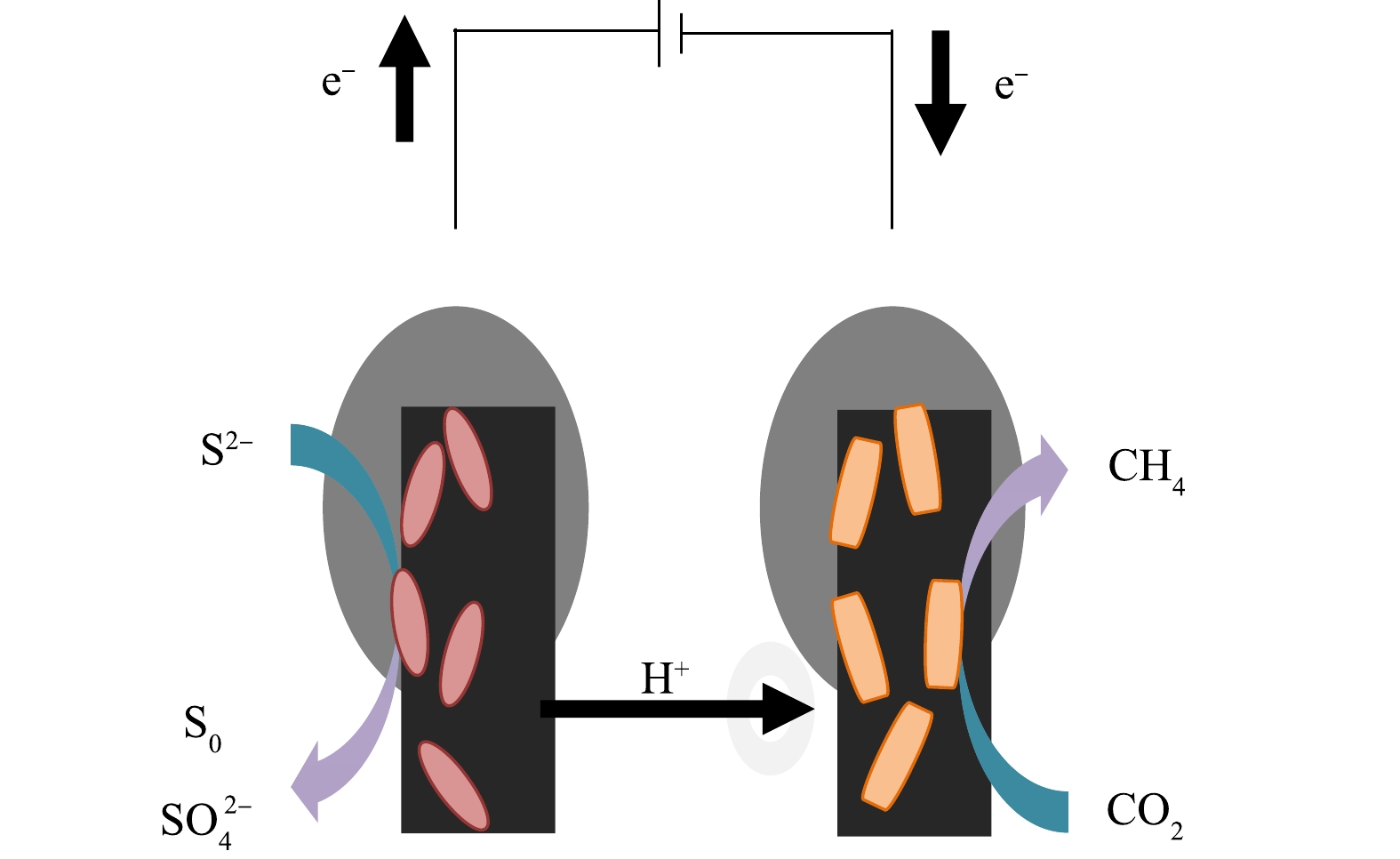
基于上述背景,本研究构建了双室MEC脱硫反应体系,在MEC阳极施加正电压,构建氧化环境,在阳极富集具备胞外电子传递功能的硫氧化菌;在MEC阴极施加负电压,构建还原环境,在阴极富集具有电活性的产甲烷菌。MEC阳极的S2−在硫氧化菌和电极的协同作用下发生氧化反应形成高价态硫,产生的电子和质子流向阴极,在阴极发生CO2转化为甲烷的直接电子传递过程。通过脱硫MEC的启动、运行,开展了S2−去除和硫价态转化规律的研究,结合阳极微生物群落演替,探明了MEC生物阳极脱硫机理,以期为沼气脱硫新工艺的研发提供参考。
-
实验所用的MEC为双室H型(图1),是由2个相同的圆柱体高硼玻璃构成,每室有效容积为200 mL,中间采用质子交换膜(Nafion 117,杜邦)隔开[11],质子交换膜直径为3.6 cm,阳极和阴极电极材料均为的石墨毡,电极尺寸为6 cm×2.8 cm,电极与钛丝连接,反应器上方通过注射器接100 mL金属嘴气袋。反应器采用直流电源(Keithley 2280S-32-6,KEITHLEY)提供外电压,使用数据采集器(Keithley DAQ6510,KEITHLEY)每隔10 min收集1次电压数据。
-
MEC阳极室添加0.6 g·L−1 Na2S·9H2O,同时设置不加S2-的对照实验。实验温度为35 ℃。有研究[12]表明,30~35 ℃为微生物电化学系统中硫化物去除的最佳温度,且35 ℃更利于电活性产甲烷菌的生长[13]。因此,选择该温度可在保持硫化物得到有效去除的情况下便于与阴极产甲烷相耦合。当外加电压为0.8 V、阳极培养液pH为9、阴极培养液pH为7时,MEC两室均接种50 mL厌氧颗粒污泥和150 mL培养液。1 L培养液包括50 mmol·L−1磷酸缓冲溶液、0.31 g NH4Cl、0.13 g KCl、10 mL维生素、10 mL微量元素[14]。阳极室以0.68 g·L−1的CH3COONa·3H2O作为碳源[8,15-16],阴极室以0.84 g·L−1的NaHCO3作为碳源。反应器均采用间歇进料的方式,当监测到反应器外电路电压降至1 mV以下时更换培养液。当连续3个周期电压随时间规律变化、硫化物去除率相近、电流峰值达到1 mA且保持稳定时[17],视为反应器启动成功。启动后的反应器运行1个周期为75 h,其余实验条件与启动期一致。
-
1) S2-浓度采用GB/T16489-1996《水质 硫化物的测定 亚甲基蓝分光光度法》测定。X-射线光电子能谱(XPS)分析:将反应器充分振摇使固体物质均匀分散于溶液中,然后取该悬浮液冻干后作XPS分析。分析仪器为Thermo Scientific K-AlpHa+型光电子能谱分析仪(ThermoFisher 厂家),采用单色化的AlKα射线(1 486.6 eV)作为激发源,分析室的真空度约为5×10−11 mPa。以表面污染C1s结合能284.8 eV作为标准对其他谱线校正。电流密度:利用直流电源(Keithley 2280S-32-6型号)在MEC两端外加电压为0.8 V,MEC阴极与电源负极之间连有外部电阻器(10 Ω),使用数据采集器(Keithley DAQ6510型号)每隔10 min采集1次外电路电阻上的电压。电路电流根据电阻电压通过欧姆定律计算得到,并根据式(1)换算成基于MEC阳极面积的电流密度。
式中:J为电流密度,mA·m−2;U为数据采集器监测到的电压,mV;R为外部电阻,Ω;A为电极面积,m2。
2)微生物群落分析。将石墨毡电极剪碎,反应器悬浮液经冷冻离心机12 000 r·min−1离心15min后在−80 ℃保存,后进行DNA抽提和PCR扩增,使用16S rRNA基因引物338F(5’-ACTCCTACGGGAGGCAGCAG-3’)和806R(5’-GGACTACHVGGGTWTCTAAT-3’)来扩增细菌DNA[18]。每个样本3个重复。使用NEXTflexTM Rapid DNA-Seq Kit(Bioo Scientific,美国)进行建库,利用Illumina公司的Miseq PE300平台进行测序。使用UPARSE软件(
http://drive5.com/uparse/ ,version 7.1),根据97%[19]的相似度对序列进行OTU聚类并剔除嵌合体。利用RDP classifier[20](http://rdp.cme.msu.edu/,version 2.2) 对每条序列进行物种分类注释,比对Silva 16S rRNA数据库(version 138),设置比对阈值为70%。3)生物量密度分析。取1 cm×1 cm样品置于6 mL 0.1 mol·L−1的NaOH溶液中,振荡1 h,以保证电极上的微生物被完全溶解。然后加入2 mL无菌超纯水,振荡清洗10 min,重复3次,共得到12 mL蛋白溶液;将所得蛋白溶液用BCA蛋白定量分析试剂盒测定阳极的生物量密度(Solarbio, Beijing),每组实验做3个平行。
-
脱硫MEC反应器启动与运行过程的电流密度变化如图2所示。反应器在启动阶段最大电流密度呈逐渐上升趋势,当对照组最大电流密度增加到953 mA·m−2、脱硫组最大电流密度增加到581 mA·m−2后,趋于稳定。这可能与电活性微生物在电极上富集有关,此时电极生物膜基本形成,反应器启动成功。在启动阶段,对照组与脱硫组的电流密度随时间均呈周期性变化,反应器运行较为稳定(图2(b))。在脱硫组每次进料初始,有瞬时电流产生,后逐渐降低。这可能是因为S2–的加入使体系内发生了自发的电化学反应[21]。随着乙酸钠及和S2−的氧化,阳极产生的电子流向阴极,电流密度有所升高,由于阳极乙酸钠和S2−逐渐被消耗,电路中流通的电子减少,电流密度达到峰值后也逐渐降低。其中,脱硫组较对照组电流密度持续时间更长,是对照组的2倍左右。这可能是因为S2−的存在使得乙酸盐的消耗变慢。结合第2.2节中硫价态变化结果可知,在1个间歇周期内,硫价态先降低后升高。这说明,硫在MEC阳极先被还原后被氧化,还原过程得电子,氧化过程则释放电子到外电路。因此,外电路电流密度呈现先降后升的趋势,到达最大值后又有所下降。
启动阶段S2−去除率如图3所示。稳定运行阶段,MEC阳极的S2−去除率均达到90%以上,并随着运行时间的延长逐渐提高。在第2、3周期,S2−去除率分别比第1周期分别提高了4.97%和8.83%,第3周期比第2周期提高了3.68%。在第3周期,当反应器运行96 h后,出水中已检测不到S2−。
-
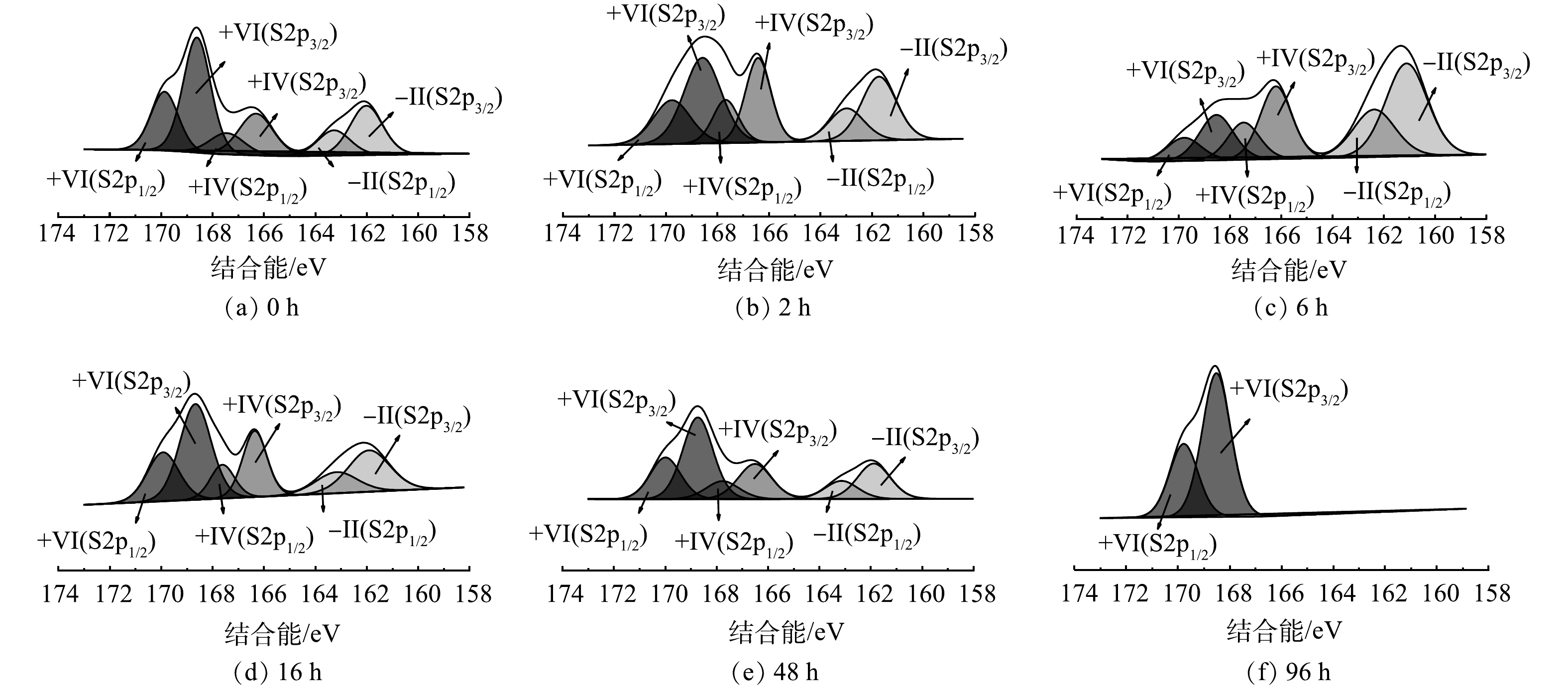
脱硫MEC阳极溶液中硫价态随时间变化的XPS图谱如图4所示。通过对硫元素进行分峰拟合,得到3组双峰。根据已有研究[22-23]及XPS结合能库,可将S2p3/2161.72 eV和S2p1/2162.98 eV归为S2−,可将S2p3/2166.42 eV和S2p1/2167.68 eV归为S4+价,可将S2p3/2168.58 eV和S2p1/2169.77 eV归为S6+。
XPS谱图中各价态硫的峰面积变化如图5所示。在0 h,S2− : S4+ : S6+为0.26 : 0.23 : 0.51;在6 h,S2− : S4+ : S6+为0.51 : 0.29 : 0.2;在96 h,S2−和S4+峰均为分出,S6+占比达到100%。S2−在6 h比0 h提高了96%,而在96 h时降低为0;S2−总体呈现先升高后降低趋势;而S6+则先降低后升高,其在6 h比0 h降低了61%,而在96 h比0 h增加了96%,比6 h增加了4倍。上述结果表明,反应初期S2−增加是由于阳极生物膜上富集的硫酸盐还原菌将接种污泥中少量SO42−还原为S2−,发生还原反应,此时电流密度呈现下降趋势;6 h之后,S2−呈下降趋势,而SO42−呈上升趋势,表明硫氧化菌的活性逐渐增强,从而将S2−氧化为SO42−,释放电子到外电路,此时电流密度也逐渐上升,随着反应的进行达到最大值后下降。在SUN等[24]构建的脱硫MFC反应结束时,S2p的相对峰面积分析结果也表明SO42−为主要的存在形态。
-
1)阳极微生物群落多样性。表1列举了接种污泥、阳极生物膜、阳极溶液的Alpha多样性分析统计数据。结果表明,接种厌氧颗粒污泥的多样性、丰富度最大,微生物经过驯化后,多样性、丰富度均有所降低。这会使菌群功能更专一[10],也表明脱硫MEC阳极的驯化过程对微生物群落结构有重要影响。
2)阳极微生物群落物种差异。对接种污泥、阳极电极、阴极电极、阳极溶液、阴极溶液进行了PCoA分析,以探索群落组成的差异性(图6)。结果表明,接种污泥、电极和溶液中的群落组成差异较明显。这表明当反应器接种厌氧颗粒污泥启动成功后,生物膜和溶液中细菌的群落组成发生了显著的演替变化。此外,电极与溶液的群落组成差异较为明显。这可能是由于石墨毡电极的特殊结构富集了与溶液中不同的微生物[25]。
3)阳极微生物群落演替。接种污泥、阳极生物膜和溶液细菌在门、属上的分布如图7、图8所示。当反应器接种污泥启动后,阳极生物膜和溶液中细菌各种群的丰度发生了明显的变化。由图7可以看出,在门水平上,接种污泥的优势菌群是丰度为16.7%的Chloroflexi和丰度为16.4%的Bacteroidota。Synergistota、Caldatribacteriota和Firmicutes的丰度也相对较高,分别为10.4%、9.4%和8%。这与YANG等[26]的研究结果一致。当反应器接种厌氧颗粒污泥启动成功后,阳极生物膜和溶液中细菌各种群的丰度发生了明显的变化。阳极溶液中的Proteobacteria菌群丰度(56.5%)占绝对优势,其次为Bacteroidota,丰度为19.8%;阳极生物膜上Desulfobacterota丰度最高,为32.7%,其次也为Bacteroidota,其丰度为13.2%。Desulfobacterota为脱硫杆菌门,属于硫还原功能菌群[27]。这也解释了前面提到的阳极出水中SO42-含量在前期上升的现象。
由图8可见,在属水平上,群落分布差异更为显著。阳极生物膜丰度最大的是Trichlorobacter(16.2%),其次是Geobacter(11.3%),该菌属为电活性细菌,其在整个反应体系内起到了传递电子的作用[28],与电流密度的周期性变化关系密切。Pseudomonas在阳极生物膜上丰度排名第4,为7.6%;而该菌属在阳极溶液中丰度排名第2,为15.8%。Pseudomonas为硫氧化功能菌属,能够氧化阳极的S2–[22,29]。该菌属在阳极生物膜和溶液中的丰度分别比接种污泥提高了252倍和526倍。这表明该菌属能够很好地适应MEC体系,以S2–和有机物作为能源,在驯化的过程中迅速富集增加,可有效去除S2–,且最终阳极出水中SO42–上升。
4)阳极生物量密度。采用BCA试剂盒对实验组及对照组阳极生物膜的生物量进行了分析。脱硫MEC阳极生物膜的生物量为(0.18±0.008) mg· cm−2。这说明经过驯化培养,在MEC阳极形成了成熟的生物膜[16]。未加硫的MEC阳极生物膜的生物量为(0.22±0.017) mg· cm−2,略高于脱硫MEC。这说明阳极在硫化钠底物的驯化下菌群功能更专一[10],能有效去除S2−。
-
1) 脱硫MEC反应器经启动和运行后,可在阳极形成稳定的具有高效脱硫功能的生物膜,阳极S2−去除率达90%以上,最大电流密度为658 mA·m−2;阳极硫价态由S2−经S4+最终氧化为S6+,SO42−为阳极S2−氧化的主要产物。
2) Pseudomonas为参与阳极硫氧化的优势功能微生物。该微生物在阳极生物膜和溶液中的相对丰度分别为7.6%和15.8%,分别比接种污泥提高了252倍和526倍。
3) 采用脱硫双室MEC工艺可实现S2−的有效脱除。由于产物以SO42–为主,不会造成电极单质硫沉积,可提高运行寿命,降低运维成本。
脱硫MEC的硫价态变化与微生物群落演替规律
Sulfur valence variety and microbial community succession of desulphurization MEC
-
摘要: 采用微生物电解池(microbial electrolysis cell,MEC)进行沼气脱硫研究,探究了阳极的S2−去除率、硫价态的变化及微生物群落演替规律。结果表明,该方法中S2−去除率可达到90%以上,SO42−为阳极硫氧化反应的主要产物。微生物群落分析结果表明,Pseudomonas为脱硫优势微生物,在阳极生物膜与溶液中的丰度分别为7.6%和15.8%;电活性细菌Geobacter在阳极生物膜与溶液中的丰度分别为11.3%和5.26%,参与了Pseudomonas的种间电子传递,从而提高硫氧化效率。本研究结果可为MEC沼气脱硫工艺提供参考。Abstract: Microbial electrolysis cell (MEC) was used to perform biogas desulfurization in this study. The removal rate of S2−, the variety of different valence of sulfur and the microbial community in MEC anode were analyzed. The results suggested that S2− removal rate was higher than 90% and SO42− was the main product of the anodic sulfur oxidation reaction. The microbial community analysis results showed that Pseudomonas was the predominate bacterial genus for desulfurization. The relative abundance of Pseudomonas in anodic biofilm and anolyte were 7.6% and 15.8%, respectively. The relative abundance of electroactive bacterium Geobacter in anodic biofilm and anolyte were 11.3% and 5.26%, respectively. Geobacter participated in the interspecies electron transfer with Pseudomonas, thus the of sulfur oxidation efficiency increased. The above results can provide technical support for the MEC biogas desulfurization process, which can be combined with MEC biogas decarbonization to achieve biogas purification and upgrading.
-
Key words:
- microbial electrolysis cell /
- biogas desulfurization /
- S2− /
- sulfur valence /
- microbial community
-
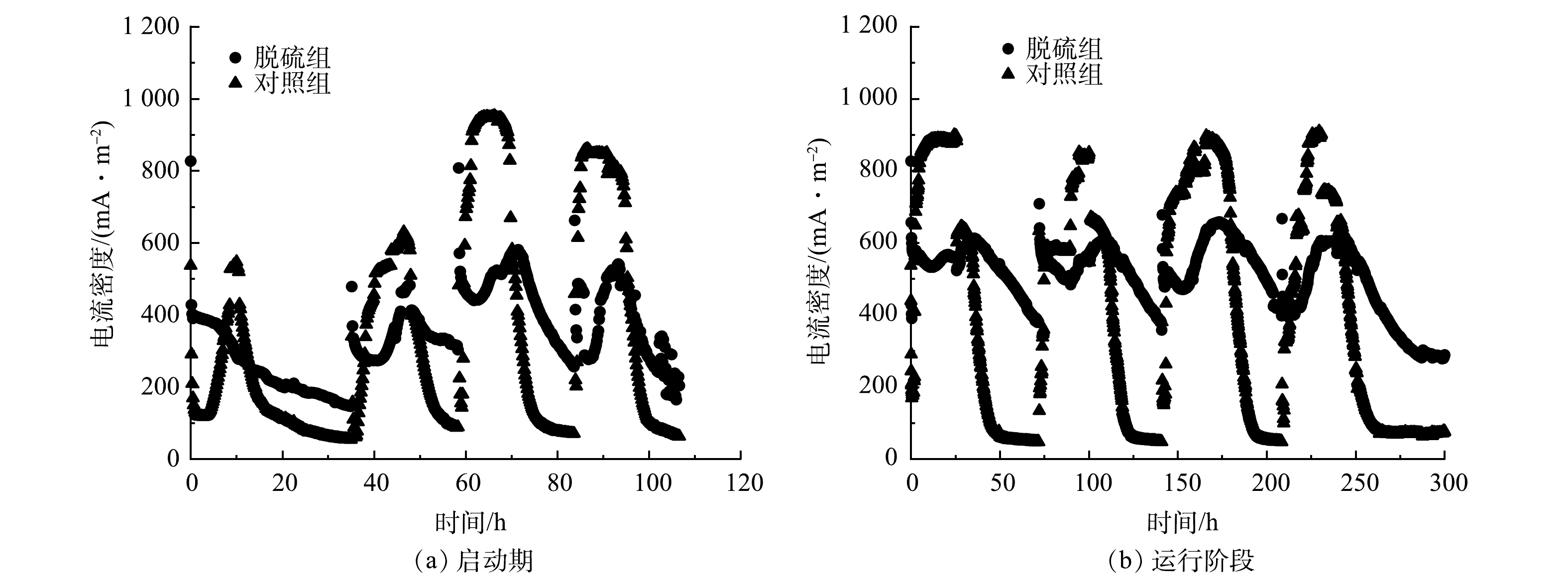
图 2 MEC启动期及运行阶段电流密度随时间的变化
Figure 2. Change of current density during MEC start-up and operation phases
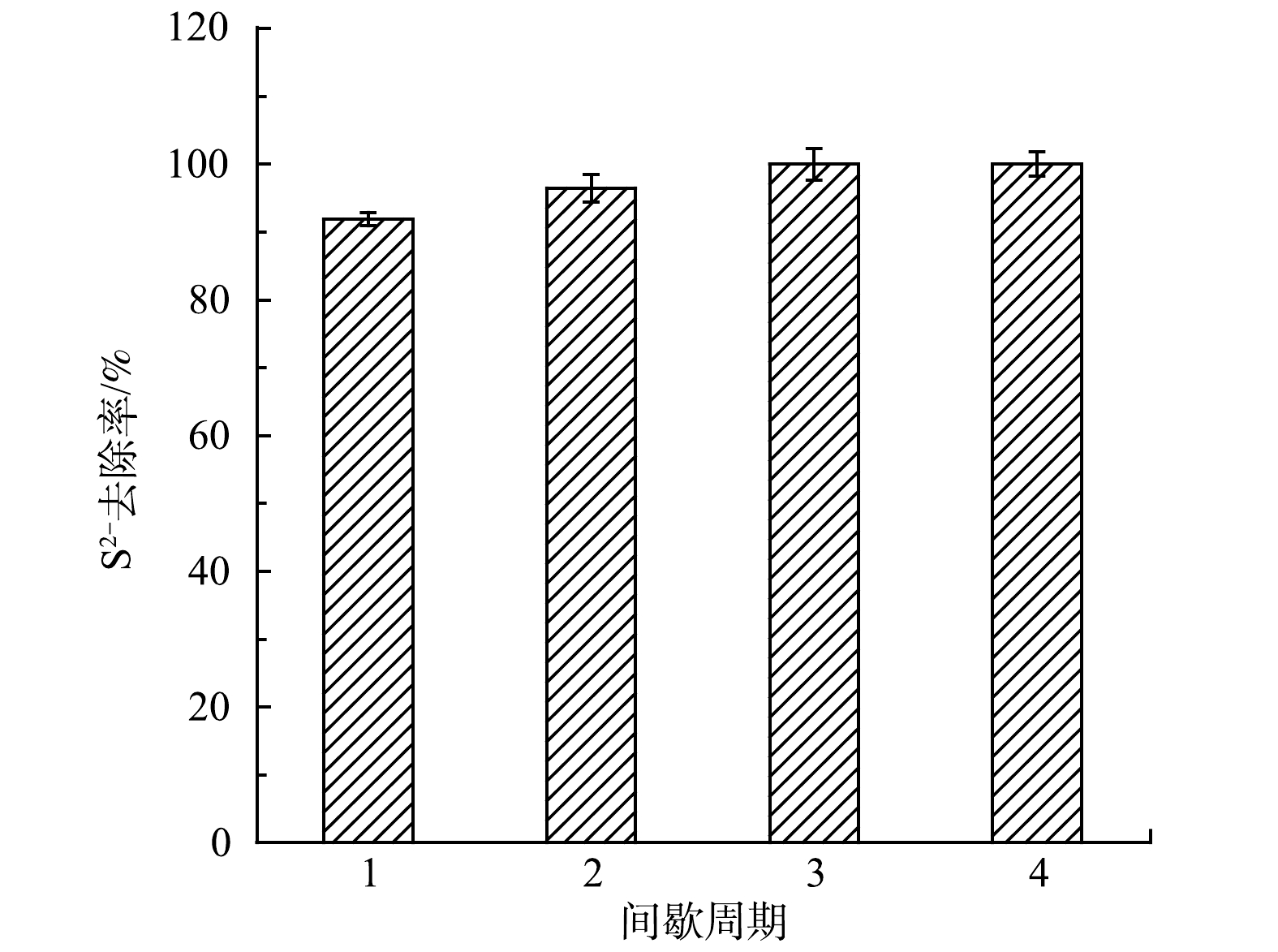
图 3 MEC连续4个周期的S2–去除率
Figure 3. Removal rate of S2– in MEC during four consecutive cycles
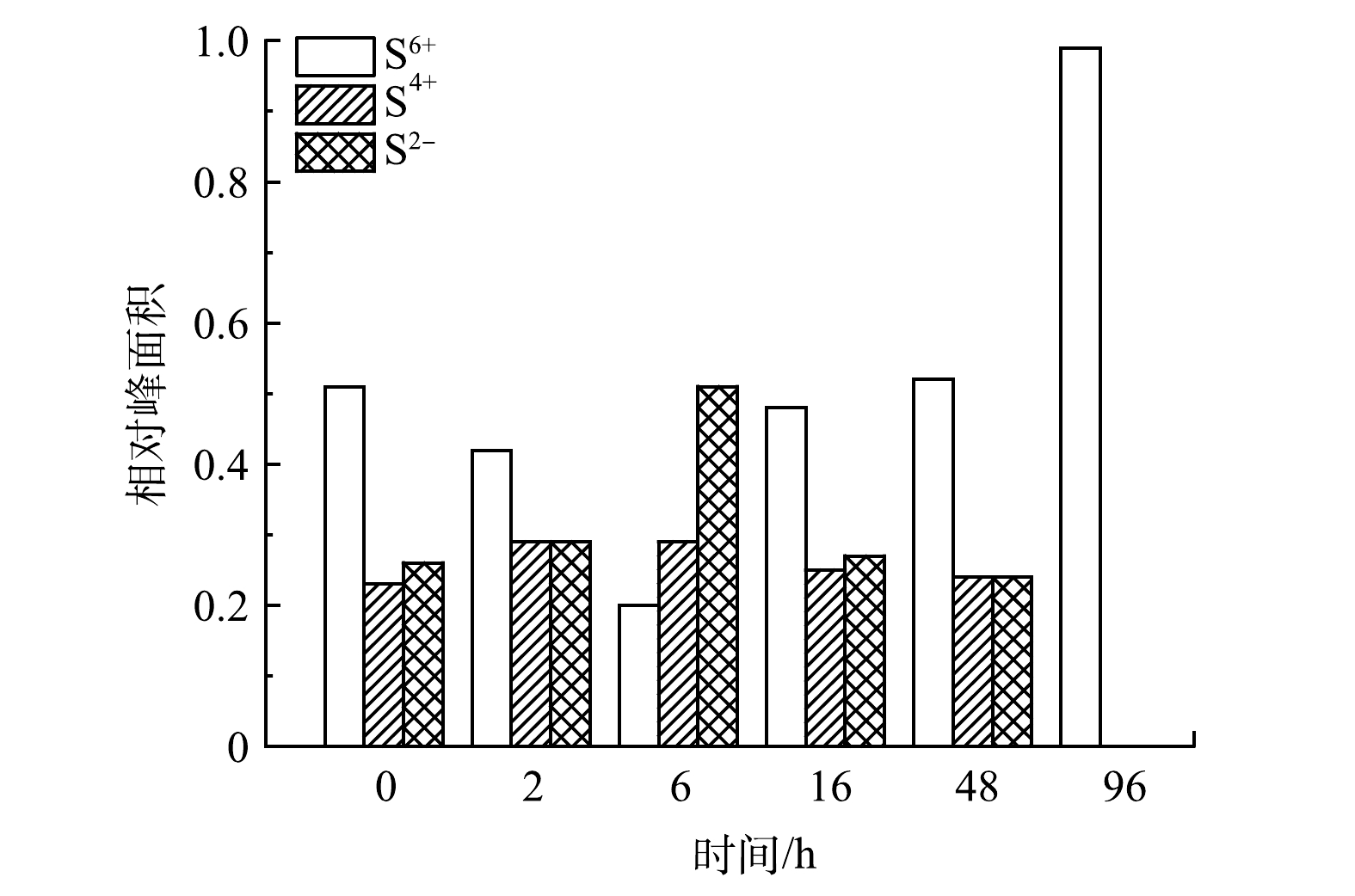
图 5 XPS谱图中各价态硫的相对峰面积变化
Figure 5. Relative peak area variety of different valence sulfur in XPS spectra

图 6 污泥、生物膜及溶液细菌群落的主成分分析
Figure 6. The bacterial community principal component analysis of sludge, biofilm and solution

图 7 门水平上污泥、阳极生物膜及阳极溶液的细菌群落分布
Figure 7. The bacterial community of sludge, anode biofilm and anolyte at the phylum level

图 8 属水平上污泥、阳极生物膜及阳极溶液的细菌群落分布
Figure 8. The bacterial community of sludge, anode biofilm and anolyte at the genus level
表 1 污泥、阳极生物膜、阳极液微生物群落Alpha多样性比较
Table 1. Microbial community by Alpha diversity comparison of sludge, anode biofilm and anolyte
样本 Shannon Simpson Ace Chao 覆盖率/% 接种污泥 4.15 0.04 693.05 720 99.759 9 阳极生物膜 3.59 0.06 448.49 449.02 99.705 2 阳极溶液 2.87 0.1 319.62 302.95 99.795 6  下载: 导出CSV
下载: 导出CSV
-
[1] YOUNAS M, SOHAIL M, LEONG L K, et al. Feasibility of CO2 adsorption by solid adsorbents: A review on low-temperature systems[J]. International Journal of Environmental Science and Technology, 2016, 13(7): 1839-1860. doi: 10.1007/s13762-016-1008-1 [2] 李洪言, 赵朔, 刘飞, 等. 2040年世界能源供需展望: 基于《BP世界能源展望(2019年版)》[J]. 天然气与石油, 2019, 37(6): 1-8. doi: 10.3969/j.issn.1006-5539.2019.06.001 [3] ZHENG L, CHENG S, HAN Y, et al. Bio-natural gas industry in China: Current status and development[J]. Renewable & Sustainable Energy Reviews, 2020, 128: 109925. [4] 武传涛, 韩严和, 符一鸣. 内循环微电解对天然气中H2S的处理及其工艺的优化[J]. 环境工程学报, 2020, 14(3): 721-729. doi: 10.12030/j.cjee.201905175 [5] 蔡靖, 王凯权, 孙月, 等. 生物强化自发电化学氧化含硫废水及关键功能微生物研究[J]. 环境科学学报, 2021, 41(5): 1840-1846. [6] HAMELERS H V M, HEIJNE T A, SLEUTELS T H J A, et al. New applications and performance of bioelectrochemical systems[J]. Applied Microbiology and Biotechnology, 2010, 85(6): 1673-1685. doi: 10.1007/s00253-009-2357-1 [7] 李莉, 代勤, 张赛, 等. 不同pH下微生物燃料电池降解含硫偶氮染料废水的效能及其机理[J]. 环境工程学报, 2021, 15(1): 115-125. doi: 10.12030/j.cjee.202004125 [8] GUO Y L, WEI X, ZHANG S H. Simultaneous removal of organics, sulfide and ammonium coupled with electricity generation in a loop microbial fuel cell system[J]. Bioresource Technology, 2020, 305: 123082. doi: 10.1016/j.biortech.2020.123082 [9] CAI J, QAISAR M, SUN Y, et al. Coupled substrate removal and electricity generation in microbial fuel cells simultaneously treating sulfide and nitrate at various influent sulfide to nitrate ratios[J]. Bioresource Technology, 2020, 306: 123174. doi: 10.1016/j.biortech.2020.123174 [10] NI G, HARNAWAN P, SEIDEL L, et al. Haloalkaliphilic microorganisms assist sulfide removal in a microbial electrolysis cell[J]. Journal of Hazardous Materials, 2019, 363: 197-204. doi: 10.1016/j.jhazmat.2018.09.049 [11] ZEPPILLI M, VILLANO M, MAJONE M. Microbial electrolysis cell to enhance energy recovery from wastewater treatment[J]. Chemical Engineering Transactions, 2015, 43: 2341-2346. [12] ZHANG S H, BAO R, LU J, et al. Simultaneous sulfide removal, nitrification, denitrification and electricity generation in three-chamber microbial fuel cells[J]. Separation and Purification Technology, 2018, 195: 314-321. doi: 10.1016/j.seppur.2017.12.027 [13] FU X, LI J, PAN X, et al. A single microbial electrochemical system for CO2 reduction and simultaneous biogas purification, upgrading and sulfur recovery[J]. Bioresource Technology, 2020, 297: 122448. doi: 10.1016/j.biortech.2019.122448 [14] AHN Y, LOGAN B E. A multi-electrode continuous flow microbial fuel cell with separator electrode assembly design[J]. Applied Microbiology and Biotechnology, 2012, 93(5): 2241-2248. doi: 10.1007/s00253-012-3916-4 [15] DAI Q, ZHANG S, LIU H, et al. Sulfide-mediated azo dye degradation and microbial community analysis in a single-chamber air cathode microbial fuel cell[J]. Bioelectrochemistry, 2020, 131: 107349. doi: 10.1016/j.bioelechem.2019.107349 [16] DYKSTRA C M, PAVLOSTATHIS S G. Hydrogen sulfide affects the performance of a methanogenic bioelectrochemical system for biogas upgrading[J]. Water Research, 2021, 200: 117268. [17] WANG B, LIU W, ZHANG Y, et al. Bioenergy recovery from wastewater accelerated by solar power: Intermittent electro-driving regulation and capacitive storage in biomass[J]. Water Research, 2020, 175: 115696. doi: 10.1016/j.watres.2020.115696 [18] YU J, HUANG Z, WU P, et al. Performance and microbial characterization of two-stage caproate fermentation from fruit and vegetable waste via anaerobic microbial consortia[J]. Bioresource Technology, 2019, 284: 398-405. doi: 10.1016/j.biortech.2019.03.124 [19] EDGAR R C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads[J]. Nature Methods, 2013, 10(10): 996-998. doi: 10.1038/nmeth.2604 [20] WANG Q, GARRITY G M, TIEDJE J M, et al. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy[J]. Applied and Environmental Microbiology, 2007, 73(16): 5261-5267. doi: 10.1128/AEM.00062-07 [21] 孙敏. 微生物燃料电池的功能拓展和机理解析[D]. 合肥: 中国科学技术大学, 2009. [22] SUN M, ZHAI L F, MU Y, et al. Bioelectrochemical element conversion reactions towards generation of energy and value-added chemicals[J]. Progress in Energy and Combustion Science, 2020, 77: 100814. doi: 10.1016/j.pecs.2019.100814 [23] IKUMAPAY F, MAKITATO M, JOHANSSON B, et al. Recycling of process water in sulphide flotation: Effect of calcium and sulphate ions on flotation of galena[J]. Minerals Engineering, 2012, 39: 77-88. doi: 10.1016/j.mineng.2012.07.016 [24] SUN M, MU Z X, CHEN Y P, et al. Microbe-assisted sulfide oxidation in the anode of a microbial fuel cell[J]. Environmental Science & Technology, 2009, 43(9): 3372-3377. [25] ZHEN G, LU X, KUMAR G, et al. Microbial electrolysis cell platform for simultaneous waste biorefinery and clean electrofuels generation: Current situation, challenges and future perspectives[J]. Progress in Energy and Combustion Science, 2017, 63: 119-145. doi: 10.1016/j.pecs.2017.07.003 [26] YANG H, BAO B, LIU J, et al. Temperature dependence of bioelectrochemical CO2 conversion and methane production with a mixed-culture biocathode[J]. Bioelectrochemistry, 2018, 119: 180-188. doi: 10.1016/j.bioelechem.2017.10.002 [27] PANG Y, GU T, ZHANG G, et al. Experimental study on volatile sulfur compound inhibition using a single-chamber membrane-free microbial electrolysis cell[J]. Environmental Science and Pollution Research International, 2020, 27(24): 30571-30582. doi: 10.1007/s11356-020-09325-8 [28] LOGAN B E, ROSSI R, RAGAB A, et al. Electroactive microorganisms in bioelectrochemical systems[J]. Nature Reviews Microbiology, 2019, 17(5): 307-319. doi: 10.1038/s41579-019-0173-x [29] DENG H, CHEN Z, ZHAO F. Energy from plants and microorganisms: Progress in plant-microbial fuel cells[J]. ChemSusChem, 2012, 5(6): 1006-1011. doi: 10.1002/cssc.201100257 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4620
- HTML全文浏览数: 4620
- PDF下载数: 92
- 施引文献: 0
