




 下载:
下载:

</td><td class="table_top_border" align="center" valign="middle"><inline-formula><tex-math id="M4">${\rm{NH}}_4^ + $</tex-math><img class="inline-formula" style="display:none;" src="data:image/svg+xml,<svg xmlns='http://www.w3.org/2000/svg' width='350' height='600'><foreignObject width='2000' height='100%'><div xmlns='http://www.w3.org/1999/xhtml' style='font-size:16px;'><table>
<thead><tr><td class="table_top_border" align="center" valign="middle">数值类型</td><td class="table_top_border" align="center" valign="middle">COD/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle"><inline-formula><tex-math id="M4">${\rm{NH}}_4^ + $</tex-math><alternatives><img class="graphic" src="202010033-heliang_M4.jpg"><img class="graphic" src="202010033-heliang_M4.png"></alternatives></inline-formula>-N/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle"><inline-formula><tex-math id="M5">${\rm{NO}}_3^ - $</tex-math><alternatives><img class="graphic" src="202010033-heliang_M5.jpg"><img class="graphic" src="202010033-heliang_M5.png"></alternatives></inline-formula>-N/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle"><inline-formula><tex-math id="M6">${\rm{NO}}_2^ - $</tex-math><alternatives><img class="graphic" src="202010033-heliang_M6.jpg"><img class="graphic" src="202010033-heliang_M6.png"></alternatives></inline-formula>-N/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle">TN/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle">pH</td></tr></thead>
<tbody><tr><td class="table_top_border2" align="center" valign="middle">平均值</td><td class="table_top_border2" align="center" valign="middle">3 588.54</td><td class="table_top_border2" align="center" valign="middle">1 615.41</td><td class="table_top_border2" align="center" valign="middle">36.42</td><td class="table_top_border2" align="center" valign="middle">0.11</td><td class="table_top_border2" align="center" valign="middle">1 991.21</td><td class="table_top_border2" align="center" valign="middle">8.13</td></tr><tr><td align="center" valign="middle">最大值</td><td align="center" valign="middle">5 600.84</td><td align="center" valign="middle">1 722.24</td><td align="center" valign="middle">48.27</td><td align="center" valign="middle">0.28</td><td align="center" valign="middle">2 198.51</td><td align="center" valign="middle">8.61</td></tr><tr><td class="table_bottom_border" align="center" valign="middle">最小值</td><td class="table_bottom_border" align="center" valign="middle">2 096.62</td><td class="table_bottom_border" align="center" valign="middle">1 057.56</td><td class="table_bottom_border" align="center" valign="middle">14.19</td><td class="table_bottom_border" align="center" valign="middle">未检出</td><td class="table_bottom_border" align="center" valign="middle">1 763.11</td><td class="table_bottom_border" align="center" valign="middle">7.51</td></tr></tbody>
</table></div></foreignObject></svg>"></inline-formula>-N/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle"><inline-formula><tex-math id="M5">${\rm{NO}}_3^ - $</tex-math><img class="inline-formula" style="display:none;" src="data:image/svg+xml,<svg xmlns='http://www.w3.org/2000/svg' width='350' height='600'><foreignObject width='2000' height='100%'><div xmlns='http://www.w3.org/1999/xhtml' style='font-size:16px;'><table>
<thead><tr><td class="table_top_border" align="center" valign="middle">数值类型</td><td class="table_top_border" align="center" valign="middle">COD/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle"><inline-formula><tex-math id="M4">${\rm{NH}}_4^ + $</tex-math><alternatives><img class="graphic" src="202010033-heliang_M4.jpg"><img class="graphic" src="202010033-heliang_M4.png"></alternatives></inline-formula>-N/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle"><inline-formula><tex-math id="M5">${\rm{NO}}_3^ - $</tex-math><alternatives><img class="graphic" src="202010033-heliang_M5.jpg"><img class="graphic" src="202010033-heliang_M5.png"></alternatives></inline-formula>-N/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle"><inline-formula><tex-math id="M6">${\rm{NO}}_2^ - $</tex-math><alternatives><img class="graphic" src="202010033-heliang_M6.jpg"><img class="graphic" src="202010033-heliang_M6.png"></alternatives></inline-formula>-N/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle">TN/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle">pH</td></tr></thead>
<tbody><tr><td class="table_top_border2" align="center" valign="middle">平均值</td><td class="table_top_border2" align="center" valign="middle">3 588.54</td><td class="table_top_border2" align="center" valign="middle">1 615.41</td><td class="table_top_border2" align="center" valign="middle">36.42</td><td class="table_top_border2" align="center" valign="middle">0.11</td><td class="table_top_border2" align="center" valign="middle">1 991.21</td><td class="table_top_border2" align="center" valign="middle">8.13</td></tr><tr><td align="center" valign="middle">最大值</td><td align="center" valign="middle">5 600.84</td><td align="center" valign="middle">1 722.24</td><td align="center" valign="middle">48.27</td><td align="center" valign="middle">0.28</td><td align="center" valign="middle">2 198.51</td><td align="center" valign="middle">8.61</td></tr><tr><td class="table_bottom_border" align="center" valign="middle">最小值</td><td class="table_bottom_border" align="center" valign="middle">2 096.62</td><td class="table_bottom_border" align="center" valign="middle">1 057.56</td><td class="table_bottom_border" align="center" valign="middle">14.19</td><td class="table_bottom_border" align="center" valign="middle">未检出</td><td class="table_bottom_border" align="center" valign="middle">1 763.11</td><td class="table_bottom_border" align="center" valign="middle">7.51</td></tr></tbody>
</table></div></foreignObject></svg>"></inline-formula>-N/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle"><inline-formula><tex-math id="M6">${\rm{NO}}_2^ - $</tex-math><img class="inline-formula" style="display:none;" src="data:image/svg+xml,<svg xmlns='http://www.w3.org/2000/svg' width='350' height='600'><foreignObject width='2000' height='100%'><div xmlns='http://www.w3.org/1999/xhtml' style='font-size:16px;'><table>
<thead><tr><td class="table_top_border" align="center" valign="middle">数值类型</td><td class="table_top_border" align="center" valign="middle">COD/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle"><inline-formula><tex-math id="M4">${\rm{NH}}_4^ + $</tex-math><alternatives><img class="graphic" src="202010033-heliang_M4.jpg"><img class="graphic" src="202010033-heliang_M4.png"></alternatives></inline-formula>-N/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle"><inline-formula><tex-math id="M5">${\rm{NO}}_3^ - $</tex-math><alternatives><img class="graphic" src="202010033-heliang_M5.jpg"><img class="graphic" src="202010033-heliang_M5.png"></alternatives></inline-formula>-N/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle"><inline-formula><tex-math id="M6">${\rm{NO}}_2^ - $</tex-math><alternatives><img class="graphic" src="202010033-heliang_M6.jpg"><img class="graphic" src="202010033-heliang_M6.png"></alternatives></inline-formula>-N/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle">TN/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle">pH</td></tr></thead>
<tbody><tr><td class="table_top_border2" align="center" valign="middle">平均值</td><td class="table_top_border2" align="center" valign="middle">3 588.54</td><td class="table_top_border2" align="center" valign="middle">1 615.41</td><td class="table_top_border2" align="center" valign="middle">36.42</td><td class="table_top_border2" align="center" valign="middle">0.11</td><td class="table_top_border2" align="center" valign="middle">1 991.21</td><td class="table_top_border2" align="center" valign="middle">8.13</td></tr><tr><td align="center" valign="middle">最大值</td><td align="center" valign="middle">5 600.84</td><td align="center" valign="middle">1 722.24</td><td align="center" valign="middle">48.27</td><td align="center" valign="middle">0.28</td><td align="center" valign="middle">2 198.51</td><td align="center" valign="middle">8.61</td></tr><tr><td class="table_bottom_border" align="center" valign="middle">最小值</td><td class="table_bottom_border" align="center" valign="middle">2 096.62</td><td class="table_bottom_border" align="center" valign="middle">1 057.56</td><td class="table_bottom_border" align="center" valign="middle">14.19</td><td class="table_bottom_border" align="center" valign="middle">未检出</td><td class="table_bottom_border" align="center" valign="middle">1 763.11</td><td class="table_bottom_border" align="center" valign="middle">7.51</td></tr></tbody>
</table></div></foreignObject></svg>"></inline-formula>-N/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle">TN/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle">pH</td></tr></thead>
<tbody><tr><td class="table_top_border2" align="center" valign="middle">平均值</td><td class="table_top_border2" align="center" valign="middle">3 588.54</td><td class="table_top_border2" align="center" valign="middle">1 615.41</td><td class="table_top_border2" align="center" valign="middle">36.42</td><td class="table_top_border2" align="center" valign="middle">0.11</td><td class="table_top_border2" align="center" valign="middle">1 991.21</td><td class="table_top_border2" align="center" valign="middle">8.13</td></tr><tr><td align="center" valign="middle">最大值</td><td align="center" valign="middle">5 600.84</td><td align="center" valign="middle">1 722.24</td><td align="center" valign="middle">48.27</td><td align="center" valign="middle">0.28</td><td align="center" valign="middle">2 198.51</td><td align="center" valign="middle">8.61</td></tr><tr><td class="table_bottom_border" align="center" valign="middle">最小值</td><td class="table_bottom_border" align="center" valign="middle">2 096.62</td><td class="table_bottom_border" align="center" valign="middle">1 057.56</td><td class="table_bottom_border" align="center" valign="middle">14.19</td><td class="table_bottom_border" align="center" valign="middle">未检出</td><td class="table_bottom_border" align="center" valign="middle">1 763.11</td><td class="table_bottom_border" align="center" valign="middle">7.51</td></tr></tbody>
</table></div></foreignObject></svg>)





-
很长时间以来,我国城市垃圾的处理主要是以卫生填埋为主[1]。相对于其他处理方式来说,卫生填埋处理成本低,操作简便[2]。但长年的实践经验表明,填埋不仅会占用大量土地,而且带来填埋场周围空气的污染;更重要的是,填埋过程中会产生大量的垃圾渗滤液需要处理[3]。而垃圾渗滤液具有很高的氨氮浓度,特别是老龄垃圾渗滤液具有高氨氮和低碳氮比的特征,采用传统的脱氮方法往往面临着能耗高和碳源不足等问题。如何经济有效地处理垃圾渗滤液仍是一个挑战。
生物脱氮是处理垃圾渗滤液较为经济有效的方法。在传统的生物处理过程中,高浓度的氨氮通过完全硝化与反硝化反应去除。然而,由于氨氮浓度很高且碳氮比低,导致硝化过程中需要大量曝气,能耗较高;同时,反硝化过程中需要投加大量的碳源,使得脱氮成本较高[4]。短程硝化反硝化通过将硝化过程控制在亚硝化阶段,直接利用亚硝态氮进行反硝化,缩短了反应过程,能够加快反应速度,可节约20%左右的曝气量和40%左右的有机碳源[5-11]。在高氨氮废水处理中,常利用游离氨(FA)对氨氧化菌(AOB)和亚硝酸盐氧化菌(NOB)的抑制差异性实现短程硝化[12]。然而,老龄垃圾渗滤液的碱度较高,水质偏碱性,进入生物脱氮系统后容易造成高浓度的FA,而FA具有较强的生物毒性,其对AOB和NOB均具有明显的抑制作用,从而导致系统亚硝化速率低。为此,在生物脱氮系统中合理降低FA浓度至关重要,而分点进水方式提供了一种比较可行的策略。采用分点进水方式合理分配进入系统各部分的基质浓度,可以在降低营养负荷和提高污泥浓度的同时降低FA浓度,以减少FA对AOB的抑制,提高亚硝化微生物的比生长优势[13],增强系统亚硝化性能。
基于此,本研究采用分点进水短程硝化反硝化工艺对老龄垃圾渗滤液进行了处理,通过合理分配营养负荷,以改变系统中FA浓度和微生物生长环境,并提高AOB种群生长优势,提高生物亚硝化性能以及脱氮效率,以期为实现垃圾渗滤液的高效低耗处理提供技术参考。
全文HTML
-
实验用水为老龄垃圾渗滤液,取自无锡市某垃圾渗滤液处理中心调节池,具体水质指标如表1所示。实验水质波动较大,偏碱性,具有高浓度的氨氮和总氮,C/N约为1.5。
-
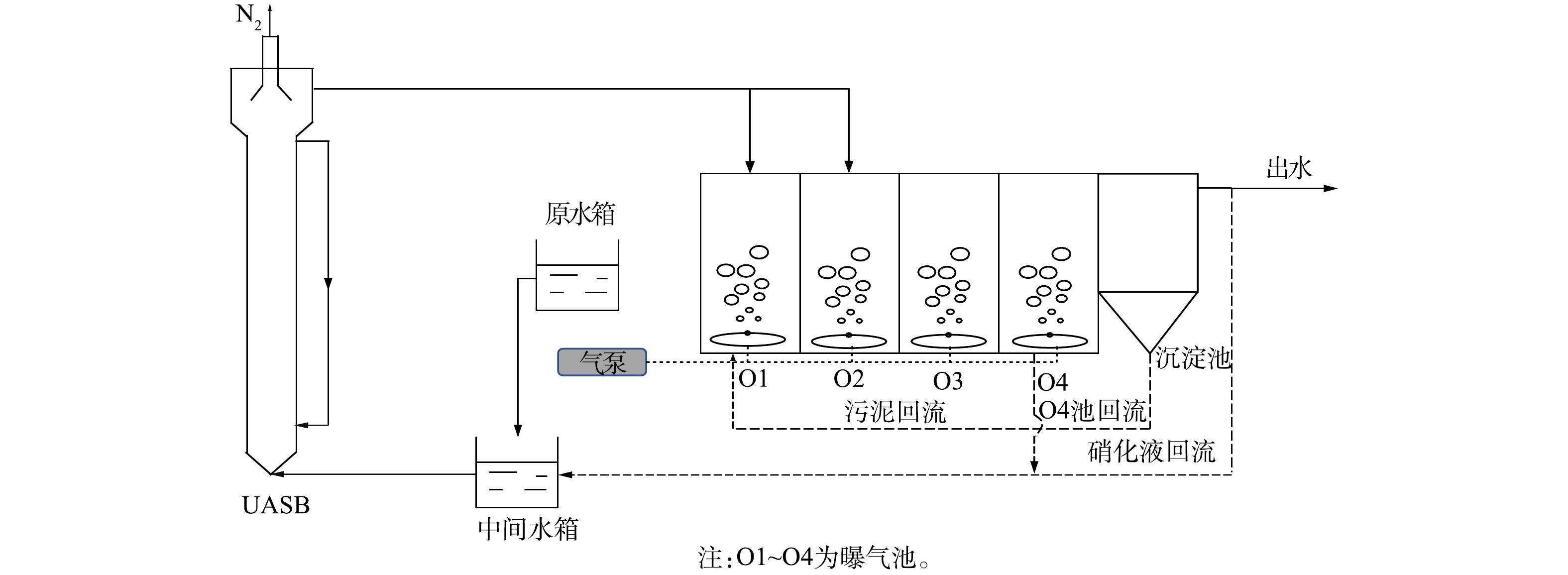
实验工艺过程如图1所示。核心实验装置主要包括UASB和曝气池2个部分,均由有机玻璃制成,总有效容积为50 L。UASB反应器高为2 m,内径为0.1 m,有效容积为10 L;曝气池长为0.6 m,高为0.5 m,宽为0.2 m,有效容积40 L,内装有生物填料以便挂膜。好氧区采用曝气泵曝气,进水、硝化液及污泥回流均采用蠕动泵进行精确控制。实验中反应器温度控制在(30±1) ℃。反应器用黑色材料覆盖,以防止光线穿透。
本研究工艺采用UASB作为前置反硝化单元,垃圾渗滤液原水首先与回流硝化液混合进入到脱氮反应器UASB中,利用原水中的有机物作为反硝化的电子供体,去除硝化液中部分亚硝态氮和硝态氮,同时为后续硝化反应提供碱度[14]。随后反硝化单元出水通过单点或分点进水的方式进入到好氧系统中进行生物硝化反应。最后通过硝化液的回流与微生物反硝化实现脱氮。
-
接种污泥取自无锡市某垃圾渗滤液处理中心的好氧池。其中曝气池(O1~O4)中接种污泥浓度为3 g·L−1(以MLSS计),UASB中接种污泥浓度为10 g·L−1(以MLSS计)。实验过程分为2个阶段。在阶段1(1~45 d),通过逐步提高进水量的方式快速启动短程硝化反硝化系统,在实验期间控制DO浓度低于1.0 mg·L−1,污泥回流比为100%,硝化液和O4池回流比各为100%。一直提升负荷至反应器崩溃,其后恢复系统至稳定运行(系统崩溃后恢复至稳定状态(46~54 d)这阶段数据未展示)。在阶段2,UASB出水按流量比1∶1分点进入硝化系统O1和O2池中,探究分点进水对于系统脱氮性能的影响。实验共设置5种工况:工况Ⅰ取未分点进水条件下系统运行时(40~45 d)的结果作为参照,此时HRT为5.5 d;工况Ⅱ为未分点进水运行(55~64 d),控制HRT为6.25 d;工况Ⅲ为分点进水运行(65~74 d),控制HRT为5.5 d;工况Ⅳ为分点进水运行(75~84 d),并提高系统负荷,控制HRT为5.0 d;工况Ⅴ为分点进水运行(85~99 d),HRT为5.0 d,按C/N=4在水中添加碳源。
-
进出水水样经 0.45 μm滤膜过滤后,滤液采用国标方法测试各项指标[15]。
NH+4 -N 采用纳氏试剂分光光度法测定,硝态氮(NO−3 -N)采用紫外分光光度法测定,亚硝态氮(NO−2 -N)采用N-(1-萘基)-乙二胺分光光度法测定,TN采用过硫酸钾氧化-紫外分光光度法测定;污泥浓度(MLSS)按重量法测定;DO采用便携式溶氧仪测定。亚硝态累积率(NAR)、游离氨(FA)、氨氧化速率(AOR)、亚硝态氮氧化速率(NOR)、总氮去除率(NRE)、反应器总氮去除负荷(NRR)、总氮容积负荷(NLR)的计算[16-18]如式(1)~(7)所示。式中:ηNAR为亚硝态氮积累率;
CNO−2-N 为反应器中亚硝态氮的质量浓度,mg·L−1;CNO−3-N 为反应器中硝态氮的质量浓度,mg·L−1。式中: CFA为游离氨的质量浓度,mg·L−1;T为该系统的温度,℃;
CNH+4-N 为反应器中氨氮的质量浓度,mg·L−1。式中: VAOR为氨氧化速率,mg·(L·h)−1;VNOR为亚硝态氮氧化速率,mg·(L·h)−1;
CNH+4-Ninf 和CNH+4-Neff 分别为进水和出水氨氮质量浓度,mg·L−1;CNO−3-Ninf 和CNO−3-Neff 分别为进水和出水硝态氮质量浓度,mg·L−1;t为水力停留时间,d。式中:ηNRE为总氮去除率;NNRR为总氮去除负荷,kg·(m3·d)−1;NNLR为总氮容积负荷,kg·(m3·d)−1;
CTNinf 和CTNeff 分别为进水和出水总氮质量浓度,mg·L−1;Vinf和V分别为进水体积和反应器容积,L,t为水力停留时间,d。 -
分别在运行第1、40 (分点进水运行前)和84天(分点进水运行后)取悬浮污泥样品进行分析。首先,采用试剂盒(PowerSoil DNA isolation kit,Mobio,USA)对样品微生物DNA进行提取,然后采用细菌的通用引物(338F:5′-ACTCCTACGGGAGGCAGCAG-3′和806R:5′-GGACTACHVGGGTWTCTAAT-3)对V3-V4可变区域进行PCR扩增,扩增片段长度约为200 bp;最后利用QuantiFluor™-ST(Promega,USA)进行检测定量,用分析软件QIIME1.8.0对微生物序列进行处理。
1.1. 垃圾渗滤液水质
1.2. 实验装置与工艺流程
1.3. 实验方法
1.4. 测定项目与方法












1.5. 微生物种群结构变化分析
-
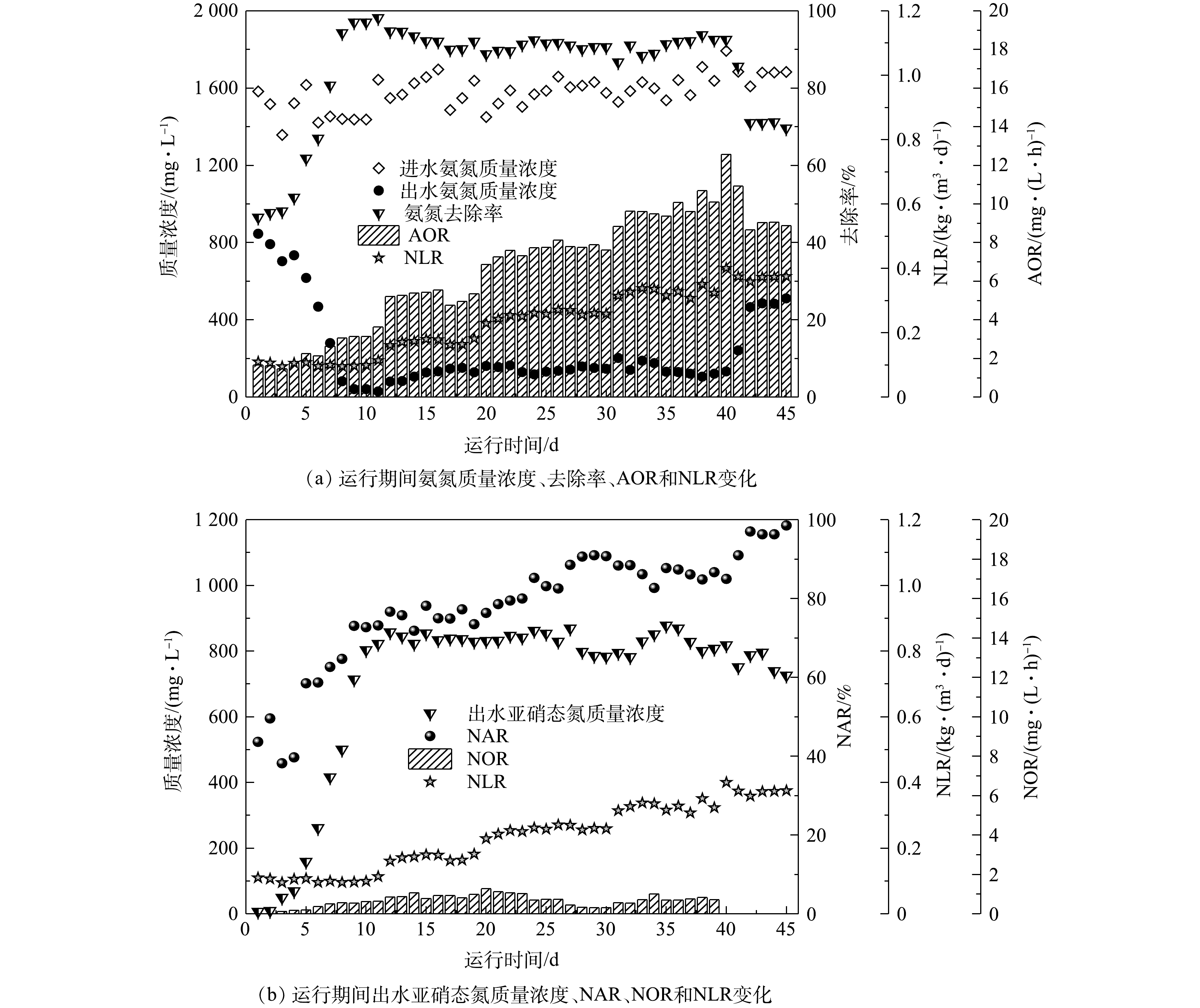
1)短程硝化反硝化效果。在生物脱氮系统启动过程中,随时间变化,垃圾渗滤液中氨氮的去除效果如图2(a)所示。在1~4 d内,反应器对氨氮去除效果不高,去除率低于60%,平均氨氧化速率(AOR)仅为1.63 mg·(L·h)−1,出水中氨氮浓度高于700 mg·L−1。由于反应系统接种污泥为全程硝化污泥,对低DO浓度环境敏感,部分菌体自溶,菌体活性变差[19]。然而,经过一段时间的适应,氨氮去除率迅速上升。到第12天时,氨氮去除率上升到94.78%,AOR上升为5.21 mg·(L·h)−1。进一步提高总氮容积负荷(NLR为0.23 kg·(m3·d)−1)后,出水中氨氮浓度始终保持较低水平,微生物活性逐步上升。到第40天时,氨氮去除率达到92.57%,AOR从一开始的1.63 mg·(L·h)−1增加到12.57 mg·(L·h)−1。然而,当NLR提升为0.38 kg·(m3·d)−1时,系统处理氨氮能力迅速下降,出水中氨氮浓度迅速上升,氨氮去除率降至69.63%。
脱氮系统内的亚硝态氮变化如图2(b)所示。由于接种污泥为全程硝化污泥,在开始的前几天,系统内氨氮大部分被转化为硝态氮,出水中亚硝态氮含量较低(<100 mg·L−1)。当系统运行一段时间后,由于溶解氧的限制,NOB活性被限制,亚硝态氮氧化速率(NOR)始终维持在较低水平(0.78 mg·(L·h)−1),而AOB具有较高活性。因此,系统内氨氮主要被转化成亚硝态氮,水中亚硝态氮含量迅速升高,系统NAR提高。到第40天时,NAR达到约84.97%,出水中亚硝态氮浓度达到818.66 mg·L−1。因此,通过控制DO浓度小于1.0 mg·L−1成功启动了短程硝化反硝化系统。
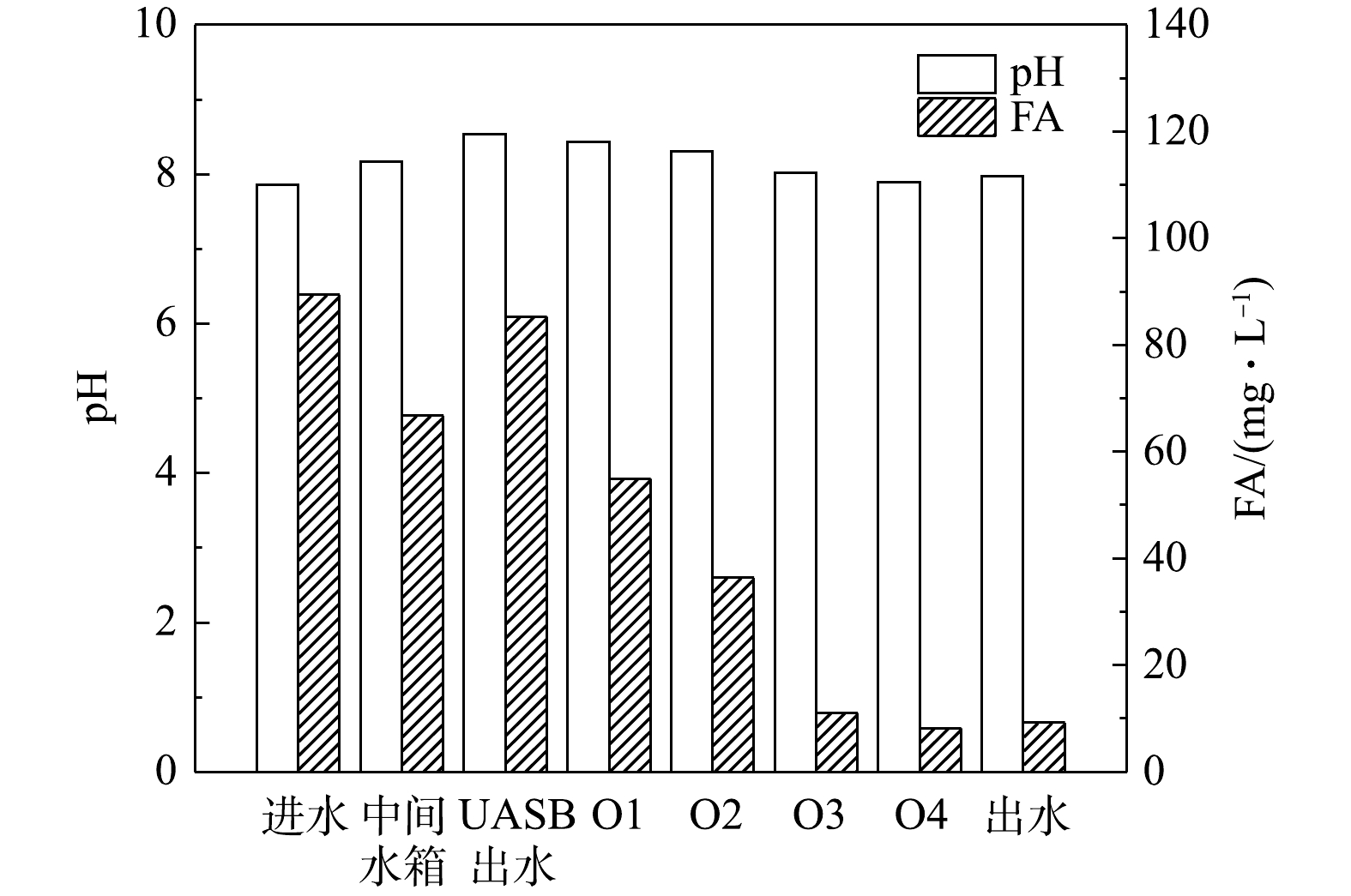
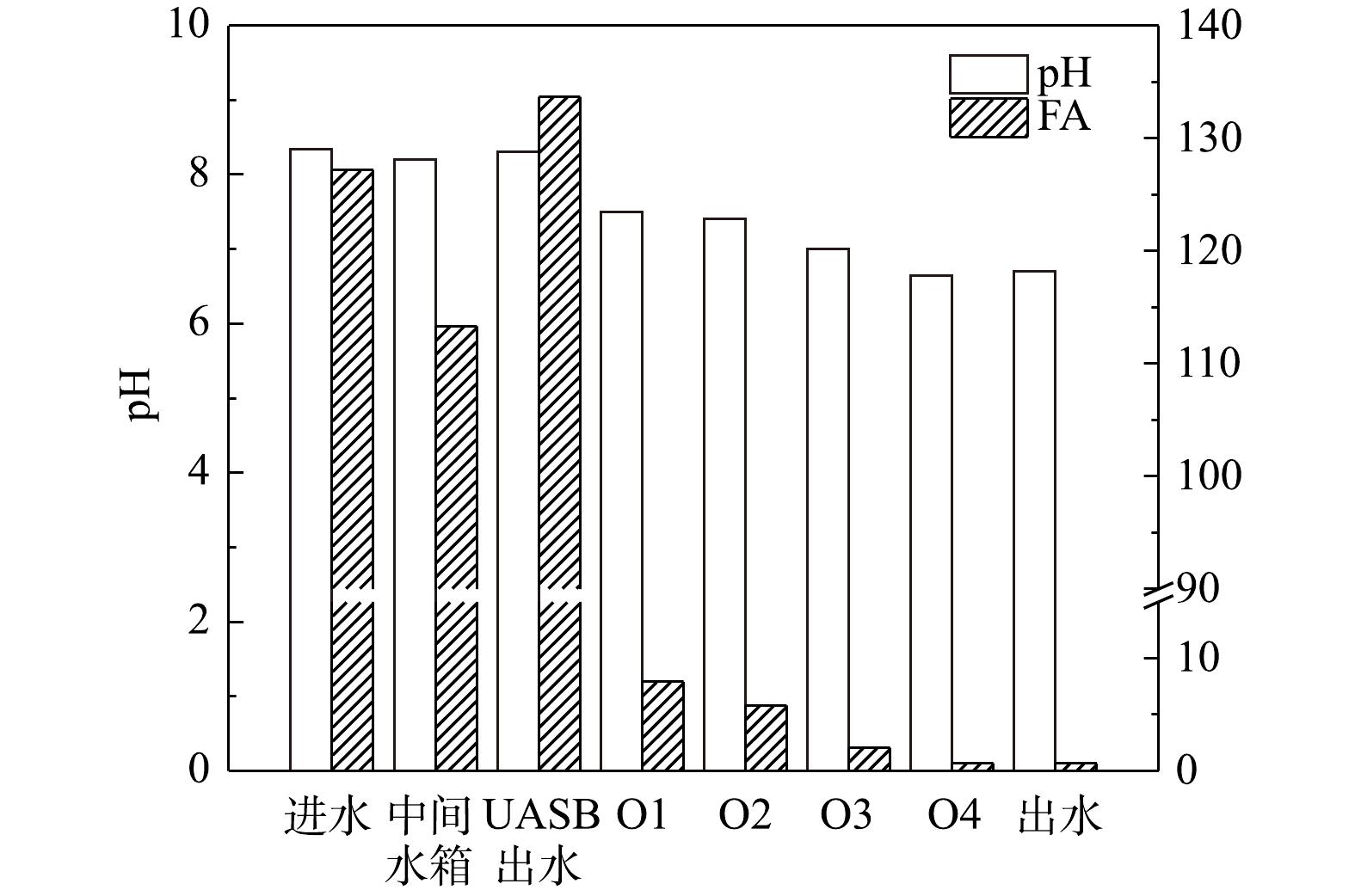
2)FA和pH的变化分析。短程硝化反硝化脱氮系统内各取样点的FA和pH变化如图3所示。原水与回流硝化液混合后进入UASB中进行反硝化反应,并提高后续硝化反应的碱度。在硝化段即好氧段,氨氮浓度不断下降,碱度不断被消耗,pH不断下降,pH由O1的8.43降至O4的7.89,然后出水中pH略有上升,这可能是由于在沉淀池中发生了反硝化反应[20]。有研究[16]表明,FA是维持垃圾渗滤液短程硝化的重要因素之一。WEI等[12]的研究表明,当FA浓度在0.1~1.0 mg·L−1时,即可以抑制NOB的活性;与NOB相比,AOB承受FA的能力更强,当FA超过10 mg·L−1时会对AOB产生抑制。因此,可以通过FA对AOB和NOB的选择性抑制实现短程硝化。由图3中可以看出,虽然随着微生物反应的进行好氧池中FA浓度逐渐降低,但在好氧阶段FA浓度最低也有8.13 mg·L−1 (O4池),说明在硝化阶段系统内全程均存在FA对NOB的抑制,从而降低亚硝态氮的进一步转化,实现稳定的短程硝化效果。此外,在O1、O2池中,FA浓度大于36.32 mg·L−1,这不仅对NOB不利,而且远超过了AOB的耐受范围,即在抑制NOB的同时,也抑制了AOB的活性,从而不利于整体脱氮系统的亚硝化过程,限制了亚硝化效率的提升。
-
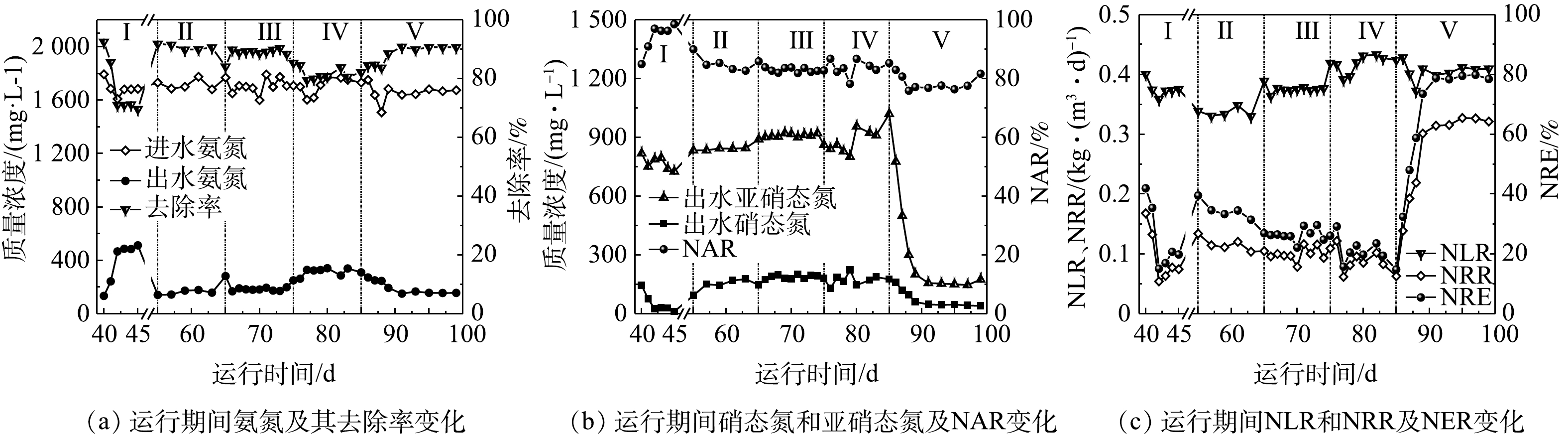
1)系统的稳定运行情况。为了在抑制NOB的同时尽量避免FA对AOB的抑制,提高亚硝化速率,缩短反应时间,提高脱氮效率,本研究探讨了分点进水对系统脱氮性能的影响。分点进水后脱氮系统的运行结果如图4所示。分点进水前,脱氮系统的HRT缩短到5.5 d时(工况Ⅰ),氨氮去除率快速下降,低于80%(图2(a))。为了保持较高的氨氮氧化效率,在采用分点进水方式前,将脱氮系统的HRT调整为6.25 d(工况Ⅱ),并实现系统稳定的运行,氨氮的去除率达到90%以上(图4(a))。以分点进水前HRT为5.5 d时(工况Ⅰ)脱氮系统的运行效果为对照,此时,氨氮去除率迅速下降到69.63%,总氮去除率(NRE)仅有18.06%(图4(c));分点进水后,HRT为5.5 d时(工况Ⅲ),氨氮去除率仍有89.86%,NRE为27.62%,比分点进水前提高了52.93%。可见,通过分点进水的方式提高了系统的脱氮效率。当进一步提升负荷,HRT缩短为5.0 d时(工况Ⅳ),出水中氨氮浓度上升,氨氮去除率开始下降,但氨氮去除率仍高于81.25%,出水中亚硝态氮含量先下降后上升,微生物经过几天适应后,亚硝态氮浓度开始上升,NAR高于80%。由图4(c)也可以看出,分点进水前,系统总氮容积负荷率(NLR)平均值为0.34 kg·(m3·d)−1,分点进水后在85~99 d(工况Ⅳ),系统平均NLR为0.42 kg·(m3·d)−1,相比而言提升了23.53%。AOR从分点进水前平均10.37 mg·(L·h)−1(55~64 d)提升到了11.98 mg·(L·h)−1 (75~84 d),这意味着将相同量的氨氮亚硝化所需要的时间缩短了15.52%。在工况Ⅴ(85~99 d),按C/N=4额外补充碳源后,出水中氨氮浓度从第86天的271.32 mg·L−1下降到了155.25 mg·L−1,氨氮去除率上升到90.84%,这主要是由于补充了碳源,反硝化过程能够快速进行,系统中亚硝态氮浓度快速降低(图4(b)),从而降低产物抑制作用,促进氨氮氧化过程顺利进行。添加碳源后,系统总氮去除率NRE迅速上升,最终达到80%左右。
2)系统内FA和pH的变化分析。为了探究分点进水后反应系统负荷提高并能实现稳定的短程硝化的原因,在反应器65~74 d(工况Ⅲ)的运行过程中,对系统内FA和pH的变化进行沿程分析,结果如图5所示。垃圾渗滤液进水在经过反硝化系统后,由于微生物的反硝化作用使得污水pH升高,FA浓度上升到133.70 mg·L−1,这严重超出了AOB对FA的耐受范围。通过分点进水的方式将反硝化系统出水分别进入到O1和O2池后,在稀释作用下FA的浓度分别降低到7.91 mg·L−1和5.81 mg·L−1。CHUNG等[21]的研究表明,当FA浓度为5~10 mg·L−1时,能有效的抑制NOB的活性,而不对AOB产生抑制作用。比较分点进水前后脱氮系统中FA浓度的变化可以发现,未分点进水前,O1和O2池中的FA浓度超过36.32 mg·L−1 (图3),在抑制NOB的同时也严重抑制了AOB活性。而通过分点进水的方式降低了O1、O2池内的基质浓度,使得FA浓度维持在抑制NOB活性的同时,却对AOB的活性不产生抑制,从而有利于提高AOB的比生长优势,提高系统的亚硝化反应速率。
系统内FA浓度主要受氨氮浓度、pH和温度的影响[16],在本研究中温度保持不变,氨氮进行硝化反应浓度会降低,且硝化反应会消耗碱度降低pH。进一步分析不同进水方式对系统FA的影响发现,分点进水方式降低FA主要通过以下2方面。一方面,进水被稀释,氨氮和碱度(pH)降低,从而导致FA降低:在单点进水时,UASB中的出水全部进入到O1池中,造成较高的氨氮浓度和较高pH,这使得FA浓度较高;而在分点进水时,UASB出水通过分点方式进入硝化系统使得进入O1池中污水的量只有50%,从而O1池的氨氮浓度和碱度(pH)比单点进水模式要低,降低了FA浓度。另一方面,在分点进水模式下,首先,脱氮系统中形成污泥浓度梯度分布,前端污泥浓度高,硝化速度快,抗冲击能力强;而且整个系统的微生物平均浓度得到提高[22],硝化性能提升,氨氮浓度降低,从而FA浓度降低;其次,由于UASB出水中还含有大量有机物[23],进入好氧池后有利于异养菌的繁殖,进而与自养型硝化菌竞争氧气;而分点进水下营养物质被稀释,限制了异养菌的大量繁殖竞争氧气,从而有利于提高硝化菌的种群优势,提高硝化效果[24]。因此,采取分点进水方式,可以使FA保持在适宜范围内,利于AOB生长,进而提高系统亚硝化速率。
-
1)物种多样性和种群结构的分析。对接种污泥和驯化后的污泥微生物DNA进行提取和高通量测序,得到优化序列66 268和71 679,聚类相似度97%。共识别出3 148个和3 673个OTUs(operational taxonomic units),如表2所示。2个样本微生物群落多样性的覆盖度均在99%以上。
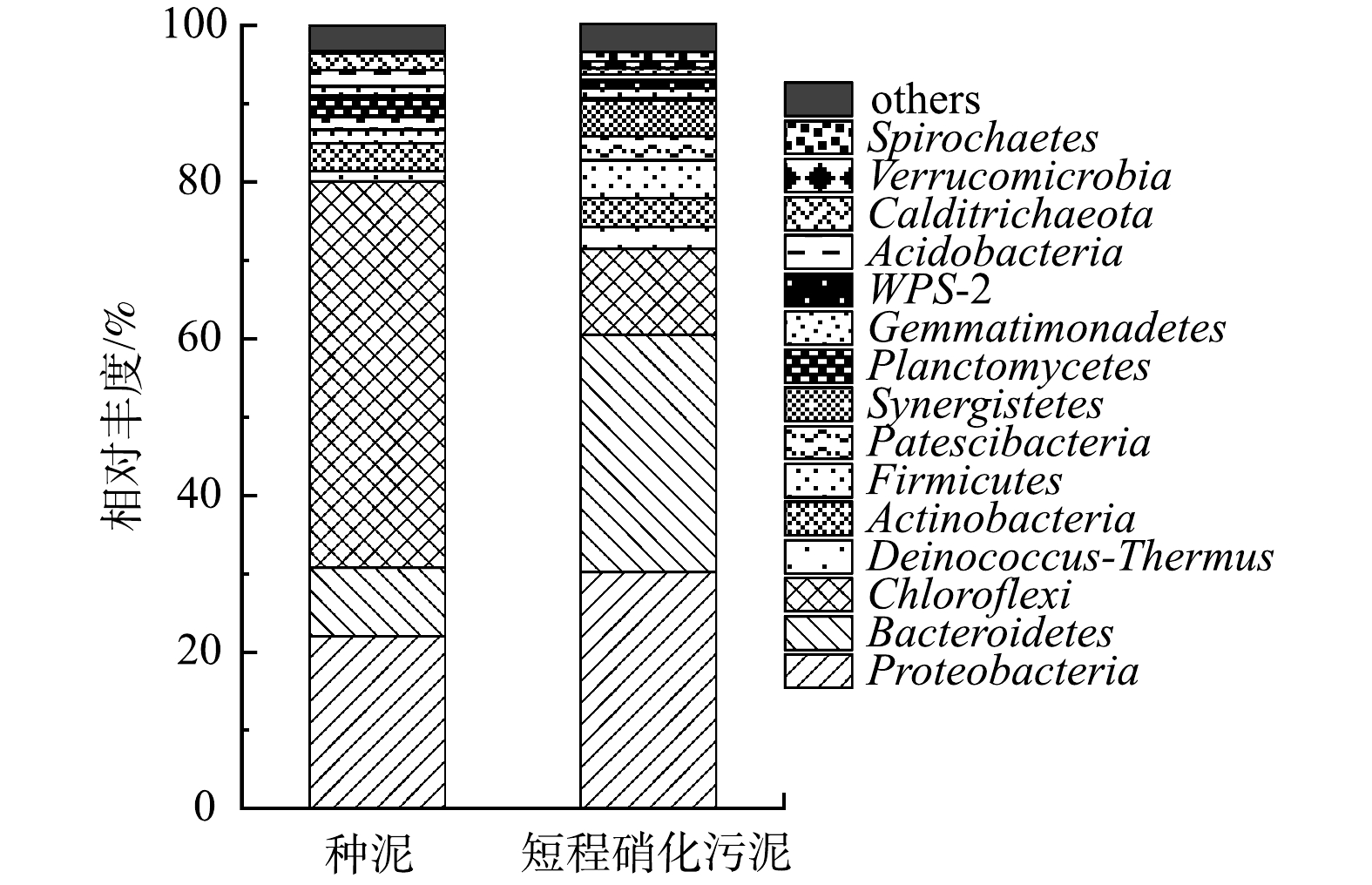
如表2所示,驯化后污泥的Shannon多样性指数(5.45)、基于丰度的覆盖度估计量(ACE)(1 468.29)和Chao丰富度估计量(1 468.73)均高于种泥 (4.76、1 215.82和1 215.80)。驯化后的污泥和种泥Simpson指数分别为0.01和0.03。利用Shannon指数和Simpson指数可以分别表示微生物群落的丰富度和多样性,Shannon、ACE、Chao指数越高,Simpson越低,说明微生物种群丰富度和多样性越高[22]。上述结果表明,经过一段时间培养驯化后,微生物适应了进水水质,微生物多样性提高。有研究[25]表明,微生物多样性越高,反应器的稳定性越好。
如图6所示,2个污泥样本中Proteobacteria、Bacteroidetes、Chloroflexi为优势菌门,占比超过65%。Proteobacteria包含了多种代谢类型的菌种,包括AOB、NOB和反硝化细菌等[26]。国内外的一些研究[27-28]表明,大多数在生物脱氮过程中起重要作用的微生物均属于Proteobacteria菌门。驯化后污泥中的Proteobacteria微生物相对丰度增大,也是系统脱氮能力增强的主要原因之一。Bacteroidetes菌门为化能有机营养菌,可以降解多种复杂有机物,代谢碳水化合物,主要进行有机物的降解,广泛存在于活性污泥系统中[29]。驯化后Bacteroidetes菌相对丰度升高,这也与韩亚琳等[30]的研究结果相似。
2)分点后系统内脱氮功能菌的变化。为了评估分点进水对脱氮系统内微生物种群的影响,对系统内O1、O2、O3、O4池内微生物群落的属级别上的变化进行了分析,结果如图7所示。在所有样品中都检测到了Saprospiraceae和Thauera,这些菌种与有机物的降解有关[31]。Saprospiraceae可以降解多糖和蛋白质,然后利用水解产物作为能量和碳源来进行生长[32]。Nitrosomonas属于AOB菌属,与氨氮的氧化有关。分点进水前O1、O2、O3、O4中Nitrosomonas相对丰度分别为2.02%、1.05%、1.75%、0.86%,分点进水模式下运行后细菌营养负荷被降低,Nitrosomonas的相对丰度增加到2.27%、1.77%、2.75%、1.37%。相较于分点进水前,4个好氧区内Nitrosomonas的相对丰度分别提高了12.37%、68.57%、57.14%、59.30%。以上结果表明,分点进水后由于FA和基质浓度均有所降低,AOB菌属受到的抑制减弱,微生物生长优势提升,从而表现出亚硝化速率提高,系统脱氮能力增强。
2.1. 短程硝化反硝化系统的启动与稳定运行
2.2. 分点进水下脱氮系统的运行和分析
2.3. 微生物种群分析
-
1)控制DO浓度小于1.0 mg·L−1,可成功实现短程硝化反硝化系统的快速启动,亚硝态氮累积率(NAR)能达到84.97%;NOB在低氧环境下活性受到抑制,NOR仅有0.78 mg·(L·h)−1;同时,由于O1、O2池中FA浓度较高,在抑制NOB的同时也抑制了AOB活性,从而降低了系统亚硝化速率。
2)分点进水运行有效降低了O1、O2中的FA浓度,可使其对NOB产生抑制的同时不抑制AOB的活性,提高了亚硝化速率,系统总氮容积负荷相比未分点进水时提升了23.53%。
3)分点进水运行后,O1、O2、O3、O4中AOB相对丰度均有提高,进一步表明分点进水模式下运行可以降低FA对AOB的抑制,提高了AOB的生长优势和亚硝化速率,实现高效短程硝化效果;同时,亚硝化过程形成的高浓度亚硝态氮经外加碳源的反硝化过程后浓度快速降低,总氮去除率达到80%左右。
