


 DownLoad:
DownLoad:


 百度学术
百度学术


-
降雨径流是城市面源污染物空间迁移的主要载体。在降雨过程中,雨水及其形成的地表径流冲刷地面污染物,通过排水沟渠或分流制排水系统直接进入江河湖泊中,对受纳水体的水质保障带来较大压力。为保护水资源、改善水生态、优化水环境、确保水安全,海绵城市建设理念应运而生。为海绵城市主流措施之一的生物滞留系统凭借其应用灵活、径流调控成效突出的优势得到了广泛应用。本课题组前期的研究结果[1]表明,回填填料、种植植物对生物滞留系统的氮磷去除效果具有显著影响,海绵铁的添加有助于提升硝态氮的去除效果。生物滞留系统的填料组成和填料层的深度也会影响污染物的去除效率[2]。同时,植物的栽种可以提升系统对有机物和营养性污染物的去除效率[3]。
为进一步探明填料和植物对生物滞留系统滞净效能的强化情况,国内外学者开展了一系列相关研究。高晓丽等[4]、王书敏等[5]对生物滞留设施填料研究进展和生物滞留介质土理化性质进行了综述分析,介绍了国内外相关研究中填料的组成及配比、去污效果、渗透性能、填料的改良及填料深度,分析了介质土的粒径时空分布特点,可为生物滞留系统填料的筛选提供依据。仇付国等[2, 6-7]尝试用铝污泥和沸石对传统基质填料进行改良以提高系统对氨氮和磷的吸附效果,并在系统底部设置或增加淹没区高度,创造缺氧环境以提高系统对硝态氮的去除效果。为研究季节及植物对生物滞留系统径流污染物去除的影响,HERMAWAN等[8]研究了热带条件下生物滞留系统的季节特性,评估了其性能并筛选出适宜条件生长的植物。余雪花等[3]和陈韬等[9]对生物滞留系统中能有效去除多种污染物的最佳植物进行了研究,比较了雨水中各类污染物在不同种植植物下的去除效果,结果表明,植物对TP的去除影响不显著,对TN去除率良好,对硝态氮去除有一定效果,但并不稳定。
尽管诸多业界学者已在生物滞留系统回填介质填料方面做了一定程度的研究,然而,生物滞留系统滞净效果影响因素众多,研究基础仍然薄弱,在区域环境背景下开展生物滞留系统滞净效能的研究仍需加强。鉴于此,本研究以重庆地区常用的海绵设施种植植物为供试对象,以改良回填介质为装填基质,在模拟径流条件下开展了场次降雨实验,旨在阐明径流污染物在改良填料生物滞留系统中不同季节的去除情况,获得改良填料系统污染物去除的季节分布特征,以期为生物滞留系统优化建设提供参考。
全文HTML
-
如图1所示,设计生物滞留是实验装置为长方体柱状,柱子高约1.2 m,底面长宽均为36 cm,前后两面从上到下设置5个出水阀门,左右两面设置4个出水阀门;综合设计生物滞留装置内填料由下往上依次为 21 cm海绵铁、7 cm高度的3~5 mm火山岩、9 cm高度的5~8 mm火山岩、5 cm高度的河砂和50 cm高度的沙土比7∶3的人工配土[1, 5];栽种植物为金叶女贞(Ligustrum × vicaryi Rehder)和麦冬(Ophiopogon japonicus (Linn. f.) Ker-Gawl.)。土壤取自重庆文理学院校内实验外场地旁,自然风干后,打磨成100目左右的细土。
-
模拟雨水径流实验工况如表1所示,参考王书敏等[10-11]山城雨水径流相关研究,计算出各污染物进水浓度。实验模拟雨水采用磷酸二氢钾、硫酸铵、硝酸钾等药剂配制。TN进水浓度为4.76~10.04 mg·L−1,硝态氮进水浓度为0.3~2.64 mg·L−1,氨氮进水浓度为1.98~3.96 mg·L−1,TP进水浓度为1.51~2.53 mg·L−1,磷酸盐进水浓度为1.33~2.88 mg·L−1,TOC进水浓度为7.9~21.68 mg·L−1。进水开始后约2 h,打开最下面的阀门,其连接的高度与出水高度(30 cm/40 cm/50 cm)一致的导管开始出水,而后不间断接水,取5~10 L水样后摇匀从中取500 mL左右,现场测pH、氧化还原电位等参数,冷藏备用。
-
水样使用0.45 μm的滤膜过滤处理,氨氮和硝态氮分别采用紫外分光光度计(岛津UV-2600, 日本)和全自动间断化学分析仪(CleverChem380, 德国)进行测定;水样中TN用紫外分光光度计检测;TOC测定采用TOC分析仪(Analytikjena Multi N/C 2100 s,德国);总磷用分光光度计测定;磷酸盐则添加抗坏血酸和钼酸铵后,用分光光度计测定。数据处理和绘图采用软件Excel 2016和Origin 2018。
1.1. 构建生物滞留系统
1.2. 进出水设计及取样
1.3. 分析方法
-
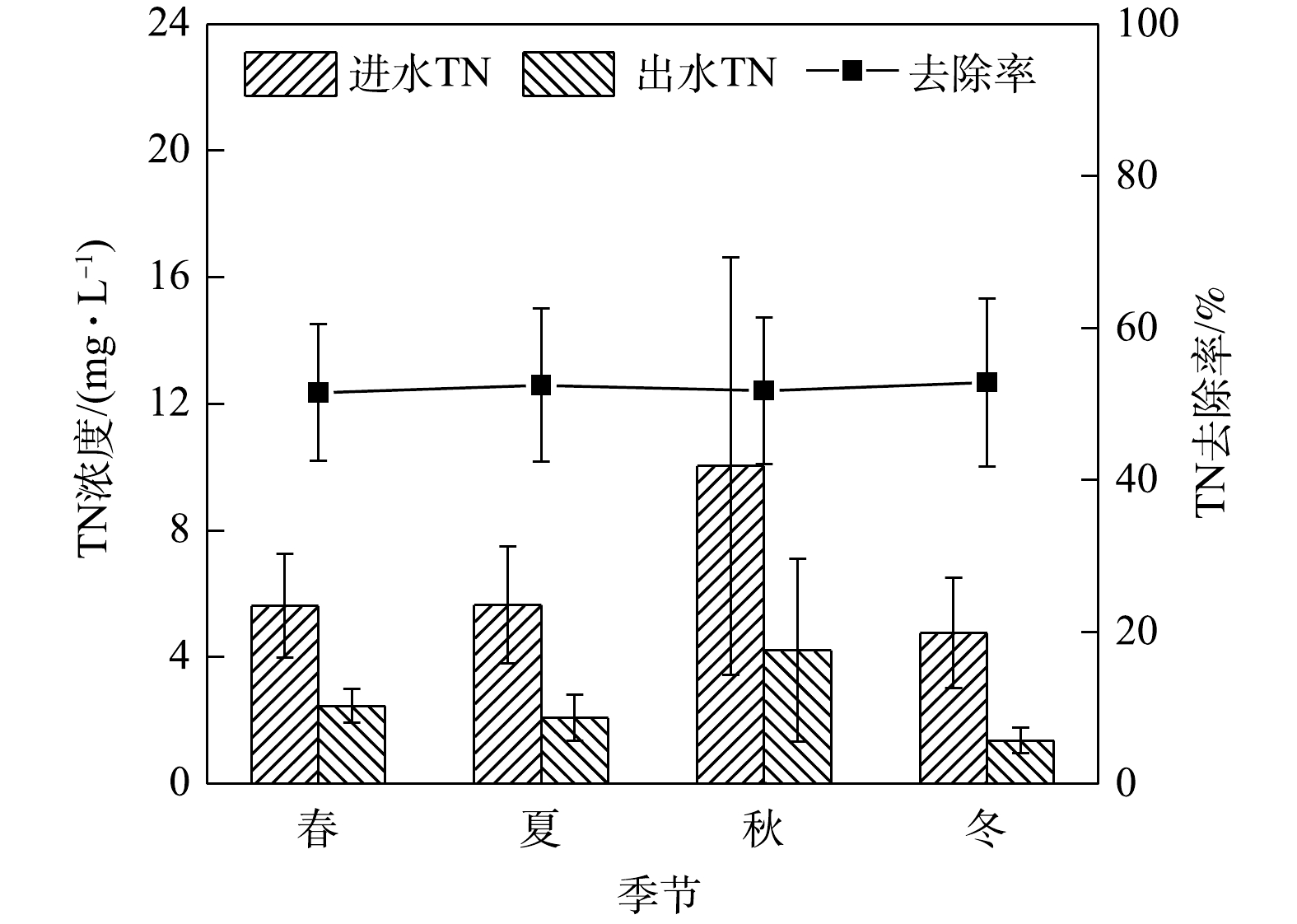
1)季节变化对TN去除效果的影响。由图2可知,在春夏秋冬四季,秋季径流的总氮平均进水浓度最高为10.04 mg·L−1,平均出水浓度为4.21 mg·L−1;冬季平均进水浓度为4.76 mg·L−1,出水浓度为1.37 mg·L−1,夏冬两季出水水质均满足地表水环境质量标准Ⅴ类限值,春秋两季不满足。生物滞留系统对总氮的去除率冬季表现最高,夏季,秋季与春季表现略差,整体在20%以内变化。
TN在冬季去除率较高的原因可能是因为春季开始种植的麦冬生长到冬季,根系较其他季节多。GOH等[12]发现,繁盛的根系在吸收TN、保持介质的导水性以及减少系统径流方面发挥着重要作用。生物滞留系统对氮污染物的去除机理主要有吸附、过滤、硝化反硝化、微生物同化和植物吸收等作用。TN去除率高则说明系统的反硝化能力较强。有研究[1]表明,海绵铁对TN的去除有良好效果。海绵铁促进硝化作用的机理主要是:硝化细菌的细胞膜具有复杂的内褶结构,铁离子通过加大对其的渗透性进而加快硝化的进行;同时由于海绵铁对
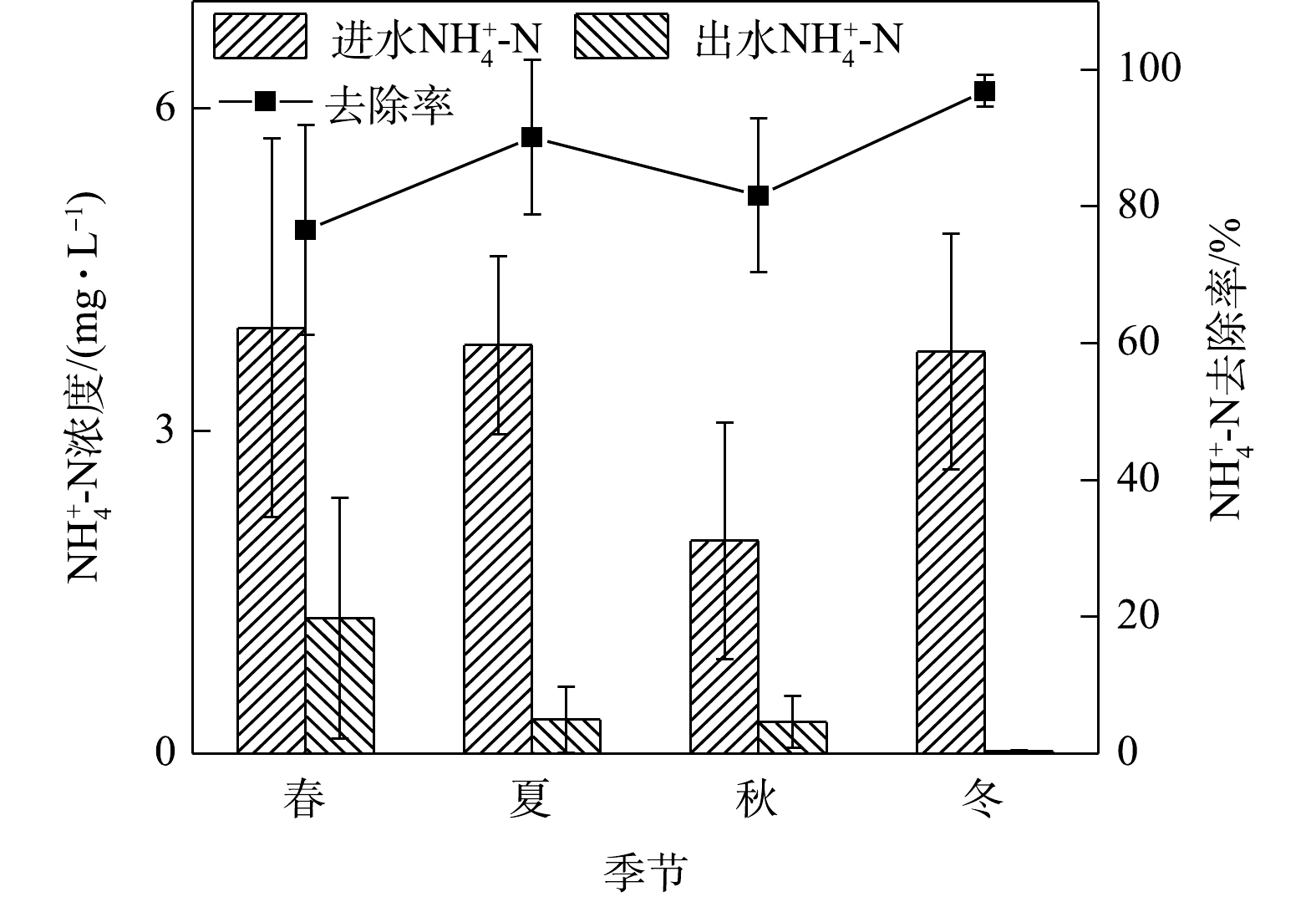
NO−3 -N和有机氮有一定的还原作用,从而使有机氮和NO−3 -N转化为NH+4 -N。而植物对氮素的去除效果受植物种类、干湿条件、气候变化、植物与根际微生物协同作用等因素的综合影响,因此,选择合适的植物,对生物滞留系统的设计具有重要的实际意义[13]。植被的存在可以极大地提高TN去除率或其吸收利用率[2, 13-15]。本研究的实验系统填料中有海绵铁,种植植物为适应性强、根系较长且多的金叶女贞和麦冬。从整体来看,系统全年TN去除率平均值为60.58%,冬夏季去除率较春秋两季偏高。这可能是因为在干燥条件下,对于根长的物种,非饱和(自由排水)设计的本地植物的脱氮性能较高,且存在饱和带的情况下,植物生长速度慢、叶面积大,与出水氮浓度较低显著相关[16]。2)季节变化对氨氮去除效果的影响。如图3所示,在春夏秋冬四季,出水氨氮平均浓度在冬季最低,春季最高。春季平均进水浓度为6.08 mg·L−1,出水浓度平均为1.26 mg·L−1,冬季平均进水浓度为3.74 mg·L−1,平均出水浓度为0.02 mg·L−1,春夏秋冬平均出水浓度均满足地表水环境质量标准Ⅳ类限值。在实验系统中,氨氮最高去除率在冬季,为96.88%,最低在春季,为69.34%。
硝化作用是指氨氮在有氧条件下,经亚硝酸细菌和硝酸细菌的作用转化为硝酸的过程。植物通过根系储氧,在根毛周围形成好氧区,离根毛较远的区域则呈现缺氧状态,更远的区域则是完全厌氧,进而形成内部好氧、缺氧、厌氧区的多串联单元,使硝化反硝化作用同时发生[17]。城市径流中的有机氮可经过吸附、沉淀等物理作用被介质截留,在微生物作用下转化为
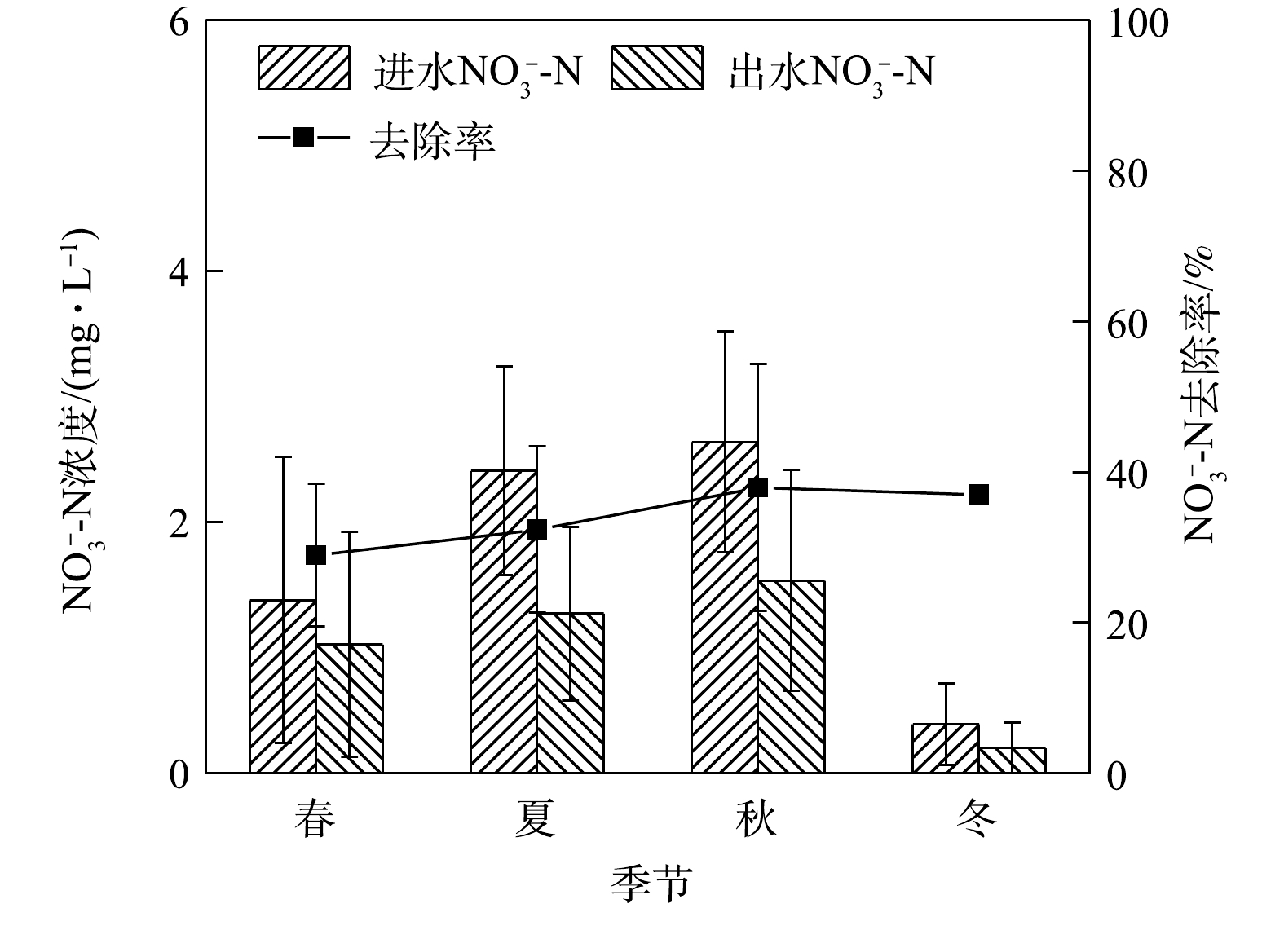
NH+4 -N和NO−3 -N等形态的无机氮。NH+4 -N可被带负电荷的介质颗粒吸附而去除,也可经硝化细菌在好氧条件下氧化成NO−3 -N,进而先被植物吸收进行生物同化,剩余部分则在缺氧条件下经反硝化作用还原成气态氮并排出系统[18]。在这个过程中,NO−3 -N前期去除的主要途径是生物同化,此时反硝化作用对氮污染物去除的贡献率不到10%[19]。本研究中,系统对氨氮的去除率比较理想,这可能是因为填料层中微生物的硝化作用以及种植植物对氨氮的去除效果优于硝态氮所致[13]。氨氮在冬季去除率较高,这与王超等[20]研究的结果“夏季氨氮去除率最高”有所不同。其原因可能是,本实验模拟的山城雨水径流,与文献[20]报道的河水原水相比,进水水质和条件不同。3)季节变化对硝态氮去除效果的影响。硝态氮的去除是生物滞留系统的一大技术难点。其原因是系统中硝态氮的去除不稳定或者说去除效能过低。如图4所示,硝态氮在秋季平均进水浓度最高为2.64 mg·L−1,平均出水浓度为1.54 mg·L−1,不满足地表水环境质量标准限值。本研究系统对硝态氮的去除率较低,在春季去除率最低为28.98%,夏、秋、冬三季则在5%左右浮动,且相对稳定,全年整体去除效能偏低。
有研究[21-22]表明,硝态氮在春季的去除与植物无关,而可能是由微生物群落驱动的,这是因为土壤微生物量C、N、P在春季和冬季含量较高。在实验系统中,硝态氮在冬季出水浓度最低,这可能是因为植物在种植几个月后达到一定的生物量水平,而根质量增长与硝态氮去除率息息相关[23]。此外,增加饱和带深度可以显著提高
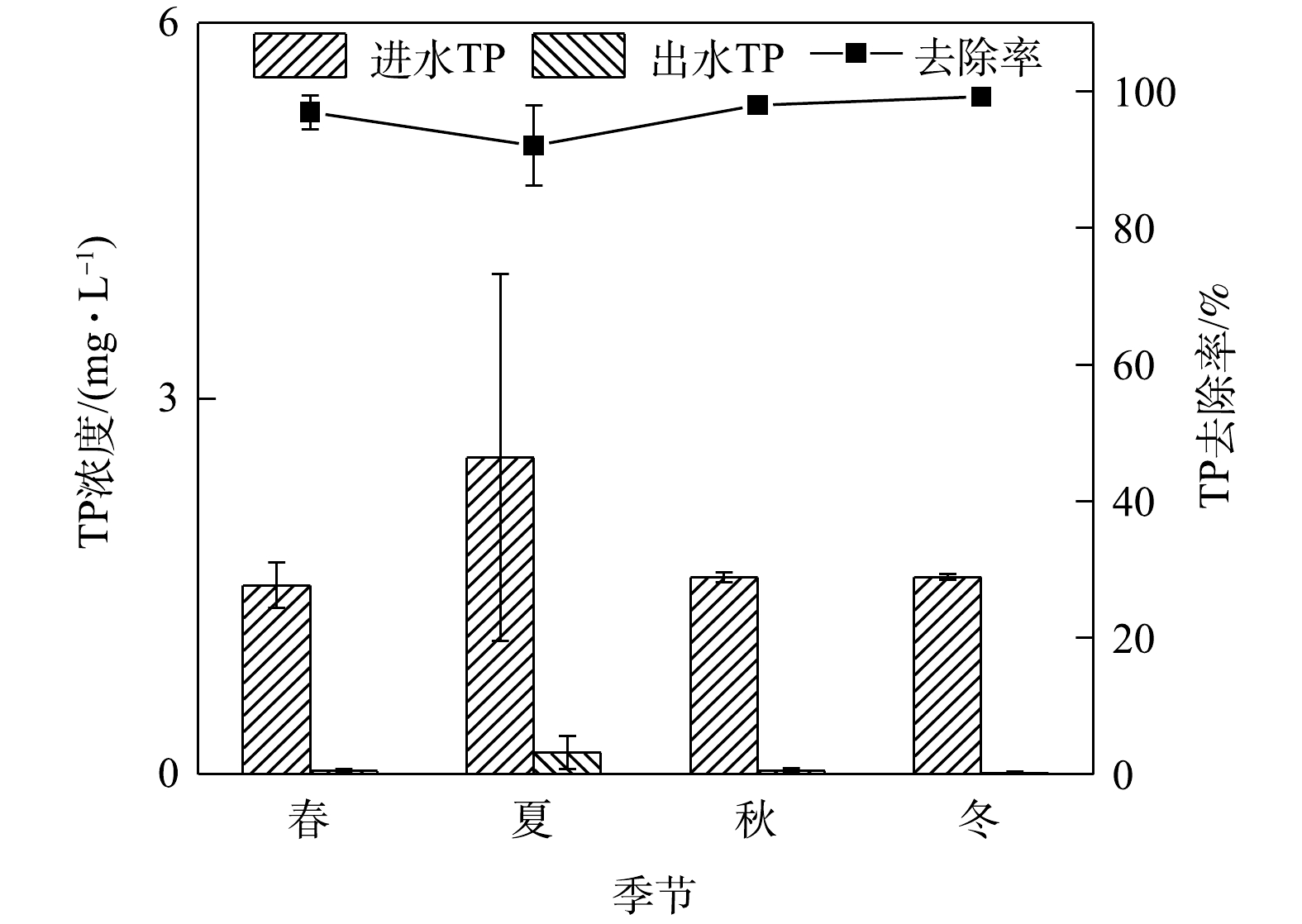
NO−3 的去除率,通过改造饱和区为反硝化过程创造厌氧条件,从而可以提高脱氮效果[24]。在饱和区深度由0 cm增加到60 cm的过程中,NO−3 的去除率显著提高,而NH+4 的去除率不受饱和区深度的影响[25-26]。本研究的实验系统参考其他研究[1, 27]的优化方案,为创造更好的反硝化环境而设置有不同深度淹没区(30、40、50 cm),且将海绵铁作为淹没区材料以加强硝态氮的去除。结果表明,氨氮去除效果良好,硝态氮去除效果略差。这说明实验系统淹没区的设置没有为反硝化菌创造良好的环境,可能是由于淹没区高度设置不够高所致。4)季节变化对TP去除效果的影响。如图5所示,在春夏秋冬四季,秋季最低平均进水浓度为1.57 mg.L−1,平均出水浓度为0.01 mg.L−1,春夏秋冬平均出水浓度均满足地表水环境质量标准Ⅲ类限值。实验系统对雨水径流中总磷的去除效率全年处于高水平状态,对TP去除率较为理想,全年最高去除率为99.24%。
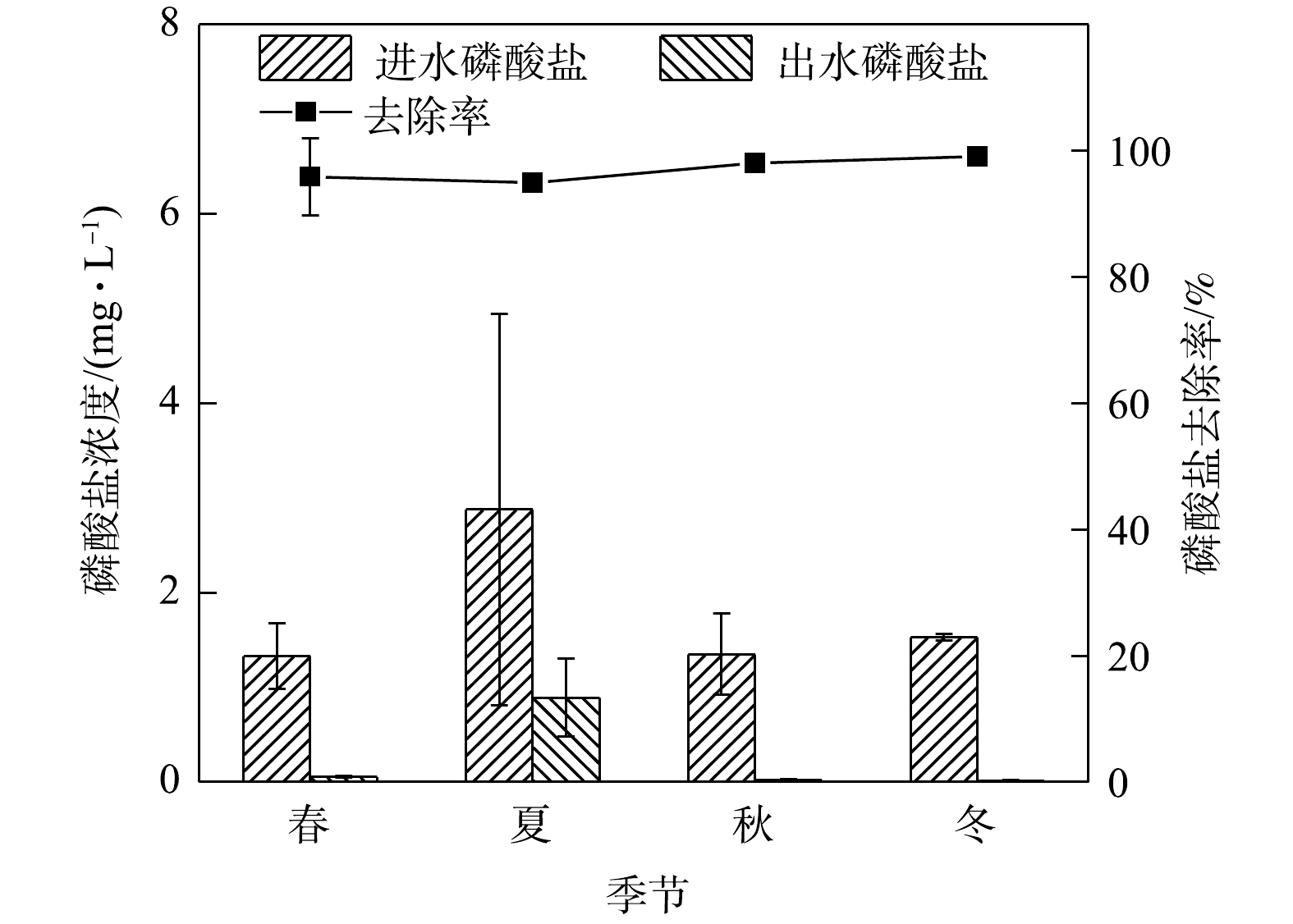
结果表明,TP的去除率与季节关系不大,这可能是因为TP的去除主要由填料层介质驱动的。AHMAD等[28]对土壤中磷的组分进行季节性分析发现,各磷组分的定位-季节互作效应不显著,但从夏季到冬季土壤中的TP含量会增加。有研究[3, 8, 23]表明,TP的去除与植物种类无关或者关联较小。TP在不同降雨期迁移到土壤中,然后在土壤中转化为不同的磷形态,植物在根系和微生物的作用下,不断地转化、吸收和利用磷,实现水-土-根系/生物系统中磷的动态平衡[29]。通常生物滞留系统对径流雨水中TP的去除率在80%以上,而改变填料类型、配比及添加改良剂则去除率增至90%以上[8, 29-32]。本研究的实验系统在淹没层设置海绵铁的情况下,还设有2种粒径火山岩,此设计填料层极好地提高了TP的去除率,这与匡颖等[33]、林修咏[1]的研究结果一致,且系统TP的去除率受季节影响较小。
5)季节变化对磷酸盐去除效果的影响。如图6所示,实验系统对磷酸盐的去除率极为理想。在春夏秋冬四季,夏季平均进水浓度最高为2.88 mg.L−1,相应平均出水浓度为0.89 mg.L−1,去除率高达96%。磷酸盐去除率同TP情况相似,全年去除率整体水平较高,各季度去除率变化差别不大。磷酸盐是生物滞留系统去除的主要营养元素之一。城市径流中的磷主要分布在溶解态(一般为有机磷和磷酸盐)和附属于颗粒物的磷之间[34]。溶解态磷中最主要的正磷酸盐通过填料的吸附和植物的吸收而去除[35]。实验系统选用的火山岩填料,其主要成份为硅、铝、锰、铁等几十种矿物质和微量元素,而Al和Fe与
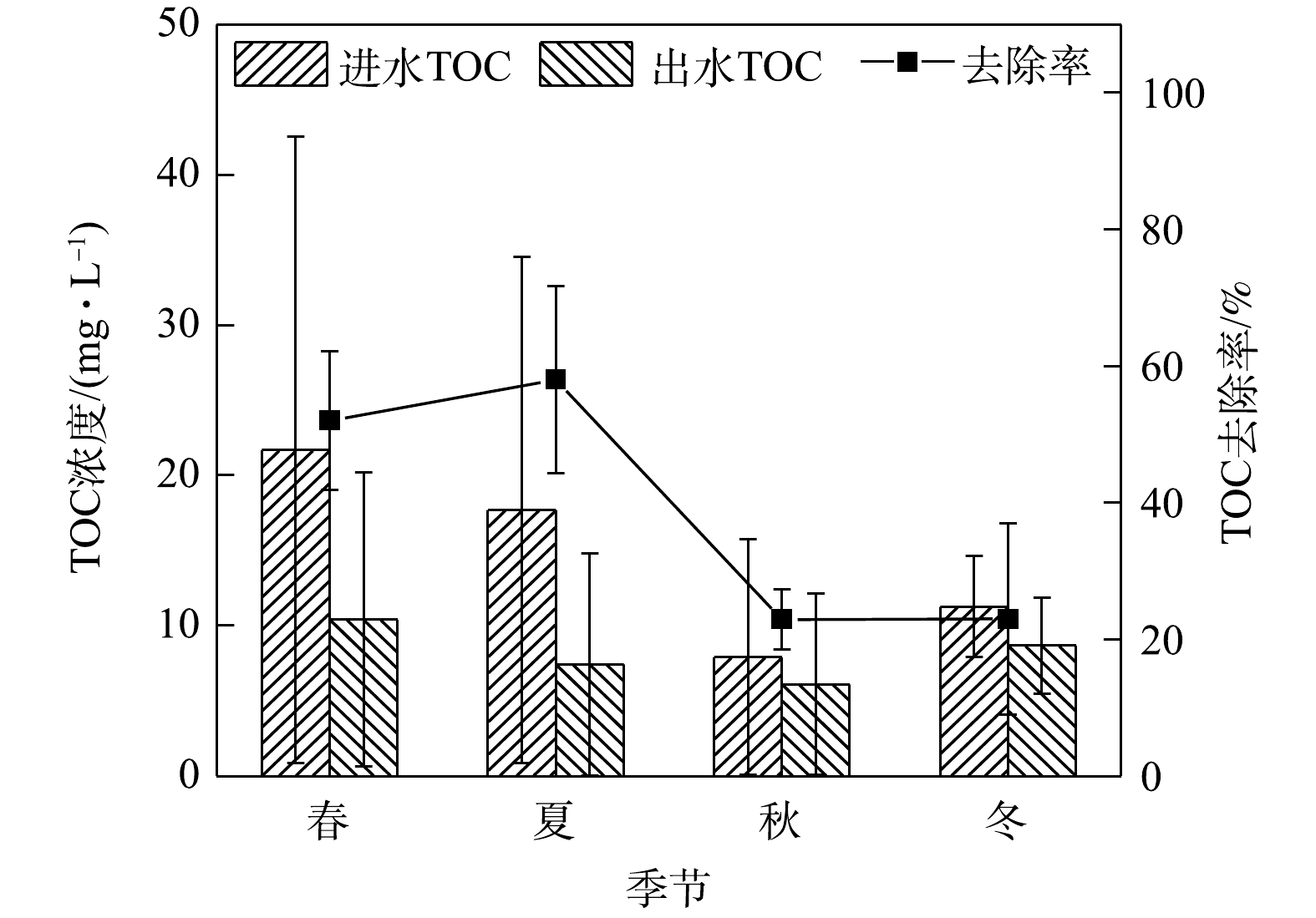
PO3−4 可通过化学键或离子键结合而生成磷酸盐沉淀,从而实现除磷。从整体来说,生物滞留系统在磷酸盐去除方面的研究已经取得良好成果。其他学者的研究结果显示,相比普通生物滞留系统,填料改良、淹没区设置、高负荷进水等条件下改造的生物滞留系统磷酸盐去除率在90%及以上[31, 36-37]。综上所述,本研究中系统磷酸盐去除率高的原因可能是实验种植的麦冬是根系发达的植物,且填料层添加有火山岩。6)季节变化对TOC去除效果的影响。如图7所示,在实验系统中,TOC在春夏两季进水浓度高,春季平均进水浓度最高为21.68 mg·L−1,平均出水浓度为10.41 mg·L−1,相应去除率为52.01%,较秋冬两季高。这可能是因为秋季植物枯萎凋谢增加了土壤中的有机碳[38]。整体来看,系统对TOC的去除率并不理想。据了解,不同种植基质对雨水径流中的TOC浓度具有非常显著的影响[39]。总有机碳(TOC)是以碳的含量表示水中有机物的总量,水的TOC值越高,说明水中有机物含量越高。雨水径流中TOC的来源主要有2个方面:一是自然界本身,如腐烂的动植物;二是人类活动,如工业有机试剂,人畜排泄物等。实验系统中的种植层是沙土比为7∶3的人工配土,土壤则选取当地土壤以便于植物生长以及适应当地气温环境。有研究[40]表明,在土壤中掺入木质有机质后,土壤水分、肥力、有机碳含量均可得到改善进而改善微生物活性和植物生长。TOC去除率越高,说明其中的微生物对有机物的需求越高[1]。可能是系统中微生物活性的缘故,出水TOC浓度在实验系统中四季均不稳定,尤其是在春季达到极端异常值。
-
生物滞留系统对径流污染物的去除情况除了受季节影响外,填料基质也是其主要影响因素之一。有研究[4, 14, 41-42]表明,填料的类型、填料层的深度、填料组成及配比等均会影响TN的去除效果。本实验系统以火山岩矿物为填料,通过设置内部缺氧段,采用海绵铁强化除磷,可以提高其脱氮除磷能力[33]。就本实验系统的实验结果来说,其TN和TP去除率同文献报道的其他实验结果[1, 26]基本一致,且磷酸盐去除率也很理想。生物滞留系统填料层中采用的火山岩具有天然蜂窝多孔性结构,孔径间隙分布不均、表面粗糙且亲水性强,适宜微生物在其表面繁殖生长形成生物膜,有利于硝化菌的增殖生长,提高介质层的硝化能力,从而使填料层能去除更多的氨氮[18]。因为硝化菌增殖生长,对反硝化菌可能造成了一定影响,故系统硝态氮去除效果不是很理想。有研究[1]表明,海绵铁填料对TOC(约94.53%)的去除率较高。本研究实验系统未能达到理想去除效果,这可能是因为pH过高和水温过低,从而影响了TOC的去除[43]。
对比了本研究与其他改良填料基质下径流污染物的去除情况,结果如表2所示。由表2可知,本研究实验系统相比其他研究,在脱氮除磷方面同其他单一填料较占优势。但同其他改良填料相比,氮污染物及TOC去除效果相对不是很好,磷污染物去除效果与其他改良填料相差无几甚至普遍偏高。
2.1. 季节对污染物浓度去除效果影响












2.2. 填料对污染物去除的对比分析
-
1)生物滞留系统对总氮和氨氮的去除效果趋势均为夏冬两季高于春秋两季,去除率均在冬季最高,分别达到69.53%和96.88%;而对于硝态氮来说,整体去除率较低,在28.98%~47.18%内波动,且受季节的影响较大。
2)生物滞留系统对总磷和磷酸盐的去除效果受到季节的影响最小,四季去除率波动幅度不大,且长时间稳定保持在90%以上;TOC全年平均去除率为38.96%,夏季去除率最高为58.01%,春夏两季去除率较秋冬两季去除率高,受季节影响较大。
3)本研究的实验系统的脱氮除磷性能较其他单一填料更有优势,但同其他改良填料相比,氮污染物及TOC去除效果并不是很好,磷污染物去除情况与其相差不大且去除率较高。
