


 下载:
下载:
-
目前,国内外普遍在水源水体中检测到了不同种类的微量污染物[1-2]。这些微量污染物经过传统水处理工艺包括混凝、沉淀、过滤很难有效去除[3-4]。因此,高级氧化工艺如催化臭氧、紫外过氧化氢、紫外氯联合、芬顿等对微量污染物的强化去除方面的研究成为热点[5-8]。
多相芬顿技术作为高级氧化技术的一种,与活性炭工艺联用不仅可以去除水中天然有机物,还可以去除水中的有机农药、抗生素、内分泌干扰物等难降解的微量污染物,降低由微量污染物导致的水体毒性[9-12]。与其他深度处理技术相比,其不需要增加光、声、电等辅助设施,通常对温度和压力无要求,故具有非常广阔的应用前景[13]。
双酚A作为一种微量污染物,会影响人的内分泌系统,目前,对其降解的相关研究较多,除了化学法之外,也有采用微生物方法对其进行降解的研究[14-15]。在本研究中,首先以工业化合成的多相芬顿催化剂建立了多相芬顿催化柱,在北京某水厂进行了多相芬顿催化对天然有机物(NOM)的降解去除研究。在经过200 d运行后,通过对催化柱不同位置的催化剂进行取样,利用小试实验考察了生物膜对不同浓度双酚A去除效果的影响,最后为了分析生物膜的作用,对催化剂表面生物膜群落结构等进行了表征。本研究结果表明多相芬顿技术是一种很有应用前景的去除微量污染物的实用技术。
-
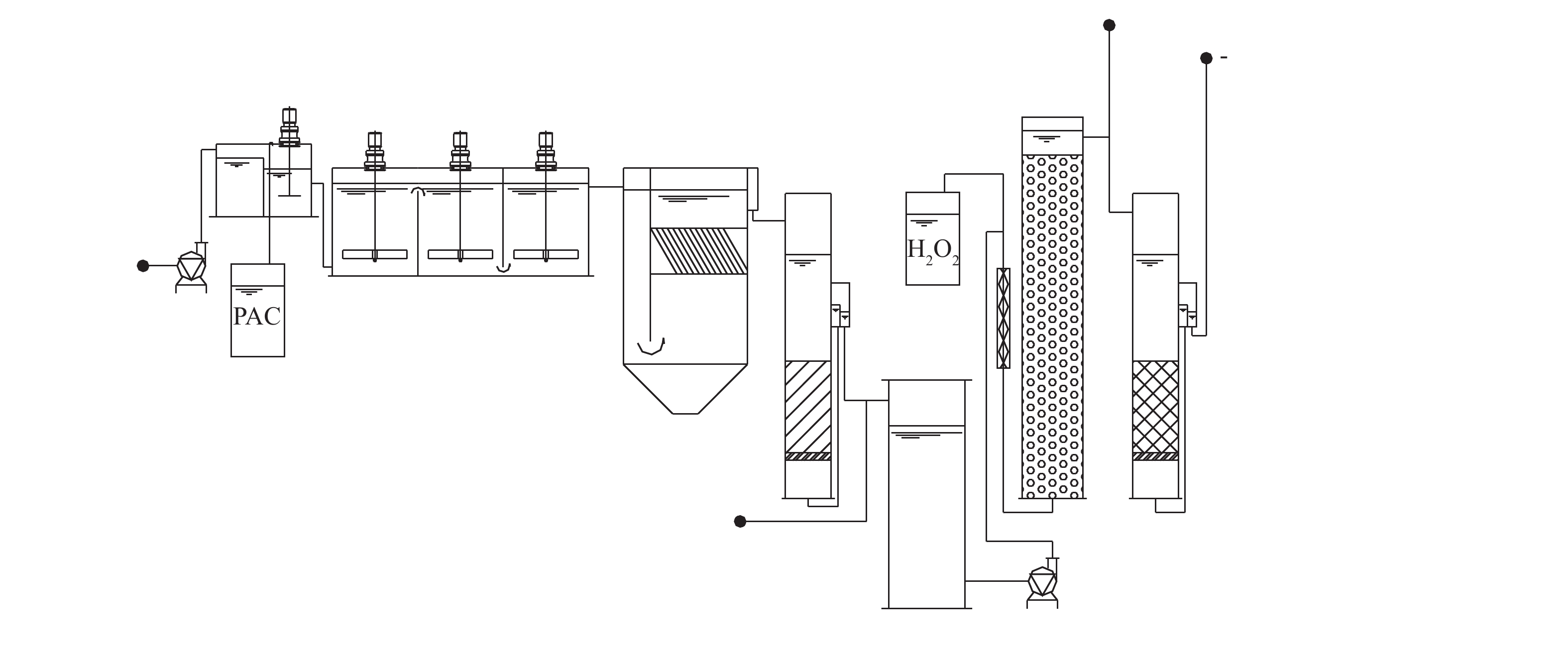
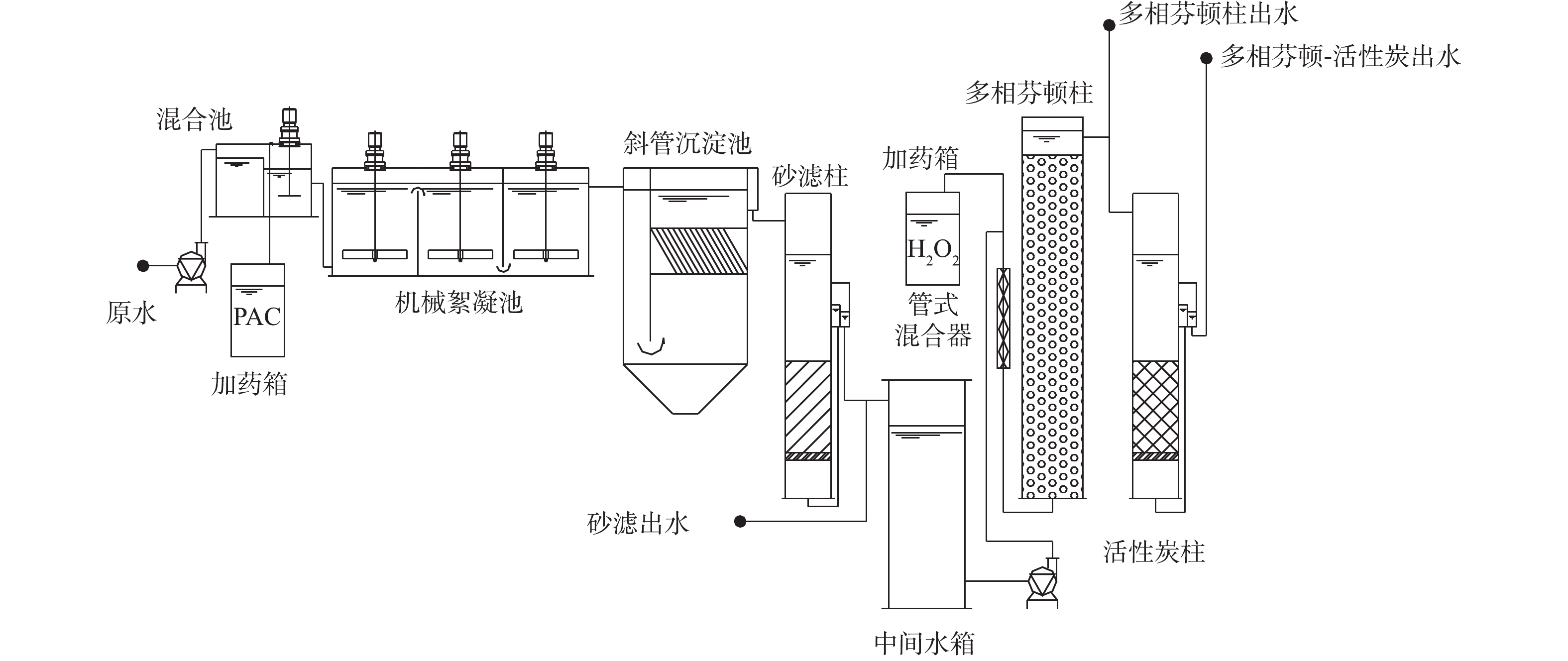
中试装置建在北京市某自来水厂实验基地内,装置采用的水处理工艺流程包括混凝-沉淀-砂滤-多相芬顿催化氧化-活性炭过滤,装置流程如图1所示。本研究所使用的多相芬顿催化剂以氧化铝小球为载体,以铜、钴等化合物为主要成分,并以吨级规模工业合成(目前尚未商品化生产),LYU等[16]对该类催化剂已有较多的研究报道。多相芬顿催化柱流量30 L·min−1、水力停留时间为15 min、投加过氧化氢量为0.15 mmol·L−1、活性炭柱流量为30 L·min−1、水力停留时间为15 min、反冲周期为7 d。
-
由于该水厂原水中没有检测到双酚A的存在,而且考虑到在水厂中人为添加微量污染物实验排水会影响环境,因此,在中试现场对水体中NOM进行了为期200 d的去除研究,同时考察了对消毒副产物和致病微生物控制效果,相关的研究结果[17]已发表。芬顿催化柱高度为4.0 m,经过长时间运行之后,在芬顿催化柱上、中、下层(柱高分别为3.6、1.8和0.5 m处)分别取适量材料,之后在实验室进行双酚A去除的小试研究。把上、中、下3层催化剂材料分别填装,共填装6根小柱(柱子尺寸40 mm×30 cm):3根小柱分别装填上、中、下3层的多相芬顿催化剂,另外3根小柱装填上、中、下3层经过超声振荡并清洗后无生物膜的多相芬顿催化剂。
实验分别在2种条件下运行,一种配制进水双酚A浓度为10 μg·L−1,运行条件与中试相同,停留时间为15 min;另一种条件配制进水双酚A浓度为5 mg·L−1、停留时间为40 min。2种实验条件分别连续运行7 d,并于第2、4和6天取样,分析2种条件下双酚A的去除率,且考察了生物膜对双酚A去除效果的影响。
-
中试实验流程进水采用水厂水源水经预氯化后水,经检测水质如下:温度为(20±2) ℃、浊度为3.0 NTU、DOC为(3.5±0.6) mg·L−1、pH=7.6~8.6、硬度为80~126 mg·L−1(以CaCO3计)、碱度为75~126 mg·L−1(以CaCO3计)、其他各种离子(2~3 mg·L−1
NO−3 -N、4~8 mg·L−1 Cl−、0.2~0.5 mg·L−1 F−、21~36 mg·L−1SO2−4 )。 -
水中溶解性有机碳 (DOC) 采用TOC分析仪(TOC-VCPH,SHIMADZU, Japan)。H2O2浓度利用辣根过氧化氢酶采用分光光度法分析。
在进行催化剂上生物膜采样时,首先把催化剂进行冷冻干燥然后称取一定量的催化剂利用灭菌棉签擦拭催化剂表面,并用磷酸盐缓冲溶液(PBS)进行冲洗,然后把棉签和磷酸盐缓冲溶液一起进行超声震荡,收集PBS溶液[17]。催化柱上中下层出水各采集1 L溶液。上、中、下3层出水及生物膜各取样3个,分别标记为生物膜上层(RS1.1、RS1.2、RS1.3),生物膜中层(RS2.1、RS2.2、RS2.3),生物膜下层(RS3.1、RS3.2、RS3.3),上层出水(RS4.1、RS4.2、RS4.3),中层出水(RS5.1、RS5.2、RS5.3)和下层出水(RS6.1、RS6.2、RS6.3)。
把溶液过0.45 µm聚碳酸酯膜,然后用Fast DNA spin kit for soil(Qbiogene,Solon,OH)试剂盒进行DNA基因提取。利用7300实时荧光定量聚合酶链式反应系统仪器(7300 Real Time PCR System)分析检测总细菌(16S rRNA),用于表征催化剂表面的微生物量。同时基于IonS5TMXL测序平台,利用单端测序方法构建小片段文库进行单端测序。通过对Reads剪切过滤,OTU(operational taxonomic units)聚类,β多样性分析采用算术均值的非加权配对平均法(unweighted pair-group method with arithmetic mean, UPGMA)聚类分析。最后根据16S测序数据进行基于KEGG数据库的功能预测。
对传统的热提取方法进行了优化,以提取生物膜中微生物的胞外多聚物(EPS),具体步骤参照文献中的方法[18]。然后用Folin苯酚比色法测定EPS中的蛋白质,以牛血清白蛋白为标准物质。用苯酚硫酸比色法测定EPS中的多糖,以葡萄糖为标准物质,并使用三维荧光(EEM)对EPS进行成分分析。
低浓度双酚A首先利用固相萃取柱进行浓缩,然后利用超高效液相色谱四极杆-飞行时间质谱联用仪(UPLC-Q-TOF-MS,ACQUITY UPLC/Xevo G2 Q-TOF,Waters)。
-
多相芬顿催化柱中试实验连续运行200 d,运行周期内多相芬顿催化柱进出水溶解性有机碳(DOC)变化如图2所示。砂滤出水(多相芬顿催化柱进水)的DOC为(3.12±0.51) mg·L−1,经过多相芬顿催化剂处理后DOC值降低至(1.96±0.49) mg·L−1,在相同运行时间条件下比砂滤出水降低了42%~63%。由图2可以看出,多相芬顿催化柱对NOM有很好的去除效果。另外,在中试运行120 d以后,多相芬顿出水DOC出现稳中有降的趋势,这可能与生物膜的形成有关。
-
为了分析生物膜对芬顿催化剂去除双酚A的影响,我们对上、中、下3层具有不同生物膜的催化剂材料分别进行了小试实验。根据国家标准《生活饮用水卫生标准》(GB 5749-2006)中生活饮用水水质参考指标及限值规定,饮用水中双酚A浓度不得高于10 µg·L−1,因此,我们选择此浓度为低浓度进行研究。图3反映了不同起始浓度下双酚A的降解率。如图3(a)所示,在停留时间仍然为15 min且在不加H2O2的条件下,低浓度(10 µg·L−1)双酚A在无生物膜的上、中、下3层催化剂中的去除率分别为24.9%、27.6%和28.3%,而在有生物膜的上、中、下3层催化剂上去除率分别为36.0%、37.4%和37.8%。该结果说明,上、中、下3层催化剂洗去生物膜后,对双酚A去除效果基本相同,而具有不同生物膜群落组成的上、中、下3层催化剂对其去除效果也基本相同。但是,有生物膜的催化剂明显比没有生物膜的催化剂去除效果更好。
除了低浓度双酚A,本研究还进行了高浓度5 mg·L−1双酚A的去除实验,由于其浓度较高,控制停留时间为40 min,结果如图3(b)所示。与低浓度双酚A的实验结果类似,有生物膜的催化剂比没有生物膜催化剂对双酚A的去除率较高,而且有生物膜的催化剂从上层到下层其对双酚A的去除能力有逐渐增强趋势,在下层中对双酚A的去除率达到了39%。
因此,在中试条件下,控制多相芬顿催化柱流量30 L·min−1,水力停留时间为15 min,投加过氧化氢量为0.15 mmol·L−1,冲洗周期为7 d的条件下,多相芬顿催化柱表面形成的生物膜影响了小试过程中双酚A的去除,使得双酚A去除率有所提高,同时发现,随着停留时间的延长,下层生物膜影响较大。
-
1)微生物群落多样性。根据实验分析方法描述,对催化柱上中下3层进行生物膜和出水中微生物采样分析,图4为6组样品的所有平行样,共18个,在微生物门水平(phylum level)上对其多样性基于进化序列差异(unweighted unifrac)距离矩阵进行的UPGMA聚类树分析。由图4可以看出,生物膜和水中微生物各自聚类在一起,这说明生物膜和水中微生物存在差别。有研究[19]表明,过滤过程中生物膜对其出水中微生物存在决定作用,该结果上层出水与生物膜相似性高,中下层出水与生物膜差别大,这可能是由于H2O2从催化剂底部投加,中、下层为H2O2初始反应阶段,产生的羟基自由基对水体中微生物生长影响较大。另外,中层生物膜RS2.1、RS2.2、RS2.3和下层生物膜RS3.1、RS3.2、RS3.3聚类在一起,相似度较高,与上层生物膜有一定差别,该结果同样说明,随着反应不断进行,H2O2浓度有搜降低,从而影响了生物膜群落组成。有研究[20]表明,加氯消毒过程对微生物群落结构影响较大,该结果也说明,多相芬顿反应投加H2O2能够影响微生物群落结构组成。
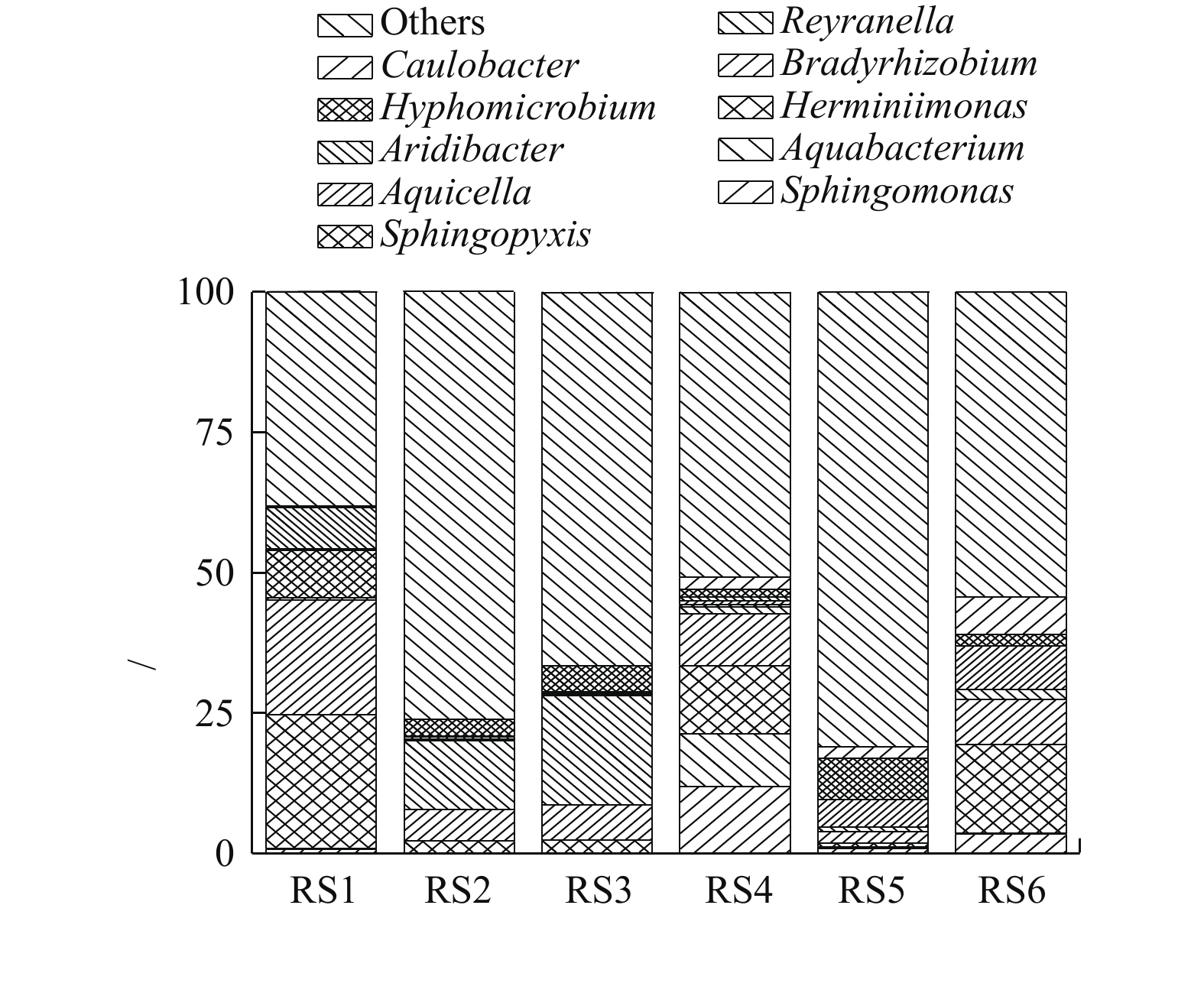
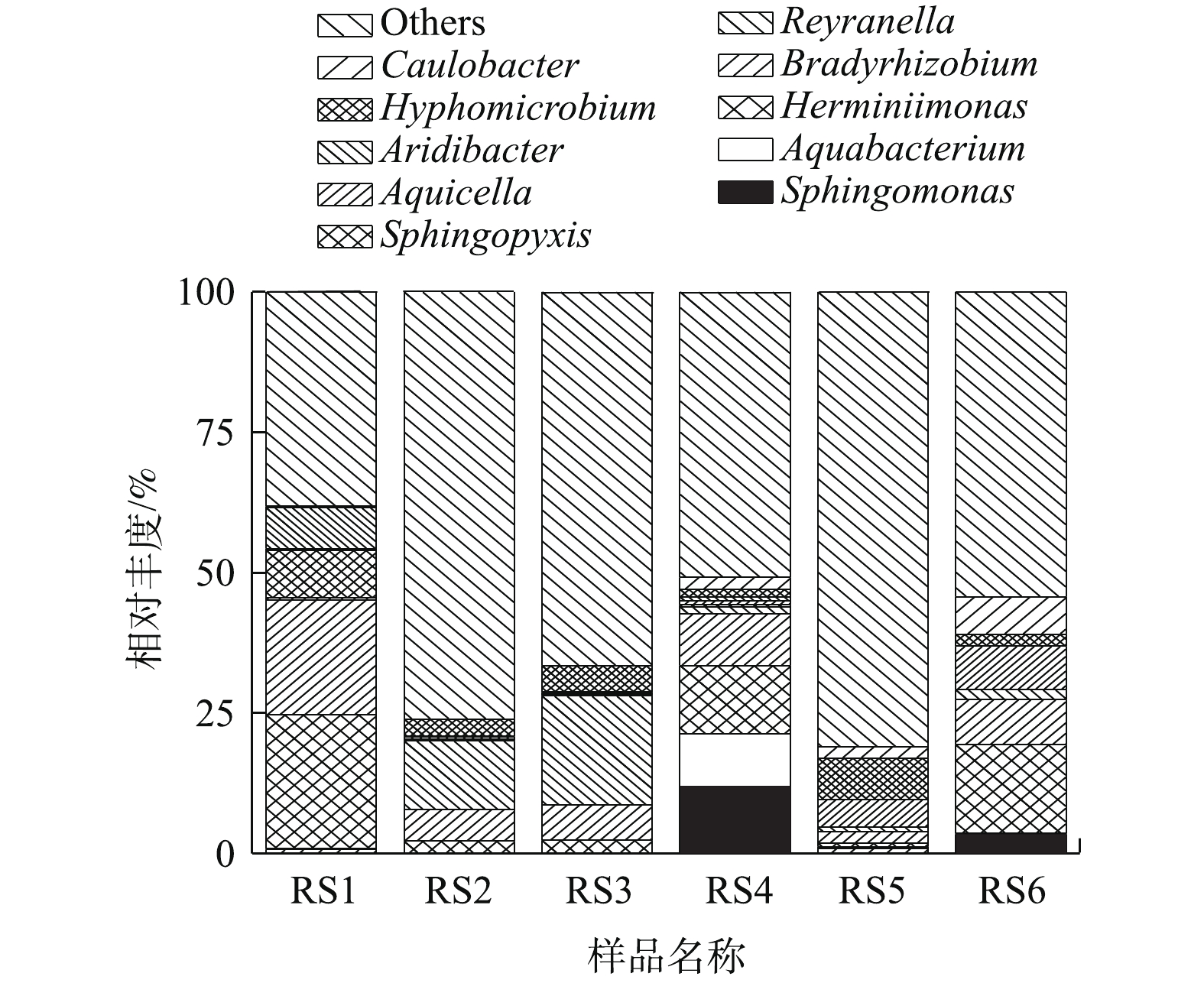
2)微生物群落组成。为了进一步分析生物膜及出水中微生物群落组成,对生物膜及出水微生物在属水平上相对丰度在前10的微生物进行作图分析(每组样品3个平行样取平均值作图),如图5所示。通过对比分析发现,上层生物膜RS1中赫山单胞菌属(Herminiimonas)、慢生根瘤菌属(Bardyrhizobium)、鞘氨醇盒菌属(Sphingopyxis)和旱杆菌属(Aridibacter)4种微生物菌属较多,其对应的相对丰度分别为23.9%、20.5%、8.5%和7.3%。中层生物膜RS2中这4种菌属分别降低至2.1%、5.6%、0.3%和0.06%,下层生物膜RS3这4种微生物菌种降低至2.2%、6.3%、0.3%和0.2%。但是在中层RS2和下层RS3生物膜中Reyranella菌属和生丝微菌属(Hyphomicrobium)含量明显升高。RS2中相对丰度分别为12.2%和3.1%,RS3中分别为19.4%和4.7%。另外,催化柱上、中、下3层出水微生物变化较大,在上层出水RS4中鞘氨醇单胞菌属(Sphingomonas)、水杆菌属(Aquabacterium)、赫山单胞菌属(Herminiimonas)和慢生根瘤菌属(Bardyrhizobium)相对丰度较高。中层出水RS5中Aquicella和生丝微菌属(Hyphomicrobium)相对丰度较高。下层出水RS6中赫山单胞菌属(Herminiimonas)、慢生根瘤菌属(Bardyrhizobium)、柄杆菌属(Caulobacter)相对丰度高。结果表明,相比于生物膜,不同层出水中微生物群落组成变化较大。另外,投加H2O2是从催化柱下面投加,H2O2接触到催化剂表面生成羟基自由基(∙OH)对有机物进行降解并灭活微生物,随着反应进行从下层到上层H2O2浓度逐渐降低。因此,生物膜中赫山单胞菌属(Herminiimonas)、慢生根瘤菌属(Bardyrhizobium)、鞘氨醇盒菌属(Sphingopyxis)和旱杆菌属(Aridibacter)相对丰度从下层到上层逐渐增加,但是芬顿催化柱中层和下层生物膜群落组成相似,其中,Reyranella菌属和生丝微菌属(Hyphomicrobium)含量明显比上层高。
3)微生物功能分析。根据测序样品在数据库中的功能注释及丰度信息,从功能查阅层面进行聚类分析。其中,微生物功能分析包括细胞过程、环境信息处理、遗传信息处理、关于人类疾病、新陈代谢、有机系统、无法归类和无功能共8种。在环境信息处理和新陈代谢方面,中层和下层催化剂生物膜的微生物功能更强。以上结果说明,受周围环境H2O2和有机物浓度的影响,中层和下层催化剂生物膜的微生物表现出更强的代谢能力。
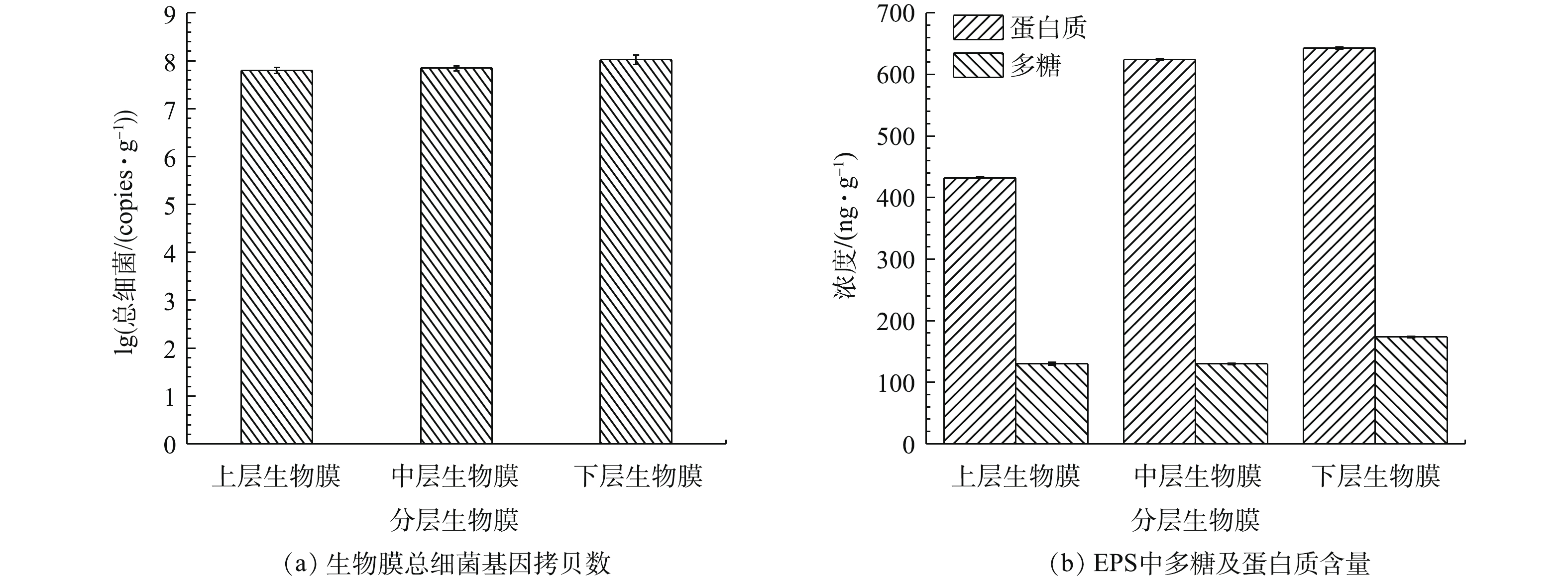
通过对生物膜上微生物16S rRNA进行定量分析,发现下层微生物总细菌含量最多,微生物基因拷贝数高达8个对数量级;中层其次,上层最少,分别为7.8个和7.7个对数量级(图6(a))。由对催化剂表面生物膜上胞外多聚物EPS表征结果可以看出,在中、下层的生物膜上微生物EPS中蛋白质含量较高,分别高达624.3 ng·g−1和642.8 ng·g−1(图6(b))。
因此,中试过程中从催化柱下部投加H2O2溶液,起始投加量为5 mg·L−1,随着反应的进行,从下到上的H2O2浓度逐渐降低,出水H2O2浓度为0.45 mg·L−1。随着催化柱从下到上H2O2浓度变化及其对有机物的降解,多相芬顿催化柱上、中、下3层出水中微生物群落组成变化较大,但是生物膜上微生物群落存在相似性,其中,中层和下层相似度高。上层生物膜中微生物以赫山单胞菌属(Herminiimonas)和慢生根瘤菌属(Bardyrhizobium)为主,而中层和下层生物膜中微生物以Reyranella菌属和生丝微菌属(Hyphomicrobium)为主。另外,中层和下层生物膜中微生物量相比上层多,代谢能力强,生物膜上微生物分泌的EPS多,蛋白质含量高,微生物吸附性能有所增强[21],这可能会影响催化柱对天然有机物和微量污染物的去除。小试实验结果证明,生物膜的形成影响了对双酚A的去除。在停留时间为15 min时,有生物膜的催化柱明显比没有生物膜的催化柱对10 µg·L−1双酚A的去除率高,但对于有生物膜的上、中、下层,其对双酚A的去除率差别不大。当停留时间在40 min后,有生物膜的催化柱比没有生物膜的催化柱对5 mg·L−1双酚A去除率要高,而且下层对双酚A的去除率最高,可能与下层生物膜中Reyranella菌属和生丝微菌属(Hyphomicrobium)含量较高有关。
在多相芬顿催化体系中,H2O2除了与释放到溶液中的少量金属离子发生链反应外,主要与催化剂发生界面反应,多相芬顿反应需要高活性的催化剂,其可提高H2O2的利用率[13]。本研究结果表明,在多相芬顿催化柱运行过程中,很难避免生物膜的形成,而对于微量污染物双酚A,生物膜可以提高对其的去除效果。但是,生物膜对不同种类和浓度的污染物去除的影响还需要进行深入的研究和探索,从而更好的优化运行多相芬顿催化柱,以便能尽早的解决实际问题。另外,建议今后要深入开展生物膜对芬顿反应去除不同微量污染物的影响研究。
-
1)多相芬顿催化柱在中试运行过程中对天然有机物表现出良好的去除效果,而实验过程中形成的生物膜可提高对双酚A的去除率。在无生物膜条件下,多相芬顿反应对双酚A的去除率低于30%,而在生物膜形成后,对双酚A的去除率可提高到36%~39%。
2)通过对催化柱上生物膜进行表征,发现催化柱生物膜上微生物群落存在相似性,其中,中层和下层相似度高。上层生物膜中微生物以赫山单胞菌属(Herminiimonas)和慢生根瘤菌属(Bardyrhizobium)为主,而中层和下层生物膜中微生物以Reyranella菌属和生丝微菌属(Hyphomicrobium)为主。
3)随着生物膜上微生物群落组成的变化,多相芬顿催化柱由上层到下层生物膜上生物量有所增加,微生物代谢活性增强,从而分泌更多的胞外多聚物,这可能是有生物膜的催化柱特别是下层催化柱可更好地去除双酚A的主要原因。
多相芬顿催化剂表面生物膜对去除双酚A的影响及其微生物群落表征
Effects of the surface biofilms of heterogeneous Fenton catalyst on the bisphenol A removal and the characterization of its bacterial community
-
摘要: 为了分析经长期运行的多相芬顿催化柱上生物膜的群落结构特征及其对双酚A去除的影响,利用中试实验对多相芬顿催化降解天然有机物(NOM)进行了为期200 d的跟踪检测,并利用小试实验对比研究了在有无生物膜以及催化柱不同位置上的生物膜对多相芬顿反应去除双酚A的影响效果。同时采集了多相芬顿催化柱不同位置的催化剂样品,对样品表面的生物膜群落结构、微生物量、代谢活性以及胞外多聚物(EPS)进行了系统的表征。结果表明:多相芬顿对NOM有很好的去除效果,而且生物膜的形成使得对双酚A的去除率可提高到36%~39%;此外,催化柱生物膜上微生物群落存在相似性,其中层和下层相似度更高;上层生物膜中微生物以赫山单胞菌属(Herminiimonas)和慢生根瘤菌属(Bardyrhizobium)为主,而中层和下层生物膜中微生物以Reyranella菌属和生丝微菌属(Hyphomicrobium)为主;随着微生物群落的变化,中试催化柱由上层到下层生物量有所增加,微生物代谢活性增强,可分泌更多的EPS,这可能是生物膜特别是下层生物膜提高对双酚A去除效果的主要原因。以上研究结果证实,多相芬顿技术是一种很有应用前景的去除微量污染物的实用技术。Abstract: In order to investigate the biofilms communities on the surface of heterogeneous Fenton catalyst after long-time running and their effects on the removal of bisphenol A, the heterogeneous Fenton was studied in a pilot scale to test its effect on the removal of natural organic matter (NOM) for 200 d. The effects of biofilms and the biofilms at different places of catalyst column on the removal of bisphenol A were also studied using a bench scale test. Meanwhile, different catalyst samples at the different places of the catalyst column were collected, and the biofilms bacterial community structure, bacterial biomass, bacterial metabolism, and the composition of extracellular polymeric substances (EPS) on the sample surface were analyzed. The results showed that heterogeneous Fenton had a good performance on the removal of NOM, and the formed biofilms improved the removal rate of bisphenol A to 36%~39%. Moreover, the results also indicated that the biofilms showed great similarity, especially for the biofilms in the middle and lower layers. The bacterial genera in upper layers were dominated by Herminiimonas and Bardyrhizobium, while in middle and lower layers, the bacterial genera were dominated by Reyranella Hyphomicrobium. Along with the changes of bacterial community, the biomass increased in the biofilms of catalyst from top to bottom. The function of bacterial metabolism also increased, and the bacteria could produce more EPS. This may be the main reason for the more removal of bisphenol A by biofilms adsorption, especially for the biofilms in the lower layers. The results indicated that the heterogeneous Fenton was a practical technique with promising applications for removing micropollutants.
-
目前,国内外普遍在水源水体中检测到了不同种类的微量污染物[1-2]。这些微量污染物经过传统水处理工艺包括混凝、沉淀、过滤很难有效去除[3-4]。因此,高级氧化工艺如催化臭氧、紫外过氧化氢、紫外氯联合、芬顿等对微量污染物的强化去除方面的研究成为热点[5-8]。
多相芬顿技术作为高级氧化技术的一种,与活性炭工艺联用不仅可以去除水中天然有机物,还可以去除水中的有机农药、抗生素、内分泌干扰物等难降解的微量污染物,降低由微量污染物导致的水体毒性[9-12]。与其他深度处理技术相比,其不需要增加光、声、电等辅助设施,通常对温度和压力无要求,故具有非常广阔的应用前景[13]。
双酚A作为一种微量污染物,会影响人的内分泌系统,目前,对其降解的相关研究较多,除了化学法之外,也有采用微生物方法对其进行降解的研究[14-15]。在本研究中,首先以工业化合成的多相芬顿催化剂建立了多相芬顿催化柱,在北京某水厂进行了多相芬顿催化对天然有机物(NOM)的降解去除研究。在经过200 d运行后,通过对催化柱不同位置的催化剂进行取样,利用小试实验考察了生物膜对不同浓度双酚A去除效果的影响,最后为了分析生物膜的作用,对催化剂表面生物膜群落结构等进行了表征。本研究结果表明多相芬顿技术是一种很有应用前景的去除微量污染物的实用技术。
1. 材料与方法
1.1 中试装置及运行条件
中试装置建在北京市某自来水厂实验基地内,装置采用的水处理工艺流程包括混凝-沉淀-砂滤-多相芬顿催化氧化-活性炭过滤,装置及流程图如图1所示。本研究所使用的多相芬顿催化剂以氧化铝小球为载体,以铜、钴等化合物为主要成分,并以吨级规模工业合成(目前尚未商品化生产),LYU等[16]对该类催化剂已有较多的研究报道。多相芬顿催化柱流量30 L·min−1、水力停留时间为15 min、投加过氧化氢量为0.15 mmol·L−1、活性炭柱流量为30 L·min−1、水力停留时间为15 min、反冲周期为7 d。
1.2 小试装置及运行条件
由于该水厂原水中没有检测到双酚A的存在,而且考虑到在水厂中人为添加微量污染物实验排水会影响环境,因此,在中试现场对水体中NOM进行了为期200 d的去除研究,同时考察了对消毒副产物和致病微生物控制效果,相关的研究结果[17]已发表。芬顿催化柱高度为4.0 m,经过长时间运行之后,在芬顿催化柱上、中、下层(柱高分别为3.6、1.8和0.5 m处)分别取适量材料,之后在实验室进行双酚A去除的小试研究。把上、中、下3层催化剂材料分别填装,共填装6根小柱(柱子尺寸40 mm×30 cm):3根小柱分别装填上、中、下3层的多相芬顿催化剂,另外3根小柱装填上、中、下3层经过超声振荡并清洗后无生物膜的多相芬顿催化剂。
实验分别在2种条件下运行,一种配制进水双酚A浓度为10 μg·L−1,运行条件与中试相同,停留时间为15 min;另一种条件配制进水双酚A浓度为5 mg·L−1、停留时间为40 min。2种实验条件分别连续运行7 d,并于第2、4和6天取样,分析2种条件下双酚A的去除率,且考察了生物膜对双酚A去除效果的影响。
1.3 中试进水水质
中试实验流程进水采用水厂水源水经预氯化后水,经检测水质如下:温度为(20±2) ℃、浊度为3.0 NTU、DOC为(3.5±0.6) mg·L−1、pH=7.6~8.6、硬度为80~126 mg·L−1(以CaCO3计)、碱度为75~126 mg·L−1(以CaCO3计)、其他各种离子(2~3 mg·L−1
NO−3 -N、4~8 mg·L−1 Cl-、0.2~0.5 mg·L−1 F-、21~36 mg·L−1SO2−4 )。1.4 分析方法
水中溶解性有机碳 (DOC) 采用TOC分析仪(TOC-VCPH,SHIMADZU, Japan)。H2O2浓度利用辣根过氧化氢酶采用分光光度法分析。
在进行催化剂上生物膜采样时,首先把催化剂进行冷冻干燥然后称取一定量的催化剂利用灭菌棉签擦拭催化剂表面,并用磷酸盐缓冲溶液(PBS)进行冲洗,然后把棉签和磷酸盐缓冲溶液一起进行超声震荡,收集PBS溶液[17]。催化柱上中下层出水各采集1 L溶液。上、中、下3层出水及生物膜各取样3个,分别标记为生物膜上层(RS1.1、RS1.2、RS1.3),生物膜中层(RS2.1、RS2.2、RS2.3),生物膜下层(RS3.1、RS3.2、RS3.3),上层出水(RS4.1、RS4.2、RS4.3),中层出水(RS5.1、RS5.2、RS5.3)和下层出水(RS6.1、RS6.2、RS6.3)。
把溶液过0.45 µm聚碳酸酯膜,然后用Fast DNA spin kit for soil(Qbiogene,Solon,OH)试剂盒进行DNA基因提取。利用7300实时荧光定量聚合酶链式反应系统仪器(7300 Real Time PCR System)分析检测总细菌(16S rRNA),用于表征催化剂表面的微生物量。同时基于IonS5TMXL测序平台,利用单端测序方法构建小片段文库进行单端测序。通过对Reads剪切过滤,OTU(operational taxonomic units)聚类,β多样性分析采用算术均值的非加权配对平均法(unweighted pair-group method with arithmetic mean, UPGMA)聚类分析。最后根据16S测序数据进行基于KEGG数据库的功能预测。
对传统的热提取方法进行了优化,以提取生物膜中微生物的胞外多聚物(EPS),具体步骤参照文献中的方法[18]。然后用Folin苯酚比色法测定EPS中的蛋白质,以牛血清白蛋白为标准物质。用苯酚硫酸比色法测定EPS中的多糖,以葡萄糖为标准物质,并使用三维荧光(EEM)对EPS进行成分分析。
低浓度双酚A首先利用固相萃取柱进行浓缩,然后利用超高效液相色谱四极杆-飞行时间质谱联用仪(UPLC-Q-TOF-MS,ACQUITY UPLC/Xevo G2 Q-TOF,Waters)。
2. 结果与讨论
2.1 多相芬顿对溶解性有机碳(DOC)的去除效果
多相芬顿催化柱中试实验连续运行200 d,运行周期内多相芬顿催化柱进出水溶解性有机碳(DOC)变化如图2所示。砂滤出水(多相芬顿催化柱进水)的DOC为(3.12±0.51) mg·L−1,经过多相芬顿催化剂处理后DOC值降低至(1.96±0.49) mg·L−1,在相同运行时间条件下比砂滤出水降低了42%~63%。由图2可以看出,多相芬顿催化柱对NOM有很好的去除效果。另外,在中试运行120 d以后,多相芬顿出水DOC出现稳中有降的趋势,这可能与生物膜的形成有关。
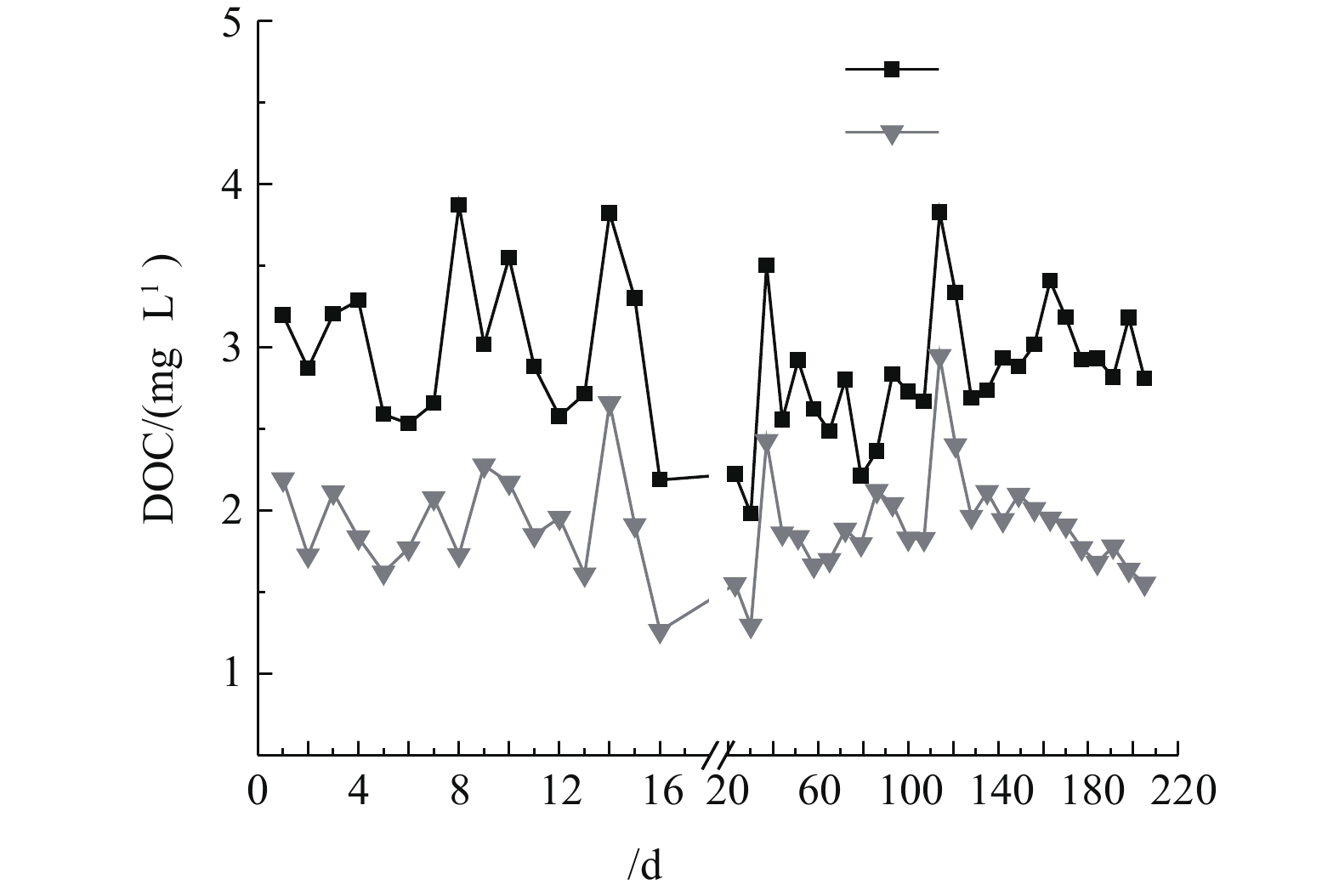 图 2 多相芬顿催化柱运行期间进出水DOC的变化Figure 2. Changes of DOC in influents and effluents during the running process of the heterogeneous Fenton column
图 2 多相芬顿催化柱运行期间进出水DOC的变化Figure 2. Changes of DOC in influents and effluents during the running process of the heterogeneous Fenton column2.2 生物膜对双酚A去除的影响
为了分析生物膜对芬顿催化剂去除双酚A的影响,我们对上、中、下3层具有不同生物膜的催化剂材料分别进行了小试实验。根据国家标准《生活饮用水卫生标准》(GB 5749-2006)中生活饮用水水质参考指标及限值规定,饮用水中双酚A浓度不得高于10 µg·L−1,因此,我们选择此浓度为低浓度进行研究。图3反映了不同起始浓度下双酚A的降解率。如图3(a)所示,在停留时间仍然为15 min且在不加H2O2的条件下,低浓度(10 µg·L−1)双酚A在无生物膜的上、中、下3层催化剂中的去除率分别为24.9%、27.6%和28.3%,而在有生物膜的上、中、下3层催化剂上去除率分别为36.0%、37.4%和37.8%。该结果说明,上、中、下3层催化剂洗去生物膜后,对双酚A去除效果基本相同,而具有不同生物膜群落组成的上、中、下3层催化剂对其去除效果也基本相同。但是,有生物膜的催化剂明显比没有生物膜的催化剂去除效果更好。
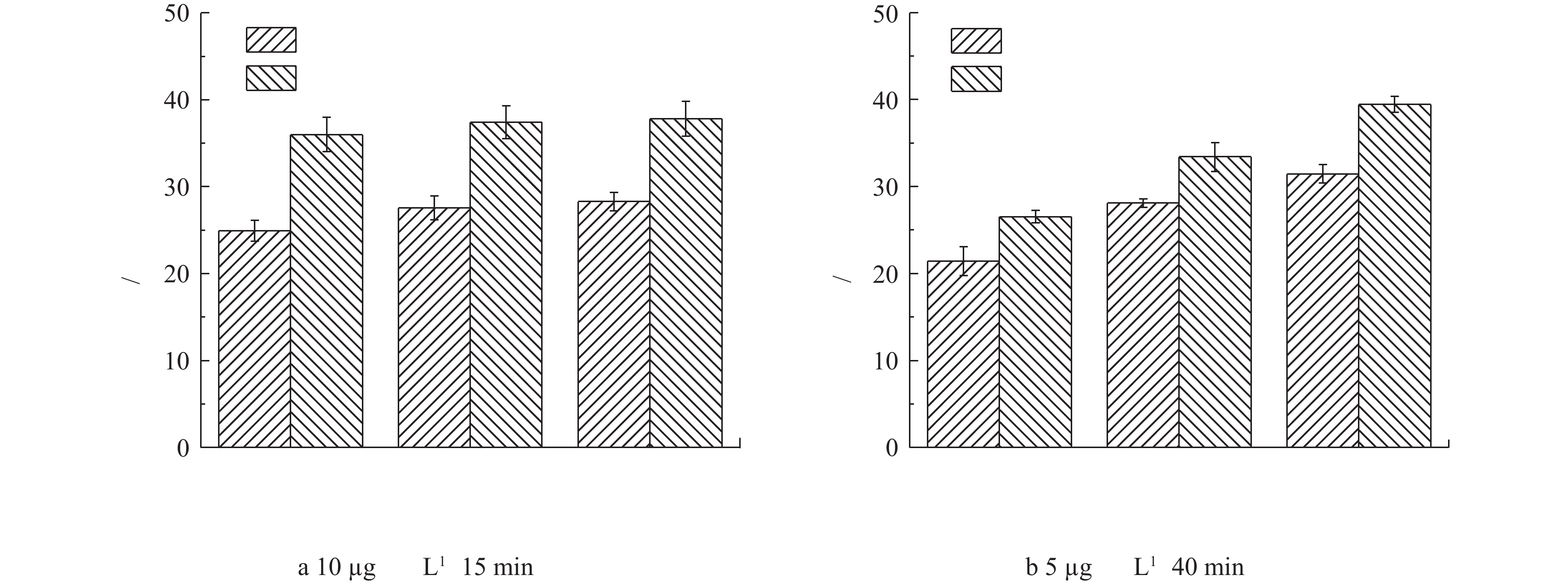 图 3 不同起始浓度下双酚A的去除率Figure 3. Removal rate of bisphenol A at different initial concentrations
图 3 不同起始浓度下双酚A的去除率Figure 3. Removal rate of bisphenol A at different initial concentrations除了低浓度双酚A,本研究还进行了高浓度5 mg·L−1双酚A的去除实验,由于其浓度较高,控制停留时间为40 min,结果如图3(b)所示。与低浓度双酚A的实验结果类似,有生物膜的催化剂比没有生物膜催化剂对双酚A的去除率较高,而且有生物膜的催化剂从上层到下层其对双酚A的去除能力有逐渐增强趋势,在下层中对双酚A的去除率达到了39%。
因此,在中试条件下,控制多相芬顿催化柱流量30 L·min−1,水力停留时间为15 min,投加过氧化氢量为0.15 mmol·L−1,冲洗周期为7 d的条件下,多相芬顿催化柱表面形成的生物膜影响了小试过程中双酚A的去除,使得双酚A去除率有所提高,同时发现,随着停留时间的延长,下层生物膜影响较大。
2.3 多相芬顿催化剂表面生物膜表征
1)微生物群落多样性。根据实验分析方法描述,对催化柱上中下3层进行生物膜和出水中微生物采样分析,图4所示为6组样品的所有平行样,共18个,在微生物门水平(phylum level)上对其多样性基于进化序列差异(unweighted unifrac)距离矩阵进行的UPGMA聚类树分析。由图4可以看出,生物膜和水中微生物各自聚类在一起,这说明生物膜和水中微生物存在差别。有研究[19]表明,过滤过程中生物膜对其出水中微生物存在决定作用,该结果上层出水与生物膜相似性高,中下层出水与生物膜差别大,这可能是由于H2O2从催化剂底部投加,中、下层为H2O2初始反应阶段,产生的羟基自由基对水体中微生物生长影响较大。另外,中层生物膜RS2.1、RS2.2、RS2.3和下层生物膜RS3.1、RS3.2、RS3.3聚类在一起,相似度较高,与上层生物膜有一定差别,该结果同样说明,随着反应不断进行,H2O2浓度有搜降低,从而影响了生物膜群落组成。有研究[20]表明,加氯消毒过程对微生物群落结构影响较大,该结果也说明,多相芬顿反应投加H2O2能够影响微生物群落结构组成。
 图 4 基于Unweighted Unifrac距离的UPGMA聚类树Figure 4. UPGMA cluster tree based on Unweighted Unifrac distance matrix
图 4 基于Unweighted Unifrac距离的UPGMA聚类树Figure 4. UPGMA cluster tree based on Unweighted Unifrac distance matrix2)微生物群落组成。为了进一步分析生物膜及出水中微生物群落组成,对生物膜及出水微生物在属水平上相对丰度在前10的微生物进行作图分析(每组样品3个平行样取平均值作图),如图5所示。通过对比分析发现,上层生物膜RS1中赫山单胞菌属(Herminiimonas)、慢生根瘤菌属(Bardyrhizobium)、鞘氨醇盒菌属(Sphingopyxis)和旱杆菌属(Aridibacter)4种微生物菌属较多,其对应的相对丰度分别为23.9%、20.5%、8.5%和7.3%。中层生物膜RS2中这4种菌属分别降低至2.1%、5.6%、0.3%和0.06%,下层生物膜RS3这4种微生物菌种降低至2.2%、6.3%、0.3%和0.2%。但是在中层RS2和下层RS3生物膜中Reyranella菌属和生丝微菌属(Hyphomicrobium)含量明显升高。RS2中相对丰度分别为12.2%和3.1%,RS3中分别为19.4%和4.7%。另外,催化柱上、中、下3层出水微生物变化较大,在上层出水RS4中鞘氨醇单胞菌属(Sphingomonas)、水杆菌属(Aquabacterium)、赫山单胞菌属(Herminiimonas)和慢生根瘤菌属(Bardyrhizobium)相对丰度较高。中层出水RS5中Aquicella和生丝微菌属(Hyphomicrobium)相对丰度较高。下层出水RS6中赫山单胞菌属(Herminiimonas)、慢生根瘤菌属(Bardyrhizobium)、柄杆菌属(Caulobacter)相对丰度高。结果表明,相比于生物膜,不同层出水中微生物群落组成变化较大。另外,投加H2O2是从催化柱下面投加,H2O2接触到催化剂表面生成羟基自由基(∙OH)对有机物进行降解并灭活微生物,随着反应进行从下层到上层H2O2浓度逐渐降低。因此,生物膜中赫山单胞菌属(Herminiimonas)、慢生根瘤菌属(Bardyrhizobium)、鞘氨醇盒菌属(Sphingopyxis)和旱杆菌属(Aridibacter)相对丰度从下层到上层逐渐增加,但是芬顿催化柱中层和下层生物膜群落组成相似,其中,Reyranella菌属和生丝微菌属(Hyphomicrobium)含量明显比上层高。
3)微生物功能分析。根据测序样品在数据库中的功能注释及丰度信息,从功能查阅层面进行聚类分析。其中,微生物功能分析包括细胞过程、环境信息处理、遗传信息处理、关于人类疾病、新陈代谢、有机系统、无法归类和无功能共8种。在环境信息处理和新陈代谢方面,中层和下层催化剂生物膜的微生物功能更强。以上结果说明,受周围环境H2O2和有机物浓度的影响,中层和下层催化剂生物膜的微生物表现出更强的代谢能力。
通过对生物膜上微生物16S rRNA进行定量分析,发现下层微生物总细菌含量最多,微生物基因拷贝数高达8个对数量级;中层其次,上层最少,分别为7.8个和7.7个对数量级(图6(a))。由对催化剂表面生物膜上胞外多聚物EPS表征结果可以看出,在中、下层的生物膜上微生物EPS中蛋白质含量较高,分别高达624.3 ng·g−1和642.8 ng·g−1(图6(b))。
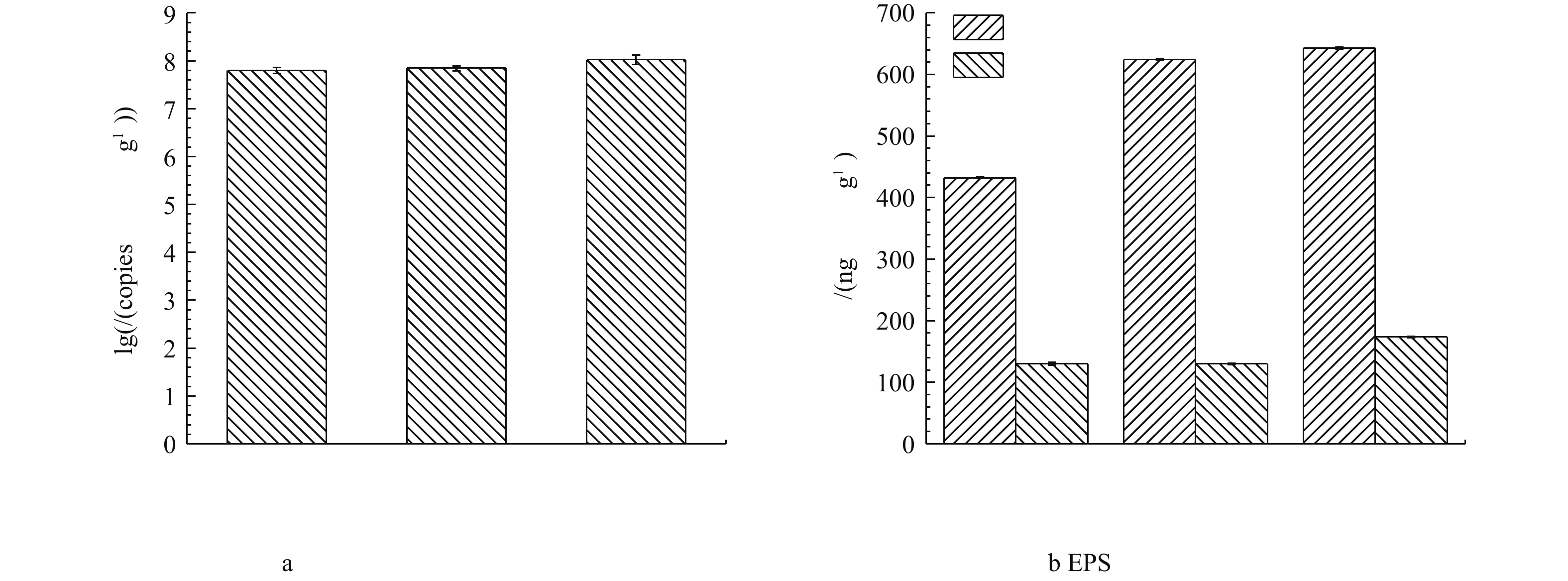 图 6 生物膜总细菌基因拷贝数和EPS中多糖及蛋白质含量Figure 6. Gene copies of total bacteria and concentration of polycarbonate and proteins in EPS of biofilm
图 6 生物膜总细菌基因拷贝数和EPS中多糖及蛋白质含量Figure 6. Gene copies of total bacteria and concentration of polycarbonate and proteins in EPS of biofilm因此,中试过程中从催化柱下部投加H2O2溶液,起始投加量为5 mg L−1,随着反应的进行,从下到上的H2O2浓度逐渐降低,出水H2O2浓度为0.45 mg L−1。随着催化柱从下到上H2O2浓度变化及其对有机物的降解,多相芬顿催化柱上、中、下3层出水中微生物群落组成变化较大,但是生物膜上微生物群落存在相似性,其中,中层和下层相似度高。上层生物膜中微生物以赫山单胞菌属(Herminiimonas)和慢生根瘤菌属(Bardyrhizobium)为主,而中层和下层生物膜中微生物以Reyranella菌属和生丝微菌属(Hyphomicrobium)为主。另外,中层和下层生物膜中微生物量相比上层多,代谢能力强,生物膜上微生物分泌的EPS多,蛋白质含量高,微生物吸附性能有所增强[21],这可能会影响催化柱对天然有机物和微量污染物的去除。小试实验结果证明,生物膜的形成影响了对双酚A的去除。在停留时间为15 min时,有生物膜的催化柱明显比没有生物膜的催化柱对10 µg·L−1双酚A的去除率高,但对于有生物膜的上、中、下层,其对双酚A的去除率差别不大。当停留时间在40 min后,有生物膜的催化柱比没有生物膜的催化柱对5 mg·L−1双酚A去除率要高,而且下层对双酚A的去除率最高,可能与下层生物膜中Reyranella菌属和生丝微菌属(Hyphomicrobium)含量较高有关。
在多相芬顿催化体系中,H2O2除了与释放到溶液中的少量金属离子发生链反应外,主要与催化剂发生界面反应,多相芬顿反应需要高活性的催化剂,其可提高H2O2的利用率[13]。本研究结果表明,在多相芬顿催化柱运行过程中,很难避免生物膜的形成,而对于微量污染物双酚A,生物膜可以提高对其的去除效果。但是,生物膜对不同种类和浓度的污染物去除的影响还需要进行深入的研究和探索,从而更好的优化运行多相芬顿催化柱,以便能尽早的解决实际问题。另外,建议今后要深入开展生物膜对芬顿反应去除不同微量污染物的影响研究。
3. 结论
1)多相芬顿催化柱在中试运行过程中对天然有机物表现出良好的去除效果,而实验过程中形成的生物膜可提高对双酚A的去除率。在无生物膜条件下,多相芬顿反应对双酚A的去除率低于30%,而在生物膜形成后,对双酚A的去除率可提高到36%~39%。
2)通过对催化柱上生物膜进行表征,发现催化柱生物膜上微生物群落存在相似性,其中,中层和下层相似度高。上层生物膜中微生物以赫山单胞菌属(Herminiimonas)和慢生根瘤菌属(Bardyrhizobium)为主,而中层和下层生物膜中微生物以Reyranella菌属和生丝微菌属(Hyphomicrobium)为主。
3)随着生物膜上微生物群落组成的变化,多相芬顿催化柱由上层到下层生物膜上生物量有所增加,微生物代谢活性增强,从而分泌更多的胞外多聚物,这可能是有生物膜的催化柱特别是下层催化柱可更好地去除双酚A的主要原因。
-
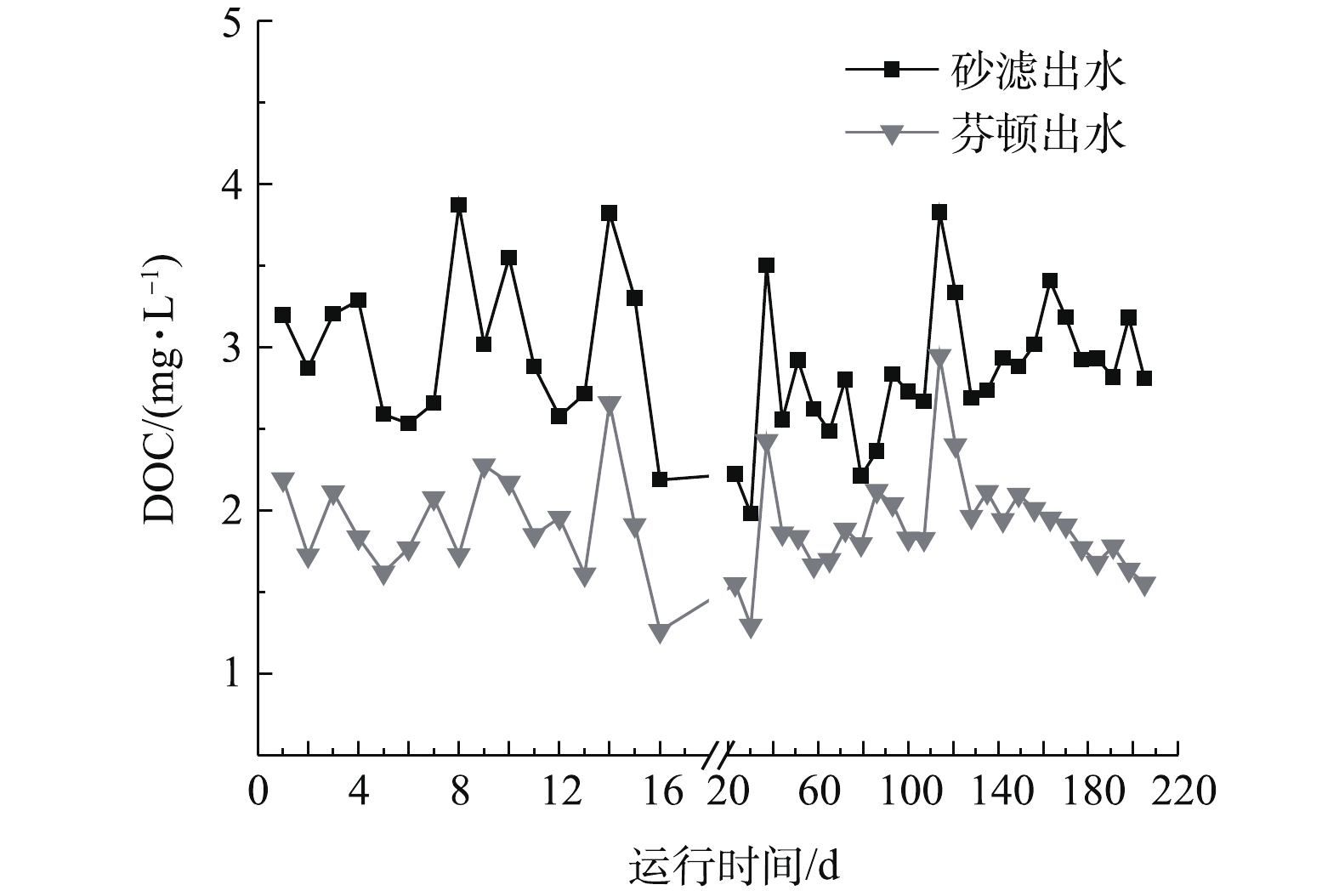
图 2 多相芬顿催化柱运行期间进出水DOC的变化
Figure 2. Changes of DOC in influents and effluents during the running process of the heterogeneous Fenton column
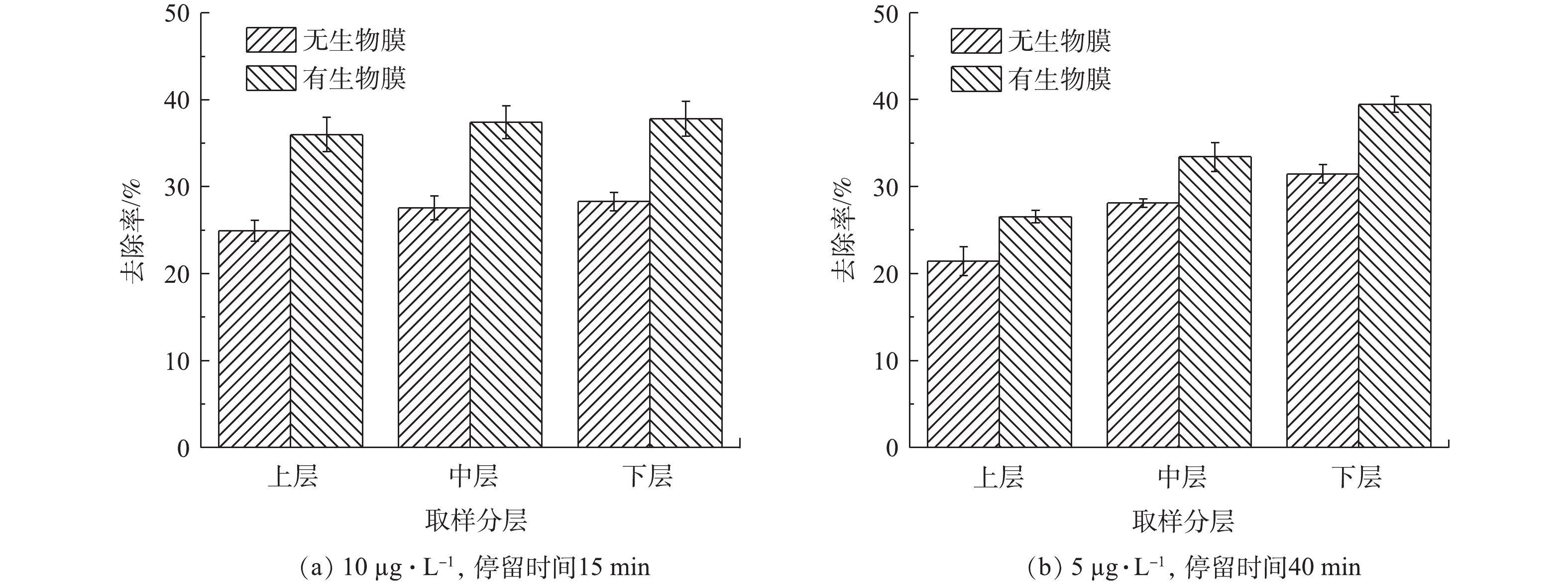
图 3 不同起始浓度下双酚A的去除率
Figure 3. Removal rate of bisphenol A at different initial concentrations

图 4 基于Unweighted Unifrac距离的UPGMA聚类树
Figure 4. UPGMA cluster tree based on Unweighted Unifrac distance matrix
-
[1] JOHNSON A C, KELLER V, DUMONT E, et al. Assessing the concentration and risks of toxicity from the antibiotics ciprofloxacin, sulfamethoxazole, trimethoprim and erythromycin in european rivers[J]. Science of the Total Environment, 2015, 511: 747-755. doi: 10.1016/j.scitotenv.2014.12.055 [2] WANG H, HU C, LIU L, et al. Interaction of ciprofloxacin chlorination products with bacteria in drinking water distribution systems[J]. Journal of Hazardous Materials, 2017, 339: 174-181. doi: 10.1016/j.jhazmat.2017.06.033 [3] YE Z, WEINBERG H S, MEYER M T. Trace analysis of trimethoprim and sulfonamide, macrolide, quinolone, and tetracycline antibiotics in chlorinated drinking water using liquid chromatography electrospray tandem mass spectrometry[J]. Chemosphere, 2007, 79(3): 1135-1144. [4] JIA S, SHI P, HU Q, et al. Bacterial community shift drives antibiotic resistance promotion during drinking water chlorination[J]. Environmental Science & Technology, 2015, 49(20): 12271-12279. [5] GUO Y, WANG H, WANG B, et al. Prediction of micropollutant abatement during homogeneous catalytic ozonation by a chemical kinetic model[J]. Water Research, 2018, 142: 383-395. doi: 10.1016/j.watres.2018.06.019 [6] IKE I A, LEE Y, HUR J. Impacts of advanced oxidation processes on disinfection byproducts from dissolved organic matter upon post-chlor(am)ination: A critical review[J]. Chemical Engineering Journal, 2019, 375: 121929. doi: 10.1016/j.cej.2019.121929 [7] FAN C, HORNG C Y, LI S J. Structural characterization of natural organic matter and its impact on methomyl removal efficiency in Fenton process[J]. Chemosphere, 2013, 93: 178-183. doi: 10.1016/j.chemosphere.2013.05.027 [8] WANG W, ZHANG X, WU Q, et al. Degradation of natural organic matter by UV/chlorine oxidation: Molecular decomposition, formation of oxidation byproducts and cytotoxicity[J]. Water Research, 2017, 124: 251-258. doi: 10.1016/j.watres.2017.07.029 [9] ZHAO L, CHEN Y, LIU Y, et al. Enhanced degradation of chloramphenicol at alkaline conditions by S(II) assisted heterogeneous Fenton-like reactions using pyrite[J]. Chemosphere, 2017, 188: 557-566. doi: 10.1016/j.chemosphere.2017.09.019 [10] ZHANG Y, CHEN Z, ZHOU L, et al. Heterogeneous Fenton degradation of bisphenol A using Fe3O4@β-CD/rGO composite: Synergistic effect, principle and way of degradation[J]. Environmental Pollution, 2019, 244: 93-101. doi: 10.1016/j.envpol.2018.10.028 [11] HOU X J, HUANG X P, JIA F, et al. Hydroxylamine promoted goethite surface Fenton degradation of organic pollutants[J]. Environmental Science & Technology, 2017, 51(9): 5118-5126. [12] XIE Z R, WANG C X, YIN L W. Nickel-assisted iron oxide catalysts for the enhanced degradation of refractory DDT in heterogeneous Fenton-like system[J]. Journal of Catalysis, 2017, 353: 11-18. doi: 10.1016/j.jcat.2017.03.024 [13] 吕来, 胡春. 多相芬顿催化水处理技术与原理[J]. 化学进展, 2017, 29(9): 981-999. doi: 10.7536/PC170552 [14] YAO J Y, QU R J, WANG X H, et al. Visible light and fulvic acid assisted generation of Mn(III) to oxidize bisphenol A: The effect of tetrabromobisphenol A[J]. Water Research, 2020, 169: 115273. doi: 10.1016/j.watres.2019.115273 [15] TAGHIZADEH T, TA0LEBIAN-KIAKALAIEH A, JAHANDAR H, et al. Biodegradation of bisphenol A by the immobilized laccase on some synthesized and modified forms of zeolite Y[J]. Journal of Hazardous Materials, 2020, 386: 121950. doi: 10.1016/j.jhazmat.2019.121950 [16] LYU L, ZHANG L L, WANG Q Y, et al. Enhanced fenton catalytic efficiency of γ-Cu-Al2O3 by σ-Cu2+-ligand complexes from aromatic pollutant degradation[J]. Environmental Science & Technology, 2015, 49: 8639-8647. [17] 刘小琳, 刘文君, 金丽燕, 等. 北京市给水管网管壁微生物膜群落[J]. 自然科学版, 2008, 48(9): 78-81. [18] 鲁智礼, 张堯, 黄俊亮, 等. 多相芬顿-活性炭工艺强化饮用水消毒效果[J]. 环境工程学报, 2019, 13(4): 792-799. doi: 10.12030/j.cjee.201812123 [19] MA X, VIKRAM A, CASSON L, et al. Centralized drinking water treatment operations shape bacterial and fungal community structure[J]. Environmental Science & Technology, 2017, 51(13): 7648-7657. [20] WANG H B, HU X X, HU C. Effects of chlorine and pipe material on biofilm development and structure in a reclaimed water distribution system[J]. Water Science and Technology: Water Supply, 2012, 12(3): 362-371. doi: 10.2166/ws.2012.005 [21] XING X C, WANG H B, HU C, et al. Effects of phosphate-enhanced ozone/biofiltration on formation of disinfection byproducts and occurrence of opportunistic pathogens in drinking water distribution systems[J]. Water Research, 2018, 139: 168-176. doi: 10.1016/j.watres.2018.03.073 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4487
- HTML全文浏览数: 4487
- PDF下载数: 49
- 施引文献: 0
