

 DownLoad:
DownLoad:





-
随着水质排放标准的提高,如何使脱氮更加经济有效受到社会各界的广泛关注。在众多方法中,生物硝化反硝化法是目前最理想的脱氮方法[1]。生物脱氮一般采用异养反硝化法。该方法通常存在碳源不足,须外加有机碳源的问题,运行成本高,且处理后存在二次污染风险[2-4]。相较于异养反硝化过程,自养反硝化法以硫自养反硝化研究居多,利用单质硫提供电子,在缺氧条件下达到氮还原的目的[5-7]。其主要具有以下优势:1)无须外加有机碳源,无二次污染问题;2)剩余污泥产量低,污泥处理成本显著降低[8-9];3)硫自养反硝化过程中N2O等温室气体产量低[10]。此外,硫自养反硝化也存在2点技术缺陷:一方面,硫自养反硝化过程中会产生H+,导致系统pH降低,须补充碱度以维持硫自养反硝化细菌生长最适pH[11];另一方面,产生的硫酸盐是须控制的环境污染物之一[12-13]。
近年来,有研究者针对2种反硝化存在的问题,在异养与硫自养反硝化的基础上,结合两者的优缺点,实现协同脱氮的目的[14-16]。李祥等[17]将单质硫投加至异养反硝化反应器中进行协同作用,污泥产量缩减一半,且无须外加碱度。陈川[18]利用自养/异养菌的协同作用进行反硝化脱硫,以单质硫为终产物,实现资源的回用。张超[19]向硫自养反硝化系统中适当投加碳源,研究了甲醇、乙醇2种碳源对硫自养/异养协同反硝化去除效果及最终产物的影响,达到了提高脱氮能力和抑制过多硫酸盐生成的效果。
硫自养反硝化效果在碳源投加后得到强化,但关于强化后的反硝化系统中自养和异养反硝化贡献度的研究相对较少。本研究在连续稳定运行的硫自养反硝化反应器内,投加少量碳源,实现硫自养/异养协同反硝化过程;通过分析投加碳源后的脱氮效果、碱度、硫酸盐和COD的变化,进一步探讨碳源强化硫自养过程脱氮效果,并对协同反硝化作用机制进行分析讨论,以期为更高效的脱氮处理提供参考。
全文HTML
-
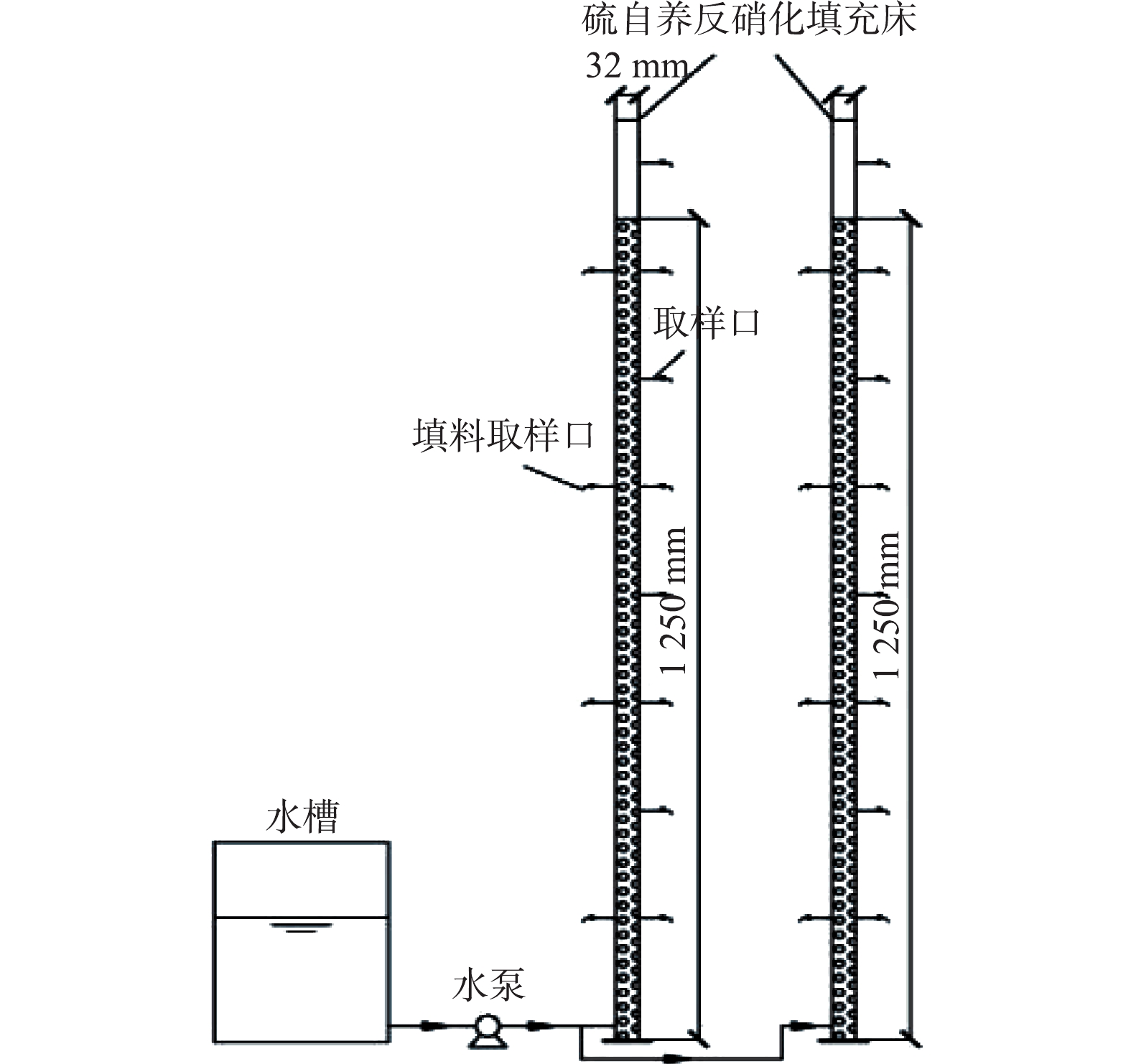
实验采用单质硫填充床反应器,实验装置如图1所示。反应器内径32 mm,高度1 400 mm,填料填充高度1 250 mm,有效体积1 L,进水口距反应器底部20 mm,沿程设置8个取样口用于获取沿程水样,另在沿程方向平均设置4个填料取样口。
实验共设置2组行反应器,分别用于硫自养反硝化和协同反硝化效能对比研究。硫自养反硝化反应器与协同反硝化反应器内部均装填直径1~3 mm片状单质硫,进水采用上向流连续进水。
-
实验接种污泥取自深圳市某污水处理厂缺氧段污泥,经硫代硫酸钠驯化富集培养20 d后,污泥浓度MLSS=3 000 mg·L−1,接种至硫自养反应器中,接种量500 mL。
实验进水采用人工配水,碱度投加量对运行效能的影响实验和碳源投加量对运行效能的影响实验中水质情况分别如表1和表2所示。
硫自养反硝化实验通过改变进水碱度和水力停留时间(HRT)来探究协同反硝化的实验条件,协同反硝化实验通过改变乙酸钠投加量以探究碳源投加量对反硝化效能的影响。
-
实验仪器:紫外分光光度计(UV-2600型,岛津);电子天平(BSA2245-CW型,德国赛多利斯);电热鼓风干燥箱(DHG-9145A型,上海一恒科学仪器有限公司);调温电热套(KDM型,山东华鲁电热仪器有限公司)。
实验试剂:无水乙酸钠(CH3COONa),分析纯;酚酞(C20H14O4);甲基橙(C14H14N3NaO3S);98%碳酸氢钠(NaHCO3);铬酸钾(K2CrO4),分析纯;氯化钡(BaCl2·2H2O),分析纯;无水硫酸钠(Na2SO4),优级纯;氨基磺酸(NH2SO3H),分析纯;对氨基苯磺酰胺(C6H8N2O2S),分析纯;N-(1-萘基)-乙二胺二盐酸盐(C10H7NHC2H4NH2·2HCl),分析纯;重铬酸钾(K2Cr2O7),优级纯;硫酸亚铁铵((NH4)2Fe(SO4)2·6H2O),分析纯;硫酸铝钾(KAl(SO4)2·12H2O),分析纯;钼酸铵((NH4)6Mo7O24),分析纯;硫酸银(Ag2SO4),分析纯;七水合硫酸亚铁(FeSO4·7H2O),分析纯;99%无水邻菲罗啉(C12H8N2);硫酸汞(HgSO4),优级纯。
-
采用紫外分光光度法[20]测定
NO−3-N 浓度;采用N-(1-萘基)-乙二胺光度法[20]测定NO−2-N 浓度;采用铬酸钡光度法[20]测定SO2−4 浓度;采用酸碱指示剂滴定法[20]测定碱度;采用快速密闭催化消解法[20]测定CODMn。
1.1. 实验装置
1.2. 实验水质及接种污泥
1.3. 实验仪器和试剂
1.4. 分析方法
-
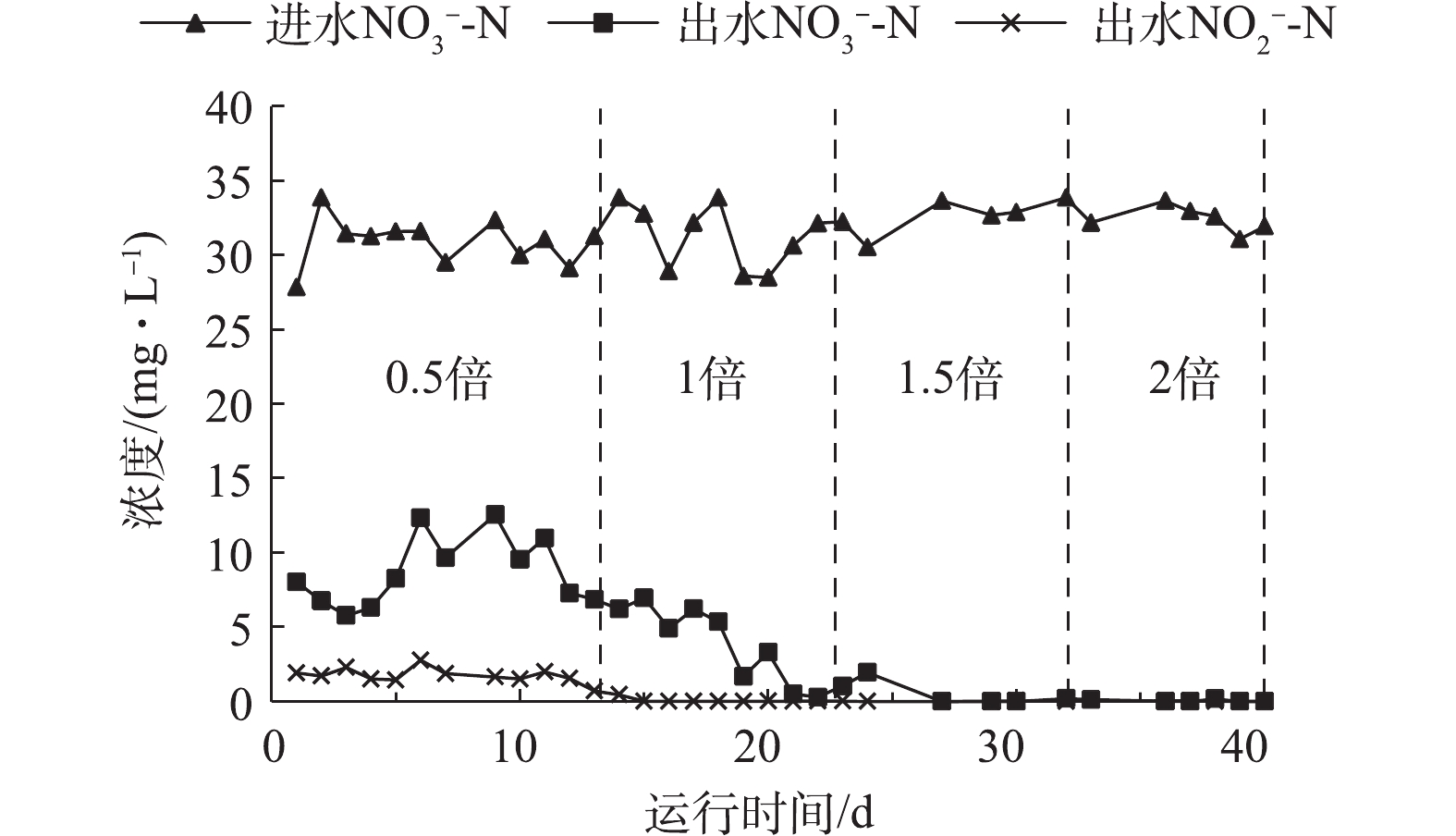
实验初期进行硫自养反硝化培养,进水硝氮浓度30.00 mg·L−1,进水碱度(以CaCO3计)分别设置为理论投加量的0.5、1、1.5和2倍,即实际碱度投加量分别为83.37、167.82、259.42和352.13 mg·L−1,检测进出水中
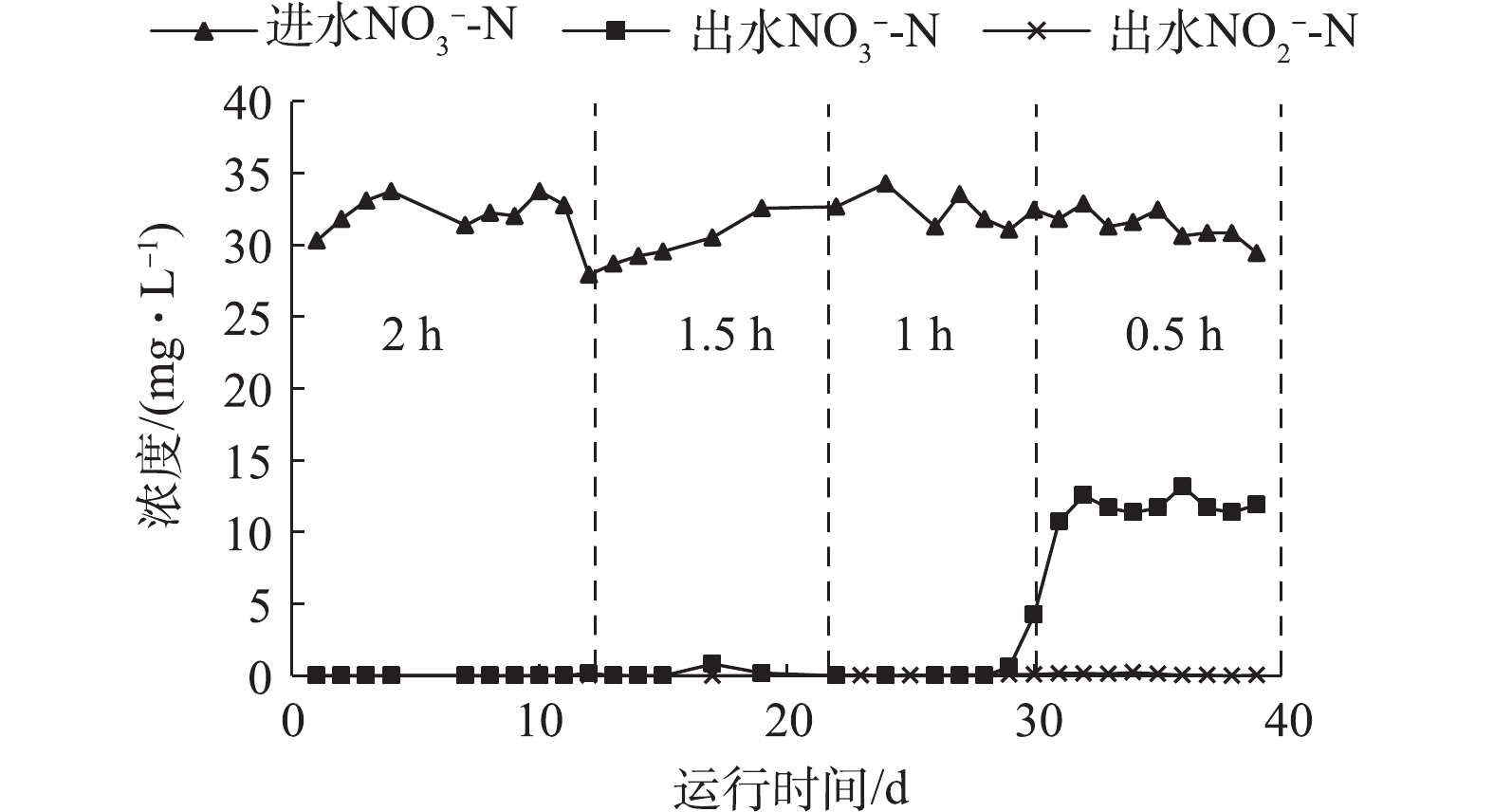
NO−3-N 和NO−2-N 浓度。不同碱度对自养反硝化效能的影响如图2所示。从图2可以看出,协同反硝化最适碱度投加量为理论值1.5倍时即可实现完全反硝化,出水NO−3-N 浓度为0.15 mg·L−1,NO−2-N 浓度小于0.1 mg·L−1。在1.5倍碱度投加量条件下,控制HRT分别为0.5、1、1.5和2 h,测定进出水NO−3-N 和NO−2-N 浓度,结果如图3所示。可以看出,当HRT为0.5 h时,自养反硝化过程正常进行且硝氮有剩余,硝氮去除率为62%,这有利于后续研究硫自养/异养协同反硝化强化脱氮的效能及反应机理。在HRT为0.5 h,1.5倍碱度投加量条件下,当硫自养反硝化反应器稳定运行至出水
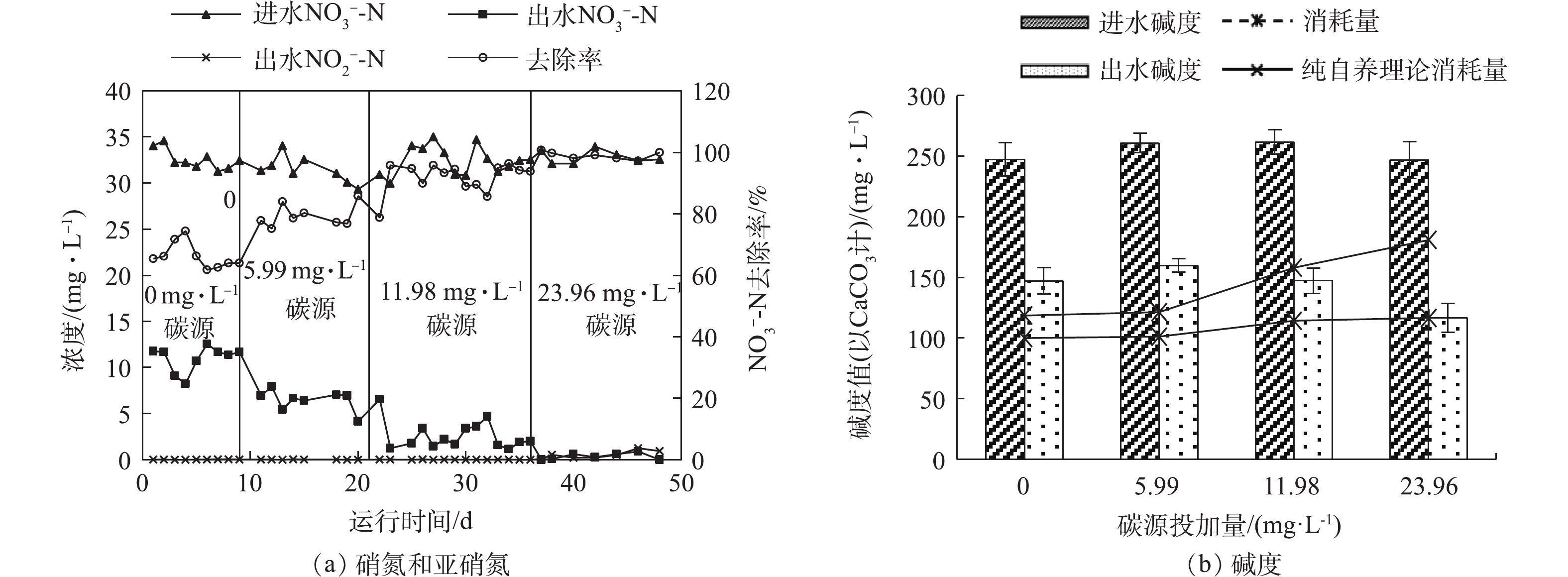
NO−3-N 浓度约为10 mg·L−1时,向反应器内投加碳源,探究了硫自养/异养协同反硝化运行效能。选取完全降解上述自养反硝化剩余10 mg·L−1NO−3-N 所需的0.125、0.25、0.5倍碳源投加量(以乙酸钠计),即实际投加的碳源量分别为5.99、11.98、23.96 mg·L−1,检测了协同反硝化条件下进出水NO−3-N 、NO−2-N 浓度以及碱度变化,结果如图4所示。从图4(a)可以看出,在自养反硝化阶段,出水
NO−3-N 浓度为10.98 mg·L−1,去除率为66.3%;在5.99、11.98、23.96 mg·L−1碳源期各阶段出水NO−3-N 浓度分别为6.46、2.32、0.35 mg·L−1,去除率分别为79.4%、92.9%、99.0%。分析出水NO−2-N 浓度变化发现,在碳源投加量不超过理论值的0.25倍时,出水NO−2-N 低于0.02 mg·L−1;当投加量增加到理论值的0.5倍时,出水NO−2-N 浓度升高至0.65 mg·L−1。有研究[12]发现,在反硝化过程中,反应分2步进行:首先,NO−3-N 转化为NO−2-N ;然后,NO−2-N 转化为N2。在自养反硝化过程中,NO−3-N 转化为NO−2-N 的反应速率小于NO−2-N 转化为N2的反应速率,而异养反硝化过程恰好相反。因此,硫自养反硝化过程中投加碳源可增强异养反硝化效果,从而显著提高反硝化速率。在不同碳源投加量下,耗碱量的对比结果如图4(b)所示。根据运行期间进出水碱度和硝氮去除量,可以得到协同反硝化的实际耗碱量以及同等去除量条件下自养反硝化的耗碱量。5.99 mg·L−1碳源投加量时的碱度消耗量较自养反硝化阶段无明显变化。自养反硝化阶段耗碱量比理论值减少18 mg·L−1,而5.99 mg·L−1碳源投加量条件下,耗碱量只比理论值减少20.19 mg·L−1。SAHINKAYA等[12]通过批次实验证实了在碳源量投加量较低时,会出现碳源被完全利用却无任何异养反硝化过程发生的现象,这可能是兼性自养/异养反硝化菌作用的结果,说明当碳源投加量仅为5.99 mg·L−1时,异养反硝化并无明显贡献。而11.98 mg·L−1和23.96 mg·L−1碳源投加量条件下,耗碱量变化较明显,分别比理论值减少了43.87 mg·L−1和64.31 mg·L−1。理论上,自养条件去除1 g
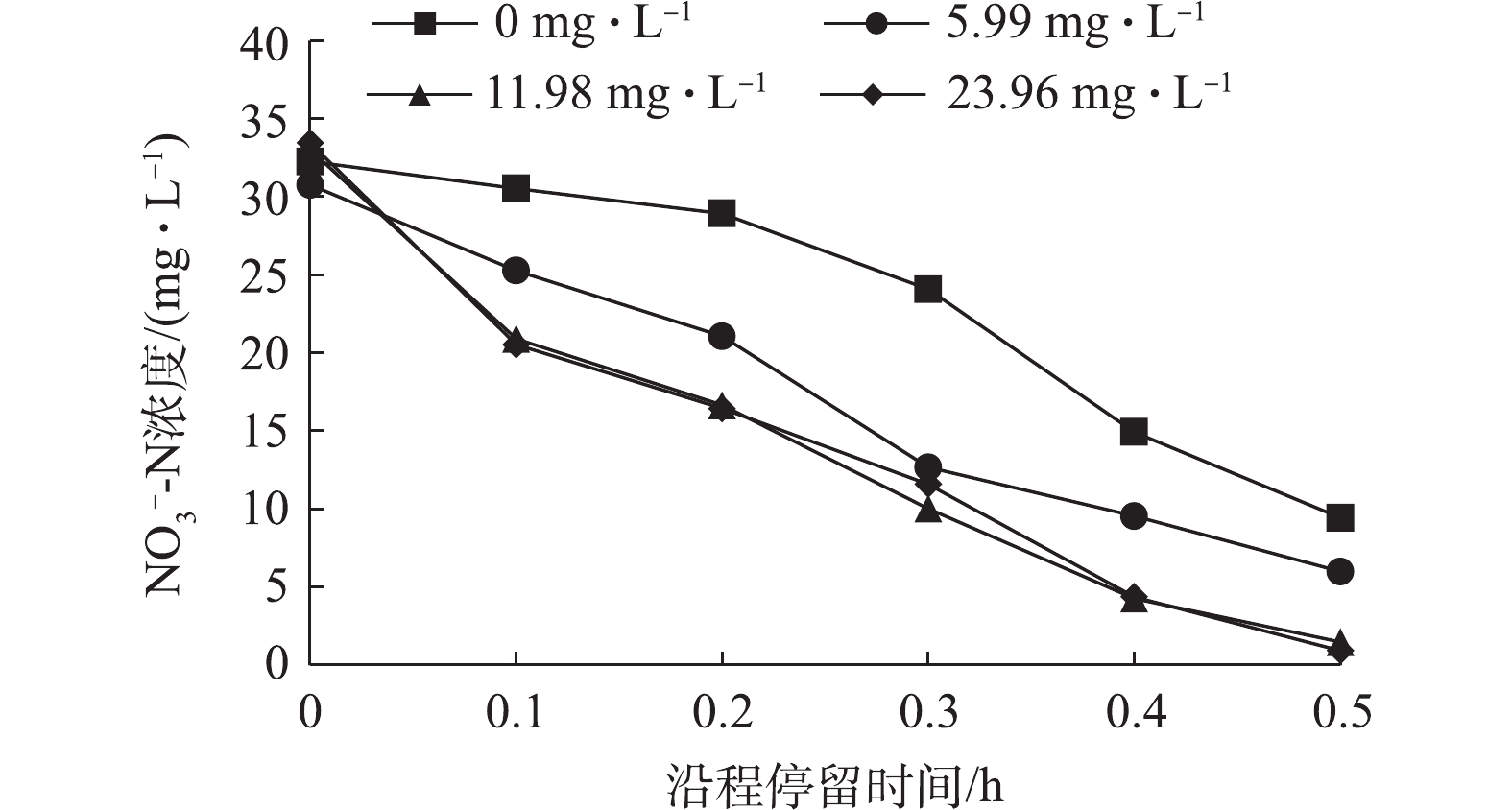
NO−3-N 将会消耗4.57 g CaCO3 碱度,而异养条件下每去除1 gNO−3-N 将会产生3.57 g CaCO3 碱度。在11.98 mg·L−1和23.96 mg·L−1碳源投加量条件下,碱度理论消耗量与实际消耗量的差值随投加量增加而增加,这说明反应中出现了明显的异养反硝化现象。在一个完整水力停留时间期间内,测定不同碳源投加量下沿程出水
NO−3-N 浓度变化,结果如图5所示。由图5可以看出,自养反硝化过程中前半段反硝化速率较低,反应主要在中后段。随着碳源投加量的增加,前半段反硝化速率明显提高,5.99 mg·L−1碳源期整个沿程过程中反应速率均匀;11.98 mg·L−1和23.96 mg·L−1碳源期前半段NO−3-N 的降解速率明显提高,这表明向硫自养反硝化系统中投加适量碳源可显著提高反硝化效率,且随碳源投加量的增加,反硝化速率逐渐增强。 -
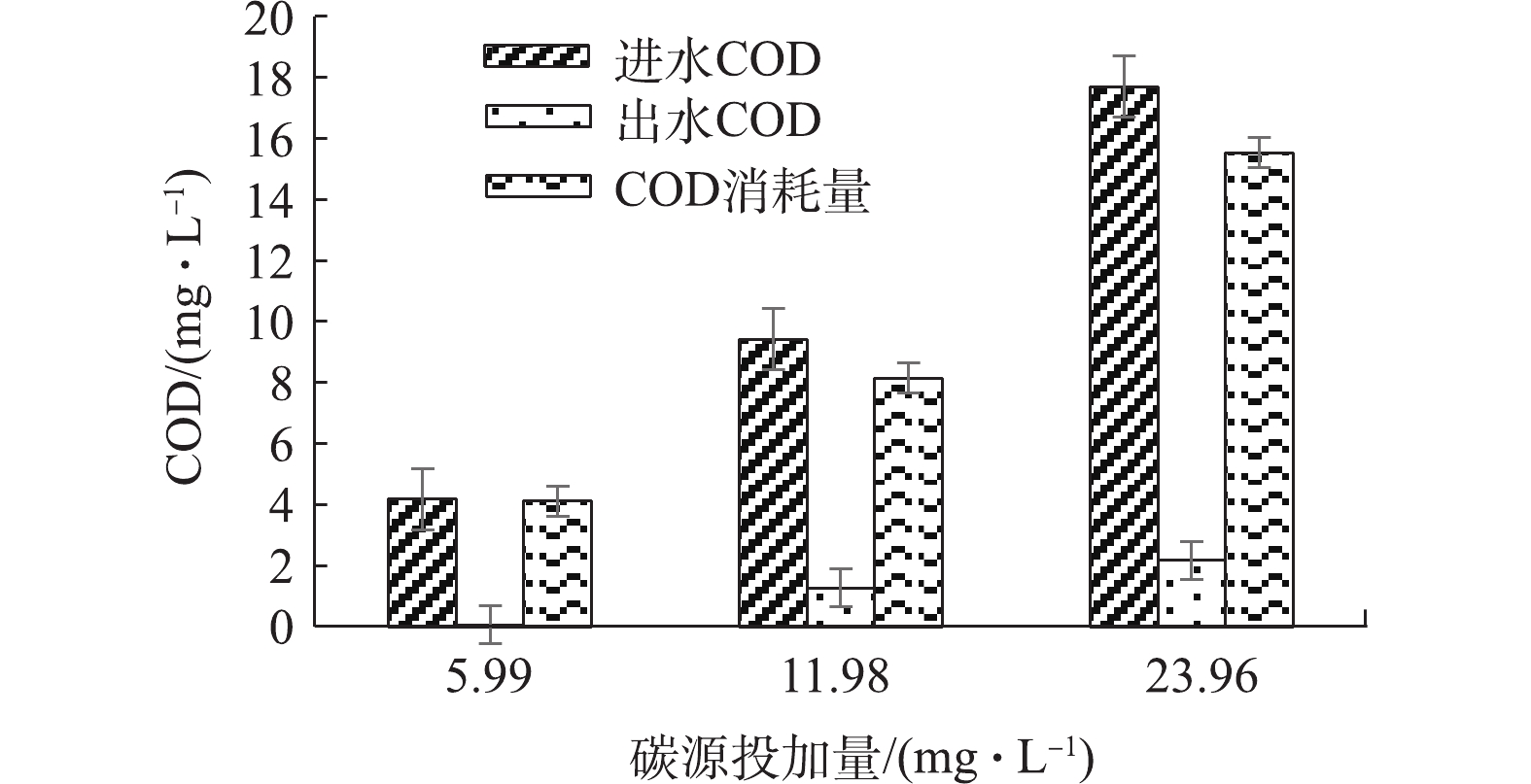
1)协同条件下进出水COD对比分析。为探究不同碳源投加量条件下,协同反硝化过程中COD消耗量同脱氮效率之间的关系。分别对5.99、11.98和23.96 mg·L−1碳源投加量条件下的进出水COD进行了检测,结果如图6所示。计算得到3种条件下COD消耗量分别为4.11、8.15、15.54 mg·L−1,相对应的异养反硝化硝氮去除量分别为1.12、2.18、4.16 mg·L−1。分析各阶段COD利用率发现,除5.99 mg·L−1碳源投量条件下COD被完全消耗,11.98 mg·L−1和23.96 mg·L−1碳源投加量条件下COD均有剩余,利用率分别为86.5%和87.7%,COD利用率有所降低,这表明自氧反硝化作用或者微生物可能影响COD的利用。
-
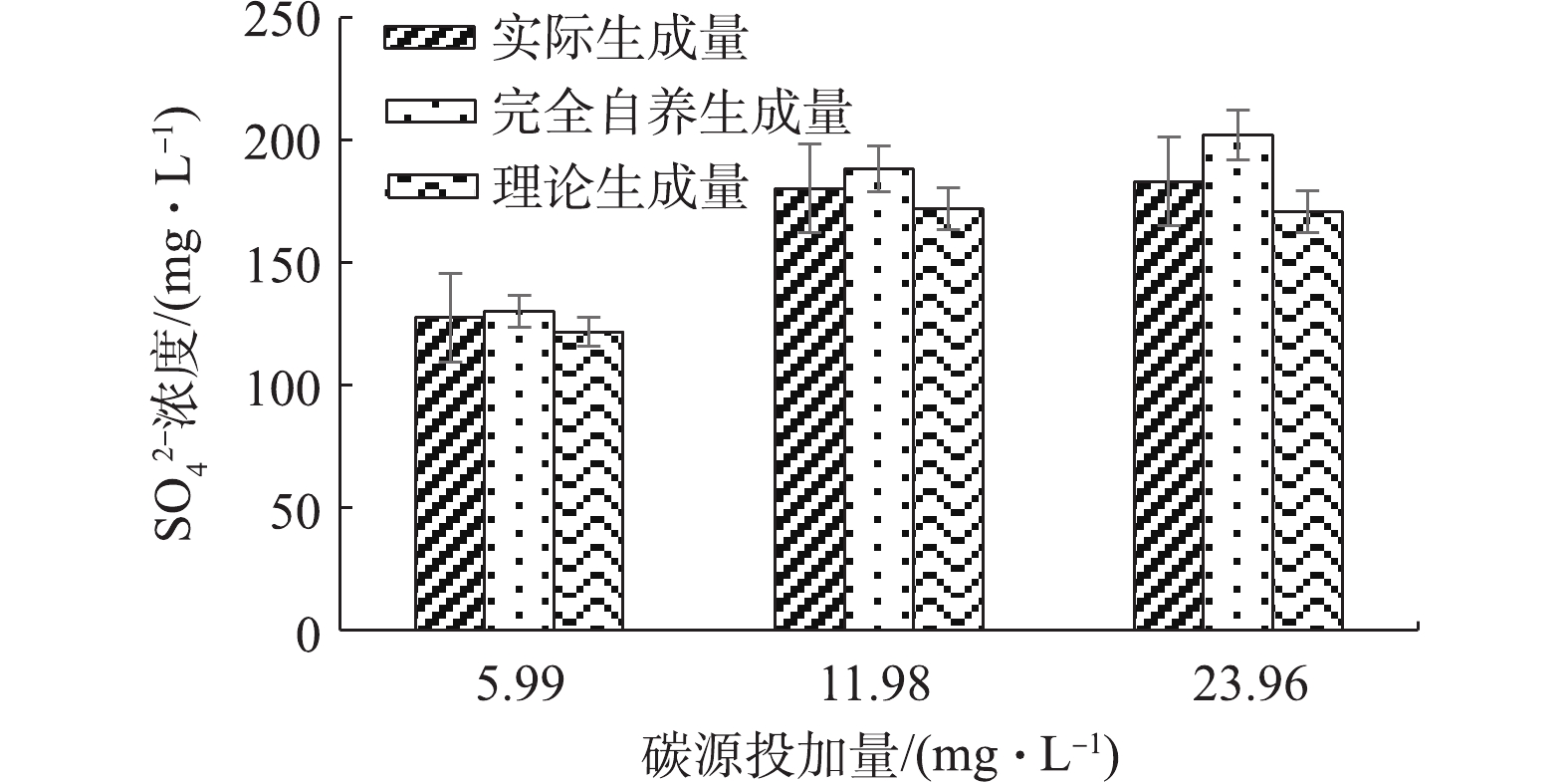
2)协同条件下进出水硫酸盐影响。为确定协同条件下自养部分实际的硝氮量去除,分别测定了不同反应条件下出水硫酸盐浓度,结果如图7所示。在3种碳源投加量条件下的硫酸盐生成量分别为127.55、180.27和183.18 mg·L−1。根据实际出水硝氮值,完全自养反硝化应生成的硫酸盐的量分别为130.15、185.87和202.11 mg·L−1。根据COD消耗量对应的硝氮去除量值可确定随碳源投加而减少的硫酸盐生成量,分别为8.41、16.44和31.34 mg·L−1。因此,协同条件下的硫酸盐实际生成量同理论值相比,分别增加了5.81、10.84和12.40 mg·L−1;另一方面,协同过程中硫酸盐实际生成量和纯自养理论生成量并无明显差异,这说明协同条件下与自养条件下的硫酸盐生成量基本一致。通过COD消耗量计算得出的协同条件下的硫酸盐理论生成量亦比实际生成量少。COD被消耗表明有异养反硝化反应发生,而硫酸盐生成量则反映自养反硝化程度。由于协同反硝化过程中异养反硝化的存在,会一定程度上减少硫酸盐的生成量,而实验的结果则显示,硫酸盐生成量随着碳源消耗无明显减少。
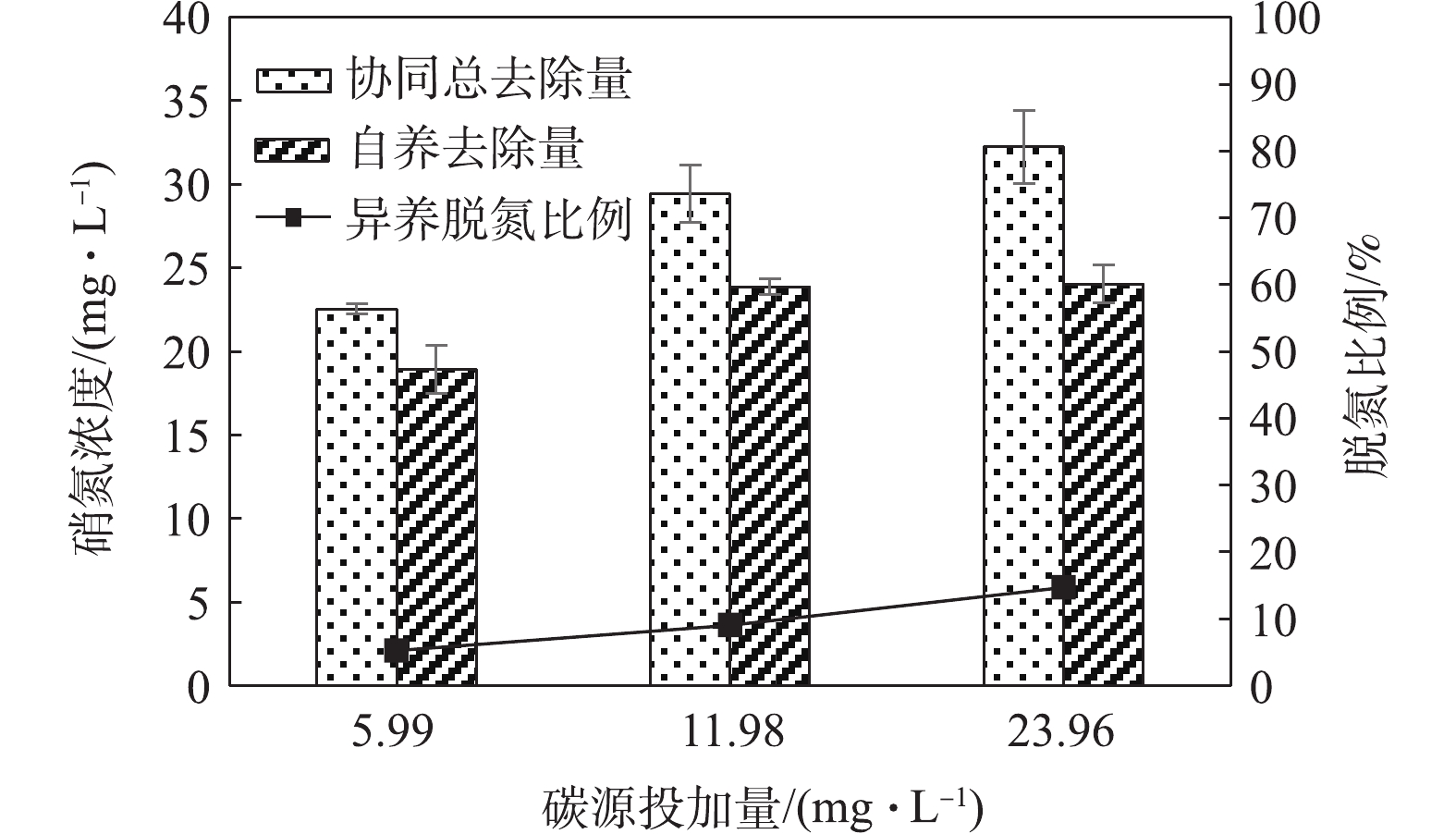
对根据硫酸盐生成量计算得出的硝氮去除量进行分析。图8反映了根据出水
SO2−4 含量所得的协同条件下的自养去除量和异养脱氮比例,以及根据COD消耗量得到的自养脱氮比例。其中,异养脱氮比例(rh)与自养脱氮比例(ra)的计算方法如式(1)和式(2)所示。式中:S为实际硫酸根生成量,mg·L−1; N为实际硝氮去除量,mg·L−1; SN为去除1 mg·L−1硝氮的
SO2−4 生成量,mg·L−1; R为COD消耗量,mg·L−1; RN为去除1 mg·L−1硝氮的COD生成量,mg·L−1。自养和异养脱氮比例呈现良好的负相关关系,3种碳源投加量条件下的异养脱氮比例分别为5.3%、9.1%和15.5%,自养脱氮比例为95.1%,92.6%和87.1%。随着碳源投加量的增加,碳源被用于异养反硝化的比重也在不断增加。3种条件下自养与异养脱氮比例加和后的总和分别为100.3%、101.7%和102.6%,表明随着碳源投加量的增加,自养与异养的去除效果有叠加效应。
-
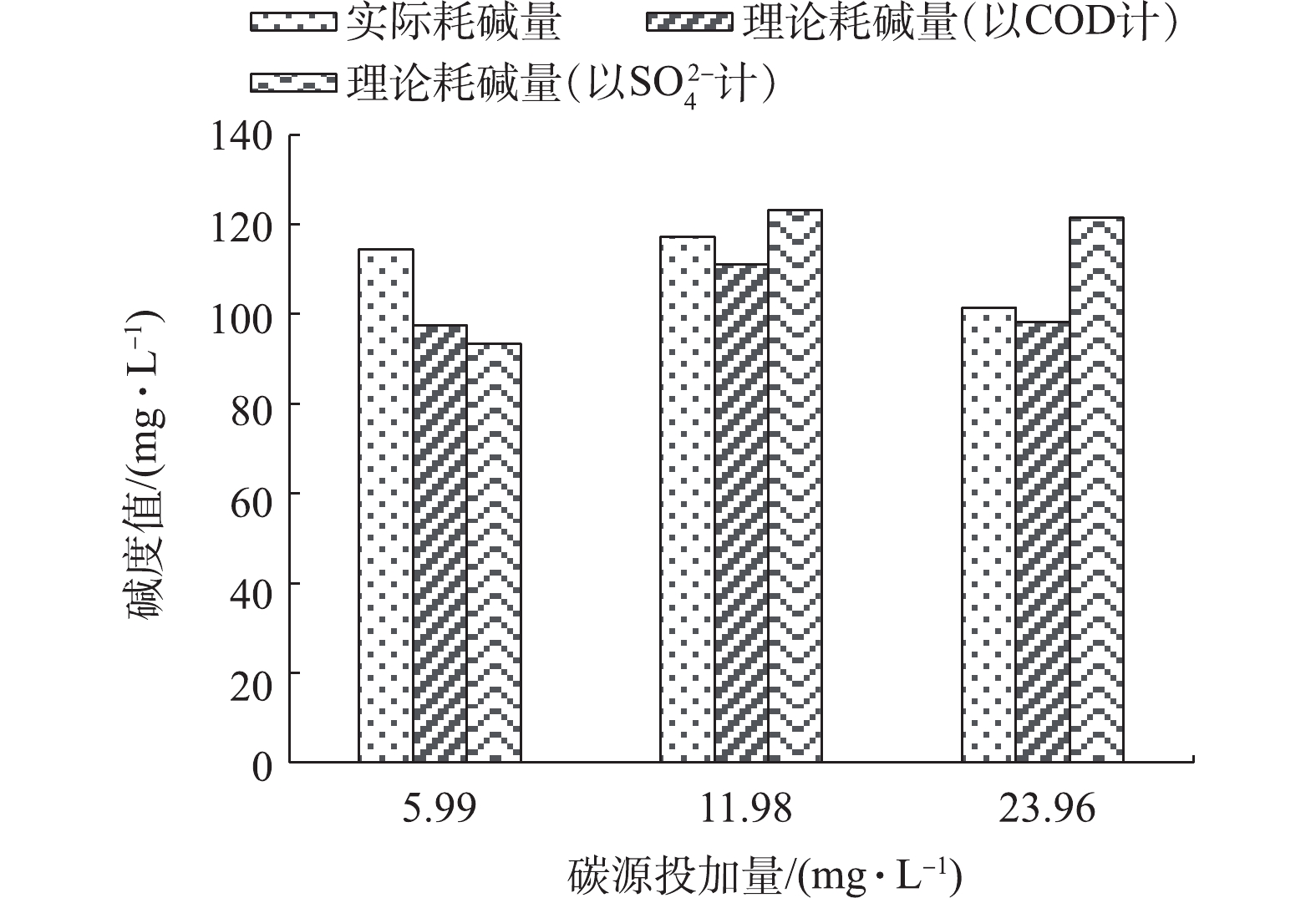
3)协同条件下碱度分析。为进一步探究协同反硝化过程异养反硝化的作用,考察了硝氮去除量与碱度消耗量之间的关系。对协同反硝化条件下耗碱量进行分析,排除自养本身耗碱量与理论值的偏差,投加碳源之后均在一定程度上减少了碱度消耗量,如图9所示。图9中的实际耗碱量表示由实际硝氮去除量得到相同去除量在完全自养条件下需要的碱度,理论耗碱量(以COD计)表示从COD消耗量推算理论上异养反硝化产生碱度的量,与自养反硝化叠加即可得到1.5倍碱度投加量下协同反硝化过程的碱度消耗值,理论耗碱量(以
SO2−4 计)表示从硫酸盐生成量推算消耗的理论碱度。3种碳源投加量条件下的实际耗碱量分别为114.40、117.26、101.49 mg·L−1。在5.99 mg·L−1碳源投加量条件下,根据COD消耗量推算得出的耗碱量为93.35 mg·L−1,根据硫酸盐生成量得到的耗碱量为97.43 mg·L−1,均小于实际耗碱量。结果表明,虽然反应过程中COD有被利用,但实际并无明显的碱度产生,因而耗碱量大于理论值。而实际耗碱量和理论耗碱量(以SO2−4 计)的差值则表明在碱度被消耗的同时,没有产生硫酸盐,说明存在兼性反硝化细菌导致有机碳源在被利用的同时消耗了碱度[8]。在11.98 mg·L−1碳源投加量条件下,根据COD消耗量得出的耗碱量为111.06 mg·L−1,小于实际耗碱量,而根据硫酸盐生成量得到的耗碱量为123.22 mg·L−1,大于实际耗碱量,即在COD被利用的同时又消耗了部分碱度,自养与异养现象同时存在,结合硫酸盐的生成情况,这一阶段投加碳源显著增强了自养反硝化的效果。实际耗碱量值介于2种理论值之间,表明虽然有异养现象存在,但主要是由于兼性反硝化菌的存在,却并没有真正的异养反硝化反应发生。此外,23.96 mg·L−1碳源投加量条件下存在相同现象,根据COD消耗量得出的耗碱量为98.31 mg·L−1,而根据硫酸盐生成量得到的耗碱量为121.49 mg·L−1,结果表明仍无明显的异养反硝化反应发生,但理论耗碱量(以COD计)与实际耗碱量减小,说明随之碳源投加量增加异养作用有增强的趋势。
2.1. 硫自养/异养协同反硝化运行效能
2.2. 协同反硝化反应机理分析
-
1)当分别向自养反硝化系统中投加5.99、11.98和23.96 mg·L−1碳源时,相比于自养过程,协同反硝化过程分别增加了12.83、15.10、15.58 mg·L−1硝氮的去除量,表明投加少量碳源对反硝化过程有明显的强化作用。
2)在协同反硝化过程中,随着碳源的消耗,在3种碳源投加量条件下,均有超过85% COD被利用,但
SO2−4 生成量并未减少。其原因是,较少碳源投加时,虽有异养反硝化现象发生,但脱氮效果的提高主要源于自养反硝化效率的提高。3)当碳源投加量为5.99 mg·L−1时,实际耗碱量大于根据硫酸根和COD得到的理论耗碱量;而11.98 mg·L−1和23.96 mg·L−1碳源投加条件下,实际耗碱量则均处于2种理论值之间。这说明,实验过程中无单纯异养反硝化发生,但增加碳源投加量会增加异养反硝化的趋势。
