


 下载:
下载:








-
好氧颗粒污泥沉降性能好、处理效率高、生物量高、抗冲击负荷能力强,可实现同步脱氮除磷[1-2],其具有显著的节省占地、节约能源和降低化学药剂使用量等优点,是一项应用价值很高的污水处理技术[3]。除磷颗粒污泥是在厌氧好氧交替环境下,利用生长速率较慢的除磷功能菌群(聚磷菌和聚糖菌)培养的一种特殊的好氧的颗粒污泥,近年来由于其较高的稳定性受到越来越多的关注。除磷功能菌群偏好挥发性脂肪酸(VFA),因此,现有研究常常以乙酸钠、丙酸钠等挥发性脂肪酸盐为碳源[4],在短期内便可形成颗粒污泥[5]。然而,在实际污水中,VFA含量非常有限,大分子有机物(胶体态、颗粒态)才是碳源的主体,全球实际污水中大分子有机物约占总好氧性有机物(以COD计)的50%~90%[6]。尽管大分子有机物有一定的絮凝作用[2,7],但其缓慢的扩散、水解发酵过程,可能通过影响微生物的代谢过程而抑制除磷功能菌群的生长,进而减缓甚至阻碍除磷污泥的颗粒化[8]。但目前对于大分子有机物在除磷颗粒污泥系统的降解研究较少,关于在大分子有机物作用下除磷污泥的颗粒化的机理尚不清楚。
胞外聚合物(EPS)是微生物在一定条件下,由自身代谢产生并黏附于细胞壁外的一种高分子聚合物[9-10],由多糖(PS)、蛋白质(PN)、DNA、脂类、腐殖酸、氨基酸等和一些无机成分共同组成[11],其中PS和PN约占总量的70%~80%[12]。胞外聚合物是微生物聚集体的重要组成部分,在颗粒污泥的形成和结构稳定中有汲取营养、信号传递、吸附架桥等重要作用[13-15]。胞外聚合物假说是颗粒污泥形成机理的重要假说之一[16-17]。在为数不多的关于大分子有机物对除磷颗粒污泥影响的研究中,主要考察系统的运行效果(除碳、脱氮、除磷)和污泥的颗粒化进程,而对胞外聚合物的关注较少。WAGNER等[18]指出,以胰蛋白胨和颗粒态淀粉等大分子有机物为碳源时,延长厌氧时间可以提高系统的除磷效率,有利于除磷颗粒污泥的形成。LAYER等[19]发现,当进水中含有大量大分子有机物时,将进水后的厌氧静置改为厌氧搅拌,可以提高系统的脱氮除磷效率,改善污泥的沉降性能并促进污泥颗粒化。
EPS的产生及组分与基质种类密切相关[20]。李冬等[21]发现当分别以葡萄糖和生活污水为碳源时,PS在葡萄糖系统的含量较高,而PN在生活污水系统中的含量较高。HE等[22]指出,LB-EPS在乙酸钠系统的含量最高,在淀粉系统最少[23]。但是,WANG等[24]发现,以葡萄糖为基质的EPS中多糖和蛋白的含量均少于以淀粉为基质时,淀粉的水解过程会促进EPS中PN的分泌。张杰等[25]指出,向丙酸钠为主要碳源的进水中投加30%的淀粉,可以刺激微生物分泌淀粉酶,增加EPS中蛋白质的含量,有利于颗粒污泥的形成。然而,这些研究多关注多糖和蛋白质含量等传统指标的变化,且大都基于活性污泥系统,对于颗粒污泥系统中EPS的变化,以及何种组分是影响污泥颗粒化的主要因素尚未有定论。与此同时,现有研究对EPS在污泥表面的分布情况、蛋白质和多糖种类、带电性等特征关注较少。
本研究拟分别以100% NaAc、60% NaAc+40%胰蛋白胨和60%可溶性淀粉+40%胰蛋白胨为碳源,采用扫描电镜、三维荧光光谱分析、Zeta电位等检测方法考察大分子有机物作用下除磷颗粒污泥中EPS的组分差异,并结合污泥性状及颗粒化进程,探索大分子有机物作用下EPS对除磷污泥颗粒化的影响。
HTML
-
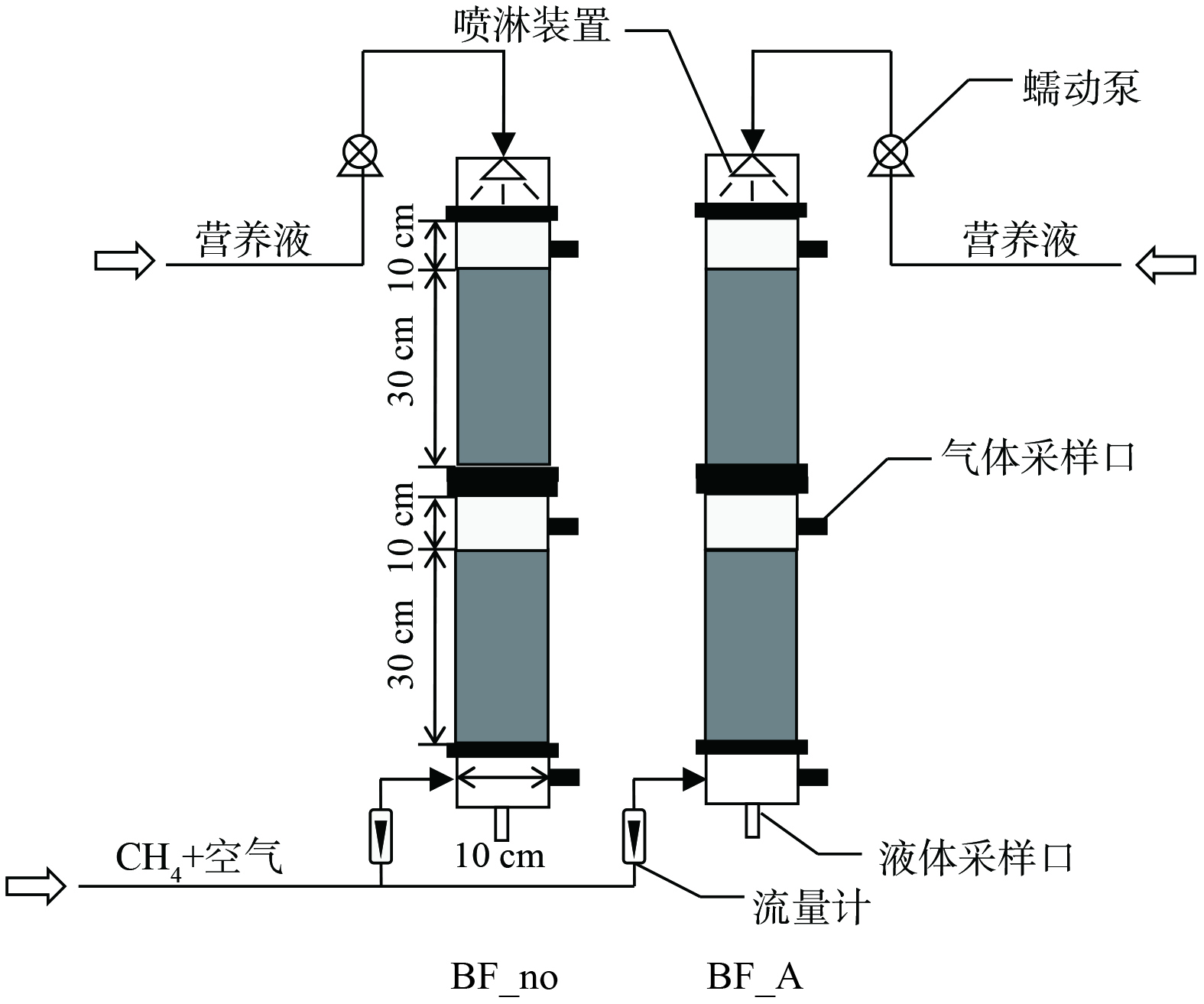
本实验采用3组构型和运行方式相同的序批式反应器(图1),反应器有效水深90 cm,内径7.5 cm,H/D=12,有效容积4 L。反应器底部设有微孔砂芯曝气头,采用电磁式空气压缩机(海利 ACO-318)供气,转子流量计控制曝气量1.2 L·min−1。实验中采用推流式进水的SBR反应器,等工作体积运行,进水从底部由布水盘以活塞流均匀进入,上部已处理好的水被逐渐置换出反应器,由出水口同步自流至出水桶。进水流量由蠕动泵(保定兰格 BT100-2J)控制,进水上升流速0.46 m·h−1,排水比50%。采用恒温水浴(西安东瑞 YHX-030)保持反应器在20 ℃运行。反应器运行周期为4 h,包括同步进出水60 min和静置30 min(进行磷的释放),曝气120 min (DO为6.5~8.5 mg·L−1),沉淀5 min(沉淀时间在30 d内逐渐由30 min减少至5 min,曝气时间对应延长)。整个系统采用时间控制器自动控制。
-
接种污泥为西安市第四污水厂回流污泥,污泥絮体中虽有大量丝状菌,SV5和SV30为99%,但污泥浓度较高(12 000 mg L−1),使得SVI30计算值偏低(82.5 mL·g−1)。采用模拟生活污水作为反应器进水,进水流量为12 L·d−1,进水COD 400 mg·L−1、总氮 40 mg·L−1、总磷 10 mg·L−1,进水成分详见表1。矿物质溶液MgSO4·7H2O 0.4 g·L−1、CaCl2 0.5 g·L−1、NaHCO3 2.5 g·L−1、FeSO4·7H2O 1.0 g·L−1,每天添加120 mL至进水箱;微量元素溶液取KI 60 mg·L−1、H3BO3 300 mg·L−1、MnCl2·4H2O 117 mg·L−1、CoCl2·6H2O 50 mg·L−1、Na2MoO4·2H2O 120 mg·L−1、CuSO4·5H2O 700 mg·L−1、ZnSO4·7H2O 380 mg·L−1,每天添加12 mL至进水箱。
-
常规水质指标监测参考文献中的方法[26]。采用光学显微镜(Olympus BX-53)每周进行1次镜检。污泥扫描电镜(SEM)预处理方法为:在好氧期末取一定量的颗粒污泥,加入磷酸盐缓冲溶液(PBS) 4 ℃下8 000 r·min−1离心清洗3次,每次5 min。加入新配的2.5%戊二醛混匀后置于4 ℃下固定4 h,再加入PBS缓冲液混匀后4 ℃下8 000 r·min−1离心清洗3次,每次离心5 min,依次用30%、50%、70%、85%、95%、100%的乙醇进行梯度脱水(每次加入乙醇后振荡混匀静置15 min,8 000 r·min−1离心5 min,弃上清液)。乙酸异戊酯8 000 r·min−1离心置换2次,每次15 min。样品采用冷冻干燥机(FD-1D-50)在−55 ℃下冷冻干燥12 h。用导电胶将污泥样品固定在样品台上并镀金膜(Pt),进行SEM(Quanta 450FEG)观察。
-
反应器运行期间分别在颗粒化前期(90 d)、颗粒化中期(130 d)、颗粒化成熟期(190 d)各提取3次EPS,EPS提取采用超声-阳离子树脂交换法[27]。采用三维荧光光谱法分别对EPS(LB-EPS和TB-EPS)中物质的种类进行分析。三维荧光光谱的测定采用荧光分光光度计(日本日立 F-7000),激发波长(Ex)为200~480 nm,间隔为5 nm,发射波长(Em)为220~550 nm,间隔为2 nm。激发光和发射光的夹缝均为5 nm,扫描速度12 000 nm·min−1。采用蒽酮比色法测定EPS中多糖类物质,采用Lowry法测定蛋白质类物质[28]。将分离出的LB-EPS和TB-EPS分别注入Zeta电位仪(英国马尔文-ZS90)样品池内测定Zeta电位,同一样品测定5次。
1.1. 反应器
1.2. 接种污泥及模拟污水
1.3. 常规分析方法
1.4. 胞外聚合物(EPS)提取及测定方法
-
对3组反应器中污泥的沉降性能进行连续监测,结果如图2所示。在污泥驯化初期,即20 d之前,各反应器中污泥浓度MLSS迅速下降,污泥流失严重。这主要是由于颗粒污泥培养初期系统选择压较大,导致较多的污泥絮体随出水排出。在20~75 d,污泥逐渐适应新环境,浓度逐渐稳定。由于基质的不同,各反应器的污泥浓度呈现不同的变化规律,R1、R2、R3系统中MLSS分别维持在2 500~4 000、4 000~5 000和5 000~6 500 mg·L−1,污泥浓度依次增高。这主要由于进水中大分子有机物的絮凝和吸附架桥作用,有利于游离细菌和污泥絮体之间的聚集[25]。随着污泥的颗粒化,培养75 d后,系统逐渐稳定,3个反应器内的MLSS基本维持在4 000~5 500 mg L−1(图2(a))。
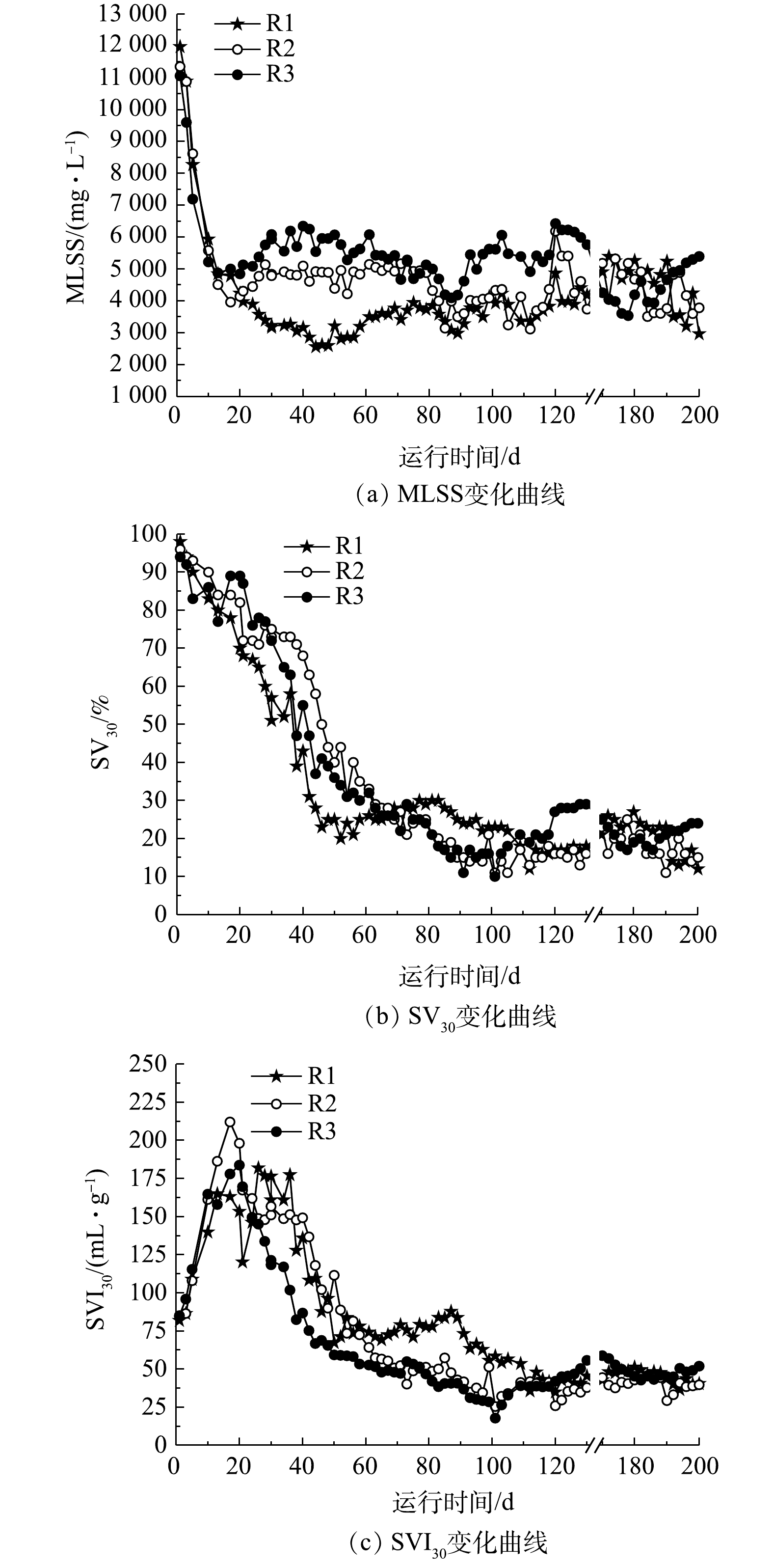
由SV30变化曲线可知:在污泥颗粒化的前60 d,3个反应器的SV30迅速降低,R1(NaAc)反应器SV30下降速度最快,R3(淀粉+胰蛋白胨)次之,R2反应器(NaAc+胰蛋白胨)最慢;在85 d后,各反应器SV30均维持在20%左右,泥水分离效果较好,污泥沉降性能较好(图2(b))。与之相类似,SVI30的变化总体呈现降低趋势,但在驯化初期出现污泥膨胀现象。在接种污泥驯化20 d内,反应器中丝状菌大量增殖,导致污泥膨胀,SVI30数值增高;36 d后,污泥初步完成驯化,丝状菌淘汰消失殆尽,污泥膨胀现象逐渐消失;在110 d后,各反应器中SVI30值均维持在50 mL·g−1左右,污泥沉降性良好(图2(c))。
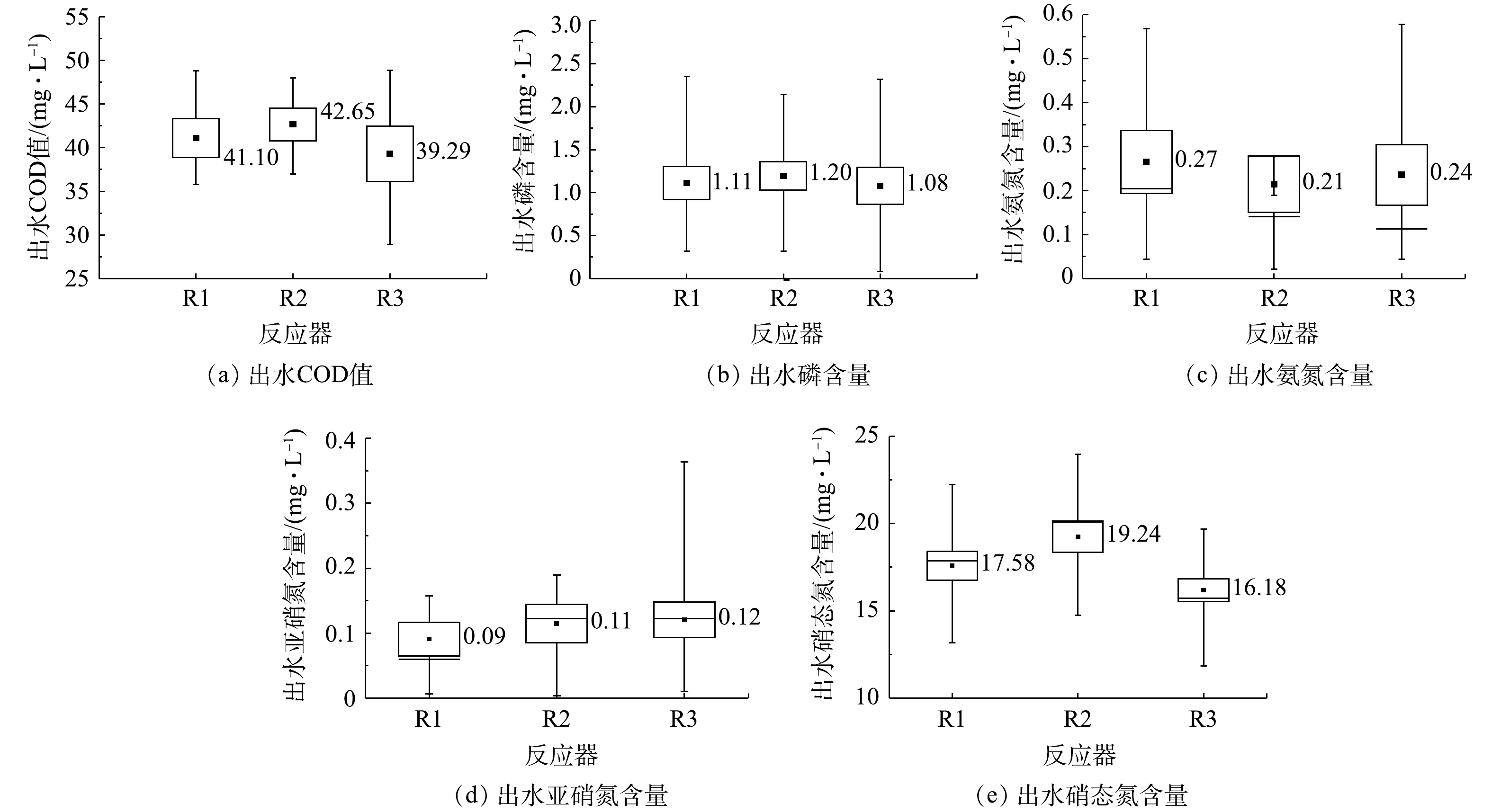
将整个培养阶段所测出水污染物含量汇总并作箱式图,结果如图3所示。3个除磷颗粒污泥系统中出水COD值和磷浓度差异较小,除碳除磷效果均良好,COD去除率分别为89.73%、89.34%和90.18%,磷的去除率分别为88.90%、88.00%和89.20%。出水中氨氮和亚硝氮含量较低,硝态氮较高,系统硝化效果较好,但反硝化效果较差。
-
由图4(a)可以看出,接种污泥为微小絮体,形状不规则,污泥聚集程度和密实度较低。污泥中含有大量的微丝菌、0041型和0092型等丝状菌(图4(b))。在R1(100% NaAc)系统前期,颗粒化速率较慢,除磷颗粒污泥粒径较小(90 d,图4(c));190 d后成形的除磷颗粒污泥形状规则圆润,边界清晰,内部密实,污泥絮体少,颗粒化程度较高,粒径最大,大约为200~500 μm (图4(d));R2(60%NaAc+40%胰蛋白胨)系统最初形成的微生物聚集体形状不规则,表面粗糙(90 d,图4(e));190 d后形成的成熟除磷颗粒污泥表面有丝状菌包裹,内部有单个或多个核心密实点,呈椭圆形,系统中污泥絮体较多,颗粒污泥粒径为200~400 μm(图4(f));R3(60%淀粉+40%胰蛋白胨)系统中颗粒化前期污泥容易聚集,同期相比颗粒污泥粒径较大,丝状菌数量较多(90 d,图4(g));190 d后系统形成的除磷颗粒污泥边界较为粗糙,表面不光滑,污泥内有多个密实点,颗粒污泥内部存在孔道,边界较为模糊,系统中污泥絮体最多,颗粒污泥粒径为200~300 μm (图4(h))。3个系统中污泥絮体含量随着大分子有机物占比的增多而增多,形成的颗粒污泥为白色或黄褐色。

-
不同基质条件下除磷颗粒污泥的表面形貌见图5。R1系统成熟的除磷颗粒污泥表面有大量胞外聚合物,光滑圆润,结构密实,球菌和杆菌由胞外聚合物黏附并固定在颗粒污泥表面。刘燕等[29]研究也发现,乙酸钠为底物时分泌的EPS最多,表面有一层包裹物质。污泥表面EPS的大量存在有助于颗粒污泥对菌胶团以及小絮体的吸附和固定,可减少液相中的污泥絮体。这与普通光学显微镜的观察结果一致。BARR等[30]将除磷颗粒污泥进行冷冻切片后通过荧光原位杂交技术发现,除磷颗粒污泥外部被一层致密且光滑的EPS所包裹,并推测由乙酸盐和丙酸盐诱导而优势生长的聚磷菌,可以通过分泌大量的胞外聚合物促进污泥的颗粒化。R2系统除磷颗粒污泥表面胞外聚合物相比R1系统较少,污泥表面较为粗糙,颗粒污泥以丝状菌为骨架,短杆菌和球菌在丝状菌的框架中聚集。R3系统成熟的颗粒污泥表面胞外聚合物更少,污泥表面疏松、粗糙,球菌、杆菌和丝状菌交错生长,颗粒污泥内部孔道明显,这些孔隙和通道有利于水中营养物质和溶解氧深入除磷颗粒污泥内部,提高大分子有机物的利用效率[31]。可见,大分子有机物导致污泥EPS含量降低,不利于污泥颗粒化。

-
为确定LB-EPS和TB-EPS的主要成分,采用三维荧光光谱对不同系统中提取的LB-EPS和TB-EPS进行分析。根据荧光区域积分法(FRI)把Ex和Em所形成的荧光区域分为5个区域:Ⅰ(Ex/Em=200~250 nm/280~330 nm)、Ⅱ(Ex/Em=200~250 nm/330~380 nm)、Ⅲ(Ex/Em=200~250 nm/380~500 nm)、Ⅳ(Ex/Em=250~280 nm/280~380 nm)、Ⅴ(Ex/Em=250~400 nm/380~500 nm)。其中,Ⅰ和Ⅱ分别代表芳香类蛋白物质Ⅰ和Ⅱ,Ⅲ代表类富里酸物质,Ⅳ代表溶解性微生物代谢产物,Ⅴ代表类腐殖酸物质[32-35]。
如图6所示,进水基质的不同导致3个系统中LB-EPS的组分存在明显差异。R1系统污泥LB-EPS中只含有峰A(芳香类蛋白物质Ⅱ,Ex/Em=225 nm/335 nm)和峰B(色氨酸类蛋白质和其他蛋白质类物质,Ex/Em=280 nm/335 nm),R2中除峰A和B,还出现峰E(类腐殖酸物质,Ex/Em=365 nm/445 nm);R3 系统LB-EPS中同样含有峰A、峰B、峰E(类腐殖酸物质,Ex/Em=345 nm/430 nm),这表明大分子碳源系统中LB-EPS的组成成分更丰富。芳香族蛋白质和色氨酸是好氧颗粒污泥的关键成分,对颗粒污泥的形成和结构稳定有重要作用[35]。李定昌等[36]发现,小粒径好氧颗粒污泥中腐殖酸类物质的荧光强度较高,随着粒径的增大而逐渐减弱。与此类似,程祯等[37]对颗粒污泥形成过程中EPS成分的监测发现,在颗粒污泥成熟之后腐殖酸类物质消失。在本研究中,R2、R3系统中一直伴随着小絮体的存在,未实现完全颗粒化,且颗粒污泥粒径呈现出R1>R2>R3的规律,推断较小的粒径可能是R2,R3系统存在腐殖酸类物质的一个原因。此外,WANG等[6]在研究大分子有机物对活性污泥系统EPS的影响时发现,大分子有机物会使活性污泥产生腐殖酸类代谢产物,这可能是污泥EPS含有腐殖酸类物质的第2个原因。3个系统的TB-EPS中均只出现峰C(酪氨酸类蛋白质,Ex/Em=230 nm/300 nm)和峰D (色氨酸类蛋白质和其他蛋白质类物质,Ex/Em=270 nm/370 nm),可见,碳源类型的差异未对TB-EPS的组分造成明显影响。值得注意的是,3个系统的LB-EPS出现的峰B和TB-EPS中出现的峰D都是色氨酸类蛋白质和其他蛋白质类物质,但其出峰位置不同,表明LB-EPS和TB-EPS中蛋白质类物质并不相同,它们可能在污泥的颗粒化过程中发挥着不同的作用。

-
EPS中PS具有支撑微生物聚合及维持污泥颗粒完整性的功能,可通过粘结、架桥作用形成稳定颗粒化结构的网状骨架,促进污泥颗粒化[38]。TAY等[39]研究表明,TB-EPS中β-多糖可提高微生物间的黏附力,强化好氧颗粒污泥结构的稳定性。也有研究表明,PN具有较高的疏水性和表面电负性,有助于颗粒间黏附絮凝,是污泥颗粒化的主要影响物质[40]。
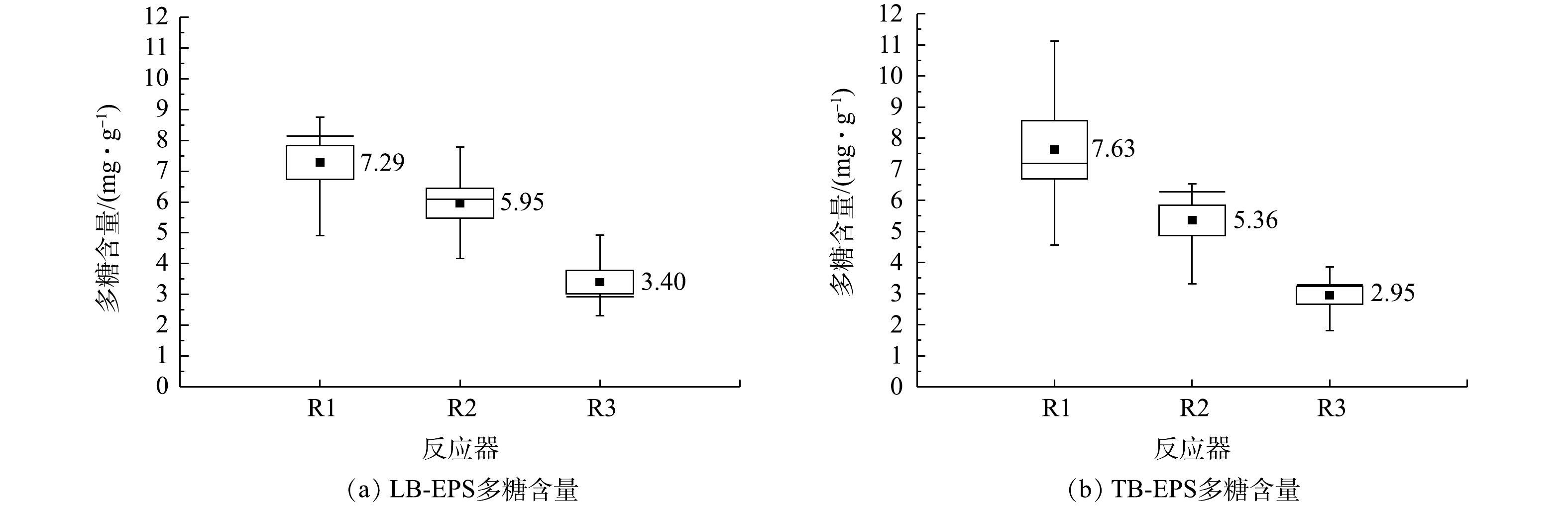
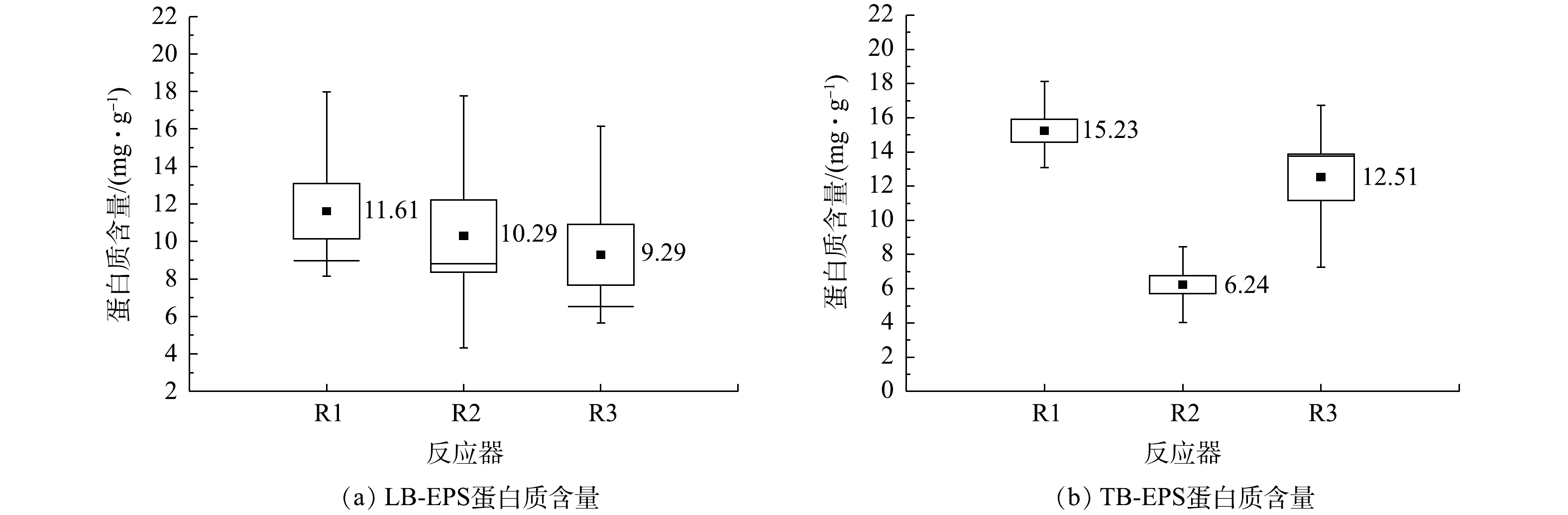
在本研究中,仅在进水碳源不同的情况下,系统内微生物分泌的EPS中PS和PN含量具有明显差异。由图7和图8可知,3个系统EPS中PS和LB-EPS中PN含量依次减少,不利于污泥的聚集和颗粒化[41]。但对于TB-EPS中的PN,R1和R3反应器的含量分别为15.23、12.51 mg·g−1(以VSS计),明显高于R2系统6.24 mg·g−1。结合3个系统颗粒污泥的粒径(R1>R2>R3)可知,TB-EPS中PN的含量与污泥颗粒化无显著相关性,推测其可能在维持污泥结构的稳定性方面具有一定的作用。
-
好氧颗粒污泥由多种带负电的微生物聚集形成,微生物的带电特性可以用Zeta电位来表示。当具有相同负电性的污泥相互靠近到一定程度时,就会发生双电层的重叠,产生静电斥力,进而阻止污泥粒子之间的进一步靠拢和聚集[42-43]。根据XDLVO理论可知,细菌表面电荷越少,静电斥力越小,总势能曲线上的“势垒”将越小,更有利于细菌落入第一极小值,促进细菌和污泥聚集体之间的相互聚集和颗粒污泥的形成[44]。絮体的Zeta电位是影响活性污泥絮凝的关键因素,不同底物条件下形成污泥的Zeta电位差异较大[45],而污泥的Zeta电位又与EPS的带电特性密切相关[46]。
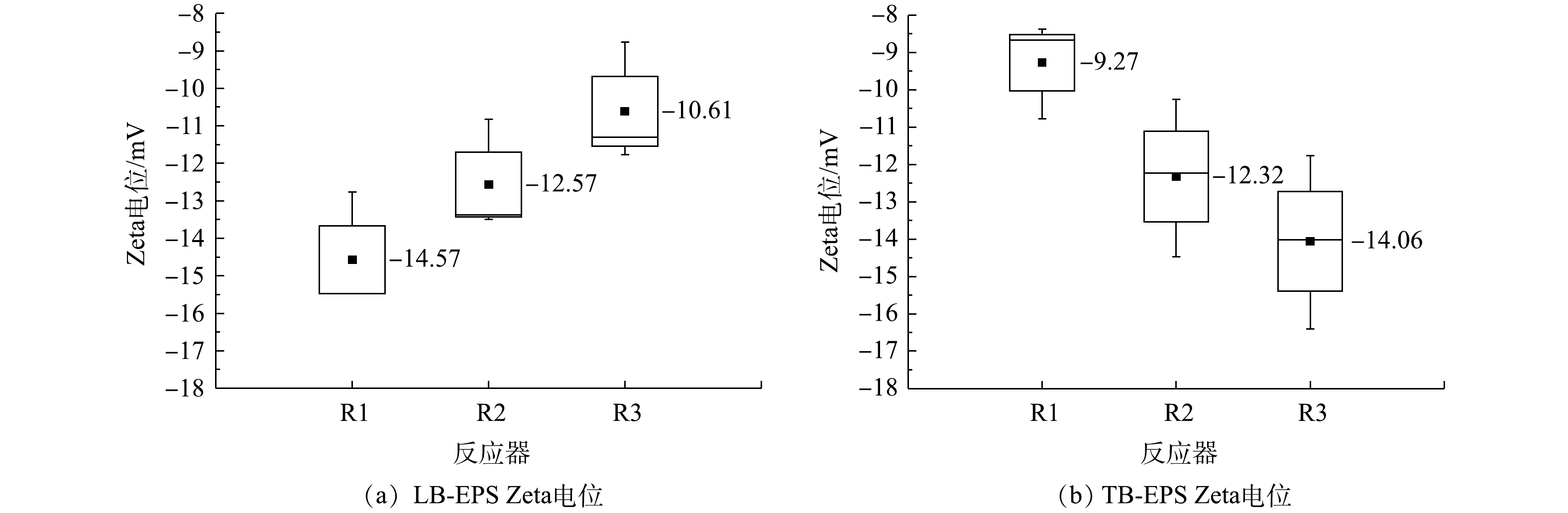
在系统运行的200 d中,在颗粒化前期(90 d)、颗粒化中期(130 d)、颗粒化成熟期(190 d) 分别对颗粒污泥LB-EPS和TB-EPS的Zeta电位进行分析,结果如图9所示。R1、R2、R3系统中TB-EPS Zeta电位的中位数分别为−9.27、−12.32、−14.06 mV,静电斥力依次增大,不利于细菌之间的聚集;LB-EPS Zeta电位的中位数依次升高,分别为−14.57、−12.57、−10.61 mV,静电斥力依次减小,有利于污泥聚集体之间的聚集和进一步的颗粒化。
-
结合研究中各项指标和显微镜观察,可提出如下大分子有机物影响下基于EPS的除磷污泥颗粒化模型(图10)。在R1溶解态有机物(NaAc)系统中,污泥表面丝状菌不断被淘汰,细菌先在多重作用下形成微小聚集体(对应TB-EPS,microcolony),再以该聚集体为中心,通过细菌分裂增殖,同时分泌大量胞外聚合物诱导其他微生物附着其上或者嵌合其内部生长,不断向外扩张。由于微小聚集体外部静电斥力较大(对应LB-EPS),各聚集体之间不易融合,因此以各自为中心独立生长,最终形成一个一个结构密实、表面光滑、形状规则的成熟颗粒污泥。可见,以溶解态小分子有机物为碳源时,除磷颗粒污泥的形成是一种从内向外逐步长大的过程。这一结论与GRACIELA-GIL等[47]和BARR等[30]采用乙酸钠或丙酸钠为碳源培养除磷颗粒污泥的过程较为相似。

在R3纯大分子有机物(淀粉+胰蛋白胨)系统中,细菌在絮体内部较大静电斥力的作用下(对应TB-EPS),形成结构较为松散的微小聚集体,由于多糖类胞外聚合物较少,诱导附着和嵌合生长的微生物较少。同时,各微小聚集体之间由于静电斥力较小(对应LB-EPS),在淀粉以及EPS的架桥作用下不断相互粘附融合,最终形成有2个或多个核心密实点的除磷颗粒污泥。成熟的颗粒污泥内部存在基质和DO传递通道,边界粗糙,形状多为椭圆形或不规则形状。由此可知,在大分子有机物为碳源的系统中,颗粒污泥的形成过程与脱稳胶体的絮凝过程相似。
在R2小分子+大分子有机物(NaAc+胰蛋白胨)系统中,污泥的聚集过程介于R1和R3系统之间,当细菌越过“势垒”时便会形成微小聚集体,由于LB-EPS的Zeta电位较高,微小聚集体可能独立生长,也可能粘附融合,最终形成具有单个或多个核心密实点的椭圆形除磷颗粒污泥。
2.1. 反应器中污泥沉降性能及除碳除磷性能分析
2.2. 光学显微镜观察
2.3. 污泥表面形貌的SEM分析
2.4. LB-EPS与TB-EPS的三维荧光光谱分析
2.5. 胞外聚合物中多糖和蛋白质含量分析
2.6. LB-EPS与TB-EPS的Zeta电位分析
2.7. 不同进水基质系统中除磷污泥颗粒化过程模型
-
1)小分子有机物使颗粒污泥结构密实,形状圆润规则,大分子有机物使污泥颗粒化进程变缓,污泥结构疏松并与小絮体共存。大分子碳源使污泥疏松型EPS(LB-EPS)组分更加丰富,碳源差异未对TB-EPS组分造成影响。随着大分子有机物占比增加,TB-EPS Zeta电位中位数依次降低,LB-EPS Zeta电位中位数依次升高。
2)进水基质会导致EPS中各组分含量发生变化,PS和LB-EPS中PN含量减少不利于污泥的聚集和颗粒化,TB-EPS中PN含量与污泥颗粒化间无显著相关性。
