

-
生物炭(biochar,BC)是生物质在限氧条件下热解(<700 ℃)形成的固态的、难熔的、高度芳香化的富含碳的材料[1],被广泛用于固碳[1-2]、土壤改良[3-4]、水体和土壤污染修复[5]等领域. 溶解性黑碳(dissolved black carbon,DBC)是BC中可溶于水的具有稠环芳烃结构的组分[6]. BC的大规模施用导致DBC释放[7],并通过灌溉、渗透、地表径流在环境中迁移[8]. 相比于BC,DBC具有较小的芳香微域结构和更多的羧基和羟基官能团[8-9],有更高的反应活性和更强的环境迁移能力[9]. 据估计,DBC全球河流通量每年可达2650万t,占河流进入海洋溶解有机碳通量的10%,是陆地BC输入海洋的一个重要来源[6, 10]. DBC的芳香微域结构是有机物和重金属吸附/络合的高能位点[9],会影响污染物的吸附、迁移、转化等关键环境过程[8]. DBC富含的羧基和羟基官能团是其在氧化-还原反应中的重要电子来源[8],参与有机污染物的光化学转化过程[10-11]. 原料和热解温度(heating temperature,HTT)是影响BC性质和用途的关键因素,也必然决定了DBC的结构特征和环境行为[12]. 研究发现,木质素含量较高的稻壳制备的BC芳香性和产量显著高于低木质素含量的甘蔗所制备的BC[3]. 另外,相比于低温(<500 ℃)制备的BC,高温(>500 ℃)BC往往具有更高的芳香性、灰分和pH[5]. 因此,深入理解水环境中不同原料和HTT的DBC与有机污染物的相互作用,有助于更好地预测BC施用环境中污染物的环境行为和评估污染物的生态风险.
光化学转化是影响水环境中有机污染物环境归趋的重要过程[13]. 水环境中光敏剂的分子结构(芳香性结构和分子大小)和环境条件(pH和盐离子)等是影响光降解的重要因素[14]. 与水环境中天然溶解有机质(natural organic matter,NOM)相比,人工来源的DBC稠环度高,芳香性强,分子量较小,这使其具有更高的光电转化效率[8, 15],更易产生羟基自由基(·OH)、单线态氧(1O2)和激发三重态DBC(3DBC*)等活性中间体(reactive intermediates,RIS)[10]. DBC的这些特性导致其对水环境中有机污染物的光降解过程的影响较NOM更为复杂[16]. 研究发现,DBC促进了塑化剂(如邻苯二甲酸二乙酯)[16]、杀虫剂(如吡虫啉)[12]、个人护理品(如阿替洛尔)[15]和性激素(如17-β雌二醇)[10]等多种有机污染物的光降解. 然而,腐殖酸(humic acid, HA)(DBC的重要组成部分)也可能会因遮光效应抑制有机物的光降解[13]. 可见,DBC参与的水环境中有机污染物的降解过程仍然存在不确定性.
新污染物是指由人类活动造成的,但尚无法律法规和标准规定或规定不完善的,危害人体健康和生态环境的污染物,主要包括环境内分泌干扰物、全氟化合物、微塑料和抗生素(antibiotics, ATs)四大类[17],而我国对于新污染物的毒理学风险评估、替代技术开发的方面还有局限性. ATs是一类由微生物分泌、化学合成或半合成的化合物,因其对微生物生长代谢的抑制而被广泛应用于医疗、畜牧业、水产养殖等领域[18]. 四环素(tetracycline, TC)因其广谱抗菌性和低廉的价格成为使用量最大的ATs之一. 2015年,全球TC使用量达6万t[19],这也导致TC在地表水、地下水和饮用水等水环境中被广泛检出,含量为ng·L−1—μg·L−1[20-21]. TC对水生动物(水生植物、浮游动物和鱼类)具有较强的生物急性毒性,能够抑制藻类和水生植物叶绿素的合成[22-23]、促进氧化应激反应的产生,影响浮游动物和鱼类的生长发育[22],同时TC还能够破坏水体微生物群落结构[21],损害水生生态系统的结构和功能[24]. 另外,ATs的长期暴露能够促进抗生素抗性基因的传播扩散,增加人体对ATs的抗药性[25],对人体健康产生极大的威胁[18, 26]. ATs在水环境中的光降解过程引起了广泛关注[13]. ATs的光降解主要受ATs种类[15]、环境条件(pH、盐离子)[27]、光敏剂(NOM、DBC)[10]等影响. 目前,水环境中DBC存在时,ATs的光降解研究以往主要集中在单一HTT制备的DBC对ATs光降解的影响上[13, 28],很少涉及不同原料和HTT的DBC对TC光降解的影响.
本研究以不同HTT(300—600 ºC)制备的湿地植物芦苇(Phragmites australis)和芦竹(Arundo donax)BC为原料提取DBC,探究HTT对DBC理化性质及结构特征的影响,系统研究了不同HTT的DBC对水环境中典型ATs TC的光降解的影响,并探讨了水中常见离子和pH对TC光降解过程的影响,以期阐明DBC影响TC光降解的主要机制. 研究结果将为评估DBC的环境行为及其对共存有机污染物的环境归趋的预测和生态风险的评估提供理论依据.
-
本研究所用BC均是实验室前期以湿地植物芦苇和芦竹为原料,利用限氧慢速热解法在300—600 ℃条件下制备[29]. DBC的提取步骤如下[10, 13]:将0.2 g BC置于20 mL超纯水中,室温条件下130 r·min−1振荡12 h,过0.45 μm醋酸纤维膜,收集滤液. 将滤膜上的BC重新分散在20 mL超纯水中,用水浴超声仪(KQ-500DE,昆山舒美,中国)在40 kHz频率下水浴超声30 min,过0.45 μm醋酸纤维膜后再次收集滤液. 重复上述超声提取过程两次,合并收集到的滤液,即为DBC溶液[10, 15]. 芦苇DBC标记为LW300、LW400和LW500,芦竹DBC标记为LZ300、LZ400、LZ500和LZ600,数字代表BC的HTT. 将DBC置于4 ℃冰箱中避光保存. TC(纯度>99%)购于生工生物工程(上海)股份有限公司,理化性质见表1.
-
总有机碳分析仪(TOC-Vcpn,Shimadzu,日本)和pH计(AB150,Thermo Fisher Scientific,美国)测定DBC的总有机碳(TOC)含量和pH. 紫外可见分光光度计(UV-9000S,上海元析,中国)测定DBC紫外-可见吸收光谱,扫描波长为100—900 nm. 荧光分光光度计(F-4600,日立,日本)分析DBC的荧光特性,测定发射波长(Em)为200—550 nm,激发波长(Ex)为220—450 nm. 傅里叶变换红外光谱仪(FTIR,Spectrum Two,PerkinElmer,英国)测定DBC的官能团,扫描区域为4000—500 cm−1,分辨率 4 cm−1 .
-
在模拟太阳光照射下,于100 mL石英管(含20 mL溶液)中进行DBC暴露下的TC光解实验[27]. 以测得TOC作为DBC原始质量浓度(表2),配置100 mg·L−1(以C的质量计)的DBC储备液. 向石英管中分别加入0.02、0.2、2 mL的DBC储备液和1 mL的TC储备液(400 mg·L−1),用超纯水定容至20 mL. 将石英管放在于以汞灯为光源的光化学反应仪(BL-GHX-V,上海比朗,中国)中,波长为290—400 nm,光功率密度为30 W·m−2,石英管以25 r·min−1匀速旋转. 反应启动后,分别在0、10、20、40、60 min时取600 µL反应溶液,过0.22 µm聚醚砜滤膜后用高效液相色谱仪(HPLC 1260,安捷伦,美国)对TC定量分析. HPLC的检测条件为:C18反向色谱柱(3.5 μm,4.6 mm×250 mm),紫外检测器,波长设为355 nm,柱温为30 ℃,流动相为1:3的乙腈和草酸(0.01 mol·L−1)溶液,流速为 1 mL·min−1,进样量为20 μL.
-
向含有2 mL DBC储备液的石英管中分别加入0、0.25、0.5、1、2 mL的TC储备液,用超纯水定容至20 mL,按照1.3.1节中条件光解、检测.
-
选择水环境中常见的阴离子(NO3−、HCO3−、SO42-、Cl−)和阳离子(Fe3+、Ca2+)进行光解实验[27]. 向含有2 mL DBC储备液和1 mL TC储备液的石英管中分别加入0、0.002、0.02、0.2、2 mL的离子储备液(100 mg·L−1),用超纯水定容至20 mL,按照1.3.1节中条件光解、检测.
-
使用0.1 mol·L−1的HCl和NaOH将含有DBC(10 mg·L−1)和TC(20 mg·L−1)混合溶液的pH分别调整为3、5、7、9和11. 按照1.3.1节中条件光解、检测.
-
为确定DBC产生的不同RIS在TC光降解中的作用,选择常用的猝灭剂异丙醇(IPA)、2,4,6-三甲基酚(TMP)和叠氮化钠(NaN3)分别验证·OH、3DBC*以及·OH和1O2在光降解过程中的作用[10, 12]. 向含有2 mL DBC储备液和1 mL TC储备液的石英管中分别添加20 μL IPA、2 mL 80 mg·L−1的NaN3和2 mL 0.545 g·L−1的TMP,用超纯水定容至20 mL,按照1.3.1节中条件光解、检测.
-
采用Excel 2016和Origin 2018进行实验结果的处理分析及图表绘制. 使用SPSS 22.0进行单因素方差分析(ANOVA),结合Duncan 检验(P < 0.05)进行显著性差异分析. 利用Origin 2018绘制三维荧光光谱图并对荧光区域积分进行定量分析. 用二级动力学模型(1/C-1/C0=kt)模拟TC光降解动力学[13],其中C和C0分别是t时刻和0时刻的TC浓度,k是二级速率常数.
-
DBC的物理化学性质如表2所示. 随着HTT升高,芦苇和芦竹DBC的TOC含量均呈先升高后降低的趋势. 这是由于在较低HTT下(300—400 ℃),BC炭化程度低,溶解性物质含量较高[3, 30];当HTT升至500 ℃时,纤维素和木质素进一步分解聚合,产生更多稳定性较强的芳烃类物质[3]. 另外,芦竹DBC的TOC含量(101—832 mg·L−1)高于芦苇DBC(98.9—152 mg·L−1),这可能是芦竹生物质较芦苇含有更多的纤维素和半纤维素[31],导致相同HTT的芦竹BC稳定性较低,含有更多的可溶性物质. 随着HTT升高,芦苇和芦竹DBC的pH分别由4.25和5.66上升至5.96和10.2,这是因为HTT升高导致BC中矿物质富集,且矿物质的溶解使得DBC的溶液的pH增加[3]. E4/E6是有机质的芳香性指标,与芳香性呈反比[12]. 随着HTT的升高,芦苇和芦竹DBC的E4/E6由7.17和5.48降低至3.88和2.00(除LZ400外),说明高HTT的DBC芳香性更高. 这是因为BC形成时,生物质发生脱水和裂解反应导致不稳定有机物分解,而聚合反应增加了芳香性物质的含量[5]. 值得注意的是,LZ400的E4/E6大幅升高,这可能是因为纤维素分解为酸醛等可溶性高的物质[32],导致芳香性组分在水相总有机质中占比下降. SUVA254是有机质的芳香性指标,与芳香性呈正比[12]. 随着HTT升高SUVA254先降低后升高,这表明芳香性在400 ℃时降低,但高温DBC芳香性更高这与E4/E6的结果一致. 相同HTT下,芦苇DBC(0.12—0.18 L·(mg·C·m-1)−1)比芦竹DBC(0.02—0.17 L·(mg·C·m-1)−1)的芳香性更强,这可能是由于芦苇生物质的木质素含量高,产生的BC芳香性物质更多[31]. E2/E3是有机质分子量大小的指标,与其分子量呈反比[12]. 两类DBC的E2/E3随HTT增加先升高后降低,表明DBC分子量先降低后上升,这与芦苇和芦竹热解形成BC的过程中脂肪烃分解和芳香烃的形成有关[30]. 另外,相同HTT下,芦竹DBC的E2/E3比值(3.81—7.28)高于芦苇DBC(3.67—5.01),表明芦竹DBC平均分子量更小,这与TOC的含量变化规律一致,表明小分子量物质可能更易从BC溶出,形成DBC[33].
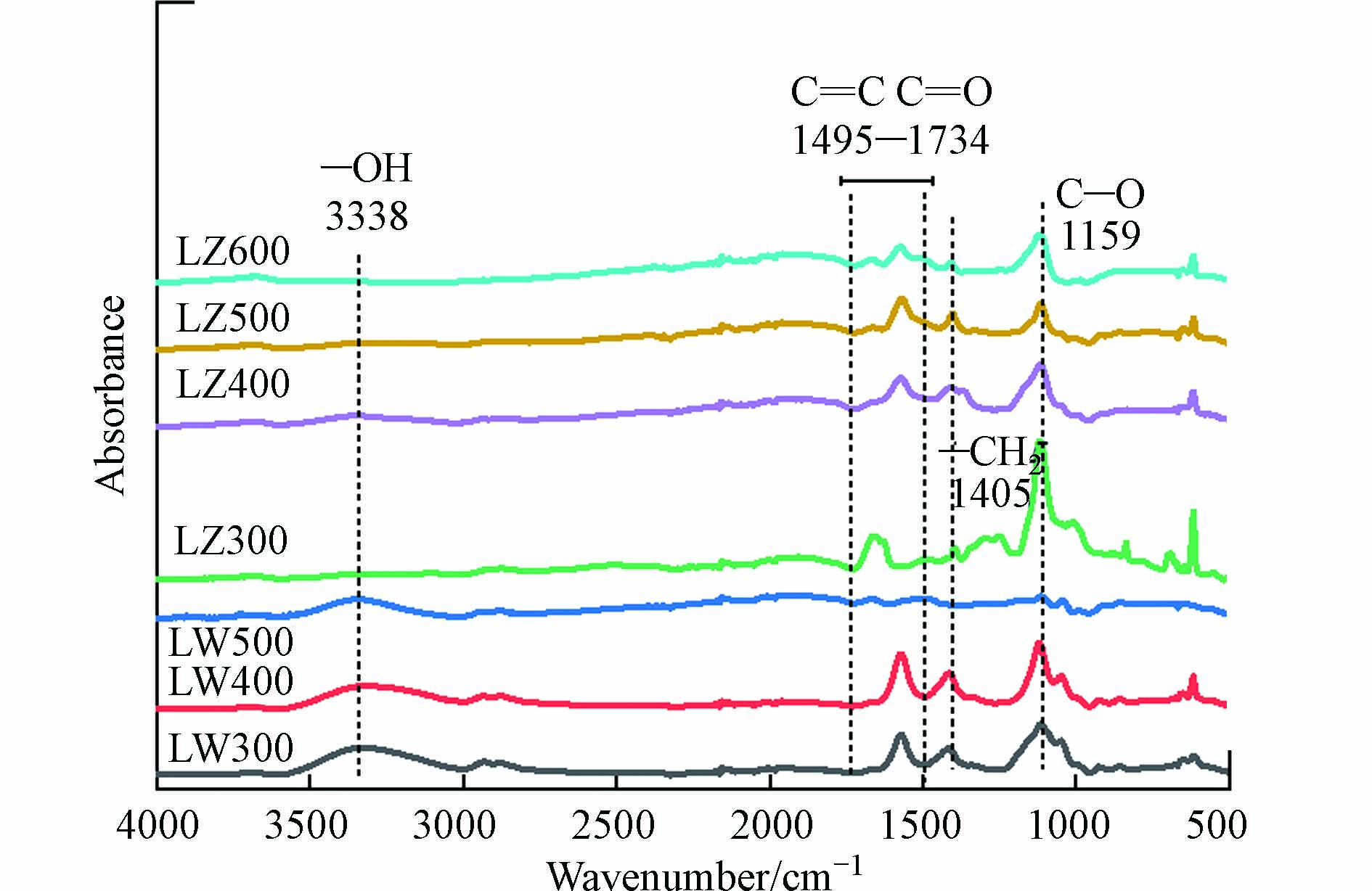
DBC的FTIR图谱如图1所示. 对于芦竹DBC,其在1495—1734 cm−1(C=C、C=O)、1405 cm−1(—CH2—)和1159 cm−1(C—O)处有明显的吸收峰[34].
随着HTT的升高,C—O的峰大幅度降低,—CH2—的峰也变小,表明不稳定的脂肪族化合物逐渐被分解,而C=C和C=O虽有小幅度降低,但仍有较大的振动峰存在,表明芦竹DBC仍具有较高的芳香性组分[34]. 对于芦苇DBC,在3338 cm−1(酚羟基,—OH)[29, 35]、1495—1734 cm−1(C=C、C=O)、1405 cm−1(—CH2—)和1159 cm−1(C—O)处有明显的吸收峰. 随着HTT升高,—CH2—和C—O的峰逐渐消失,表明不稳定的脂肪烃逐渐分解,C=C和C=O的峰虽也消失,但酚—OH的峰却无明显变化,说明芦苇DBC中的芳香性组分虽有分解,但仍有较高含量的芳香性组分存在. 总体来说,随着HTT的升高,DBC中脂肪族化合物的含量降低,芳香性组分含量增加,这与DBC的芳香性增加的结果一致(表2).
DBC发色基团的含量对于其光化学反应的发生至关重要[36]. 因此对DBC进行了三维荧光光谱(3D-EEM)分析(图2).
LW300、LW400、LZ300、LZ400和LZ500在Ⅲ(Ex:200—250 nm和Em:380—550 nm)、Ⅳ(Ex: 200—250 nm和Em: 330—380 nm)和Ⅴ(Ex: 250—400 nm和Em: 380—550 nm)处有明显的荧光峰,它们分别代表富里酸(fulvic acid, FA)类、溶解性微生物代谢产物和HA类物质. LW500和LZ600仅在Ⅲ和Ⅴ处有荧光峰,表明存在FA类和HA类物质. 所有DBC在Ⅰ(Ex:200—250 nm和Em:280—330 nm)和Ⅱ(Ex:200—250 nm和Em:330—380 nm)区域的荧光峰不明显,表明存在芳香性类蛋白质物质. 这些结果表明FA类和HA类物质是DBC的主要组分. 随HTT升高,芦苇DBC FA和HA相对含量分别由21.9%和49.7%增加至24.9%和63.6%,溶解性微生物代谢产物含量由17.1%降至7.5%,芦竹DBCFA类含量由15.6%增加到26.6%,溶解性微生物代谢产物和HA的含量分别由59.3%和18.2%减少到56.4%和9.7%. FA类和HA类物质的总含量随HTT升高呈上升趋势(71.6%—88.5%). NOM中的类FA和HA类物质是重要的光敏剂,吸收光子后会生成激发三重态HA(3HA*)和FA(3FA*),可通过能量转换或者电子转移降解有机污染物[37]. 因此,含有FA类和HA类组分的DBC也可能促进水体中ATs等有机污染物的光降解,这需要进一步的研究.
-
不同种类DBC对TC光降解的影响如图3所示. 黑暗条件下,DBC对TC浓度无明显影响(图3a),表明非光化学过程对TC降解作用不明显. 光照条件下,DBC不存在时,TC的直接光降解率较低(6.2%),但DBC的存在显著增加了TC的光解率(16.3%—97.0%)(图3b). 用二级动力学方程拟合光解速率k,在不添加DBC时,k为8.3×10−5,添加DBC后,k分别升高为1.7×10−4、1.8×10−4、2.6×10−4、8.3×10−4、8.4×10−4、9.0×10−4和8.8×10−2(图3c),表明DBC促进了TC的光解速率,且高温DBC促进效果更强. 相同HTT条件下,芦竹DBC对k的促进效果高于芦苇DBC. 7种DBC中,LZ600对TC的光解的促进最显著,且LZ600在10 mg·L−1的添加量下,TC光解率最高(图3d). 另外,当TC的初始浓度从5 mg·L−1增加至40 mg·L−1时,TC的光解速率从0.32 L·(mg·min)−1升高至1.45 L·(mg·min)−1,然后降低至0.15 L·(mg·min)−1(R2>0.91). 其中,TC浓度为20 mg·L−1时,其光解速率最快. 因此,选用10 mg·L−1 LZ600和20 mg·L−1的TC进行后续实验. ZHOU等[10]也发现竹子DBC能够将17-β雌二醇的一阶速率常数提高1个数量级,这与上述DBC增加k值的结果一致,说明DBC能够作为一种高效的光敏剂促进TC的光降解.
-
实际水环境中存在多种阴离子(如NO3−、HCO3−、SO42−、Cl−)和阳离子(如Fe3+、Ca2+),可能影响DBC的光敏性及污染物的光转化过程[38-43]. LZ600存在的条件下,与不添加离子的对照组相比(99.6%),当溶液中存在NO3−、HCO3−、SO42−、Cl−、Fe3+和Ca2+时,TC的光降解率降为76.0%—80.0%、76.3%—81.0%、71.5%—77.4%、66.8%—69.0%、16.0%—97.3%和1.2%—54.8%(图4a—g).
另外,不添加离子时,k为6.6×10−2 L·(mg·min)−1,添加离子后,k降为3.4×10−3—3.9×10−3、3.4×10−3—4.2×10−3、3.5×10−3—3.7×10−3、2.2×10−3—2.7×10−3、2.5×10−4—2.6×10−2和9.8×10−4—1.3×10−3 L·(mg·min)−1 (图4g). 结果表明,环境中常见的离子对DBC介导的TC光降解有一定抑制效果,抑制效果受离子种类和浓度的影响. ZHANG等[27]研究表明HCO3−和Cl−能够与DBC产生的RIS反应,生成活性小于RIS的次级活性中间体,抑制抗生素抗性基因的光解. MACK和ZHOU等[40-41]发现,NO3−和SO42−可以抑制DBC对光子的吸收而抑制DBC光解. OU等[42]发现,Ca2+能够与DBC形成内球配合体,促进DBC沉降进而抑制其光反应. Fe3+在0.01—0.1 mg·L−1添加量下显著抑制DBC的光解,这是由于Fe3+与DBC竞争光子抑制了RIS的生成[43]. 但当添加量为1—10 mg·L−1时,Fe3+对DBC光解的抑制效果被缓解,这可能是因为Fe3+与水分子和DBC产生了光敏剂Fe(OH)2+和Fe-DBC剂促进了RIS的产生[43-44]. 简而言之,离子通过影响DBC对光子的吸收以及RIS的活性影响TC的光降解.
pH会影响DBC的表面活性和TC的电子密度[42, 45],因此pH可能会对TC的降解产生影响. 不同pH(3、5、7、9和11)对DBC影响的TC光降解如图4h. 随着pH增加,TC光解率从3.7%增加到67.9%,k从5.0×10−5增加到2.3×10−3 L·(mg·min)−1. 这表明碱性条件下,LZ600更有助于促进TC的光降解. 这与李聪鹤等[46]研究一致,在pH为11的条件下,溶解性有机质对磺胺甲恶唑的降解效果最显著. 这可能是由于pH的增加有助于增加DBC的稳定性[42]及促进RIS的产生[46],同时高pH条件下,TC去质子化程度增大,电子密度增加,更易受到RIS的攻击[46-47].
-
DBC是水环境重要的光敏剂,会在光照下发生化学转化[15]. 为探究光解前后LZ600结构组成的变化,测定了光降解后LZ600组成及含量(图5a). 与原始LZ600相比(图2g),光照后LZ600组分Ⅰ—Ⅴ的荧光强度显著降低(14.1%—35.0%),其中FA类和HA类对荧光强度变化的贡献最大(27.1%和54.2%),表明HA类和FA类物质在光照下发生了明显的转化. 先前的研究表明,FA和HA的添加能够产生大量的RIS,将ATs的降解速率提高34%[13]. 因此,DBC导致的TC光解的增强可能归因于FA类和HA类物质的光转化而产生的强氧化性的RIS [15].
为了进一步探究DBC在光照下产生的不同RIS(·OH、1O2和3DBC*)在光解中的作用,进行了RIS猝灭实验. 如图5b所示,添加TMP(3DBC*猝灭剂)后,DBC对TC的光解率从97.0%降至84.0%,NaN3(·OH和1O2猝灭剂)和IPA(·OH猝灭剂)的加入对TC光解影响较小. 这说明TC的光降解的增强主要归因于3DBC*. 另外,TMP使得k从1.1×10−2 L·( mg·min) −1降至1.1×10−3 L·(mg·min)−1,IPA使k从1.1×10−2 L·( mg·min) −1降至8.9×103 L·(mg·min)−1,NaN3使k从1.1×10−2 L·( mg·min) −1降至6.8×103 L·(mg·min)−1,NaN3与IPA的k差值为2.1×10−3 L·(mg·min)−1. 这进一步说明DBC对TC的降解由3DBC*起主导作用,·OH和1O2也促进了光解,但贡献较小[10].
不同HTT下DBC的芳香性、分子量及FA和HA类物质的总含量均不同,为探究这些性质是否会影响DBC对TC光解的效果,进行了TC光解率与DBC的SUVA254、E2/E3、E4/E6、pH和HA和FA总含量的相关性分析(图5c). 结果表明TC光解率与pH呈显著正相关,说明pH很大程度上影响了DBC对TC的光解,可能是因为低pH时,DBC表面会形成致密的层,不易发生光反应[42];高pH时,DBC中HA和FA产生的小分子酸更容易被消耗,使化学平衡不断向RIS生成方向进行,增加了RIS含量[48]. 除pH外,DBC的芳香性(SUV254)也与TC的光解率呈显著正相关,这是因为芳香性物质是DBC吸收光子的主要组分,芳香性的升高会增加DBC光子的吸收进而促进RIS的产生[15]. 除pH和芳香性外,HA和FA含量与TC的光解率也呈正相关,也可能会影响DBC对TC的光解效果,但不是主要原因,DBC的分子量与光解率的相关性较低,对TC的光降解影响较小. 结果进一步说明,DBC促进TC的光降解主要是由于DBC具有的稠环芳烃类结构导致的RIS(3DBC*)的产生引起的(图6).
-
本研究针对不同热解温度和生物质类型的DBC对水环境中ATs光降解影响效果不明确的问题,重点评估了不同原料和热解温度条件下制备的DBC的理化性质及结构特征,探究了不同制备条件的DBC对ATs光降解的影响和机制及关键水环境条件对ATs光降解的影响. 研究发现:
(1)随着热解温度升高,DBC的TOC先上升后下降,分子量先降低后上升,芳香性逐渐增强,类FA和类HA相对含量增加.
(2)DBC对TC的光降解均有促进作用. 随着热解温度升高,芦苇和芦竹DBC对TC光降解的促进作用增加,在相同热解温度下,芦竹DBC对TC光降解的促进效果较芦苇DBC更高.
(3)水中常见阴离子(NO3−、HCO3−、SO42−、Cl−)和阳离子(Fe3+、Ca2+)均对TC的光降解有抑制作用,抑制效果受离子种类和浓度的影响. DBC在碱性条件下对TC光降解的促进作用较酸性条件更高.
(4)TC的光降解主要由DBC中的富里酸和腐殖酸类物质光转化产生的·OH、1O2和3DBC*等活性中间体诱发,其中TC的光降解由3DBC*主导,·OH、1O2对TC光降解的贡献较小.
溶解性黑碳促进水环境中四环素的光降解
Dissolved Black Carbon Enhanced the Photodegradation of Tetracycline in Aqueous Solution
-
摘要: 具有固碳功能的生物炭材料在土壤修复和水体污染治理中的大规模施用导致了溶解性黑碳的(dissolved black carbon,DBC)的大量释放,其在污染物的环境地球化学过程发挥着重要作用. 相比于天然溶解有机质,DBC稠环度高、芳香性强、分子量小,有更高的光电转化效率,更易产生活性中间体促进有机污染物的光降解. 但不同热解温度(heating temperature,HTT)和生物质类型的DBC对水环境中抗生素(antibiotics, ATs)的光降解影响尚有待深入研究. 本文选取不同HTT(300—600 ℃)的芦苇和芦竹生物炭制备DBC,表征其基本理化性质及结构特征,研究其对水环境中典型ATs四环素(tetracycline, TC)光降解过程的影响,探究关键水环境条件的影响. 结果表明,随着HTT升高,两类DBC的有机碳含量呈先升高后降低趋势,平均分子量呈先降低后增加趋势;芳香性官能团含量增加,芳香性增强,腐殖酸和富里酸类物质含量升高. 所有DBC均促进了TC的光降解(16.3%—97.0%),促进效果随HTT的升高而呈上升趋势. HTT相同时,芦竹DBC对TC光降解的促进效果高于芦苇DBC. 水环境中常见阴离子(NO3−、HCO3−、SO42−、Cl−)及阳离子(Fe3+、Ca2+)均在不同程度上抑制了TC的光降解;碱性(pH 7—11)环境中DBC对TC光降解的促进作用强于酸性(pH 3—5)环境. 活性中间体猝灭实验表明DBC对TC光降解的促进作用主要由三重激发态DBC(3DBC*)主导. 本研究拓展了人们对水环境中DBC环境行为及其对共存ATs归趋的理解,为水环境中ATs环境过程和生态风险的预测提供了理论依据.Abstract: The large-scale application of biochars with the function of carbon sequestration in soil remediation and water pollution control has resulted in the discharges of dissolved black carbon (DBC), which plays an important role in the environmental geochemical process of various pollutants. Compared with natural dissolved organic matter (NOM), DBC with higher condensed degrees, stronger aromaticity, smaller molecular weight, higher photoelectric activity, potentially generates more active intermediates to facilitate the photodegradation of organic pollutants. However, the effects of DBCs produced from different biomass under different heating temperature (HTT) on the photodegradation of antibiotics in water environment was poorly understood. Therefore, the biochars derived from reed and giant reed at 300—600 ℃ were selected to prepare the DBCs, and their basic physical, chemical properties and structural characteristics were also characterized to investigate the effects of DBCs on photodegradation process of typical antibiotic tetracycline (TC) in the water environment, and the influences of related key environmental factors were also considered. With increasing HTT, the organic carbon content of DBC increased first and then decreased, molecular weight decreased first and then increased, the aromaticity, contents of aromatic functional groups, and humic acid and fulvic acid substance increased. All the tested DBCs consistently promoted the photodegradation of TC by 16.3%—97.0%, and the promoting effect was elevated by the increasing HTT. At the same HTT, DBCs derived from giant reed showed a greater promoted effect on the TC photodegradation relative to the DBCs derived from reed. Anions (NO3−, HCO3−, SO42−, Cl−) and cations (Fe3+, Ca2+) that are frequently detected in the water environment generally inhibited the TC photodegradation to the different extent. DBC showed a greater promoted effect on TC degradation in alkaline (pH 7—11) environment than acidic (pH 3—5) environment. The quenching experiment of reactive intermediates revealed that the TC photodegradation by DBC was mainly mediated by the triple excited state DBC (3DBC*). These findings expand our understanding regarding the environmental behavior of DBC and the fate of coexisted ATs in water environment, and provide the theoretical basis for predicting ATs environmental process and ecological risk in water environment.
-
Key words:
- biochar /
- dissolved black carbon /
- heating temperature /
- antibiotics /
- photodegradation /
- reactive intermediates
-
铁是一种生物必须的营养元素,直接影响浮游植物的光合作用和碳水化合物形成,由于高含氧量和无机态铁的低溶解性,铁通常是制约HNLC海区(High nutrition low chlorophyll)初级生产力的关键微量元素[1-2]。大规模海洋施铁实验表明,水生态系统中生物可利用铁的增加可以显著提高浮游植物的生物量和光合作用率,从而提高初级生产力,并促使浮游植物的群落结构发生变化[3-7]。以往研究表明,光自养微生物对碳循环和全球气候起关键作用[8-9]。初级生产力的提高,深刻地影响着全球尺度的二氧化碳固定,对温室气体的控制具有重要意义。
水生态系统中,99%溶解性铁(dissolved iron,DFe)与有机配体结合。尽管大部分有机络合态铁不能直接被藻类利用,但通过一些地球化学转化过程,可转变为生物可利用铁[10-11]。Blazevic等[12]研究发现,海洋中腐殖酸结合态铁可以发生光还原反应,进而提高铁的生物可利用性。沼泽性河流是海洋DFe的重要来源[13]。沼泽性河流中大量存在的溶解性有机碳(DOC, dissolved organic carbon)与铁离子形成有机络合物,使水中保持较高浓度DFe。有机质中羧基和酚羟基是与铁络合的主要官能团。泥炭源中的酚酸类物质,含有稳定的芳香环结构。部分酚酸与铁有较高的配合能力,这类物质的存在保护了长距离迁移的DFe,保证了陆源DFe向水生态系统的有效输出[14]。
泥炭沼泽中存在多种类型酚酸,前人在金川泥炭中检测出了9种酚酸,包括对-羟基苯甲酸、丁香酸、香草酸、阿魏酸、对-香豆酸、没食子酸、原儿茶酸和咖啡酸,许多泥炭沼泽中都有这些酚酸的存在[14]。研究证实,酚酸等有机质可以和铁形成较为稳定的配合物,使其可以在淡水运输过程中迁移更长的距离[15]。其中,具有儿茶酚或者没食子酰基结构的原儿茶酸、没食子酸以及咖啡酸可以和Fe(Ⅱ)形成较为稳定的络合物,使得Fe(Ⅱ)在极易被氧化的碱性条件下也可以保存较长时间。而咖啡酸、没食子酸、原儿茶酸以及龙胆酸还对Fe(Ⅲ)有着明显的还原作用,同样有助于这两种铁形态之间的平衡[16]。
植物或微生物分泌代谢物质对环境中其他植物或微生物体产生不利或有利的影响,这种作用称为化感作用。在化感作用过程中分泌的物质即被称为化感物质,自然界的化感物质种类非常丰富,主要包括酚酸类、苯醌类、倍半萜类、黄酮类等几大类物质[17]。迄今发现的化感物质几乎都是植物的次生代谢物质,分子量较小,结构简单,其中酚酸类物质是一类重要的次生代谢产物,也是研究较多,被证实是化感活性较强的一类物质[18]。酚酸具有一定生物毒性。目前对于酚酸抑藻的机制还不十分清楚,其抑制作用可能通过多种方式实现。研究表明,酚酸与蛋白质分子易遵循疏水键-氢键多点键合理论结合。在酚酸存在的情况下,藻细胞的胞外磷酸酶活性受到抑制,碱性磷酸酶活性的抑制使藻利用磷的能力下降。酚酸与细胞膜蛋白的结合,会破坏生物体细胞膜结构,使植物多酚物质进一步穿过细胞膜,进入细胞体内,从而改变微生物细胞酶活性,减少藻类对外源性蛋白质的利用,并通过对细胞外酶的抑制达到抑藻的目的[19]。另外,如果酚酸进入细胞体后,通过与金属离子发生络合反应,形成沉淀而破坏微生物的正常新陈代谢也是植物多酚抑藻的原因所在[20]。尽管酚酸存在生物毒性,但适量前提下,对藻类生长有积极作用[21]。泥炭源典型酚酸与铁的络合物是否对藻类利用铁有显著影响尚待进一步研究。因此,探究酚-铁配合物络合稳定性及其生物可利用性有助于进一步了解生物对铁的吸收,更好地理解全球铁碳耦合循环。
铜绿微囊藻(microcystic aeruginosa) 是中国湖泊、水库及其他水域生态系统水体富营养化蓝藻水华的代表性藻类。本文铜绿微囊藻为培养对象,利用泥炭源典型酚酸及泥炭溶解有机质(DOM)开展了一系列培养试验,以期了解泥炭沼泽源酚酸以及酚-铁络合物对铜绿微囊藻生长的影响。
1. 材料与方法(Materials and methods)
1.1 实验材料
试验所用铜绿微囊藻藻源,由中国科学院水生生物研究所提供,采用BG-11培养基培养。
4种酚酸的配制:以1.7 g·L−1的浓度配制BG-11培养基,然后将对羟基苯甲酸、对香豆酸、水杨酸、咖啡酸加入,分别配制4份浓度为0.24 g·L−1的酚酸溶液。用0.22 μm滤膜在超净台中过滤,并用紫外光照射30 min,消除微生物的影响,现配现用。
藻种培养条件:实验前5天,将铜绿微囊藻进行扩大培养。光照强度4000 lx;光暗比24 h∶0 h;温度(25±1)°C;每天摇动培养瓶5次,使藻类生长进入对数生长期。进入对数增长期后,取铜绿微囊藻各300 mL加入到1 L锥形瓶,再加入BG-11培养基100 mL进行驯化培养。
1.2 实验设计
1.2.1 单一酚酸对铜绿微囊藻的抑制作用
由于对羟基苯甲酸和对香豆酸在泥炭中含量相对较高,水杨酸和咖啡酸和铁离子可以络合,实验选择这4种酚酸进行实验。
使用细胞计数仪确定当前藻液浓度,并根据藻液浓度取一定量的处于对数生长期的藻液加入250 mL锥形瓶内,其中分别添加稀释了不同倍数的酚酸溶液,最后用培养基补足,使得锥形瓶内的液体总体积达到150 mL。每个锥形瓶内藻的初始密度为105 cell·mL−1,酚酸的最终浓度梯度分别为0、10、20、40、60、80 mg·L−1,每组3个平行,置于光照培养箱内。培养温度为(20±1)℃,光照强度为4000 lx,24 h光照,每天震荡3—5次。
1.2.2 酚络合态铁对铜绿微囊藻生长的影响
选用水杨酸和咖啡酸与铁形成络合物,探究酚铁对藻类生长的影响。实验选择的酚酸浓度为5×10−5 mol·L−1,铁浓度为1×10−6 mol·L−1,在此条件下酚酸浓度为铁浓度的50倍,可以有效保护体系中的二价铁。此外,由于泥炭沼泽中也普遍存在草酸、柠檬酸、酒石酸、乙酸等无苯环的小分子有机酸,所以实验选择草酸、柠檬酸、乙酸作为干扰物质加入到酚铁体系中进行藻类的培养实验。
藻类的培养实验分为10组,每组添加的物质如下:A.水杨酸+硫酸亚铁;B.咖啡酸+硫酸亚铁;C.水杨酸+草酸+硫酸亚铁;D.水杨酸+乙酸+硫酸亚铁;E.咖啡酸+草酸+硫酸亚铁;F.咖啡酸+乙酸+硫酸亚铁;G.水杨酸+柠檬酸+硫酸亚铁;H.咖啡酸+柠檬酸+硫酸亚铁;I.不添加酸和铁的对照组;J.只添加铁的对照组。
以上试验均为期15 d,每隔48 h取样1次,记录藻细胞数量的变化,以及藻存活情况的变化和pH值的变化。培养周期结束后分别取10 mL和5 mL样品,测量样品中叶绿素a的含量和叶绿素荧光参数Fv/Fm(最大光能转化效率)。其中Fv/Fm常用来表征叶绿素PsⅡ(低铁环境藻类光系统Ⅱ)反应中心内禀光能转换效率,反映当时所有的PsⅡ反应中心均处于开放态时最大光量子产量。
1.2.3 生理指标的测定方法:
为测藻细胞存活率及藻细胞数量,使用5-CFDA染色,具体操作方法如下:
(1)用DMSO(二甲基亚砜)5-CFDA稀释至10 mmol·L−1。将99 μL已经配制好的BG-11培养基与1 μL的5-CFDA混合作为A液,摇晃10 s混匀;
(2)将50 μL样品与50 μL A液混匀,用移液枪至少吹打10次;
(3)在25℃条件下将上一步准备好的样品避光放置30 min;
(4)用移液枪将待测样品吹打10次或摇晃,使藻细胞分散,然后用移液枪取20 μL加入计数板内。在计数板插入仪器之前,稳定1 min,使样品在其中稳定下来。
将细胞染色后,使用细胞计数仪计数,并观察细胞的存活状态。
测量样品叶绿素a的含量:
(1)取藻液10 mL,4500 r·min-1离心15 min,去掉上层清液,将样品在4℃冰箱中放置1 d;
(2)取出后迅速加入5 mL的90%热乙醇(80℃)于80℃的热水浴萃取2 min,再用超声处理10 min,放在暗处萃取4 h后,用0.22 μm的滤头过滤。用酶标仪于波长665 nm和750 nm处测吸光值,然后滴加1滴1 mol·L−1的盐酸酸化,于波长665 nm和750 nm处再测吸光值。计算公式为:
Chla乙醇=27.9×[(E665−E750)−A665+A750)]×V乙醇/V样品 其中,Chla乙醇为热乙醇法测定的叶绿素a含量(μg·L−1);E665是乙醇萃取液于波长665 nm的吸光值;E750是乙醇萃取液于波长750 nm的吸光值;A665为乙醇萃取液酸化后在665 nm处的吸光值;A750为乙醇萃取液在750 nm处的吸光值;V乙醇为乙醇萃取液的体积(mL),V样品为所取样品的体积(mL)。
抑制率计算公式:IR=(1-N/N0×100%)抑制率为负值则有促进效果,抑制率为正则抑制。
1.2.4 泥炭沼泽源酚络合态铁对铜绿微囊藻生长的影响
培养前准备:
(1)提取泥炭中的DOM;
(2)将样品通过H型阳离子柱交换柱,去除样品中存在的金属离子;然后将DOM样品按照<1 KDa,1—3 KDa,>3 KDa分成3份;
(3)将3份样品中加入FeSO4,待稳定一段时间后,取10 mL加入培养基中,加入后Fe的浓度为5×10−6 mol·L−1。
铜绿微囊藻培养实验分为5组,每组3份平行,每组添加的物质如下:A.无添加;B.Fe,浓度为5×10−6 mol·L−1;C.DOM-Fe(<1 KDa),Fe浓度为5×10−6 mol·L−1;D.DOM-Fe(1—3 KDa),Fe浓度为5×10−6 mol·L−1;E.DOM-Fe(>3 KDa),Fe浓度为5×10−6 mol·L−1。
实验均为期15 d,每隔48 h取样1次,记录藻细胞数量的变化,以及藻存活情况的变化。
2. 结果与讨论(Results and discussion)
2.1 不同酚酸影响微囊藻生长的浓度效应
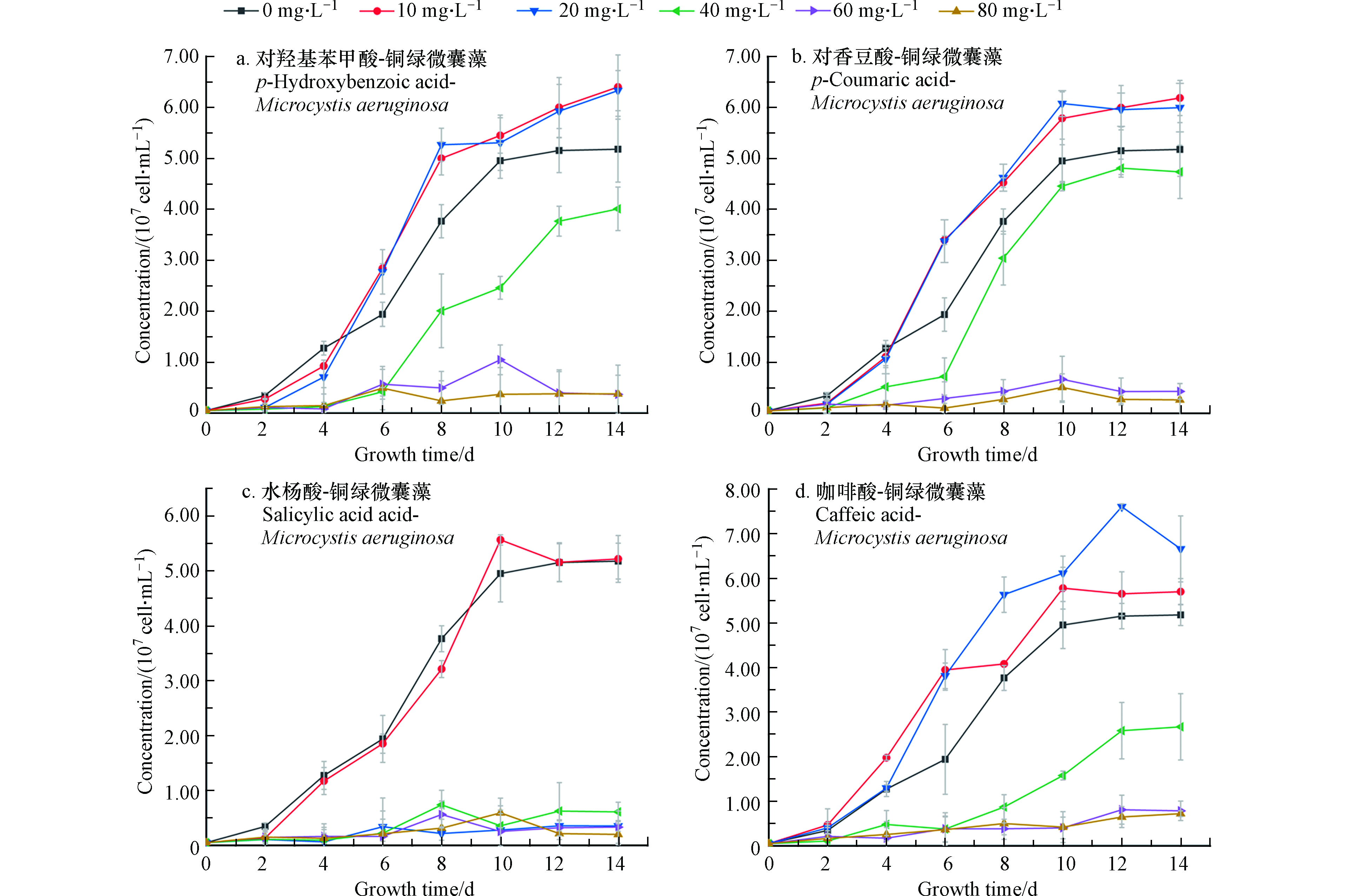
在微囊藻培养体系中分别加入不同浓度四种酚酸溶液,经过14 d培养和检测,得到微囊藻-酚酸的生长曲线如图1所示。图1中A、B为不同浓度的对羟基苯甲酸和对香豆酸对微囊藻生长情况的影响。可以看出,当酚酸浓度为10 mg·L−1和20 mg·L−1时,微囊藻的生长速率和最终达到的终点浓度明显高于控制组(0)抑制率为-23.5%—18%。从微囊藻浓度上来看,当酚酸浓度为10 mg·L−1和20 mg·L−1时,生长情况比较接近,说明在此浓度下,这两种酚酸对微囊藻的生长有一定的刺激作用。在A、B组中当酚酸浓度超过20 mg·L−1时,藻液浓度和藻的生长速率明显低于控制组;当浓度增加到40 mg·L−1时,这两种酚酸对微囊藻的生长起到了明显的抑制作用,而对羟基苯甲酸的对微囊藻生长的抑制作用更加明显;浓度继续增加到60—80 mg·L−1时,微囊藻前4—5天略有增长,然后基本停止了增长,保持在3×106 cell·mL−1左右。对香豆酸60 mg·L−1组的藻数量略高于80 mg·L−1组。
在C组水杨酸-微囊藻的实验中,水杨酸浓度为10 mg·L−1时,增长的速度与控制组接近,在8—10 d快速增长后,藻数量和控制组趋于一致。而当浓度大于10 mg·L−1时,都出现了明显的抑藻效果,抑制率约为80%。
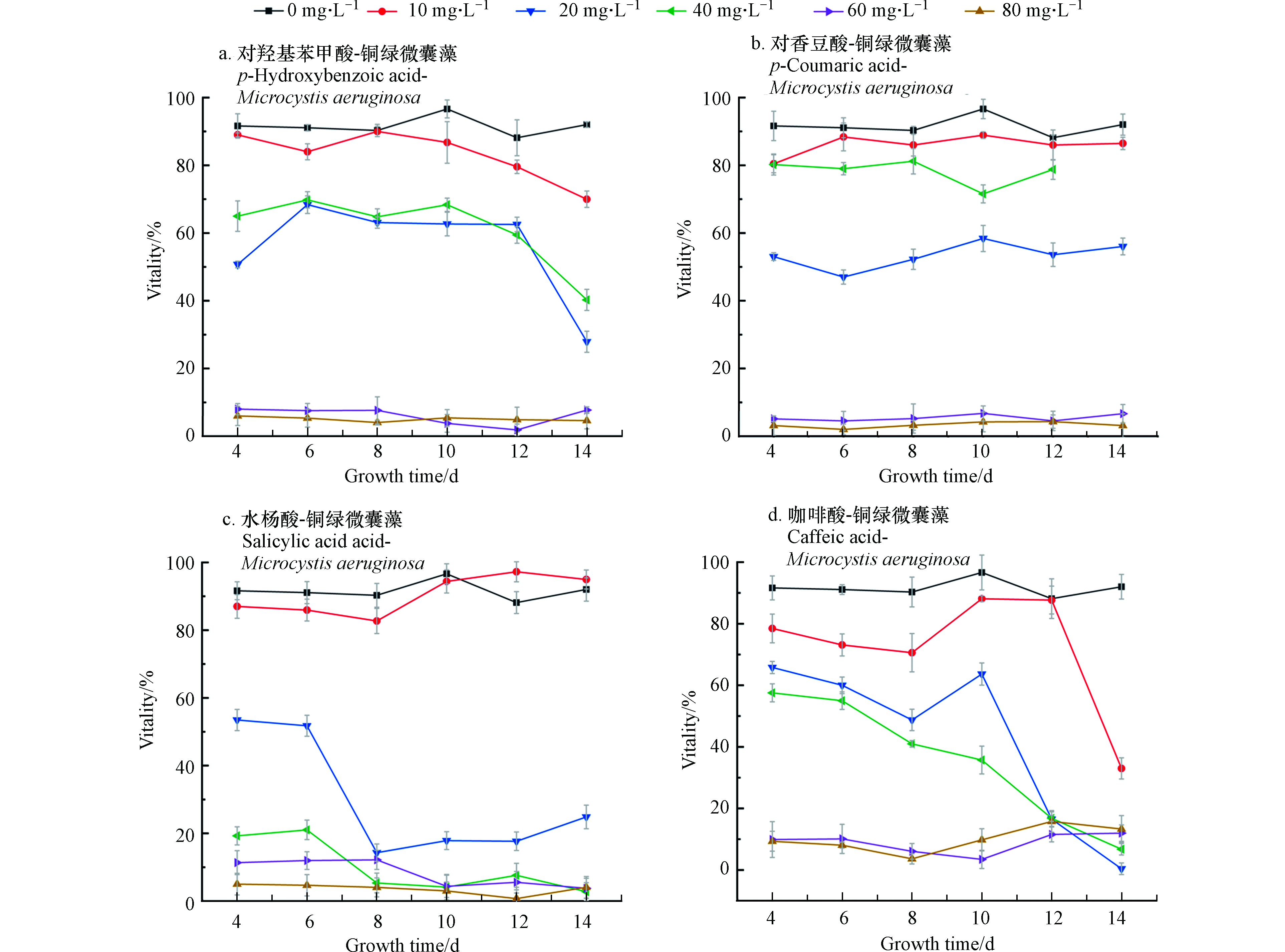
结合图2中存活率来看(从培养的第4天开始测微囊藻的存活率)当浓度为10 mg·L−1时,微囊藻的存活率都略低于控制组,差值在10%左右浮动,当浓度为20—40 mg·L−1,在开始计数时,藻类的存活率就已不同程度地低于控制组,并且A、C、D存活率在4—14 d整体处于下降的趋势,可以看出,水杨酸对微囊藻的抑制作用最强。
表1是各组样品的Fv/Fm,最大光能转化效率Fv/Fm常用来表征叶绿素PsII反应中心内禀光能转换效率,反映当时所有的PsII反应中心均处于开放态时最大光量子产量。
在非胁迫环境下,植物叶片叶绿素荧光参数Fv/Fm变化极小,表现出稳定的特点,但在胁迫条件下,该参数明显下降[22]。Fv/Fm可作为植物受环境胁迫的响应指标[23]。控制组的Fv/Fm为0.308。一般情况下,微囊藻的Fv/Fm在0.3左右,当Fv/Fm过低表明藻类受到环境胁迫,PSII中心受到损伤进而降低光合作用效率。由表1可以看出,当水杨酸浓度大于20 mg·L−1时会对微囊藻的光合作用产生明显抑制,当各组酚酸浓度超过60 mg·L−1时,藻类基本停止了光合作用,这和前文中藻类的生物量变化和存活率相吻合。表2是各个组叶绿素含量的均值,数据表明:各组中相对低浓度的酚酸,不仅对藻类数量的增长有促进作用,也促进了叶绿素含量的增加。
表 1 铜绿微囊藻的Fv/FmTable 1. Fv/Fm of Microcystis aeruginosa对羟基苯甲酸P-hydroxybenzoic acid 对香豆酸P-coumaric acid 水杨酸Salicylic acid 咖啡酸Caffeic acid 010×10−620×10−640×10−660×10−680×10−6 0.3080.3100.2920.3020.0390.000 0.3080.3100.3140.3240.0490.000 0.3080.3090.0230.0140.0000.000 0.3080.3310.2760.3600.0380.000 注:0—80×10−6分别对应添加的4种酚酸浓度,表中数据是在微囊藻培养期结束时测得的Fv/Fm。 Note: 0—80×10−6 respectively correspond to the four added phenolic acid concentrations. The data in the table are the Fv/Fm measured at the end of the Microcystis culture period. | Show Table DownLoad:
CSV
表 2 铜绿微囊藻叶绿素含量(g·L-1)Table 2. Chlorophyll content of Microcystis aeruginosa
DownLoad:
CSV
表 2 铜绿微囊藻叶绿素含量(g·L-1)Table 2. Chlorophyll content of Microcystis aeruginosa对羟基苯甲酸P-hydroxybenzoic acid 对香豆酸P-coumaric acid 水杨酸Salicylic acid 咖啡酸Caffeic acid 010×10−620×10−640×10−660×10−680×10−6 0.5681.0141.1380.7750.0000.000 0.5680.7370.8620.5680.0000.000 0.5680.7960.0090.0000.0000.000 0.5682.0932.2011.3590.0000.000 注:0—80×10−6分别对应添加的4种酚酸浓度,表中数据是微囊藻培养期结束时测得的叶绿素含量。 Note: 0—80×10−6 respectively correspond to the four phenolic acid concentrations added. The data in the table is the chlorophyll content measured at the end of the Microcystis culture period. | Show TableDownLoad:
CSV
2.2 酚络合态铁的生物可利用性
根据上述试验结果可以得出,当这4种典型泥炭源酚酸的浓度达10 mg·L−1时,从生物量、存活率以及光合作用强度来说对微囊藻的生长没有明显抑制作用。普遍认为,藻类吸收Fe主要是离子形态,而不是有机络合态Fe[24]。目前,仅观察到在产生铁载体的微生物中存在铁载体复合物的吸收及其在细胞中的还原[25]。已有研究发现,有机络合态铁中铁的释放途径不同。其中包括简单的配体-金属平衡(Ligand-Metal balance),平衡在初级生产者消耗铁后发生变化,促进铁从络合物中分离。另一个途径是基于配体的降解,这也导致了Fe和配体的分离。第三种可能是通过有机态铁的还原,降低配体对铁的亲和力,并导致Fe的释放。这个过程可能是由于络合物的自还原/氧化而发生的,这意味着配体氧化同时也释放产生Fe[26]。尤其是光还原分解对铁的生物利用度有很大影响,这一释放铁途径被称为AHS(aquatic humic substances)机制[12]。
图3是加入不同酸和Fe2+的微囊藻生长曲线。施加和不加Fe2+的对照试验表明,在培养1—9 d,两组生长速度以及生物量大致相同。在第9天后,施加Fe2+组的生长速度放缓,最终的藻密度低于对照组。图3表明在没有配体存在的情况下,加入一定量的铁对微囊藻生长促进作用不明显。同时,观察发现,在藻类指数生长阶段出现了较高的pH值(高达10.5),这与二氧化碳生物需求增加有关。在指数生长结束和平台期开始后,pH值略下降。在藻类生长平缓或生长不良的样品中,pH值无显著变化,pH值大多保持在6—8。
能促进藻类生长的酚酸应当与三价铁有较高的亲和力,与二价铁有较低的亲和力[27]。试验表明,相对其他3种酚酸铁配合物体系,水杨酸铁不能有效地为微囊藻提供生物可利用性铁,这可能与水杨酸的稳定性较强有关[27]。对照表明,用咖啡酸处理的微囊藻生长良好,最高浓度达到2.19×107 cell·mL−1。这可能是咖啡酸中的儿茶酚基结构所引起的,并且有更高的氧化还原潜力,更容易将Fe从配合物中释放。Santana等 [28]的研究也证实了在生物条件下还原络合物的可能性。总体上,酚酸的加入提高了藻类对Fe的生物可利用率。
图2显示藻类存活率从第10天开始明显下降,藻类计数可能包括了死藻,因此选择第11天数据进行显著性分析。结果表明(表3),在0.05的置信水平下,咖啡酸的加入对微囊藻生长有显著促进作用,而水杨酸在0.05的置信水平下,对微囊藻生长无显著促进作用。
 图 3 微囊藻生长曲线Figure 3. Microcystis growth curveA.水杨酸+硫酸亚铁 B.咖啡酸+硫酸亚铁 C.水杨酸+草酸+硫酸亚铁 D.水杨酸+乙酸+硫酸亚铁 E.咖啡酸+草酸+硫酸亚铁 F.咖啡酸+乙酸+硫酸亚铁 G.水杨酸+柠檬酸+硫酸亚铁 H.咖啡酸+柠檬酸+硫酸亚铁 1_3.不添加酸和铁的对照组 Fe1_3硫酸亚铁A. Salicylic acid+Ferrous sulfate B. Caffeic acid+Ferrous sulfate C. Salicylic acid+Oxalic acid+Ferrous sulfate D. Salicylic acid+Acetic acid+Ferrous sulfate E. Caffeic acid+Oxalic acid+Ferrous sulfate F. Caffeic acid+Acetic acid+Ferrous sulfate G. Salicylic acid+Citric acid+Ferrous sulfate H. Caffeic acid+Citric acid+Ferrous sulfate 1_3. Control group without acid and iron Fe1_3. Ferrous sulfate表 3 第11天不同试验组微囊藻浓度变化的相关性矩阵Table 3. Correlation matrix of changes in the concentration of Microcystis in different test groups on the 11th day
图 3 微囊藻生长曲线Figure 3. Microcystis growth curveA.水杨酸+硫酸亚铁 B.咖啡酸+硫酸亚铁 C.水杨酸+草酸+硫酸亚铁 D.水杨酸+乙酸+硫酸亚铁 E.咖啡酸+草酸+硫酸亚铁 F.咖啡酸+乙酸+硫酸亚铁 G.水杨酸+柠檬酸+硫酸亚铁 H.咖啡酸+柠檬酸+硫酸亚铁 1_3.不添加酸和铁的对照组 Fe1_3硫酸亚铁A. Salicylic acid+Ferrous sulfate B. Caffeic acid+Ferrous sulfate C. Salicylic acid+Oxalic acid+Ferrous sulfate D. Salicylic acid+Acetic acid+Ferrous sulfate E. Caffeic acid+Oxalic acid+Ferrous sulfate F. Caffeic acid+Acetic acid+Ferrous sulfate G. Salicylic acid+Citric acid+Ferrous sulfate H. Caffeic acid+Citric acid+Ferrous sulfate 1_3. Control group without acid and iron Fe1_3. Ferrous sulfate表 3 第11天不同试验组微囊藻浓度变化的相关性矩阵Table 3. Correlation matrix of changes in the concentration of Microcystis in different test groups on the 11th day1_3 Fe1_3 A B C D E F G H 1_3 1 0.963 0.971 −0.358 0.474 0.657 0.246 0.461 0.983 0.491 Fe1_3 0.963 1.00 1.000* −0.095 0.221 0.835 −0.022 0.682 0.996 0.707 A 0.971 1.000* 1.00 −0.126 0.251 0.818 0.008 0.659 0.999* 0.685 B −0.358 −0.095 −0.126 1.00 −0.992 0.468 −0.993 0.663 −0.179 0.637 C 0.474 0.221 0.251 −0.992 1.00 −0.352 0.970 −0.563 0.302 −0.534 D 0.657 0.835 0.818 0.468 −0.352 1.00 −0.569 0.972 0.785 0.979 E 0.246 −0.022 0.008 −0.993 0.970 −0.569 1.00 −0.746 0.062 −0.723 F 0.461 0.682 0.659 0.663 0.663 0.972 −0.746 1.00 0.618 0.999* G 0.983 0.996 0.999* −0.179 0.302 0.785 0.062 0.618 1.00 0.645 H 0.491 0.707 0.685 0.637 −0.534 0.979 −0.723 0.999* 0.645 1.00 注:*. 在 0.05 水平(双侧)上显著相关。Notes:*. Significant correlation at 0.05(bilateral) level. | Show TableDownLoad:
CSV
2.3 泥炭沼泽源DOM-Fe的藻类可利用性
许多泥炭源有机质同时含有酚羟基和羧基,具有酚酸性质。但由于泥炭有机质组成复杂多样,现有技术尚不能有效分离不同性质有机化合物。因此,利用不同分子量段有机质与铁的络合物,开展藻类培养试验有助于客观评估泥炭源DOM-Fe的藻类可利用性。利用不同分子量段DOM-Fe,进行培养试验,铜绿微囊藻的生长情况如图4。表4 结果表明,在0.05 的置信水平下,不同组之间差异不具统计显著性,但图4 还是反映出有机态铁对藻类生长的促进趋势.
结果显示(图3和图4),不同分子量结合态Fe均促进了铜绿微囊藻的生长,但影响程度不同。添加Fe后,铜绿微囊藻的生物量和控制组相比均有增加,微囊藻生长得到促进,最终达到107 cell·mL−1。其中,微囊藻在7—11 d增长最快;其次,对比生长终点可以发现不同DOM-Fe促进效果存在差异:E组>D组>C组>B组>A组>无Fe组;第三,添加DOM显著促进了藻的生长。研究表明,相对于Fe3+,藻类更倾向于利用Fe2+[29]。这是由于具有一定还原能力的DOM可以减缓二价铁的氧化[30],从而提高了藻类对铁的利用率。此外,不同分子量段DOM与Fe的络合稳定常数略有不同,较高分子量的DOM(>3 kD)与Fe的络合稳定常数较小[30],在光照或者其他条件下容易发生解离,产生易被藻类利用的Fe。而泥炭源低分子量DOM(<1 KD)络合态铁,由于其络合稳定的常数相对较高,在培养体系中更加稳定,相对不易被藻类利用。
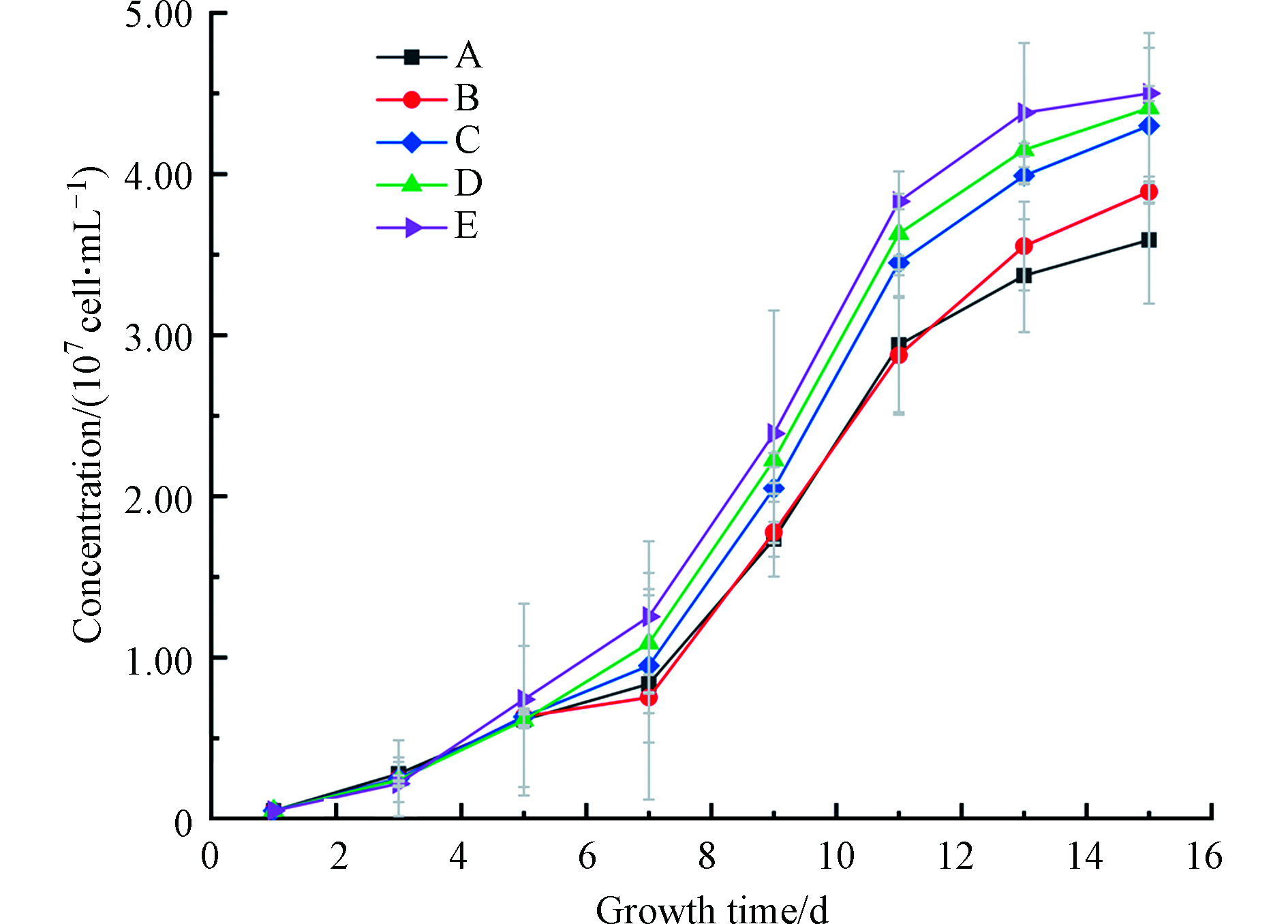 图 4 铜绿微囊藻生长曲线A.无添加 B.硫酸亚铁 C.DOM-Fe(<1KD ) D.DOM-Fe(1-3 KD) E.DOM-Fe(> 3KD)Figure 4. Microcystis aeruginosa growth curveA. No addition B. Ferrous sulfate C.DOM-Fe(<1KD) D.DOM-Fe(1-3KD) E.DOM-Fe(>3KD)表 4 第11天不同试验组铜绿微囊藻生长浓度变化的相关性矩阵Table 4. Correlation matrix of growth concentration changes of Microcystis aeruginosa in different test groups on the 11th day
图 4 铜绿微囊藻生长曲线A.无添加 B.硫酸亚铁 C.DOM-Fe(<1KD ) D.DOM-Fe(1-3 KD) E.DOM-Fe(> 3KD)Figure 4. Microcystis aeruginosa growth curveA. No addition B. Ferrous sulfate C.DOM-Fe(<1KD) D.DOM-Fe(1-3KD) E.DOM-Fe(>3KD)表 4 第11天不同试验组铜绿微囊藻生长浓度变化的相关性矩阵Table 4. Correlation matrix of growth concentration changes of Microcystis aeruginosa in different test groups on the 11th dayA B C D E A 1 −.751 .350 −.167 .770 B −.751 1 −.881 .776 −.158 C .350 −.881 1 −.982 −.327 D −.167 .776 −.982 1 .500 E .770 −.158 −.327 .500 1 注:*. 在 0.05 水平(双侧)上显著相关. Notes:*. Significant correlation at 0.05(bilateral) level. | Show TableDownLoad:
CSV
3. 结论(Conclusion)
(1)4种酚酸对藻类生长的影响均呈现“低促高抑”的规律。从藻类生物量和叶绿素含量来看,抑藻效果从高到低:水杨酸>对羟基苯甲酸>对香豆酸>咖啡酸;结合藻类的存活率,虽然低浓度酚酸刺激了藻类生物量的增长,但是也对藻类的生存产生了一定的负面影响:在添加10 mg·L−1酚酸的几组样品中,微囊藻的存活率都略低于控制组。
(2)添加酚酸的藻类样品中,当水杨酸浓度达到20 mg·L−1时,Fv/Fm明显降低(0.3降低到0.02左右),而其它3种酚酸浓度达到60 mg·L−1才出现抑制,说明水杨酸抑制作用最强。
(3)不同酚铁络合物的生物可利用性存在差异:相对咖啡酸和水杨酸,水杨酸络合态铁更难被藻类利用,除酚毒性效应外,还与其较高的络合稳定性有关。
(4)泥炭源不同分子段DOM-Fe对藻类生长的促进作用从高到低依次为:>3 KD,1—3 KD,<1 KD。高分子段DOM(>3 kD)与Fe的络合稳定常数最小,在光照或者其他条件下容易发生解离,更易释放Fe而被藻类利用;泥炭源低分子量DOM(<1 KD)络合态铁,因其络合稳定常数相对较高,相对不易被藻类利用。
致谢:感谢中国科学院水生生物研究所的大力支持。
-
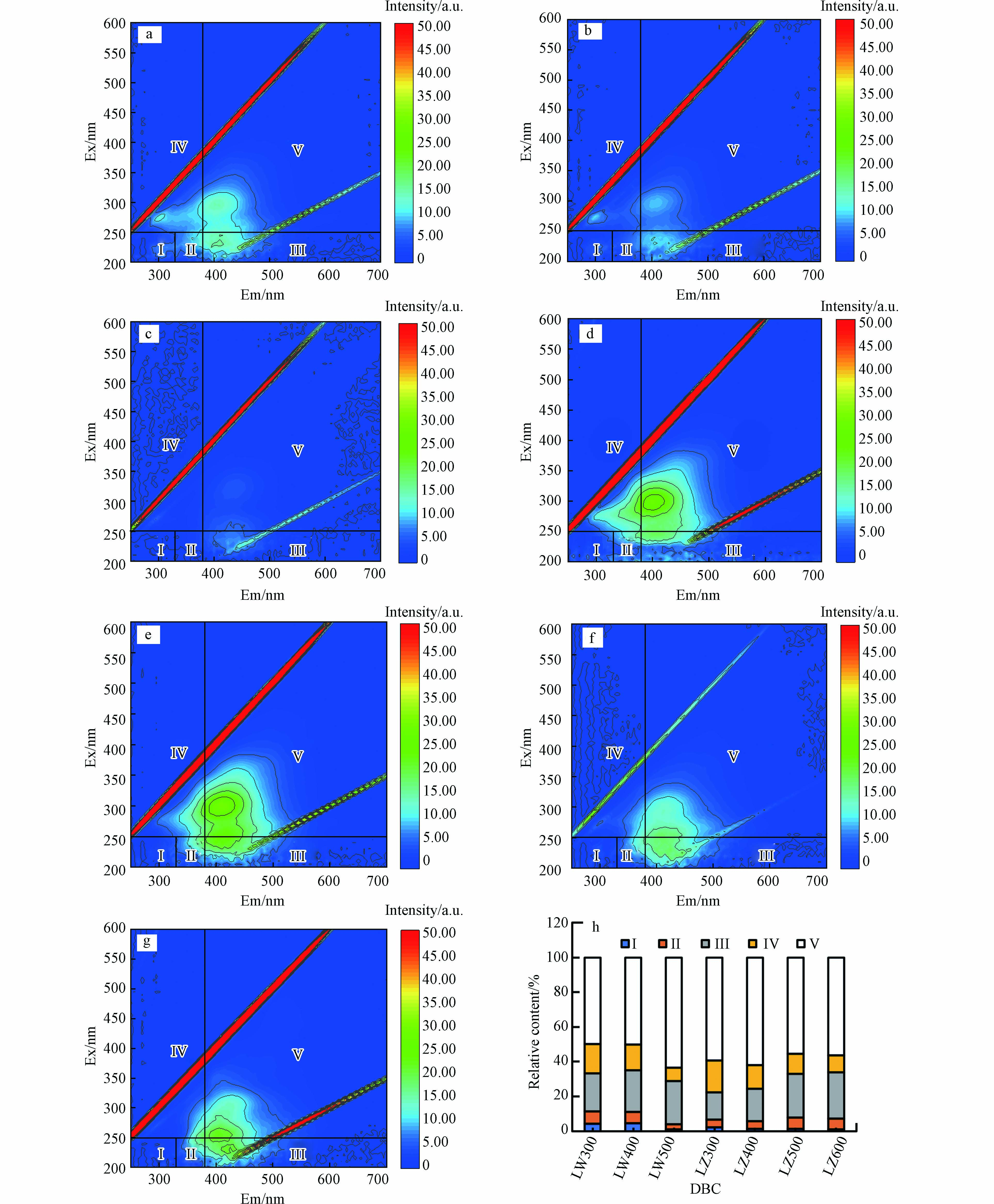
图 2 DBC的荧光组分及相对含量
Figure 2. Fluorescence components and their relative contents in DBC
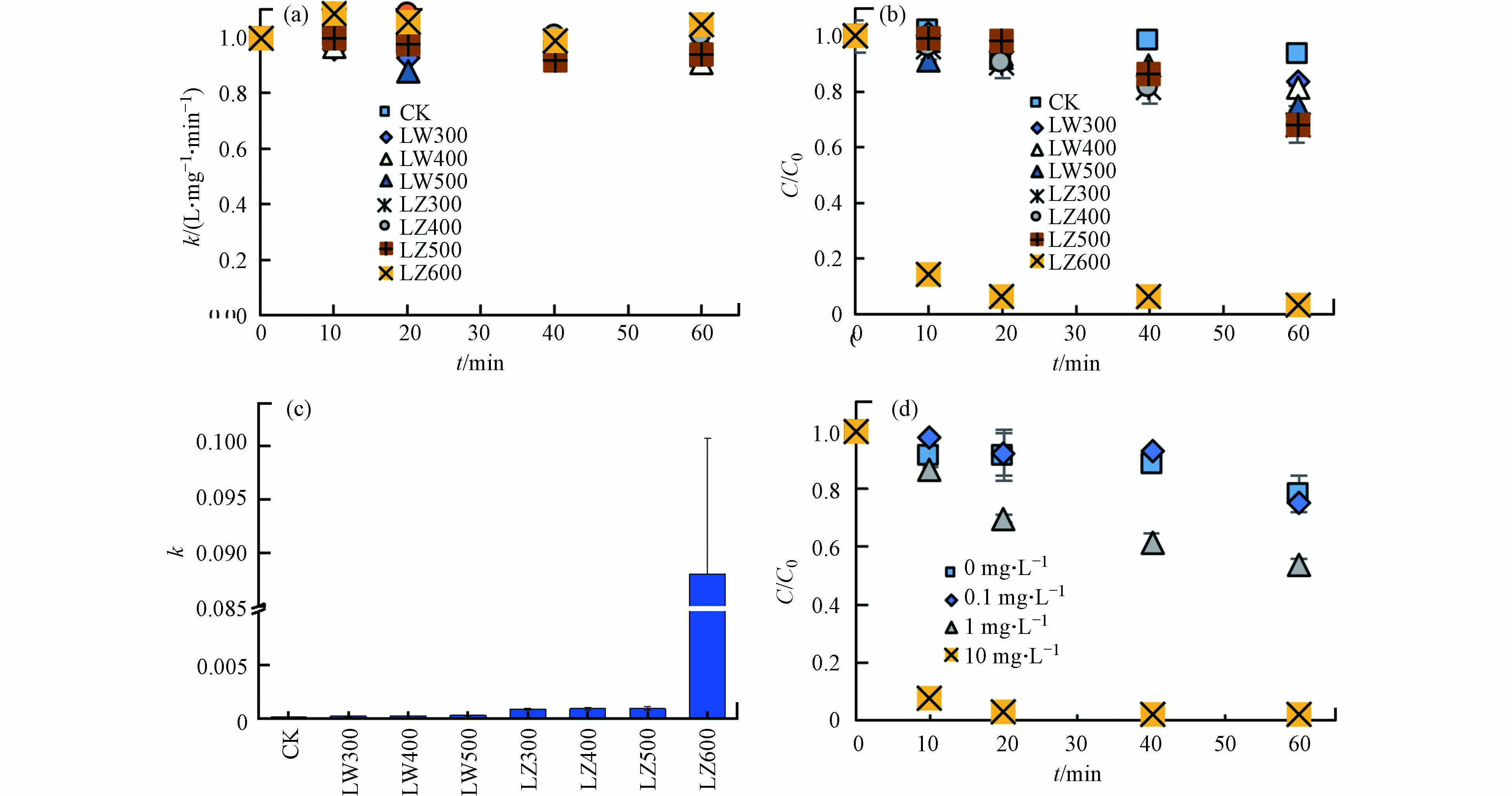
图 3 光照和黑暗条件下不同DBC和不同浓度的LZ600对TC降解的影响
Figure 3. Effects of DBC and LZ600 concentrations on TC photodegradation under light and dark conditions
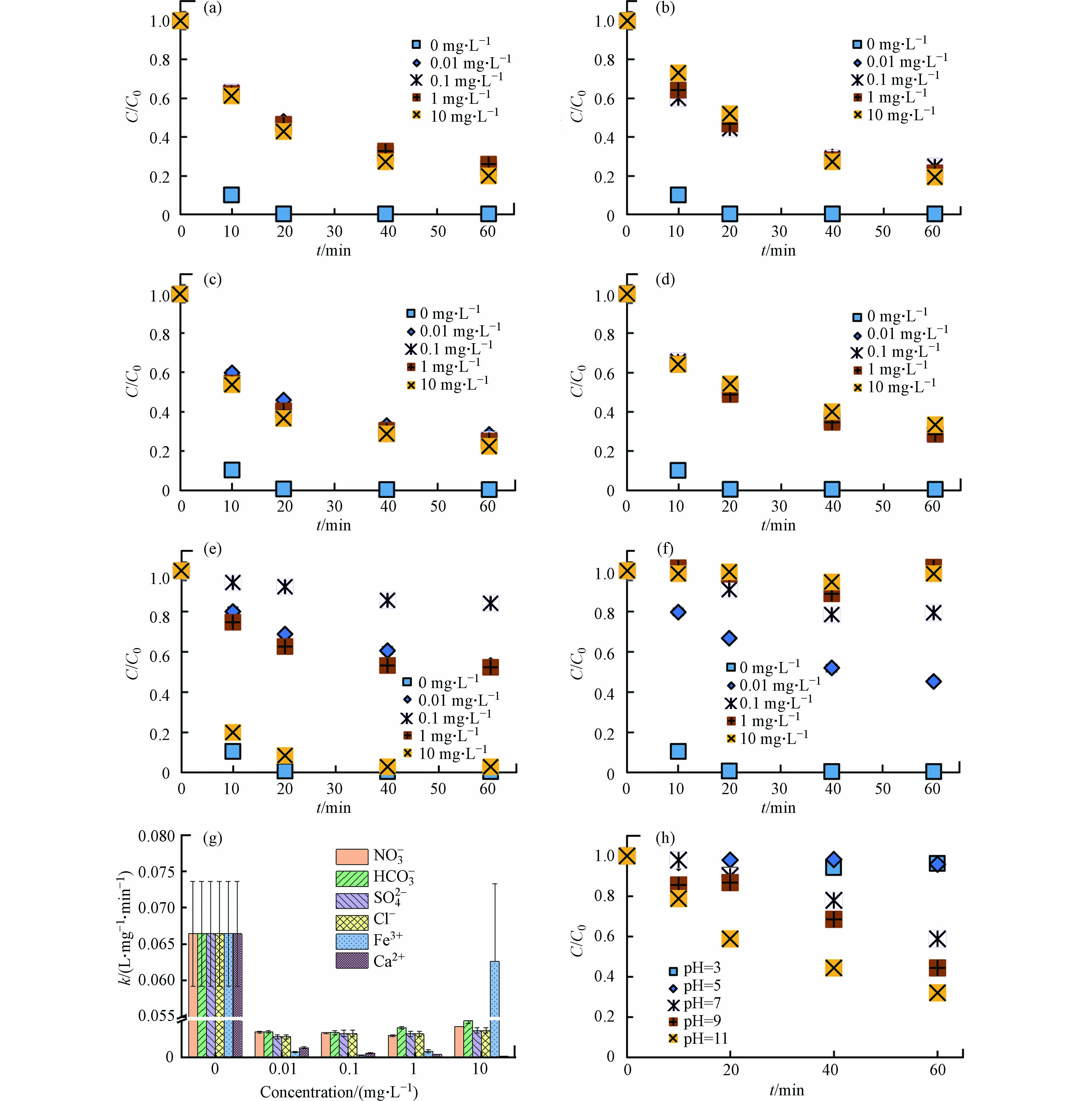
图 4 不同离子和pH条件下LZ600对TC光降解的影响
Figure 4. Effect of LZ600 on TC photodegradation under different ambient ions and pH
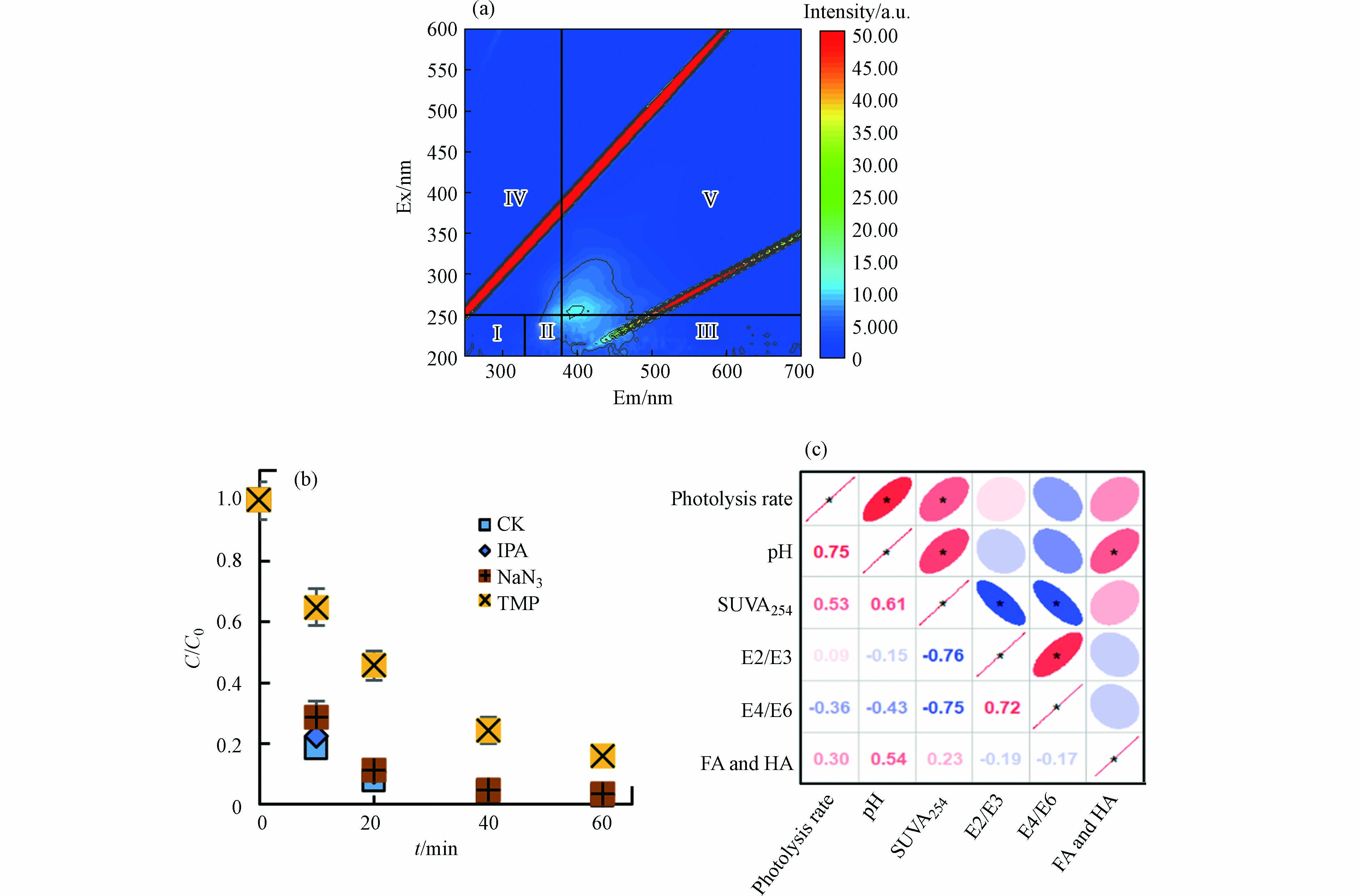
图 5 光降解对DBC性质及猝灭剂对TC光解的影响
Figure 5. Effect of photolysis on LZ600 and quenching experiment
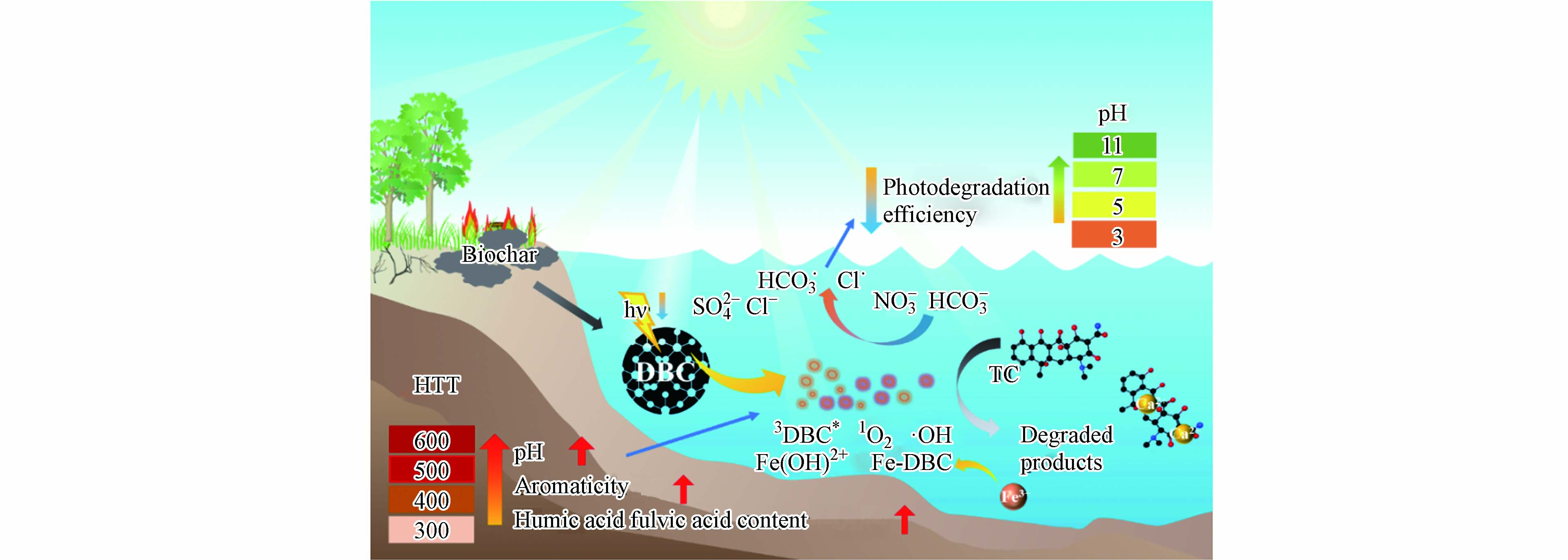
图 6 DBC影响TC降解的可能机制
Figure 6. Potential mechanisms of DBC promoting TC photodegradation
CAS 分子式Molecular formula 溶解度Solubility pKa 结构式Structural formula 64-75-5 C22H25ClN2O8 19.7 g·L−1(25 ℃) pKa1=3.30pKa2=7.68pKa3=9.68 </td><td class="table_top_border2 table_bottom_border" align="center" valign="middle">p<i>K</i><sub>a1</sub>=3.30p<i>K</i><sub>a2</sub>=7.68p<i>K</i><sub>a3</sub>=9.68</td><td class="table_top_border2 table_bottom_border" style="padding-bottom:2pt;padding-top:2pt;" align="center" valign="middle"><styled-content style="width:6cm"><img class="graphic" src="data:image/svg+xml,<svg xmlns='http://www.w3.org/2000/svg' width='350' height='600'><foreignObject width='2000' height='100%'><div xmlns='http://www.w3.org/1999/xhtml' style='font-size:16px;'><table>
<thead><tr><td class="table_top_border" align="center" valign="middle">CAS</td><td class="table_top_border" align="center" valign="middle">分子式Molecular formula</td><td class="table_top_border" align="center" valign="middle">溶解度Solubility</td><td class="table_top_border" align="center" valign="middle">p<i>K</i><sub>a</sub></td><td class="table_top_border" align="center" valign="middle">结构式Structural formula</td></tr></thead>
<tbody><tr><td class="table_top_border2 table_bottom_border" align="center" valign="middle">64-75-5</td><td class="table_top_border2 table_bottom_border" align="center" valign="middle">C<sub>22</sub>H<sub>25</sub>ClN<sub>2</sub>O<sub>8</sub></td><td class="table_top_border2 table_bottom_border" align="center" valign="middle">19.7 g·L<sup>−1</sup>(25 ℃)</td><td class="table_top_border2 table_bottom_border" align="center" valign="middle">p<i>K</i><sub>a1</sub>=3.30p<i>K</i><sub>a2</sub>=7.68p<i>K</i><sub>a3</sub>=9.68</td><td class="table_top_border2 table_bottom_border" style="padding-bottom:2pt;padding-top:2pt;" align="center" valign="middle"><styled-content style="width:6cm"><img class="graphic" src="2021122001-t1-1.jpg"></styled-content></td></tr></tbody>
</table></div></foreignObject></svg>"></styled-content></td></tr></tbody>
</table></div></foreignObject></svg>) 下载: 导出CSV
下载: 导出CSV
表 2 DBC的基本物理化学性质
Table 2. Basic physicochemical properties of DBC
DBC TOC/(mg·L−1) pH SUVA254/L·(mgC·m-1)−1 E2/E3 E4/E6 LW300 123 4.25 0.18 3.92 7.17 LW400 152 4.74 0.12 5.01 5.82 LW500 98.9 5.96 0.16 3.67 3.88 LZ300 520 5.66 0.06 6.23 5.48 LZ400 832 5.77 0.02 7.28 15.8 LZ500 451 7.75 0.17 3.81 5.09 LZ600 101 10.2 0.27 4.61 2.00 注:SUVA254为100倍的254 nm处的吸光度和DBC TOC的比值,E4/E6为465 nm处吸光度和665 nm处吸光度的比值,E2/E3为254 nm处吸光度和365 nm处吸光度的比值. Note: SUVA254 is the ratio of 100 times the absorbance at 254 nm to DBC TOC, E4/E6 is the ratio of the absorbance at 465 nm to 665 nm, E2/E3 is the ratio of the absorbance at 254 nm to 365 nm.
下载: 导出CSV
-
[1] WANG L W, O'CONNOR D, RINKLEBE J, et al. Biochar aging: Mechanisms, physicochemical changes, assessment, and implications for field applications [J]. Environmental Science & Technology, 2020, 54(23): 14797-14814. [2] LEHMANN J, COWIE A, MASIELLO C, et al. Biochar in climate change mitigation [J]. Nature Geoscience, 2021, 14(12): 883-892. doi: 10.1038/s41561-021-00852-8 [3] TOMCZYK A, SOKOŁOWSKA Z, BOGUTA P. Biochar physicochemical properties: Pyrolysis temperature and feedstock kind effects [J]. Reviews in Environmental Science and Bio/Technology, 2020, 19(1): 191-215. doi: 10.1007/s11157-020-09523-3 [4] HAGEMANN N, JOSEPH S, SCHMIDT H P, et al. Organic coating on biochar explains its nutrient retention and stimulation of soil fertility [J]. Nature Communications, 2017, 8: 1089. doi: 10.1038/s41467-017-01123-0 [5] IPPOLITO J A, CUI L Q, KAMMANN C, et al. Feedstock choice, pyrolysis temperature and type influence biochar characteristics: A comprehensive meta-data analysis review [J]. Biochar, 2020, 2(4): 421-438. doi: 10.1007/s42773-020-00067-x [6] SONG F H, LI T T, SHI Q, et al. Novel insights into the molecular-level mechanism linking the chemical diversity and copper binding heterogeneity of biochar-derived dissolved black carbon and dissolved organic matter [J]. Environmental Science & Technology, 2021, 55(17): 11624-11636. [7] WAGNER S, JAFFÉ R, STUBBINS A. Dissolved black carbon in aquatic ecosystems [J]. Limnology and Oceanography Letters, 2018, 3(3): 168-185. doi: 10.1002/lol2.10076 [8] 彭红波, 杨东, 高鹏, 等. 生物炭中溶解性炭黑的释放及环境效应 [J]. 材料导报, 2020, 34(11): 11029-11034. doi: 10.11896/cldb.19050149 PENG H B, YANG D, GAO P, et al. Releasing and the environmental implications of dissolved black carbon from biochars [J]. Materials Reports, 2020, 34(11): 11029-11034(in Chinese). doi: 10.11896/cldb.19050149
[9] 魏晨辉, 付翯云, 瞿晓磊, 等. 溶解态黑碳的环境过程研究 [J]. 化学进展, 2017, 29(9): 1042-1052. doi: 10.7536/PC170444 WEI C H, FU Y F, QU X L, et al. Environmental processes of dissolved black carbon [J]. Progress in Chemistry, 2017, 29(9): 1042-1052(in Chinese). doi: 10.7536/PC170444
[10] ZHOU Z C, CHEN B N, QU X L, et al. Dissolved black carbon as an efficient sensitizer in the photochemical transformation of 17β-estradiol in aqueous solution [J]. Environmental Science & Technology, 2018, 52(18): 10391-10399. [11] FU H Y, WEI C H, QU X L, et al. Strong binding of apolar hydrophobic organic contaminants by dissolved black carbon released from biochar: A mechanism of pseudomicelle partition and environmental implications [J]. Environmental Pollution, 2018, 232: 402-410. doi: 10.1016/j.envpol.2017.09.053 [12] ZHANG P, SHAO Y F, XU X J, et al. Phototransformation of biochar-derived dissolved organic matter and the effects on photodegradation of imidacloprid in aqueous solution under ultraviolet light [J]. Science of the Total Environment, 2020, 724: 137913. doi: 10.1016/j.scitotenv.2020.137913 [13] LI S, HU J Y. Photolytic and photocatalytic degradation of tetracycline: Effect of humic acid on degradation kinetics and mechanisms [J]. Journal of Hazardous Materials, 2016, 318: 134-144. doi: 10.1016/j.jhazmat.2016.05.100 [14] BERTILSSON S, TRANVIK L J. Photochemical transformation of dissolved organic matter in lakes [J]. Limnology and Oceanography, 2000, 45(4): 753-762. doi: 10.4319/lo.2000.45.4.0753 [15] WAN D, WANG J, DIONYSIOU D D, et al. Photogeneration of reactive species from biochar-derived dissolved black carbon for the degradation of amine and phenolic pollutants [J]. Environmental Science & Technology, 2021, 55(13): 8866-8876. [16] FANG G D, LIU C, WANG Y J, et al. Photogeneration of reactive oxygen species from biochar suspension for diethyl phthalate degradation [J]. Applied Catalysis B:Environmental, 2017, 214: 34-45. doi: 10.1016/j.apcatb.2017.05.036 [17] 王佳钰, 王中钰, 陈景文, 等. 环境新污染物治理与化学品环境风险防控的系统工程 [J]. 科学通报, 2022, 67(3): 267-277. doi: 10.1360/TB-2021-0422 WANG J Y, WANG Z Y, CHEN J W, et al. Environmental systems engineering consideration on treatment of emerging pollutants and risk prevention and control of chemicals [J]. Chinese Science Bulletin, 2022, 67(3): 267-277(in Chinese). doi: 10.1360/TB-2021-0422
[18] van BOECKEL T P, PIRES J, SILVESTER R, et al. Global trends in antimicrobial resistance in animals in low- and middle-income countries [J]. Science, 2019, 365(6459): eaaw1944. doi: 10.1126/science.aaw1944 [19] XU L Y, ZHANG H, XIONG P, et al. Occurrence, fate, and risk assessment of typical tetracycline antibiotics in the aquatic environment: A review [J]. Science of the Total Environment, 2021, 753: 141975. doi: 10.1016/j.scitotenv.2020.141975 [20] WANG J H, LU J, WU J, et al. Proliferation of antibiotic resistance genes in coastal recirculating mariculture system [J]. Environmental Pollution, 2019, 248: 462-470. doi: 10.1016/j.envpol.2019.02.062 [21] SHAO S C, WU X W. Microbial degradation of tetracycline in the aquatic environment: A review [J]. Critical Reviews in Biotechnology, 2020, 40(7): 1010-1018. doi: 10.1080/07388551.2020.1805585 [22] 侯力睿, 傅榆涵, 赵冲, 等. 兽药抗生素对生态环境的混合毒性研究进展 [J]. 环境化学, 2021, 40(1): 55-64. doi: 10.7524/j.issn.0254-6108.2020052502 HOU L R, FU Y H, ZHAO C, et al. Advance on combined toxicity of veterinary antibiotics on ecological environments [J]. Environmental Chemistry, 2021, 40(1): 55-64(in Chinese). doi: 10.7524/j.issn.0254-6108.2020052502
[23] 廖洋, 鲁金凤, 曹轶群, 等. 光催化降解对抗生素藻类毒性效应影响研究进展 [J]. 环境化学, 2021, 40(1): 111-120. doi: 10.7524/j.issn.0254-6108.2019122404 LIAO Y, LU J F, CAO Y Q, et al. Research progress on the effects of photocatalytic degradation on the algae toxicity of antibiotics [J]. Environmental Chemistry, 2021, 40(1): 111-120(in Chinese). doi: 10.7524/j.issn.0254-6108.2019122404
[24] ESCOBAR-HUERFANO F, GÓMEZ-OLIVÁN L M, LUJA-MONDRAGÓN M, et al. Embryotoxic and teratogenic profile of tretracycline at environmentally relevant concentrations on Cyprinus carpio [J]. Chemosphere, 2020, 240: 124969. doi: 10.1016/j.chemosphere.2019.124969 [25] 徐冰洁, 罗义, 周启星, 等. 抗生素抗性基因在环境中的来源、传播扩散及生态风险 [J]. 环境化学, 2010, 29(2): 169-178. XU B J, LUO Y, ZHOU Q X, et al. Sources, dissemination, and ecological risks of antibiotic resistances genes(args) in the environment [J]. Environmental Chemistry, 2010, 29(2): 169-178(in Chinese).
[26] ZHU Y G, ZHAO Y, LI B, et al. Continental-scale pollution of estuaries with antibiotic resistance genes"> [J]. Nature Microbiology, 2017, 2: 16270. doi: 10.1038/nmicrobiol.2016.270 [27] ZHANG X, LI J, FAN W Y, et al. Enhanced photodegradation of extracellular antibiotic resistance genes by dissolved organic matter photosensitization [J]. Environmental Science & Technology, 2019, 53(18): 10732-10740. [28] SONG C, ZHANG K X, WANG X J, et al. Effects of natural organic matter on the photolysis of tetracycline in aquatic environment: Kinetics and mechanism [J]. Chemosphere, 2021, 263: 128338. doi: 10.1016/j.chemosphere.2020.128338 [29] ZHENG H, WANG Z Y, DENG X, et al. Characteristics and nutrient values of biochars produced from giant reed at different temperatures [J]. Bioresource Technology, 2013, 130: 463-471. doi: 10.1016/j.biortech.2012.12.044 [30] HAN L, NIE X, WEI J, et al. Effects of feedstock biopolymer compositions on the physiochemical characteristics of dissolved black carbon from lignocellulose-based biochar [J]. Science of the Total Environment, 2021, 751: 141491. doi: 10.1016/j.scitotenv.2020.141491 [31] WANG Y T, XIN Z B, PENG F, et al. Influence of pyrolysis temperature on characteristics and nitrobenzene adsorption capability of biochar derived from reed and giant reed [J]. Science of Advanced Materials, 2019, 11(11): 1523-1530. doi: 10.1166/sam.2019.3463 [32] ZHANG J X, LUO J, TONG D M, et al. The dependence of pyrolysis behavior on the crystal state of cellulose [J]. Carbohydrate Polymers, 2010, 79(1): 164-169. doi: 10.1016/j.carbpol.2009.07.038 [33] CAO J, JIANG J. Reducing capacities in continuously released low molecular weight fractions from bulk humic acids [J]. Journal of Environmental Management, 2019, 244: 172-179. doi: 10.1016/j.jenvman.2019.05.014 [34] CHEN B L, ZHOU D D, ZHU L Z. Transitional adsorption and partition of nonpolar and polar aromatic contaminants by biochars of pine needles with different pyrolytic temperatures [J]. Environmental Science & Technology, 2008, 42(14): 5137-5143. [35] XIAO X, CHEN B L, ZHU L Z. Transformation, morphology, and dissolution of silicon and carbon in rice straw-derived biochars under different pyrolytic temperatures [J]. Environmental Science & Technology, 2014, 48(6): 3411-3419. [36] 朱静, 张朝晖. 近海CDOM光反应变化及SPM对其影响的研究 [J]. 环境科学与技术, 2019, 42(10): 9-13. doi: 10.19672/j.cnki.1003-6504.2019.10.002 ZHU J, ZHANG Z H. Photoreaction changes of CDOM in offshore and study on effects of SPM on photoreaction [J]. Environmental Science & Technology, 2019, 42(10): 9-13(in Chinese). doi: 10.19672/j.cnki.1003-6504.2019.10.002
[37] BIANCO A, MINELLA M, de LAURENTIIS E, et al. Photochemical generation of photoactive compounds with fulvic-like and humic-like fluorescence in aqueous solution [J]. Chemosphere, 2014, 111: 529-536. doi: 10.1016/j.chemosphere.2014.04.035 [38] WU S M, ZUBER F, MANIURA-WEBER K, et al. Nanostructured surface topographies have an effect on bactericidal activity [J]. Journal of Nanobiotechnology, 2018, 16(1): 20. doi: 10.1186/s12951-018-0347-0 [39] GREBEL J E, PIGNATELLO J J, SONG W H, et al. Impact of halides on the photobleaching of dissolved organic matter [J]. Marine Chemistry, 2009, 115(1/2): 134-144. [40] MACK J, BOLTON J R. Photochemistry of nitrite and nitrate in aqueous solution: A review [J]. Journal of Photochemistry and Photobiology A:Chemistry, 1999, 128(1/2/3): 1-13. [41] ZHOU L, SLEIMAN M, FERRONATO C, et al. Reactivity of sulfate radicals with natural organic matters [J]. Environmental Chemistry Letters, 2017, 15(4): 733-737. doi: 10.1007/s10311-017-0646-y [42] OU Q, XU Y H, HE Q, et al. Deposition behavior of dissolved black carbon on representative surfaces: Role of molecular conformation [J]. Journal of Environmental Chemical Engineering, 2021, 9(5): 105921. doi: 10.1016/j.jece.2021.105921 [43] WAITE T D. Role of iron in light-induced environmental processes[M]//The Handbook of Environmental Chemistry. Berlin/Heidelberg: Springer-Verlag, : 255-298. [44] 孔婧, 邓芠, 李若白, 等. 光降解酮洛芬的动力学及影响因素 [J]. 环境化学, 2016, 35(12): 2568-2574. doi: 10.7524/j.issn.0254-6108.2016.12.2016050601 KONG J, DENG W, LI R B, et al. Photolysis of Ketoprofen in aqueous solution: Kinetics and influence of environmental factors [J]. Environmental Chemistry, 2016, 35(12): 2568-2574(in Chinese). doi: 10.7524/j.issn.0254-6108.2016.12.2016050601
[45] 刘纪阳, 薛爽, 张营, 等. 水相和冰相中不同pH条件下溶解性有机质对苊光降解的影响 [J]. 环境科学学报, 2021, 41(5): 1930-1939. doi: 10.13671/j.hjkxxb.2020.0400 LIU J Y, XUE S, ZHANG Y, et al. Effect of dissolved organic matter on photodegradation of acenaphthene under different pH conditions in water and ice [J]. Acta Scientiae Circumstantiae, 2021, 41(5): 1930-1939(in Chinese). doi: 10.13671/j.hjkxxb.2020.0400
[46] 李聪鹤, 车潇炜, 白莹, 等. 水体中磺胺甲噁唑间接光降解作用 [J]. 环境科学, 2019, 40(1): 273-280. doi: 10.13227/j.hjkx.201805014 LI C H, CHE X W, BAI Y, et al. Indirect photodegradation of sulfamethoxazole in water [J]. Environmental Science, 2019, 40(1): 273-280(in Chinese). doi: 10.13227/j.hjkx.201805014
[47] LÓPEZ-PEÑALVER J J, SÁNCHEZ-POLO M, GÓMEZ-PACHECO C V, et al. Photodegradation of tetracyclines in aqueous solution by using UV and UV/H2O2 oxidation processes [J]. Journal of Chemical Technology & Biotechnology, 2010, 85(10): 1325-1333. [48] MCNALLY A M, MOODY E C, MCNEILL K. Kinetics and mechanism of the sensitized photodegradation of lignin model compounds [J]. Photochemical & Photobiological Sciences, 2005, 4(3): 268. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 5880
- HTML全文浏览数: 5880
- PDF下载数: 197
- 施引文献: 0
