




全文HTML
随着工业化发展和化石能源的大量使用,大气中CO2浓度逐渐增加并使全球气候变暖。光合微藻作为新一代的生物能源,具有光合固碳效率高、生长周期短、逆境耐受性强和油脂含量高等优点而成为生物固碳与CO2减排技术的常用藻种[1-2]。
目前,关于微藻的研究大多是在悬浮培养模式下进行的,但在悬浮培养条件下,气泡对藻细胞生长扰动大,限制了藻生物量的生长,而光的传递和CO2传质受培养液影响大,生物量采收成本相对较高[3]。藻细胞的固定化培养能改善CO2的传质效率和光的传递效率,克服悬浮培养中气泡扰动的影响,有效降低微藻生物量的采收成本。目前,研究者们设计了多种微藻固定化培养的膜式光生物反应器,如平板式生物膜反应器和转筒式生物膜培养系统等[4-5]。在这些固定化细胞反应器中,皆采用气液混合的方式培养藻细胞,仍然存在气泡对藻细胞生物膜生长的扰动作用。基于此,本课题提出了一种基于气液分离的微藻吸附式生物膜培养系统,在该系统中利用选择透过性膜将气相与液相分开以解决气泡对藻培养液的扰动作用,从而促使藻细胞有效吸附在固体基质膜材料表面,形成一层微藻生物膜,并减少水的消耗、提高光的传递效率和微藻的光合效率。
光合微藻的生长除受培养基营养成分影响外,还受多种条件如光照强度、气体流速、液体流速等参数影响[6]。微藻细胞生长过程中,光能为藻生物膜提供固定CO2的能量,通过光合作用将光能转化为储藏在有机物中的化学能。因此,光照强度显著地影响微藻细胞光合效率及其微藻生物质产率,并且对微藻细胞内的生化组分及含量产生较大影响[7]。
本文拟利用所设计的气液分离膜生物反应器开展光照强度对小球藻生物膜生长与代谢的影响研究,以评价小球藻在该系统中的生物量生长、油脂产率等特性,为探索藻细胞固定化培养和促进生物能转化提供一定的实验和理论指导。
1 实验部分
1.1 藻种及培养条件
实验藻种为购自中国科学院武汉水生生物所的蛋白核小球藻(C. pyrenoidosa FACHB-9)。实验采用培养基配方为:NaNO3 1.5 g·L-1, K2HPO4·3H2O 40 mg·L-1, MgSO4·7H2O 75 mg·L-1, CaCl2·2H2O 36 mg·L-1, 柠檬酸6 mg·L-1, EDTA 1 mg·L-1, Na2CO3 20 mg·L-1, H3BO3 2.86 mg·L-1, MnCl2·H2O 1.81 mg·L-1, ZnSO4·7H2O 0.222 mg·L-1, CuSO4·5H2O 0.079 mg·L-1, Na2MoO4 0.39 mg·L-1, Co·6H2O·2HNO3 0.049 mg·L-1。121 ℃下高压灭菌20 min,备用。将小球藻藻种接入到含有培养基的三角瓶中,150 r·min-1、30 ℃摇床培养72 h,获得藻种子液。然后将种子液按10%的接种比例接种到实验系统的储液瓶中,通过蠕动泵将种子液泵入气液分离的膜生物反应器液室侧进行藻细胞吸附培养。实验中配制含3%(体积分数)的CO2混合气体并通入反应器气室中,气体通过渗透膜进入液室,供液室侧藻细胞进行光合作用。实验中分别研究5种光照条件:23、46、92、138和230 μmol·(m2·s)-1下藻生物膜生长与代谢特性,每个参数进行3次重复实验,实验结果取平均值进行计算。
1.2 实验系统的搭建
如图 1所示,气液分离的膜式光反应器由上部的液体腔室(液室)和下部的气体腔室(气室)组成,2个腔室大小均为16 cm(长)×4 cm(宽)×0.8 cm(高),上下腔室间由具有疏水性的选择透过性PTFE膜隔开,并用有机玻璃网格支撑PTFE膜。在液室和气室的两侧分别开有8 mm的螺纹孔作为培养基和气体进出口。反应器用螺栓进行固定,密封。整个反应器水平放置,液室在上,气室在下。
图1 实验系统示意图
Fig. 1 Schematic of cultivation system
Fig. 1 Schematic of cultivation system
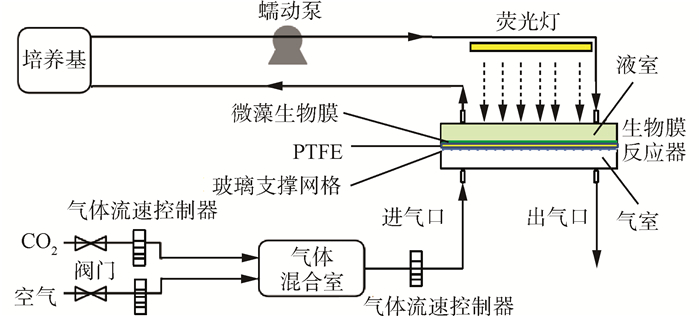
反应器系统用1 mol·L-1的氢氧化钠溶液清洗消毒杀菌,并用灭菌后的蒸馏水冲洗3次。整个实验系统包括液路循环系统和气路系统,液路循环系统依靠蠕动泵作为推动力,使藻细胞悬浮液不断地在储液瓶和反应器液室中循环流动,其中循环液中小球藻细胞在重力作用下可以有效地吸附在液室侧PTFE膜表面上,从而实现藻生物膜的挂膜和生长。气路系统由CO2钢瓶和空气泵组成,配制成具有一定浓度的CO2混合气体,并使用气体质量流量控制器控制所有气体的流量。
实验分为2个阶段:藻细胞的覆膜和生物膜生长。覆膜阶段,在蠕动泵的驱动下藻液以6 mL·min-1的流速流入液室,小球藻细胞吸附在膜材料的疏水性表面,直至膜材料表面被藻细胞覆盖即表示覆膜成功(24 h)。之后排出循环液,更换相同体积的新鲜培养液,吸附在基底膜材料表面的藻细胞在光照条件下进行生长与繁殖,并形成具有稳定厚度的生物膜。实验中进入膜生物反应器气室中混合气体的流速为40 mL·min-1(供气压力30 kPa,其中CO2分压为0.9 kPa)。
1.3 检测与分析方法
实验中,将小球藻生物膜从PTFE膜上进行刮取并冲洗下来,收集在50 mL的离心管中,8 000 r·min-1离心后,去除上清液,收集生物量进行冷冻干燥至恒重,用电子天平(EL204, 上海)测定小球藻生物膜的质量。
使用pH计测定培养基pH值(pHs-3E, 上海)。采用紫外分光光度法(722N, 上海)检测培养基中NO3--N含量[8]。蛋白质含量通过Bradford法进行测量。
小球藻生物膜内总脂含量的测定方法为氯仿-甲醇法[9]:利用液氮研磨冷冻干燥后的藻粉,然后称取100 mg藻粉,加入4 mL甲醇、2 mL氯仿、1.6 mL水;85 W、20 kHz超声(HN/650Y,上海)15 min,重复3次;然后加入6 mL氯仿、6 mL水,静置分层,吸取下层液体,60 ℃烘至恒重,并称重。
淀粉检测方法[10]如下:取4 mL藻液离心,加入0.5 mL玻璃珠(0.2~0.5 mm)和0.25 mL去离子水,旋涡振荡4 min。加入4 mL体积分数为80%的乙醇,68 ℃提取15 min,10 000 r·min-1离心5 min。上清液用于叶绿素检测。在沉淀中加入3.3 mL体积分数为30%的高氯酸,25 ℃振荡15 min,离心,收集上清液,重复3次。收集3次提取液共10 mL。取0.5 mL提取液,加入2.5 mL硫酸蒽酮溶液(2 g蒽酮溶入1 L 72%硫酸)100 ℃水浴8 min,水浴冷却2 min, 625 nm测定吸光度。
叶绿素及类胡萝卜素含量的计算公式[11]如下:
| cclha=13.95A665−6.88A649 | (1) |
| cclhb=24.96A649−7.32A665 | (2) |
| ccar=1000A470−2.05cchla−114.8cchlb245 | (3) |
式中:cclha为叶绿素a的浓度,mg·L-1;cclhb为叶绿素b的浓度,mg·L-1;ccar为类胡萝卜素的浓度,mg·L-1。A665为上述上清液在665 nm处的吸光值;A649为上述上清液在649 nm处的吸光值;A470为上述上清液在470 nm处的吸光值。
实验中采用酸水解的方法测定小球藻中油脂的脂肪酸组分。称取20 mg小球藻加入到旋盖试管中,加入2.5 mL甲醇和50 μL的浓硫酸,80 ℃恒温水浴2.5 h。待小球藻消解完全后加入1 mL饱和NaCl溶液和1 mL正己烷,振荡混匀然后静置分层,上层溶液为溶有脂肪酸的正己烷,待混合液分层完毕后收集上清液,用装有氢火焰离子检测器的气相色谱仪进行脂肪酸组分分析(GC-2010,山东),色谱柱为DB-5毛细管(30 m×0.32 nm, 0.25 μm),进行分析检测时进样口温度为230 ℃,载气为氮气,程序升温如下:190 ℃恒温10 min,然后以20 ℃·min-1的速率升温至230 ℃,并保留20 min。待温度稳定后加入待测样品,进样量为7 μL。
2 结果与分析
2.1 光照强度对小球藻生物膜生长的影响
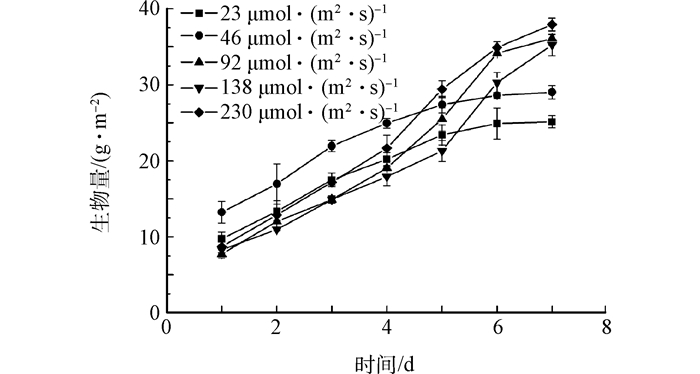
研究表明,光照强度是影响微藻生长繁殖的重要环境因素[12]。本实验也考察了光照强度对小球藻生物膜生长的影响,结果如图 2所示。当光照强度较低时(23和46 μmol·(m2·s)-1),在小球藻生长的前4 d,小球藻生物膜生物量高于其他光强条件下的生物量。但当培养到后期,在光照强度低于46 μmol·(m2·s)-1的条件下,小球藻生物膜的生长速率下降;而在光照强度为92、138和230 μmol·(m2·s)-1时,小球藻生物膜的生长速率增大。当光照强度为230 μmol·(m2·s)-1时,小球藻的生物膜生物量在第7天时达到了37.92 g·m-2。这是因为在小球藻生长初期,生物量较少,细胞之间没有相互遮挡,在光照强度低于46 μmol·(m2·s)-1的条件下,光为限制性条件,生物量和光照强度成正相关;相反,在光照强度高于46 μmol·(m2·s)-1时,超过了小球藻生物膜生长的光饱和点,对小球藻生长产生一定光抑制现象,导致生物膜生长较慢。随着培养时间延续,小球藻生物膜厚度逐渐增加,细胞之间的屏蔽效应增强,低光照强度条件下,光能不能满足生物膜下层小球藻细胞光合作用所需,影响了小球藻细胞的生长繁殖;而在光照强度较大时,小球藻生物膜中下层细胞能获得足够光能进行光合作用,表现为生物量显著增加。
图2 光照强度对小球藻生物膜生长的影响
Fig. 2 Effects of light intensity on growth of microalgae biofilm
Fig. 2 Effects of light intensity on growth of microalgae biofilm
2.2 光照强度对培养基中NO3--N的及pH的影响
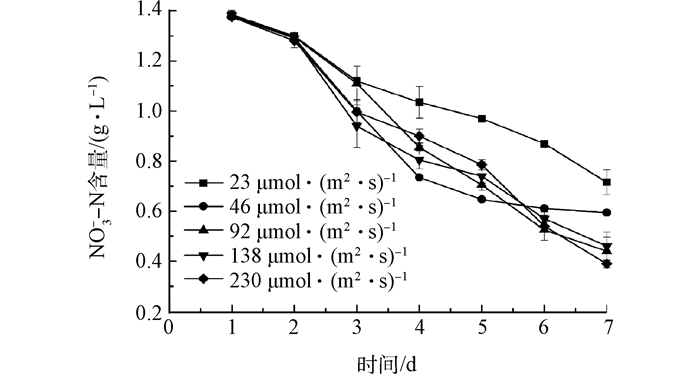
硝酸盐作为氮源对小球藻的生长代谢以及油脂积累具有重要影响[13]。光照条件影响下培养基中硝酸盐的利用情况如图 3所示。随着培养进行,培养基中硝酸盐含量呈明显下降趋势,硝酸盐的利用情况与反应器中小球藻生物膜生长具有一定相关性。在培养前期,所有实验条件下培养基中残余硝酸盐浓度下降较快,但随着培养时间延续,当光照强度为46 μmol·(m2·s)-1以下时,硝酸盐的利用呈明显减缓趋势,光照强度为23 μmol·(m2·s)-1时,培养液中NO3--N在微藻生物膜生长的整个阶段消耗较为平缓。当光照强度为230 μmol·(m2·s)-1时,其硝酸盐利用率最大,达到71.7%。结果表明,随着光照强度增加,硝酸盐消耗量逐渐增加,并与小球藻生长具有明显的正相关性,因此,氮源的消耗主要用于生物膜生物量的生长。
图3 光照强度对培养基中NO3--N利用的影响
Fig. 3 Effect of light intensities on utilization of NO3--N in medium
Fig. 3 Effect of light intensities on utilization of NO3--N in medium
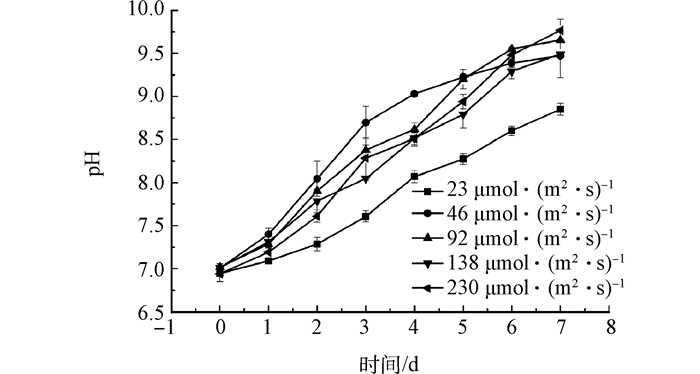
光照射条件下,微藻生物膜细胞吸收CO2进行光合作用,因固碳速率受光照强度影响较大,并进而影响培养液中pH值变化,而微藻细胞内碳酸酐酶和核酮糖-1, 5-二磷酸羧化酶等相关酶的活性受pH影响,并对微藻细胞的生长代谢产生影响[14]。因此,实验调查了光照强度影响下培养基中pH值变化,结果如图 4所示。所有实验条件下,培养基pH值均逐渐上升,这归咎于微藻生物膜生长过程中对CO2的不断消耗。研究还发现,随光照强度增加,培养液中最终pH值基本呈逐渐递增趋势,这也表明随光照强度递增,微藻生物膜固定CO2的速率呈递增趋势。
图4 光照强度对培养pH的影响
Fig. 4 Effect of light intensities on pH
Fig. 4 Effect of light intensities on pH
2.3 光照强度对小球藻细胞组分的影响
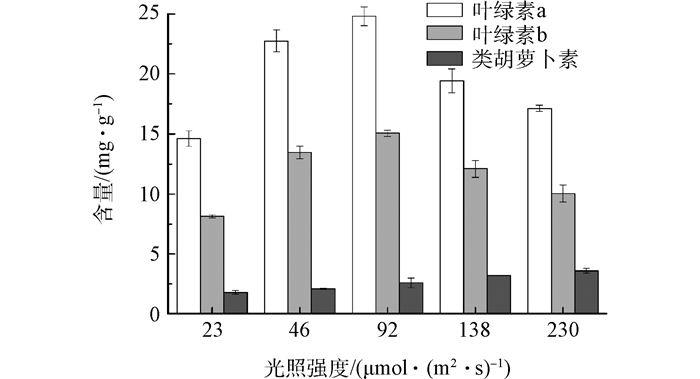
叶绿素是微藻进行光合作用的重要色素,其含量对光合速率的大小有直接影响。叶绿素作为光合色素中重要的色素,是反映微藻生理状况的一个重要指标。光照强度对小球藻细胞内叶绿素含量的影响如图 5所示。小球藻细胞内叶绿素a和叶绿素b的含量随着光照强度的增加呈先上升后下降的趋势,而类胡萝卜素的含量呈稳定上升趋势。在光照强度为92 μmol·(m2·s)-1时,叶绿素a和叶绿素b的含量达到最大,分别为24.81 mg·g-1和15.08 mg·g-1。当小球藻进行光合作用时,叶绿素a和b捕获光能并将能量传递至光反应中心,但激发态叶绿素产生的能量不能完全传递,类胡萝卜素作为光保护色素则吸收剩余的能量,避免产生激发态的活性氧伤害膜体,从而达到光保护作用[15]。因此,当光照强度为限制性条件时,随光照强度增加,细胞活性增强,叶绿素a和叶绿素b的合成量增加以捕获足够的光能供细胞生长代谢,类胡萝卜素含量相对较低。而当光照强度超过小球藻生长所需的光饱和点时,细胞活性下降,叶绿素a和叶绿素b合成量减少,类胡萝卜素的含量相应升高,以提高其保护机制,具体的作用机制尚需深入探索。
图5 光照强度对小球藻细胞内叶绿素含量的影响
Fig. 5 Effects of light intensity on chlorophyll content of microalgae
Fig. 5 Effects of light intensity on chlorophyll content of microalgae
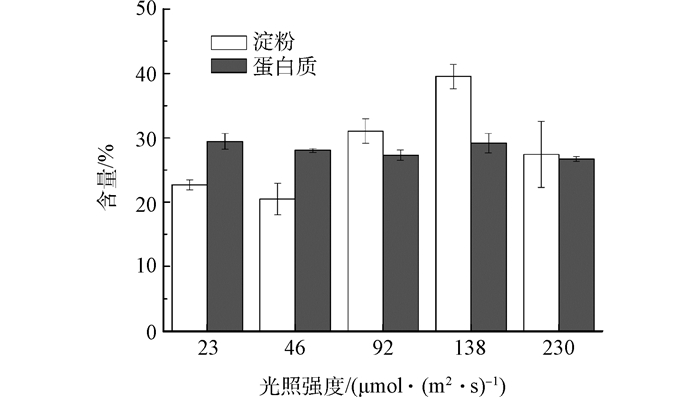
淀粉和脂肪酸是小球藻细胞内重要的能量物质和生理代谢物质。图 6为光照强度影响下藻生物膜内淀粉和蛋白质含量的变化趋势。在低光照强度(23 μmol·(m2·s)-1和46 μmol·(m2·s)-1)下,小球藻生物膜细胞内淀粉含量较低,占细胞干重的20.0%~22.0%。当光照强度为138 μmol·(m2·s)-1时,淀粉含量达到最大,占细胞干重的39.0%。随着光强进一步增强(230 μmol·(m2·s)-1),小球藻细胞内淀粉含量下降,但仍占细胞干重的27%。而光照强度对小球藻细胞内蛋白质含量影响较小,为26.7%~29.4%。小球藻通过光合作用将CO2固定并转化为胞内有机物,细胞中的有机碳首先用于小球藻的生长和代谢,其次合成碳水化合物和脂肪酸,碳水化合物中大部分为淀粉。研究发现,适当增加光照强度有利于小球藻细胞中淀粉合成[16],但是当光照强度过高时,更多的碳将用于脂肪酸的合成,从而导致淀粉含量的降低[17]。研究还发现,随光照强度的变化,微藻生物膜细胞内蛋白质含量变化较小。这主要是由于当培养基中氮源充足时,光能对蛋白质合成无直接影响,所以表现为微藻生物膜细胞内蛋白质含量受光照强度影响较小。
图6 光照强度对小球藻细胞内淀粉、蛋白质含量的影响
Fig. 6 Effects of light intensity on starch and protein content of microalgae
Fig. 6 Effects of light intensity on starch and protein content of microalgae
图6 光照强度对小球藻细胞内淀粉、蛋白质含量的影响
Fig. 6 Effects of light intensity on starch and protein content of microalgae
Fig. 6 Effects of light intensity on starch and protein content of microalgae
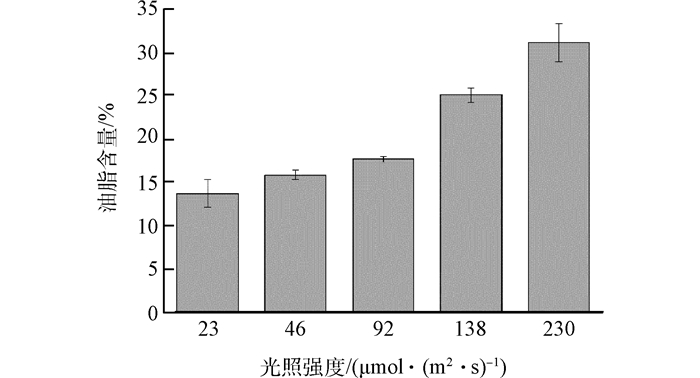
光照强度不仅影响藻细胞的色素含量、光合活性,而且还影响细胞内油脂含量[18]。图 7为小球藻生物膜细胞中油脂含量随着光照强度的变化趋势。由图 7可知,随光照强度增加,藻生物膜细胞内油脂含量呈递增趋势。对比图 2、图 6和图 7发现,在光强为230 μmol·(m2·s)-1时,获得最大的生物量产率(5.50 g·(m2·d)-1)和油脂产率(1.71 g·(m2·d)-1);但当光照强度为138 μmol·(m2·s)-1时,获得最大的淀粉含量,继续增大光照强度,淀粉含量反而下降。结果进一步表明,一定光照强度的胁迫有利于藻生物膜细胞内油脂积累的正调控,促进碳流向油脂的合成。
图7 光照强度对小球藻细胞油脂含量的影响
Fig. 7 Effect of light intensity on lipid content of Chlorella
Fig. 7 Effect of light intensity on lipid content of Chlorella
淀粉和脂肪酸的代谢过程都利用2个共同的碳前体:3-磷酸甘油酸和3-磷酸甘油醛[19]。适当地增加光照强度有利于小球藻细胞中淀粉的合成,但是当光照强度达到一定数值时,更利于脂肪酸的合成,导致淀粉含量的降低。YOO等[20]研究发现,Botryococcus braunii KMITL2的油脂含量随光照强度的增大而增加,并在光强为538 μmol·(m2·s)-1达到最大。因此,对于小球藻油脂含量而言,提高光照强度能促进小球藻油脂的积累。
小球藻细胞内脂肪酸含量以及组分含量可随环境因素的改变而改变[21]。不同光照强度下小球藻微油脂的组成如表 1所示。小球藻生物膜细胞内主要脂肪酸组成仍为C16~C18碳链脂肪酸。随着光照强度增加,C16~C18的含量呈先增加后下降趋势。微藻细胞内饱和脂肪酸比例维持相对稳定,不饱和脂肪酸比例略呈下降趋势,分别占总油脂含量的30.0%和60.0%左右。除表 1所列出的6种脂肪酸以外,其他脂肪酸含量随着光强增加,从2.6%增加至16.92%,但C16~C18碳链脂肪酸的含量仍占总油脂含量的80.0%以上,表明是制备生物柴油的较理想原料[22]。
表1 不同光照强度下小球藻微油脂的组成
Table 1 1 Fatty acid compositions of C. pyrenoidosa versus different light intensities
Table 1 1 Fatty acid compositions of C. pyrenoidosa versus different light intensities
表1 不同光照强度下小球藻微油脂的组成
Table 1 1 Fatty acid compositions of C. pyrenoidosa versus different light intensities
Table 1 1 Fatty acid compositions of C. pyrenoidosa versus different light intensities
| 光照强度/ (μmol·(m2·s)-1) |
FACHB-9中各脂肪酸含量/% | |||||||||
| 十六烷酸 (C16:0) |
十六碳一烯酸 (C16:1) |
十八烷酸 (C18:0) |
十八碳一烯酸 (C18:1) |
十八碳二烯酸 (C18:2) |
十八碳三烯酸 (C18:3) |
饱和脂肪酸 | 不饱和脂肪酸 | C16~C18 碳链脂肪酸 |
其他脂肪酸 | |
| 23 | 16.4 | 8.4 | 15.2 | 3 | 27.3 | 23.6 | 31.6 | 62.3 | 93.8 | 6.2 |
| 46 | 18.2 | 11.7 | 14.5 | 3.3 | 29.7 | 19.9 | 32.7 | 64.7 | 97.4 | 2.6 |
| 92 | 15 | 5.1 | 15.5 | 4.1 | 16.6 | 33.6 | 30.5 | 59.3 | 89.8 | 10.2 |
| 138 | 17.2 | 5.7 | 13.7 | 3 | 17.3 | 31.7 | 30.9 | 57.7 | 88.6 | 11.4 |
| 230 | 18.2 | 5.4 | 12.4 | 2.1 | 16.9 | 28.1 | 30.6 | 52.5 | 83.1 | 16.9 |
3 结论
1) 研究了在吸附生长模式下,光照强度对小球藻的生长与代谢的调控特性。小球藻在吸附生长模式下,较弱的光照强度有利于小球藻前期生长,但是随着小球藻生物膜厚度的增加,细胞之间相互遮蔽,较强的光照条件更利于小球藻后期生物膜生长。
2) 细胞内叶绿素a和b及淀粉含量随光照强度增加呈先增加后减少的趋势,但类胡萝卜素和蛋白质的含量变化较小。
3) 高光强胁迫有利于胞内油脂的积累,实验中当光照强度在230 μmol·(m2·s)-1时得到最大的总油脂含量(1.71 g·(m2·d)-1)。在小球藻的油脂组成中,在光照强度为46 μmol·(m2·s)-1时,得到最大的C16~C18碳链脂肪酸含量及不饱和脂肪酸含量,而除C16~C18外的其他脂肪酸组分则随光照强度的增加逐渐增加。
 下载:
下载:
