



-
农田中的氮磷难以被农作物完全吸收,并容易随农田排水进入自然环境之中,这会导致了区域水体的富营养化从而对生态系统产生严重威胁[1-2]。根据2015年中国农业部的相关数据,我国农业化肥的使用量高达6022万吨,但利用率仅为35.2%,这导致自然水体中的氮磷主要来自于农业中的化肥施用,其贡献率高达57.2 %和67.4 %[3]。与此同时,抗菌药物已经广泛应用于农业生产中,但它们在动物体内的降解效率较低从而大量残留于畜禽粪便中,而畜禽粪便的土地利用导致了耐药基因在自然环境中的富集与传播,其所引发的抗生素抗性问题已经成为了当今世界最大的公共健康问题之一[4-7]。农田土壤作为地球上微生物多样化程度最高的栖息地之一,是耐药基因的巨大储存库,其中部分耐药基因能够随着农田排水进入自然环境之中,而上述耐药基因能够以环境微生物为宿主从而进一步地传播与扩散[8-9]。已有研究表明农业灌溉系统中含有丰富的耐药基因,而农田排水向自然水体的释放可能会成为耐药基因在水体环境中积累与传播的主要途径之一[10]。综上所述,农田排水中的耐药基因能够以微生物为宿主在环境中富集与传播,而其中的氮磷则会导致自然水体富营养化,这对自然环境和公共健康产生极大的潜在威胁。
农田排水沟渠系统作为农田与自然水体的过渡带,已经成为了控制农田中氮磷与耐药基因向自然环境中传播与扩散的关键。传统的农田排水沟渠系统采用混凝土构筑,无法有效拦截农田排水中的氮磷,同时它们较差的水土保持能力容易引起水土流失[11-13]。生态沟渠是对传统农田排水沟渠系统的升级与改造,它通过土壤-微生物-植物形成生态链平衡系统,并利用其新陈代谢协同降解农田排水中的有机物、氮、磷等污染物,同时辅助脱氮除磷装置,拦截转化池,底泥污染捕获系统等功能性设备,进一步提高农田排水中污染物的拦截效果[14-15]。然而,生态沟渠对于农田排水中耐药基因的拦截效果尚未被研究,同时能够有效提高农田排水水质并降低其中氮磷浓度及耐药基因丰度的生态沟渠建设模式仍需探索。
开展生态沟渠系统建设是农业水环境治理的迫切需要,也是促进农业绿色发展、建设美丽田园的重要举措。因此,浙江省积极开展生态沟渠系统的建设工作,并在宁波市、湖州市、嘉兴市、绍兴市和台州市建设了20条高标准建设模式的生态沟渠。然而,生态沟渠的运行效果容易受多种因素影响,包括农田排水水力负荷、沟渠内部的水深、沟渠内部植物的种类以及水力停留时间等[1]。因此,本文全面评价20条高标准建设模式的生态沟渠对农田排水中氮磷的拦截效果,从而明确生态沟渠的建设模式对农田排水中氮磷拦截的影响。同时,生态沟渠系统中的水生植物和生态拦截功能设备能够有效控制来自于农田排水中的微生物,而耐药基因往往是被微生物所携带的[6-7, 16]。因此,生态沟渠系统具有降低农田排水中耐药基因的丰度,从而限制抗生素抗性的传播的应用潜质,但生态沟渠对农田排水中菌群和耐药基因的影响却鲜有研究。综上所述,本文评价浙江省生态沟渠对农田排水中氮磷的拦截效果,并进一步探究其对农田排水中耐药基因和细菌群落的影响,从而为生态沟渠的优化运行及推广建设、构建农业面源污染防控生态工程提供理论依据。
-
本研究供试生态沟渠为浙江省2018年在宁波市、湖州市、嘉兴市、绍兴市和台州市建设的20条高标准建设模式的生态沟渠,其中宁波市4条、湖州市5条、嘉兴市4条、绍兴市2条、台州市5条。上述生态沟渠汇水面积均在600亩以上,主要以水生植物修复为主体,其中主要包含挺水植物(美人蕉、再力花、野茭白等)、浮水植物(浮萍、荷花等)和沉水植物(金鱼藻、苦草、狐尾藻等),利用它们新陈代谢协同降解农田排水中的氮磷。与此同时,上述生态沟渠基本包含反硝化脱氮池、沉泥池、生态透水坝等标准化功能设施,进一步对农田排水中的氮磷进行拦截,从而尝试长期保持生态沟渠出水的良好水质。然而,宁波市生态沟渠由于受到台风影响,其中标准化功能设施的结构受损,其相关功能性也部分缺失。
-
浙江省5—6月为南方梅雨季节,生态沟渠中的流水主要以降雨产生的径流为主;6—10月为水稻生长季,而10月上中旬水稻收获,这个时间段农田无排灌水,因此生态沟渠中基本无水体样品。本研究于2019年9月10日至20日对浙江省20条高标准建设规模的生态沟渠进行水样采集,每条沟渠的采样点分别位于其上游阶段、中游阶段和下游阶段,每个采样点进行3次水样的采集,每次收集的样品为1000 mL,并在24 h内送回实验室,最后置于- 4 ℃冰箱中保存待用,测试数据均为3次水样的平均值。本研究针对采集到的湖州市生态沟渠-1的水体样品进行DNA提取,具体方法如下[6-7]。首先,利用0.22 µm 微孔滤膜对上述生态沟渠水样(100 mL)进行过滤,针对平行采集的水体样品进行3次重复过滤处理,从而将其中微生物细胞分别截留在滤膜上。然后,收集上述微孔滤膜,利用 PowerSoil®DNA 提取试剂盒(MOBIO, USA)提取其中DNA,然后将提取后的 DNA 平行样品充分混合并置于- 20 ℃的条件下保存待用,其浓度和纯度如表1所示。
-
水样过滤后,总氮浓度采用碱性过硫酸钾氧化-紫外分光光度法、总磷浓度采用钼酸铵分光光度法测定,氨氮测定参照《水与废水水质监测方法》第四版方法。
-
本研究将湖州市生态沟渠水体DNA样品送至上海美吉生物科技有限公司进行DNA宏基因组检测,具体方法如下。通过Covaris M220(基因公司,中国)将DNA片段化,并将其筛选打断为约400bp的片段。使用NEXTFLEX Rapid DNA-Seq(Bioo Scientific,美国)建库试剂盒构建PE文库。在桥式PCR扩增后使用 Illumina NovaSeq/Hiseq Xten(Illumina,美国)测序平台进行宏基因组测序。利用软件Fastp对原始数据进行质控,利用软件BWA将目标序列比对到宿主的DNA序列,并去除比对相似性高的污染序列。使用基于succinct de Bruijngraphs原理的拼接软件MEGAHIT对优化序列进行拼接组装。在拼接结果中筛选 ≥ 300 bp的contigs作为最终的组装结果。使用MetaGene对组装出的contig进行ORF预测,然后使用CD-HIT软件对所有样品预测出来的基因序列进行聚类,构建非冗余基因集。使用SOAPaligner软件,分别将每个样品的高质量序列与非冗余基因集进行比对(默认参数为:95% identity),统计基因在对应样品中的丰度信息。将非冗余基因集序列与Silva数据库(SSU132)进行比对(比对阈值为70 %),从而得到物种分类注释结果。利用TRIMMOMATIC、MOTHER、FLASH、QIIME等软件对高通量测序所得数据进行分析,从而评价样品中微生物多样性、物种组成和丰度、群落结构的差异。此外,使用 BLASTP(BLAST Version 2.2.28+,http://blast.ncbi.nlm.nih.gov/Blast.cgi)将非冗余基因集与CARD数据库进行比对,比对参数设置期望值 e-value 为 1e-5,获得基因对应的抗生素抗性功能注释信息,然后将样品中比对到的耐药基因序列数除以该样本中所有基因对应的序列数总和,并以PPM(Parts Per Million:每一百万条测序序列中,比对到某一基因序列的数目)的形式表示样品中不同耐药基因的丰度,具体公式如下。
式中,其中Ri代表耐药基因在某一样本中的丰度值,即该样本中比对到Genei上的序列数;
∑n1(Rj) 代表该样本中所有基因对应的序列数总和。 -
采用Microsoft Excel 2010对数据进行整理计算并用OriginPro 8.5进行相关图形的绘制,同时利用R语言软件(R 3.4.0)的pheatmap和VennDiagram程序包分别进行热图和Venn图的绘制。氮、磷的拦截率利用公式(R(%)=(Cin-Cout)/Cin×100%)计算,式中R为排水阶段氮、磷污染物拦截率(%);Cin为进水口氮、磷污染物质量浓度(mg·L−1);Cout为出水口氮、磷污染物质量浓度(mg·L−1)。
-
如图1所示,宁波市、湖州市、嘉兴市、绍兴市和台州市的生态沟渠对农田排水中的氮磷均有着明显的拦截效果。宁波市、湖州市、嘉兴市、绍兴市和台州市的生态沟渠对农田排水中的氮磷的平均拦截率在6.98%—48.7%,其中嘉兴市生态沟渠对农田排水中氨氮的平均拦截率高达48.1%,而绍兴市生态沟渠对农田排水中总氮的平均拦截率高达48.7%。然而,宁波市生态沟渠对农田排水中氮磷浓度的平均拦截率较低,基本维持在6.98%—15.1%的范围内,这可能是由于其生态沟渠标准化功能设施受台风影响部分缺失所导致。
图2全面比较了20条生态沟渠对于农田排水中氮磷的拦截效率,并通过以Bray-Curtis距离为基础的聚类分析将20条生态沟渠分为三个部分(Part1、Part2和Part3),从而突出能够同时降低农田排水中氮磷的生态沟渠。Part1部分的生态沟渠对农田排水中的氮和磷的拦截效率均相对较低。Part3部分的生态沟渠对农田排水中的总氮或总磷保持较好的拦截效率,其中湖州市生态沟渠-1对农田排水中总磷的拦截效率高达47.5%,但部分沟渠对农田排水中氨氮的拦截效率较低。Part2部分的生态沟渠对农田排水中的氨氮、总氮和总磷均有相对较好的拦截效率,其中嘉兴市生态沟渠-1对农田排水中氨氮的拦截效率高达51.3%,而绍兴市生态沟渠-1对农田排水中总氮的拦截效率高达49.2%。根据上述结果可知,嘉兴市、绍兴市、台州市和湖州市的大部分生态沟渠对农田排水中的氮磷均有着明显的拦截效果,其中40%的生态沟渠能够同时降低农田排水中氨氮、总磷和总氮的浓度。然而,宁波地区生态沟渠中的标准化设施由于受到台风损坏,导致其功能性缺失,从而影响了该地区生态沟渠的氮磷拦截效果,这也表明标准化功能设施在氮磷拦截中发挥的重要作用。
-
本研究全面评价生态沟渠对于农田排水的净化效果,分析了20条生态沟渠下游阶段出水中氨氮、总磷和总氮的浓度。如图3所示,浙江省内18条生态沟渠的下游阶段水体中氨氮浓度达到国家地表水环境质量标准Ⅰ类(氨氮≤ 0.15 mg·L−1),其他两条生态沟渠的下游阶段水体中氨氮浓度达到国家地表水环境质量标准Ⅱ类(氨氮≤ 0.50 mg·L−1)。同时,20条生态沟渠的下游阶段出水中总磷浓度在0.17—0.39 mg·L−1的范围内,均达到国家地表水环境质量标准Ⅱ类(总磷≤ 0.50 mg·L−1)。根据上述生态沟渠的下游阶段水体中总氮浓度的对比,本研究发现,其中2条生态沟渠达到国家地表水环境质量标准Ⅰ类(总氮≤ 0.15 mg·L−1),16条生态沟渠达到国家地表水环境质量标准Ⅱ类(总氮≤ 0.50 mg·L−1),2条生态沟渠达到国家地表水环境质量标准Ⅲ类(总氮≤ 1.0 mg·L−1)。综上可知,农田排水经过生态沟渠处理后,出水中氮磷(尤其是氨氮)含量很低,达到排放要求,对于环境没有明显负面影响。
开展农田氮磷生态拦截沟渠系统建设是农业水环境治理的迫切需要,也是促进农业绿色发展、建设美丽田园的重要举措。本研究选择宁波市、湖州市、绍兴市、舟山市和嘉兴市的20条标准建设规模的生态沟渠进行水体取样,研究发现生态沟渠均对农田排水中的氮和磷有着明显的拦截效果,而上述生态沟渠下游出水中氨氮、总氮和总磷的浓度很低,大部分都达到了国家地表水环境质量标准Ⅱ类,基本不会对环境所产生负面影响。生态沟渠合理地配置了沉水植物、浮水植物和挺水植物,主要包括再力花、美人蕉、茭白、浮萍、荷花、黄花水龙、灯心草和狐尾藻等,同时还建设了生态透水坝、沉泥池和反硝化池等标准化功能设施。生态沟渠内合理配置地水生植物能够直接吸收水体中的氮和磷[17],其发达的根系强泌氧作用提高微生物大量氧气来促进氮元素的转化,同时其茎秆降低了径流水流速,这导致颗粒态氮和磷地沉积和拦截[18-19]。陈英等[20]研究发现,生态沟渠中的两种水生植物组合(1:再力花、芦苇和黄花水龙;2:水芹、灯心草和菖蒲)对太湖流域农田排水中总氮和总磷的拦截率均不低于65%。田昌等[21]研究发现常德市津市市毛里湖生态沟渠对于小流域农田排水中氨氮、硝态氮、总氮和总磷的拦截率均超过50%,其中水生植物主要包括狐尾藻、茭白和芦苇等。与此同时,生态沟渠中的沉泥池和生态透水坝等标准化设施可以增加其中颗粒态氮和磷的沉积和吸附效果,而反硝化池的相对厌氧环境可以促进生态沟渠中氮磷的转化与降解。此外,宁波市地区生态沟渠由于受到台风的影响,其标准化功能设施的缺失影响了该地区生态沟渠的氮磷拦截效果,这也从侧面反映了标准化设施的功能性对生态沟渠的氮磷拦截起到了重要的效果。
-
湖州市生态沟渠-1对农田排水中总氮和总磷浓度的拦截率均接近50%,并且其下游阶段出水中氮磷的含量均低于大部分的生态沟渠,因此选择其进行生态沟渠对农田排水中耐药基因的影响研究。图4显示该生态沟渠上、中游阶段水体中耐药基因子类数量分别为257种和255种,其单独拥有的耐药基因子类数量分别为26种和9种。然而,生态沟渠下游阶段中耐药基因子类的数量为250种,其单独拥有的耐药基因种类为0种,结果表明生态沟渠下游阶段的耐药基因数量略低于其上游阶段和中游阶段,表明生态沟渠对农田排水中的耐药性基因起到了一定拦截效果。
图5表明生态沟渠上游段、中游段和下游段水体中耐药基因子类的种类及主要耐药基因的丰度,其中主要耐药基因为氟喹诺酮类、甲氧苄啶类和四环素类耐药基因。
氟喹诺酮类耐药基因在上述生态沟渠不同阶段水体中的丰度占比均最高,为41.3%—48.6%。氟喹诺酮类耐药基因在生态沟渠下游段水体相比在生态沟渠上游段水体,丰度降低了12.5%。具体而言,氟喹诺酮类耐药基因子类mfd、甲氧苄啶类耐药基因子类dfrE和四环素类耐药基因子类tetT在上述生态沟渠不同阶段水体中均有检测到且保持相对较高的丰度。因此,湖州-1生态沟渠上游段、中游段和下游段水体中主要的耐药基因子类为mfd、dfrE和tetT,它们的丰度之和占耐药基因总丰度的比例高达66.5%—69.3%。耐药基因mfd作为上述生态沟渠中丰度最高的耐药基因子类,在生态沟渠上游段水体中的丰度为133 ×10-6,而其在沟渠中游段和下游段水体的丰度分别降至129 ×10-6和117 ×10-6。与此同时,耐药基因dfrE和tetT在生态沟渠下游段水体中的丰度分别为27.8×10-6和24.4 ×10-6,相比于其在生态沟渠上游段水体中的丰度降低了41.4%和14.4%,结果表明生态沟渠能够降低农田排水中主要耐药基因的丰度。图6展示了湖州市生态沟渠-1不同阶段水体中微生物群落的组分特征。
在生态沟渠上游阶段,水体中主要包含Proteobacteria门和Verrucomicrobia门的微生物,它们的丰度分别占微生物总丰度的47.2%和16.3%。随着生态沟渠中水体从上游阶段流至中下游阶段,其中Proteobacteria门和Verrucomicrobia门的微生物分别降低15.9%—29.2 %和39.1%—67.4 %,而其中Actinobacteria门的微生物丰度增加了3.26—3.75倍。最终,生态沟渠下游水体中的微生物类别主要为Proteobacteria门和Actinobacteria门,它们的丰度分别占微生物总丰度的33.4%和21.9%。同时,生态沟渠上游阶段水体中微生物在种水平上的类别主要为Haloferula_sp._BvORR071、Vogesella_sp._EB、Hassallia_byssoidea和Hydrogenophaga_sp._Ro ot209,它们的丰度在中下游阶段明显降低。上述结果显示生态沟渠对农田排水的微生物群落结构具有显著的影响,而环境环境中耐药基因往往是被其中微生物所携带的,因此生态沟渠中微生物群落组分的改变可能与其中耐药基因丰度的变化之间存在着一定程度的联系,但具体的机制仍需进一步研究。
抗生素广泛用于农业和畜牧业,导致多种类且高丰度的耐药基因出现在水体环境。近年来,越来越多的研究发现农田灌溉系统中含有大量的耐药基因,因此农田灌溉系统被认为是环境中耐药基因富集与传播的媒介之一。为了明确耐药基因在人工农业灌溉系统中的出现及其分布,已有研究在内蒙古河套的农田灌区系统的农田排水渠中检测出多种类且高丰度的耐药基因,并发现五粮树海流域生态系统有效降低内农田灌区系统排水中耐药基因丰度,这可能是由于该生态系统内水生动植物的吸附和拦截作用所导致的[10]。本研究在湖州市生态沟渠-1上游阶段水体中发现了不同种类的耐药基因,主要包括mfd、dfrE和tetT,这是由于农田排水中的耐药基因转移至生态沟渠所致。然而,生态沟渠下游阶段水体中mfd、dfrE和tetT的丰度明显低于其上游阶段水体中对应耐药基因的丰度,这证明生态沟渠对农田排水中的耐药基因有着一定的拦截作用。此外,湖州市生态沟渠-1有效降低了农田排水中Proteobacteria门和Verrucomicrobia门的微生物丰度,并提高了其中Actinobacteria门的微生物丰度。生态沟渠中的水生植物和功能设施能够有效的拦截农田排水中的部分颗粒物及其携带的微生物群落,而耐药基因往往是由微生物所携带的,这可能有利于生态沟渠降低农田排水中耐药基因的丰度[6-7]。与此同时,氮磷等营养元素能够促进携带耐药基因的微生物的繁殖,而生态沟渠可以通过有效拦截农田排水中的氮磷来降低耐药微生物在自然环境中的富集以及耐药基因在自然环境中的传播。因此,生态沟渠不仅对农田排水中的氮与磷有着较好的拦截作用,还降低了农田排水中主要耐药基因的丰度,并改变了其中微生物群落的组分,这有利于限制耐药基因在自然环境中的富集与传播。
-
浙江省内嘉兴市、绍兴市、台州市和湖州市建设的高标准生态沟渠对农田排水中的氮磷均有着明显的拦截效果,其中40%的生态沟渠能够同时明显降低农田排水中氨氮、总磷和总氮的浓度。嘉兴市生态沟渠对农田排水中氨氮浓度的平均拦截率高达48.1%,而绍兴市生态沟渠对总氮浓度的平均降低率高达48.7%。同时,湖州市生态沟渠-1不仅有效拦截了农田排水中的氮和磷,还明显降低了农田排水中主要耐药基因的丰度(mfd、dfrE和tetT),并改变了其中微生物群落的组分(Proteobacteria门、Verrucomicrobia门和Actinobacteria门微生物)。综上可知,浙江省生态沟渠能够有效拦截农田排水中的氮磷和耐药基因,其高标准的建设模式值得推广,但生态沟渠限制耐药基因传播与扩散的机制仍需进一步研究。
生态沟渠对农田排水中菌群组分、耐药基因谱及氮磷浓度的影响
Effects of the ecological ditches on bacterial community composition, antibiotic resistance gene profiles, nitrogen and phosphorus concentrations in farmland drainage
-
摘要: 农田排水中的耐药基因能够以微生物为宿主在环境中富集与传播,而其中的氮磷则会导致自然水体富营养化。生态沟渠作为农田排水的末端处理系统,是控制农田中氮磷向自然环境中转移的关键,但其对农田排水中菌群和耐药基因的影响却鲜有研究。本研究对浙江省内20条生态沟渠的氮磷拦截效果进行了评价,然后利用宏基因组测序技术对其中效果较好的生态沟渠进行了菌群组分和耐药基因谱的分析。研究结果表明,浙江省内生态沟渠有效的拦截了农田排水中的氮磷,其中湖州市生态沟渠-1对农田排水中总氮和总磷拦截率均接近50%,相较于其他生态沟渠具有更好的氮磷拦截效果。同时,湖州市生态沟渠-1有效降低了农田排水中耐药基因的种类,并将主要耐药基因子类(mfd、dfrE和tetT)的丰度降低了12.0%—41.4%。此外,湖州市生态沟渠-1有效降低了农田排水中Proteobacteria门和Verrucomicrobia门的微生物丰度,并提高了其中Actinobacteria门的微生物丰度。综上可知,浙江省生态沟渠不仅能够拦截农田排水中的氮磷和耐药基因还能够影响其中细菌群落组分,但相关机制仍需进一步研究。Abstract:
The antibiotic resistance genes (ARGs) in farmland drainage promote the increase and spread of antibiotic resistance among environmental microbe and the anthropogenic nutrient (nitrogen and phosphorus) accelerates the eutrophication of lakes and streams. The ecological ditches, as the end treatment system of farmland drainage, are vital to mitigate the transfer of nitrogen and phosphorus from farmland drainage into natural environment, but few studies focused on the influences of the ecological ditches on ARGs and bacterial community in farmland drainage. This study comprehensively evaluated the effects of 20 ecological ditches in Zhejiang province on nutrients removal, then further applied the technology of metagenomic sequencing to analyze ARG profiles and bacterial community composition in the ecological ditch with effective nutrients removal. The related results indicated that the ecological ditches are effective in nutrients removal from farmland drainage, and the average interception efficiency of total nitrogen and total phosphorus were both closed to 50% in the Huzhou ecological ditch-1, which is more efficient comparing with that of other ecological ditches. Meantime, the Huzhou ecological ditch-1 effectively reduced the diversity of ARG subtypes in farmland drainage, and reduced the abundances of main ARGs (mfd、dfrE and tetT)by 12.0%—41.4%. In addition, the abundance of bacteria belonging to the phylum of Proteobacteri and Verrucomicrobia has been reduced, while the abundance of bacteria belonging to the phylum of Actinobacteria has been increased after the treatment of Huzhou ecological ditch-1. In summary, the ecological ditches not only contribute to the removal of nitrogen, phosphorus and ARGs in farmland drainage but also influencing its bacterial community, but the related mechanism needs to be further investigated. -
Key words:
- Antibiotic resistance genes /
- Ecological ditch /
- Farmland drainage /
- Bacterial community
-
农田中的氮磷难以被农作物完全吸收,并容易随农田排水进入自然环境之中,这会导致了区域水体的富营养化从而对生态系统产生严重威胁[1-2]。根据2015年中国农业部的相关数据,我国农业化肥的使用量高达6022万吨,但利用率仅为35.2%,这导致自然水体中的氮磷主要来自于农业中的化肥施用,其贡献率高达57.2 %和67.4 %[3]。与此同时,抗菌药物已经广泛应用于农业生产中,但它们在动物体内的降解效率较低从而大量残留于畜禽粪便中,而畜禽粪便的土地利用导致了耐药基因在自然环境中的富集与传播,其所引发的抗生素抗性问题已经成为了当今世界最大的公共健康问题之一[4-7]。农田土壤作为地球上微生物多样化程度最高的栖息地之一,是耐药基因的巨大储存库,其中部分耐药基因能够随着农田排水进入自然环境之中,而上述耐药基因能够以环境微生物为宿主从而进一步地传播与扩散[8-9]。已有研究表明农业灌溉系统中含有丰富的耐药基因,而农田排水向自然水体的释放可能会成为耐药基因在水体环境中积累与传播的主要途径之一[10]。综上所述,农田排水中的耐药基因能够以微生物为宿主在环境中富集与传播,而其中的氮磷则会导致自然水体富营养化,这对自然环境和公共健康产生极大的潜在威胁。
农田排水沟渠系统作为农田与自然水体的过渡带,已经成为了控制农田中氮磷与耐药基因向自然环境中传播与扩散的关键。传统的农田排水沟渠系统采用混凝土构筑,无法有效拦截农田排水中的氮磷,同时它们较差的水土保持能力容易引起水土流失[11-13]。生态沟渠是对传统农田排水沟渠系统的升级与改造,它通过土壤-微生物-植物形成生态链平衡系统,并利用其新陈代谢协同降解农田排水中的有机物、氮、磷等污染物,同时辅助脱氮除磷装置,拦截转化池,底泥污染捕获系统等功能性设备,进一步提高农田排水中污染物的拦截效果[14-15]。然而,生态沟渠对于农田排水中耐药基因的拦截效果尚未被研究,同时能够有效提高农田排水水质并降低其中氮磷浓度及耐药基因丰度的生态沟渠建设模式仍需探索。
开展生态沟渠系统建设是农业水环境治理的迫切需要,也是促进农业绿色发展、建设美丽田园的重要举措。因此,浙江省积极开展生态沟渠系统的建设工作,并在宁波市、湖州市、嘉兴市、绍兴市和台州市建设了20条高标准建设模式的生态沟渠。然而,生态沟渠的运行效果容易受多种因素影响,包括农田排水水力负荷、沟渠内部的水深、沟渠内部植物的种类以及水力停留时间等[1]。因此,本文全面评价20条高标准建设模式的生态沟渠对农田排水中氮磷的拦截效果,从而明确生态沟渠的建设模式对农田排水中氮磷拦截的影响。同时,生态沟渠系统中的水生植物和生态拦截功能设备能够有效控制来自于农田排水中的微生物,而耐药基因往往是被微生物所携带的[6-7, 16]。因此,生态沟渠系统具有降低农田排水中耐药基因的丰度,从而限制抗生素抗性的传播的应用潜质,但生态沟渠对农田排水中菌群和耐药基因的影响却鲜有研究。综上所述,本文评价浙江省生态沟渠对农田排水中氮磷的拦截效果,并进一步探究其对农田排水中耐药基因和细菌群落的影响,从而为生态沟渠的优化运行及推广建设、构建农业面源污染防控生态工程提供理论依据。
1. 材料与方法(Materials and methods)
1.1 生态沟渠建设概况
本研究供试生态沟渠为浙江省2018年在宁波市、湖州市、嘉兴市、绍兴市和台州市建设的20条高标准建设模式的生态沟渠,其中宁波市4条、湖州市5条、嘉兴市4条、绍兴市2条、台州市5条。上述生态沟渠汇水面积均在600亩以上,主要以水生植物修复为主体,其中主要包含挺水植物(美人蕉、再力花、野茭白等)、浮水植物(浮萍、荷花等)和沉水植物(金鱼藻、苦草、狐尾藻等),利用它们新陈代谢协同降解农田排水中的氮磷。与此同时,上述生态沟渠基本包含反硝化脱氮池、沉泥池、生态透水坝等标准化功能设施,进一步对农田排水中的氮磷进行拦截,从而尝试长期保持生态沟渠出水的良好水质。然而,宁波市生态沟渠由于受到台风影响,其中标准化功能设施的结构受损,其相关功能性也部分缺失。
1.2 生态沟渠水体样品采集及DNA提取
浙江省5—6月为南方梅雨季节,生态沟渠中的流水主要以降雨产生的径流为主;6—10月为水稻生长季,而10月上中旬水稻收获,这个时间段农田无排灌水,因此生态沟渠中基本无水体样品。本研究于2019年9月10日至20日对浙江省20条高标准建设规模的生态沟渠进行水样采集,每条沟渠的采样点分别位于其上游阶段、中游阶段和下游阶段,每个采样点进行3次水样的采集,每次收集的样品为1000 mL,并在24 h内送回实验室,最后置于- 4 ℃冰箱中保存待用,测试数据均为3次水样的平均值。本研究针对采集到的湖州市生态沟渠-1的水体样品进行DNA提取,具体方法如下[6-7]。首先,利用0.22 µm 微孔滤膜对上述生态沟渠水样(100 mL)进行过滤,针对平行采集的水体样品进行3次重复过滤处理,从而将其中微生物细胞分别截留在滤膜上。然后,收集上述微孔滤膜,利用 PowerSoil®DNA 提取试剂盒(MOBIO, USA)提取其中DNA,然后将提取后的 DNA 平行样品充分混合并置于- 20 ℃的条件下保存待用,其浓度和纯度如表1所示。
表 1 湖州市生态沟渠-1不同阶段水体样品DNA性质Table 1. The properties of DNA extracted from different water samples in the Huzhou ecological ditch-1水体样品 Water Sample DNA浓度/(μg·L−1) DNA Concentration DNA纯度 DNA Purity(OD260/280) DNA总量 Total DNA 生态沟渠上游阶段 31.0 1.81 总量满足宏基因组一次标准建库要求 生态沟渠中游阶段 6.10 2.31 总量满足宏基因组一次标准建库要求 生态沟渠下游阶段 9.70 2.01 总量满足宏基因组一次标准建库要求 | Show Table DownLoad:
CSV
DownLoad:
CSV
1.3 生态沟渠水体样品测定指标及分析方法
水样过滤后,总氮浓度采用碱性过硫酸钾氧化-紫外分光光度法、总磷浓度采用钼酸铵分光光度法测定,氨氮测定参照《水与废水水质监测方法》第四版方法。
1.4 宏基因组检测方法
本研究将湖州市生态沟渠水体DNA样品送至上海美吉生物科技有限公司进行DNA宏基因组检测,具体方法如下。通过Covaris M220(基因公司,中国)将DNA片段化,并将其筛选打断为约400bp的片段。使用NEXTFLEX Rapid DNA-Seq(Bioo Scientific,美国)建库试剂盒构建PE文库。在桥式PCR扩增后使用 Illumina NovaSeq/Hiseq Xten(Illumina,美国)测序平台进行宏基因组测序。利用软件Fastp对原始数据进行质控,利用软件BWA将目标序列比对到宿主的DNA序列,并去除比对相似性高的污染序列。使用基于succinct de Bruijngraphs原理的拼接软件MEGAHIT对优化序列进行拼接组装。在拼接结果中筛选 ≥ 300 bp的contigs作为最终的组装结果。使用MetaGene对组装出的contig进行ORF预测,然后使用CD-HIT软件对所有样品预测出来的基因序列进行聚类,构建非冗余基因集。使用SOAPaligner软件,分别将每个样品的高质量序列与非冗余基因集进行比对(默认参数为:95% identity),统计基因在对应样品中的丰度信息。将非冗余基因集序列与Silva数据库(SSU132)进行比对(比对阈值为70 %),从而得到物种分类注释结果。利用TRIMMOMATIC、MOTHER、FLASH、QIIME等软件对高通量测序所得数据进行分析,从而评价样品中微生物多样性、物种组成和丰度、群落结构的差异。此外,使用 BLASTP(BLAST Version 2.2.28+,http://blast.ncbi.nlm.nih.gov/Blast.cgi)将非冗余基因集与CARD数据库进行比对,比对参数设置期望值 e-value 为 1e-5,获得基因对应的抗生素抗性功能注释信息,然后将样品中比对到的耐药基因序列数除以该样本中所有基因对应的序列数总和,并以PPM(Parts Per Million:每一百万条测序序列中,比对到某一基因序列的数目)的形式表示样品中不同耐药基因的丰度,具体公式如下。
PPMi=Ri×106∑n1(Rj) 式中,其中Ri代表耐药基因在某一样本中的丰度值,即该样本中比对到Genei上的序列数;
∑n1(Rj) 1.5 数据处理与分析
采用Microsoft Excel 2010对数据进行整理计算并用OriginPro 8.5进行相关图形的绘制,同时利用R语言软件(R 3.4.0)的pheatmap和VennDiagram程序包分别进行热图和Venn图的绘制。氮、磷的拦截率利用公式(R(%)=(Cin-Cout)/Cin×100%)计算,式中R为排水阶段氮、磷污染物拦截率(%);Cin为进水口氮、磷污染物质量浓度(mg·L−1);Cout为出水口氮、磷污染物质量浓度(mg·L−1)。
2. 结果与讨论(Results and discussion)
2.1 生态沟渠对农田排水中氮磷的拦截效率
如图1所示,宁波市、湖州市、嘉兴市、绍兴市和台州市的生态沟渠对农田排水中的氮磷均有着明显的拦截效果。宁波市、湖州市、嘉兴市、绍兴市和台州市的生态沟渠对农田排水中的氮磷的平均拦截率在6.98%—48.7%,其中嘉兴市生态沟渠对农田排水中氨氮的平均拦截率高达48.1%,而绍兴市生态沟渠对农田排水中总氮的平均拦截率高达48.7%。然而,宁波市生态沟渠对农田排水中氮磷浓度的平均拦截率较低,基本维持在6.98%—15.1%的范围内,这可能是由于其生态沟渠标准化功能设施受台风影响部分缺失所导致。
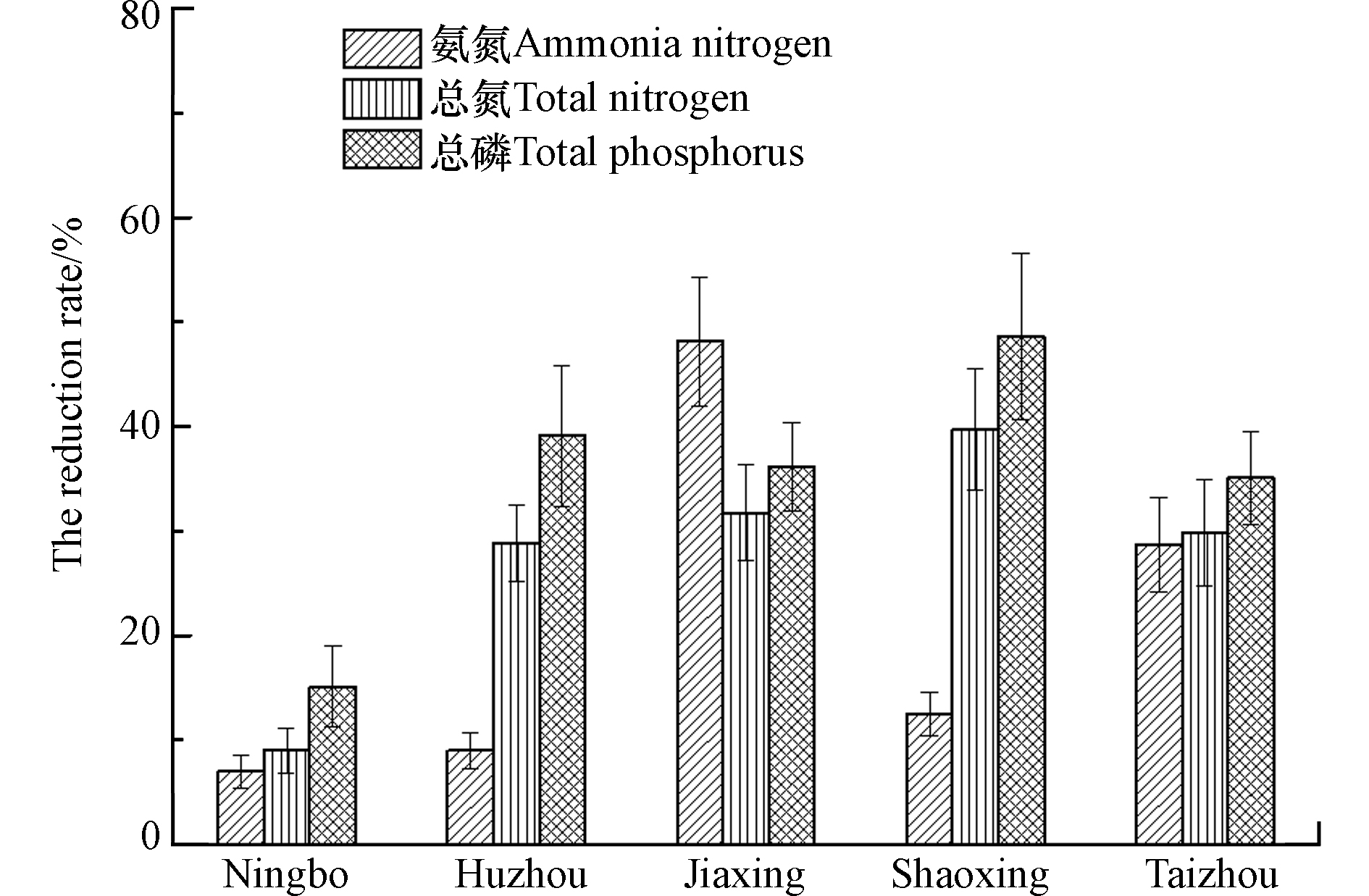 图 1 浙江省不同地市生态沟渠对农田排水中氮磷的拦截效果Figure 1. The interception effects of ecological ditches from cities in Zhejiang province on nitrogen and phosphorus in farmland drainage
图 1 浙江省不同地市生态沟渠对农田排水中氮磷的拦截效果Figure 1. The interception effects of ecological ditches from cities in Zhejiang province on nitrogen and phosphorus in farmland drainage图2全面比较了20条生态沟渠对于农田排水中氮磷的拦截效率,并通过以Bray-Curtis距离为基础的聚类分析将20条生态沟渠分为三个部分(Part1、Part2和Part3),从而突出能够同时降低农田排水中氮磷的生态沟渠。Part1部分的生态沟渠对农田排水中的氮和磷的拦截效率均相对较低。Part3部分的生态沟渠对农田排水中的总氮或总磷保持较好的拦截效率,其中湖州市生态沟渠-1对农田排水中总磷的拦截效率高达47.5%,但部分沟渠对农田排水中氨氮的拦截效率较低。Part2部分的生态沟渠对农田排水中的氨氮、总氮和总磷均有相对较好的拦截效率,其中嘉兴市生态沟渠-1对农田排水中氨氮的拦截效率高达51.3%,而绍兴市生态沟渠-1对农田排水中总氮的拦截效率高达49.2%。根据上述结果可知,嘉兴市、绍兴市、台州市和湖州市的大部分生态沟渠对农田排水中的氮磷均有着明显的拦截效果,其中40%的生态沟渠能够同时降低农田排水中氨氮、总磷和总氮的浓度。然而,宁波地区生态沟渠中的标准化设施由于受到台风损坏,导致其功能性缺失,从而影响了该地区生态沟渠的氮磷拦截效果,这也表明标准化功能设施在氮磷拦截中发挥的重要作用。
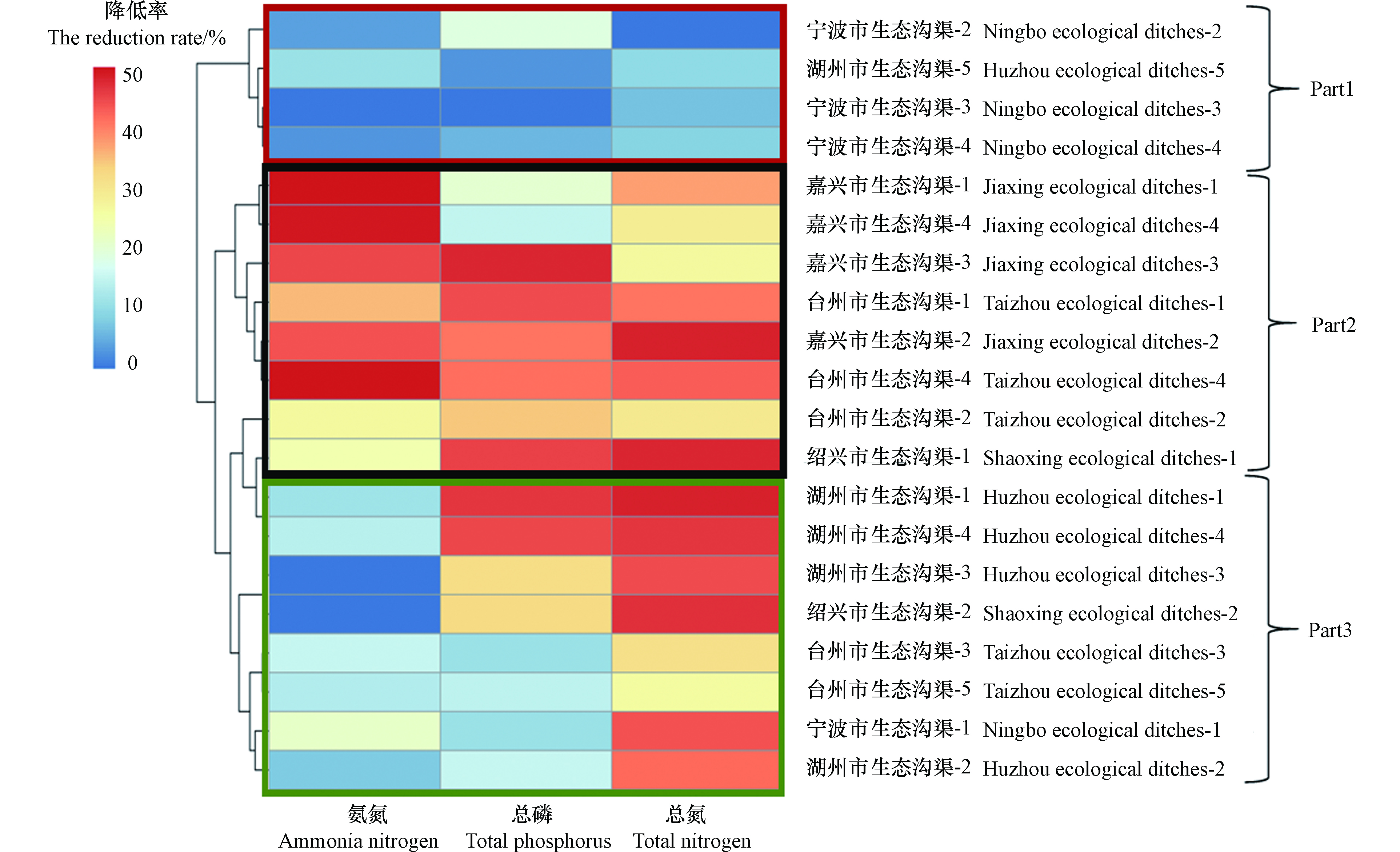 图 2 生态沟渠对农田排水中氮磷削减的热图Figure 2. Heatmap of the nitrogen and phosphorus elimination in farmland drainage by ecological ditches
图 2 生态沟渠对农田排水中氮磷削减的热图Figure 2. Heatmap of the nitrogen and phosphorus elimination in farmland drainage by ecological ditches2.2 生态沟渠处理后的农田排水中氮磷浓度
本研究全面评价生态沟渠对于农田排水的净化效果,分析了20条生态沟渠下游阶段出水中氨氮、总磷和总氮的浓度。如图3所示,浙江省内18条生态沟渠的下游阶段水体中氨氮浓度达到国家地表水环境质量标准Ⅰ类(氨氮≤ 0.15 mg·L−1),其他两条生态沟渠的下游阶段水体中氨氮浓度达到国家地表水环境质量标准Ⅱ类(氨氮≤ 0.50 mg·L−1)。同时,20条生态沟渠的下游阶段出水中总磷浓度在0.17—0.39 mg·L−1的范围内,均达到国家地表水环境质量标准Ⅱ类(总磷≤ 0.50 mg·L−1)。根据上述生态沟渠的下游阶段水体中总氮浓度的对比,本研究发现,其中2条生态沟渠达到国家地表水环境质量标准Ⅰ类(总氮≤ 0.15 mg·L−1),16条生态沟渠达到国家地表水环境质量标准Ⅱ类(总氮≤ 0.50 mg·L−1),2条生态沟渠达到国家地表水环境质量标准Ⅲ类(总氮≤ 1.0 mg·L−1)。综上可知,农田排水经过生态沟渠处理后,出水中氮磷(尤其是氨氮)含量很低,达到排放要求,对于环境没有明显负面影响。
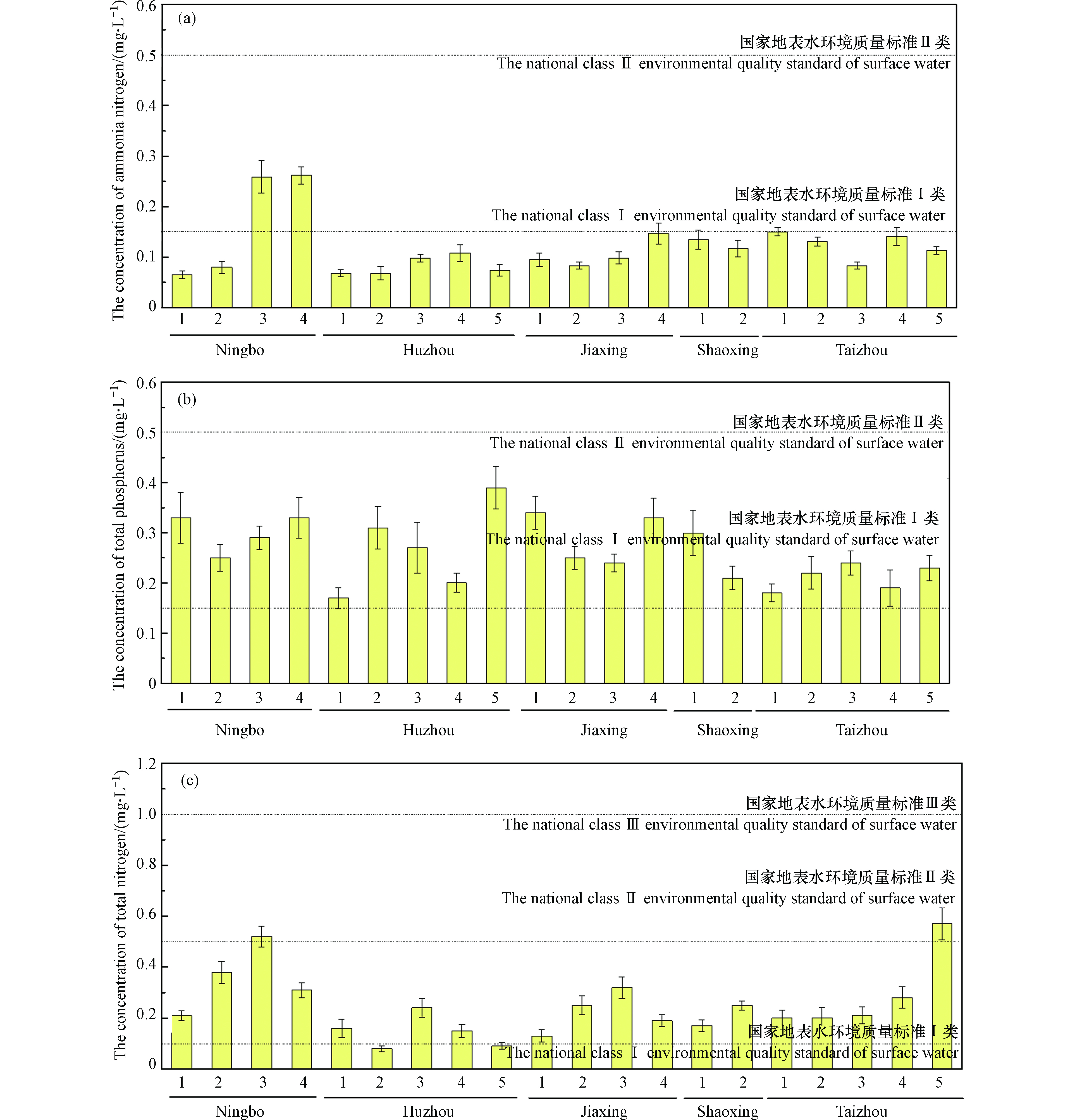 图 3 生态沟渠下游阶段出水总氮、氨氮及总磷的浓度Figure 3. The concentrations of total nitrogen, ammonia nitrogen and total phosphorus in downstream effluents of ecological ditches
图 3 生态沟渠下游阶段出水总氮、氨氮及总磷的浓度Figure 3. The concentrations of total nitrogen, ammonia nitrogen and total phosphorus in downstream effluents of ecological ditches开展农田氮磷生态拦截沟渠系统建设是农业水环境治理的迫切需要,也是促进农业绿色发展、建设美丽田园的重要举措。本研究选择宁波市、湖州市、绍兴市、舟山市和嘉兴市的20条标准建设规模的生态沟渠进行水体取样,研究发现生态沟渠均对农田排水中的氮和磷有着明显的拦截效果,而上述生态沟渠下游出水中氨氮、总氮和总磷的浓度很低,大部分都达到了国家地表水环境质量标准Ⅱ类,基本不会对环境所产生负面影响。生态沟渠合理地配置了沉水植物、浮水植物和挺水植物,主要包括再力花、美人蕉、茭白、浮萍、荷花、黄花水龙、灯心草和狐尾藻等,同时还建设了生态透水坝、沉泥池和反硝化池等标准化功能设施。生态沟渠内合理配置地水生植物能够直接吸收水体中的氮和磷[17],其发达的根系强泌氧作用提高微生物大量氧气来促进氮元素的转化,同时其茎秆降低了径流水流速,这导致颗粒态氮和磷地沉积和拦截[18-19]。陈英等[20]研究发现,生态沟渠中的两种水生植物组合(1:再力花、芦苇和黄花水龙;2:水芹、灯心草和菖蒲)对太湖流域农田排水中总氮和总磷的拦截率均不低于65%。田昌等[21]研究发现常德市津市市毛里湖生态沟渠对于小流域农田排水中氨氮、硝态氮、总氮和总磷的拦截率均超过50%,其中水生植物主要包括狐尾藻、茭白和芦苇等。与此同时,生态沟渠中的沉泥池和生态透水坝等标准化设施可以增加其中颗粒态氮和磷的沉积和吸附效果,而反硝化池的相对厌氧环境可以促进生态沟渠中氮磷的转化与降解。此外,宁波市地区生态沟渠由于受到台风的影响,其标准化功能设施的缺失影响了该地区生态沟渠的氮磷拦截效果,这也从侧面反映了标准化设施的功能性对生态沟渠的氮磷拦截起到了重要的效果。
2.3 生态沟渠对农田排水中耐药基因及微生物群落的影响
湖州市生态沟渠-1对农田排水中总氮和总磷浓度的拦截率均接近50%,并且其下游阶段出水中氮磷的含量均低于大部分的生态沟渠,因此选择其进行生态沟渠对农田排水中耐药基因的影响研究。图4显示该生态沟渠上、中游阶段水体中耐药基因子类数量分别为257种和255种,其单独拥有的耐药基因子类数量分别为26种和9种。然而,生态沟渠下游阶段中耐药基因子类的数量为250种,其单独拥有的耐药基因种类为0种,结果表明生态沟渠下游阶段的耐药基因数量略低于其上游阶段和中游阶段,表明生态沟渠对农田排水中的耐药性基因起到了一定拦截效果。
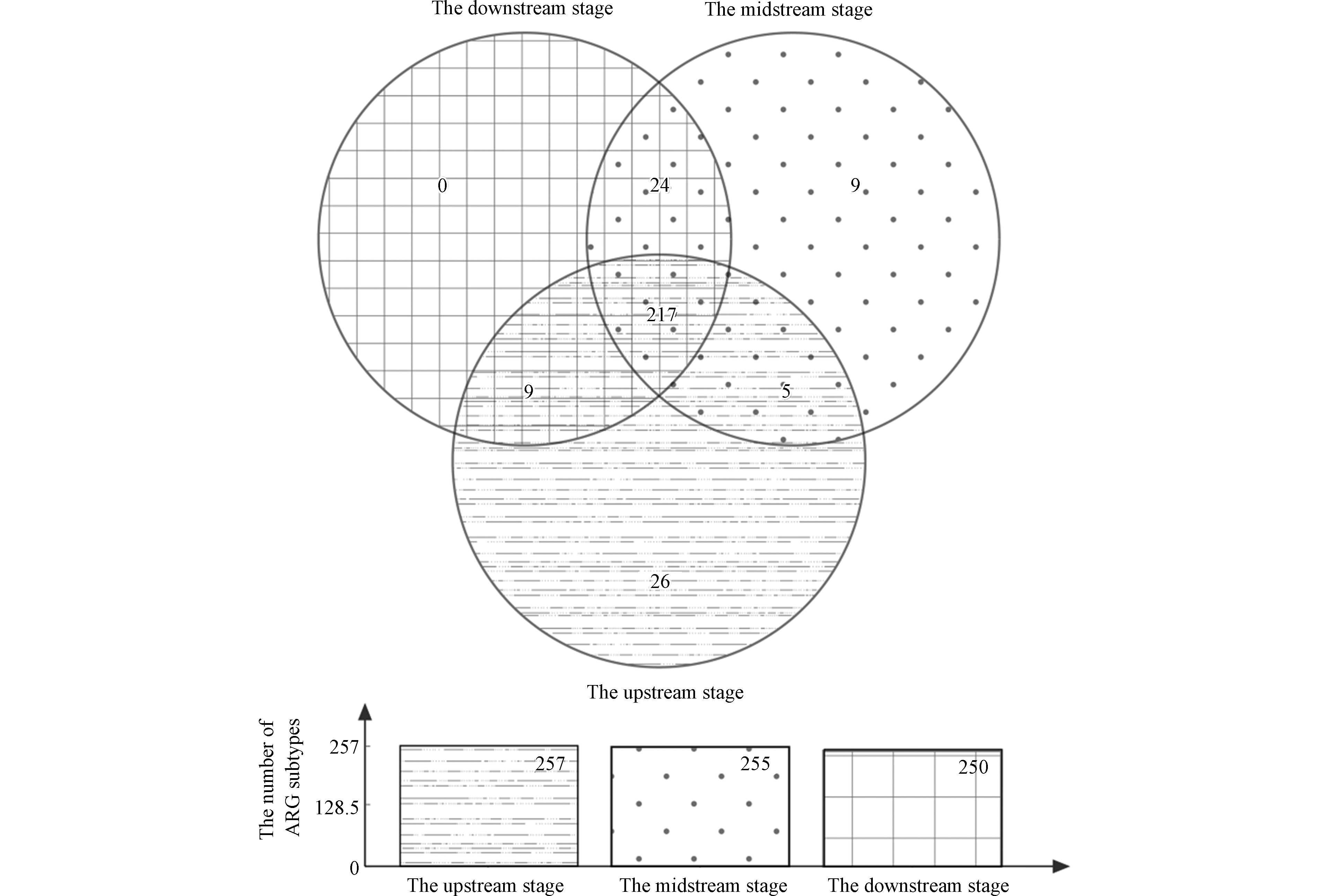 图 4 湖州市生态沟渠-1不同阶段水体中耐药基因子类数量的Venn图Figure 4. The venn diagram of ARG subtype numbers in water bodies from different stages of the Huzhou ecological ditch-1
图 4 湖州市生态沟渠-1不同阶段水体中耐药基因子类数量的Venn图Figure 4. The venn diagram of ARG subtype numbers in water bodies from different stages of the Huzhou ecological ditch-1图5表明生态沟渠上游段、中游段和下游段水体中耐药基因子类的种类及主要耐药基因的丰度,其中主要耐药基因为氟喹诺酮类、甲氧苄啶类和四环素类耐药基因。
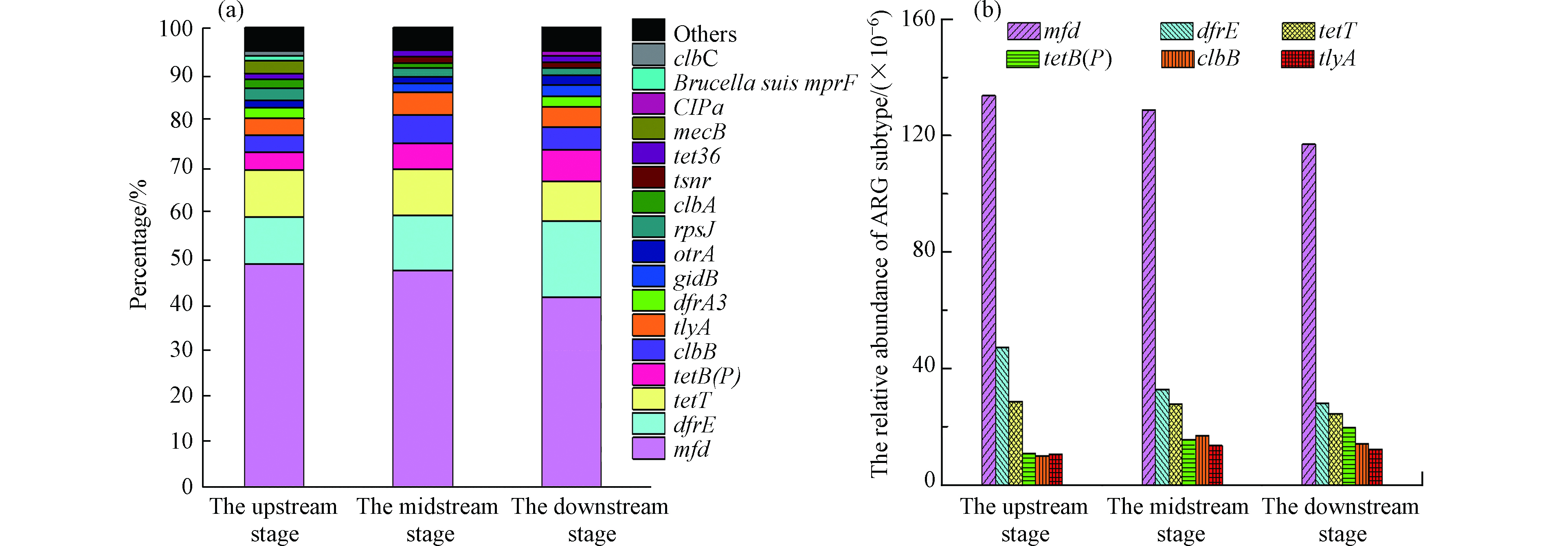 图 5 湖州市生态沟渠-1不同阶段水体中耐药基因种类(a)及主要耐药基因丰度(b)Figure 5. The diversity (a) and abundance (b) of ARG subtypes in water bodies from different stages of the Huzhou ecological ditch-1
图 5 湖州市生态沟渠-1不同阶段水体中耐药基因种类(a)及主要耐药基因丰度(b)Figure 5. The diversity (a) and abundance (b) of ARG subtypes in water bodies from different stages of the Huzhou ecological ditch-1氟喹诺酮类耐药基因在上述生态沟渠不同阶段水体中的丰度占比均最高,为41.3%—48.6%。氟喹诺酮类耐药基因在生态沟渠下游段水体相比在生态沟渠上游段水体,丰度降低了12.5%。具体而言,氟喹诺酮类耐药基因子类mfd、甲氧苄啶类耐药基因子类dfrE和四环素类耐药基因子类tetT在上述生态沟渠不同阶段水体中均有检测到且保持相对较高的丰度。因此,湖州-1生态沟渠上游段、中游段和下游段水体中主要的耐药基因子类为mfd、dfrE和tetT,它们的丰度之和占耐药基因总丰度的比例高达66.5%—69.3%。耐药基因mfd作为上述生态沟渠中丰度最高的耐药基因子类,在生态沟渠上游段水体中的丰度为133 ×10-6,而其在沟渠中游段和下游段水体的丰度分别降至129 ×10-6和117 ×10-6。与此同时,耐药基因dfrE和tetT在生态沟渠下游段水体中的丰度分别为27.8×10-6和24.4 ×10-6,相比于其在生态沟渠上游段水体中的丰度降低了41.4%和14.4%,结果表明生态沟渠能够降低农田排水中主要耐药基因的丰度。图6展示了湖州市生态沟渠-1不同阶段水体中微生物群落的组分特征。
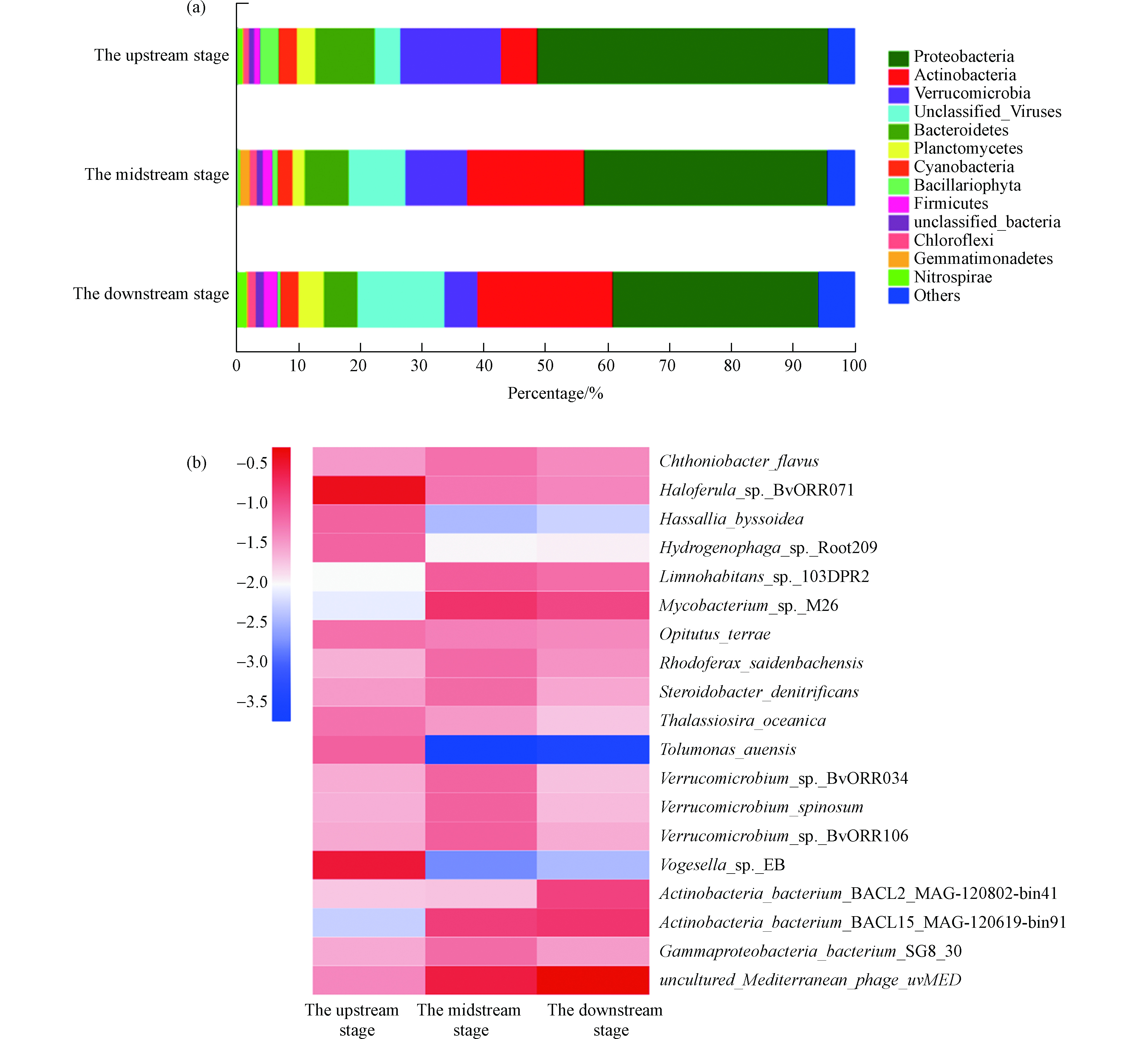 图 6 湖州市生态沟渠-1不同阶段水体中微生物群落组分特征Figure 6. The characteristic of microbial community compositions in water bodies from different stages of the Huzhou ecological ditch-1
图 6 湖州市生态沟渠-1不同阶段水体中微生物群落组分特征Figure 6. The characteristic of microbial community compositions in water bodies from different stages of the Huzhou ecological ditch-1在生态沟渠上游阶段,水体中主要包含Proteobacteria门和Verrucomicrobia门的微生物,它们的丰度分别占微生物总丰度的47.2%和16.3%。随着生态沟渠中水体从上游阶段流至中下游阶段,其中Proteobacteria门和Verrucomicrobia门的微生物分别降低15.9%—29.2 %和39.1%—67.4 %,而其中Actinobacteria门的微生物丰度增加了3.26—3.75倍。最终,生态沟渠下游水体中的微生物类别主要为Proteobacteria门和Actinobacteria门,它们的丰度分别占微生物总丰度的33.4%和21.9%。同时,生态沟渠上游阶段水体中微生物在种水平上的类别主要为Haloferula_sp._BvORR071、Vogesella_sp._EB、Hassallia_byssoidea和Hydrogenophaga_sp._Ro ot209,它们的丰度在中下游阶段明显降低。上述结果显示生态沟渠对农田排水的微生物群落结构具有显著的影响,而环境环境中耐药基因往往是被其中微生物所携带的,因此生态沟渠中微生物群落组分的改变可能与其中耐药基因丰度的变化之间存在着一定程度的联系,但具体的机制仍需进一步研究。
抗生素广泛用于农业和畜牧业,导致多种类且高丰度的耐药基因出现在水体环境。近年来,越来越多的研究发现农田灌溉系统中含有大量的耐药基因,因此农田灌溉系统被认为是环境中耐药基因富集与传播的媒介之一。为了明确耐药基因在人工农业灌溉系统中的出现及其分布,已有研究在内蒙古河套的农田灌区系统的农田排水渠中检测出多种类且高丰度的耐药基因,并发现五粮树海流域生态系统有效降低内农田灌区系统排水中耐药基因丰度,这可能是由于该生态系统内水生动植物的吸附和拦截作用所导致的[10]。本研究在湖州市生态沟渠-1上游阶段水体中发现了不同种类的耐药基因,主要包括mfd、dfrE和tetT,这是由于农田排水中的耐药基因转移至生态沟渠所致。然而,生态沟渠下游阶段水体中mfd、dfrE和tetT的丰度明显低于其上游阶段水体中对应耐药基因的丰度,这证明生态沟渠对农田排水中的耐药基因有着一定的拦截作用。此外,湖州市生态沟渠-1有效降低了农田排水中Proteobacteria门和Verrucomicrobia门的微生物丰度,并提高了其中Actinobacteria门的微生物丰度。生态沟渠中的水生植物和功能设施能够有效的拦截农田排水中的部分颗粒物及其携带的微生物群落,而耐药基因往往是由微生物所携带的,这可能有利于生态沟渠降低农田排水中耐药基因的丰度[6-7]。与此同时,氮磷等营养元素能够促进携带耐药基因的微生物的繁殖,而生态沟渠可以通过有效拦截农田排水中的氮磷来降低耐药微生物在自然环境中的富集以及耐药基因在自然环境中的传播。因此,生态沟渠不仅对农田排水中的氮与磷有着较好的拦截作用,还降低了农田排水中主要耐药基因的丰度,并改变了其中微生物群落的组分,这有利于限制耐药基因在自然环境中的富集与传播。
3. 结论(Conclusion)
浙江省内嘉兴市、绍兴市、台州市和湖州市建设的高标准生态沟渠对农田排水中的氮磷均有着明显的拦截效果,其中40%的生态沟渠能够同时明显降低农田排水中氨氮、总磷和总氮的浓度。嘉兴市生态沟渠对农田排水中氨氮浓度的平均拦截率高达48.1%,而绍兴市生态沟渠对总氮浓度的平均降低率高达48.7%。同时,湖州市生态沟渠-1不仅有效拦截了农田排水中的氮和磷,还明显降低了农田排水中主要耐药基因的丰度(mfd、dfrE和tetT),并改变了其中微生物群落的组分(Proteobacteria门、Verrucomicrobia门和Actinobacteria门微生物)。综上可知,浙江省生态沟渠能够有效拦截农田排水中的氮磷和耐药基因,其高标准的建设模式值得推广,但生态沟渠限制耐药基因传播与扩散的机制仍需进一步研究。
-
图 1 浙江省不同地市生态沟渠对农田排水中氮磷的拦截效果
Figure 1. The interception effects of ecological ditches from cities in Zhejiang province on nitrogen and phosphorus in farmland drainage
图 2 生态沟渠对农田排水中氮磷削减的热图
Figure 2. Heatmap of the nitrogen and phosphorus elimination in farmland drainage by ecological ditches
图 3 生态沟渠下游阶段出水总氮、氨氮及总磷的浓度
Figure 3. The concentrations of total nitrogen, ammonia nitrogen and total phosphorus in downstream effluents of ecological ditches
图 4 湖州市生态沟渠-1不同阶段水体中耐药基因子类数量的Venn图
Figure 4. The venn diagram of ARG subtype numbers in water bodies from different stages of the Huzhou ecological ditch-1
图 5 湖州市生态沟渠-1不同阶段水体中耐药基因种类(a)及主要耐药基因丰度(b)
Figure 5. The diversity (a) and abundance (b) of ARG subtypes in water bodies from different stages of the Huzhou ecological ditch-1
图 6 湖州市生态沟渠-1不同阶段水体中微生物群落组分特征
Figure 6. The characteristic of microbial community compositions in water bodies from different stages of the Huzhou ecological ditch-1
表 1 湖州市生态沟渠-1不同阶段水体样品DNA性质
Table 1. The properties of DNA extracted from different water samples in the Huzhou ecological ditch-1
水体样品 Water Sample DNA浓度/(μg·L−1) DNA Concentration DNA纯度 DNA Purity(OD260/280) DNA总量 Total DNA 生态沟渠上游阶段 31.0 1.81 总量满足宏基因组一次标准建库要求 生态沟渠中游阶段 6.10 2.31 总量满足宏基因组一次标准建库要求 生态沟渠下游阶段 9.70 2.01 总量满足宏基因组一次标准建库要求
下载: 导出CSV
-
[1] DIEBEL M W, MAXTED J T, ROBERTSON D M, et al. Landscape planning for agricultural nonpoint source pollution reduction III: Assessing phosphorus and sediment reduction potential [J]. Environmental Management, 2009, 43(1): 69-83. doi: 10.1007/s00267-008-9139-x [2] 侯朋福, 薛利祥, 周玉玲, 等. 掺混控释肥侧深施对稻田田面水氮素浓度的影响 [J]. 中国土壤与肥料, 2019(1): 16-21. doi: 10.11838/sfsc.1673-6257.18091 HOU P F, XUE L X, ZHOU Y L, et al. The effect of side deep fertilization for resin blending controlled-release fertilizer on nitrogen concentration in surface water of paddy field [J]. Soil and Fertilizer Sciences in China, 2019(1): 16-21(in Chinese). doi: 10.11838/sfsc.1673-6257.18091
[3] JIANG C L, FAN X Q, CUI G B, et al. Removal of agricultural non-point source pollutants by ditch wetlands: Implications for lake eutrophication control [J]. Hydrobiologia, 2007, 581(1): 319-327. doi: 10.1007/s10750-006-0512-6 [4] KÜMMERER K. Antibiotics in the aquatic environment - A review - Part I [J]. Chemosphere, 2009, 75(4): 417-434. doi: 10.1016/j.chemosphere.2008.11.086 [5] ZHOU L J, YING G G, LIU S, et al. Excretion masses and environmental occurrence of antibiotics in typical swine and dairy cattle farms in China [J]. Science of the Total Environment, 2013, 444: 183-195. doi: 10.1016/j.scitotenv.2012.11.087 [6] LU Y, XIAO Y F, ZHENG G Y, et al. Conditioning with zero-valent iron or Fe2+ activated peroxydisulfate at an acidic initial sludge pH removed intracellular antibiotic resistance genes but increased extracellular antibiotic resistance genes in sewage sludge [J]. Journal of Hazardous Materials, 2020, 386: 121982. doi: 10.1016/j.jhazmat.2019.121982 [7] LU Y, LI J M, MENG J, et al. Long-term biogas slurry application increased antibiotics accumulation and antibiotic resistance genes (ARGs) spread in agricultural soils with different properties [J]. Science of the Total Environment, 2021, 759: 143473. doi: 10.1016/j.scitotenv.2020.143473 [8] 张宁, 李淼, 刘翔. 土壤中抗生素抗性基因的分布及迁移转化 [J]. 中国环境科学, 2018, 38(7): 2609-2617. doi: 10.3969/j.issn.1000-6923.2018.07.029 ZHANG N, LI M, LIU X. Distribution and transformation of antibiotic resistance genes in soil [J]. China Environmental Science, 2018, 38(7): 2609-2617(in Chinese). doi: 10.3969/j.issn.1000-6923.2018.07.029
[9] 田其凡, 何玘霜, 陆安祥, 等. 农田土壤抗生素抗性基因与微生物群落的关系 [J]. 环境化学, 2020, 39(5): 1346-1355. doi: 10.7524/j.issn.0254-6108.2019060602 TIAN Q F, HE Q S, LU A X, et al. Relationship between antibiotic resistance genes and microbial communities in farmland soil [J]. Environmental Chemistry, 2020, 39(5): 1346-1355(in Chinese). doi: 10.7524/j.issn.0254-6108.2019060602
[10] SHI W, ZHANG H, LI J J, et al. Occurrence and spatial variation of antibiotic resistance genes (ARGs) in the Hetao Irrigation District, China [J]. Environmental Pollution, 2019, 251: 792-801. doi: 10.1016/j.envpol.2019.04.119 [11] 周俊, 邓伟, 刘伟龙. 沟渠湿地的水文和生态环境效应研究进展 [J]. 地球科学进展, 2008, 23(10): 1079-1083. doi: 10.3321/j.issn:1001-8166.2008.10.010 ZHOU J, DENG W, LIU W L. Advances on the effects of ditch wetland on hydrology and eco-environment [J]. Advances in Earth Science, 2008, 23(10): 1079-1083(in Chinese). doi: 10.3321/j.issn:1001-8166.2008.10.010
[12] 于淼, 马国胜, 赵昌平, 等. 氮磷生态拦截集成技术治理湖泊岸区农业面源污染分析研究 [J]. 环境科学与管理, 2015, 40(1): 72-74. doi: 10.3969/j.issn.1673-1212.2015.01.019 YU M, MA G S, ZHAO C P, et al. Analysis on controlling rural non-point pollution in lake shore area by eco-retain of nitrogen and phosphorus integrated technology [J]. Environmental Science and Management, 2015, 40(1): 72-74(in Chinese). doi: 10.3969/j.issn.1673-1212.2015.01.019
[13] 许明宸, 王逸超, 张文艺, 等. 生态沟渠净化稻田排水动力学分析和生物相特征 [J]. 环境化学, 2021, 40(2): 592-602. doi: 10.7524/j.issn.0254-6108.2019092701 XU M C, WANG Y C, ZHANG W Y, et al. Dynamics analysis and biofacies characteristics of drainage from paddy fields purified by ecological ditches [J]. Environmental Chemistry, 2021, 40(2): 592-602(in Chinese). doi: 10.7524/j.issn.0254-6108.2019092701
[14] GILL S L, SPURLOCK F C, GOH K S, et al. Vegetated ditches as a management practice in irrigated alfalfa [J]. Environmental Monitoring and Assessment, 2008, 144(1/2/3): 261-267. [15] SMITH D R. Assessment of in-stream phosphorus dynamics in agricultural drainage ditches [J]. Science of the Total Environment, 2009, 407(12): 3883-3889. doi: 10.1016/j.scitotenv.2009.02.038 [16] LU Y, SUN R H, ZHANG C G, et al. In situ analysis of antibiotic resistance genes in anaerobically digested dairy manure and its subsequent disposal facilities [J]. Bioresource Technology, 2021, 333: 124988. doi: 10.1016/j.biortech.2021.124988 [17] LI E H, LI W, WANG X L, et al. Experiment of emergent macrophytes growing in contaminated sludge: Implication for sediment purification and lake restoration [J]. Ecological Engineering, 2010, 36(4): 427-434. doi: 10.1016/j.ecoleng.2009.11.009 [18] 王丽莎, 李希, 甘蕾, 等. 亚热带丘陵区湿地水生植物组合模式拦截氮磷的研究 [J]. 生态环境学报, 2017, 26(9): 1577-1583. WANG L S, LI X, GAN L, et al. Study on the aquatic plant combination patterns for intercepting nitrogen and phosphorus in wetland of subtropical hilly region [J]. Ecology and Environmental Sciences, 2017, 26(9): 1577-1583(in Chinese).
[19] LACOUL P, FREEDMAN B. Relationships between aquatic plants and environmental factors along a steep Himalayan altitudinal gradient [J]. Aquatic Botany, 2006, 84(1): 3-16. doi: 10.1016/j.aquabot.2005.06.011 [20] 陈英, 邱学林, 吴钰明. 太湖流域农田生态沟渠塘不同水生植物组合净化氮磷效果研究 [J]. 江苏农业科学, 2015, 43(12): 367-369. CHEN Y, QIU X L, WU Y M. Study on nitrogen and phosphorus removal efficiency of different aquatic plant combinations in farmland ecological ditch pond of Taihu Lake Basin [J]. Jiangsu Agricultural Sciences, 2015, 43(12): 367-369(in Chinese).
[21] 田昌, 陈敏, 周旋, 等. 生态沟渠对小流域农田排水中氮磷的拦截效果研究 [J]. 中国土壤与肥料, 2020(4): 186-191. doi: 10.11838/sfsc.1673-6257.19317 TIAN C, CHEN M, ZHOU X, et al. Effects of ecological ditch on interception of nitrogen and phosphorus in farmland drainage in small watershed [J]. Soil and Fertilizer Sciences in China, 2020(4): 186-191(in Chinese). doi: 10.11838/sfsc.1673-6257.19317
-

 点击查看大图
点击查看大图
计量
- 文章访问数: 2678
- HTML全文浏览数: 2678
- PDF下载数: 115
- 施引文献: 0

