



-
饮用水安全问题对人类健康至关重要[1-2]。近些年饮用水安全方面的挑战涵盖了微生物的方方面面,通常细菌在饮用水管网输配系统(drinking water distribution systems,DWDSs)的生物膜中自然定殖,并形成稳定的微生物生态系统[3-4],这种稳态环境有利于细菌的生长并能够给其在面对消毒过程时提供一定的庇护作用,进而带来包括致病菌、抗性基因等微生物风险问题[5]。因此,探究长期运行的DWDSs生物膜的细菌群落组成对于控制饮用水微生物风险具有重要意义。
抗生素抗性基因(antibiotic resistance genes,ARGs)会增强细菌宿主对抗生素的抵抗能力[6-7],细菌产生ARGs通常需要一个稳定的环境,而生物膜恰恰能提供这种“温床”[8-9],因此长期运行的DWDSs生物膜最终可能导致微生物的ARGs风险。此外,包括磺胺甲恶唑在内的抗生素在临床上的广泛使用,不可避免地导致抗生素污染[10-11]。抗生素不仅会影响细菌的生长过程,还会对ARGs的传播造成影响[12-13]。低剂量的抗生素(0.5 mg·L−1),如土霉素和磺胺甲恶唑,会加速活性氧的生成并促进ARGs的传播[14]。因此,探究微量抗生素对长期运行的DWDSs生物膜中ARGs的影响十分重要。饮用水处理和消毒被认为是20世纪最伟大的公共卫生成就之一[15],消毒工艺通常被认为能够抑制微生物的生长以及控制ARGs传播[16]。但同样有研究报道消毒对ARGs的形成存在促进作用[17-20]。因此,探究消毒是否对长期运行的DWDSs生物膜中的ARGs起到有效控制将有助于控制管网微生物风险。此外,消毒过程作为细菌遭受到的一种环境胁迫压力,往往会影响与细菌新陈代谢有关的功能基因[21]。但是关于功能基因与ARGs之间是否存在关系,目前研究报道不多。
因此,本研究的目的是利用模拟不同处理条件下的DWDSs,研究微量磺胺甲恶唑对细菌群落组成和ARGs的影响;通过长期模拟运行探究次氯酸钠消毒对ARGs的控制效果;最后通过相关性分析探讨功能基因与ARGs之间可能存在的关系,探讨消毒过程通过改变细菌的代谢过程从而影响细菌的生长进而控制ARGs传播。
-
实验用水来自于中国东部一个饮用水处理厂(drinking water treatment plant,DWTP)。出水经过混凝、沉淀、砂滤处理。该原水的pH为7.89±0.05,浊度为(1.24±0.10) NTU,耗氧量CODMn为(3.00±0.14) mg·L−1,余氯质量浓度为0,异养菌平板计数为(120±5) CFU·mL−1,没有检测到总大肠菌群。
-
实验在1.50 m长全新的PE管材(DN 60 mm)中进行。管道进水为每日从DWTP处理后获取的60.0 L砂滤(SF)出水,并通过蠕动泵以1.50 mL·s−1的流速在各管中缓慢流过,运行为期1 a。
共采用9根管道来模拟饮用水管网输配系统(DWDSs),共设计3种实验条件,各条件有3个平行。条件A1,不使用消毒剂的SF出水作为实验进水;条件A2,在不使用消毒剂的SF出水添加2.00 μg·L−1的磺胺甲恶唑作为实验进水。条件B1,SF出水添加2.00 μg·L−1的磺胺甲恶唑并使用次氯酸钠(NaClO)消毒剂进行消毒作为实验进水。其中,磺胺甲恶唑的添加质量浓度是考虑到了可能发生的磺胺类抗生素污染情况[22];而以游离氯消毒的出厂水质量浓度一般控制在0.3~0.8 mg·L−1[23-24],因此,实验模拟管网进水游离氯质量浓度控制在0.50 mg·L−1。
-
在模拟的DWDSs运行1 a后,将每根PE管切成3段,每段长约50.0 cm。使用无菌金属刮刀从管段内部采集生物膜,刮刀距离切口至少40.0 cm,以尽量减少污染的风险[25]。使用FastDNA Spin Kits for Soil (MP Biomedicals Co., USA)试剂盒的说明进行DNA提取。用NanoDrop 2000(Thermo Fisher Scienrific., USA)和TBS-380(YPH-Bio., China)分别测量DNA质量浓度和纯度,并进行凝胶测试以验证其完整性(1%琼脂糖凝胶电泳)。每根管道3个DNA样品混合为1个样品,每个条件3个平行共3个DNA样品进行送样。
-
Illumina高通量测序是用HiSeq 2000平台进行的。宏基因组序列库由美吉生物公司构建。使用Fastp软件(http://github.com/OpenGene /fastp)对原始fastq文件进行了过滤。使用IDBA-UD/Megahit(https://github.com/voutcn/megahit)和Newbler(https://ngs.csr.uky.edu/Newbler)对序列进行多重混合拼接组装。使用Prodigal对拼接结果中的contigs进行基因预测。使用CD-HIT软件(http://www.bioinformatics.org/cd-hit/)构建非冗余基因集。
微生物群落组成分析利用NR数据库进行比对(参数blastp;E≤10−5),并通过NR库对应的分类学信息数据库获得物种注释结果,然后使用物种对应的基因丰度总和计算该物种的丰度。功能基因与京都基因与基因组百科全书(KEGG)的基因数据库进行比对(参数blastp;E≤10−5),再根据对应的基因丰度总和计算该功能类别的丰度。抗生素抗性基因(ARGs)与抗性基因数据库(CARD)进行比对(参数blastp;E≤10−5),将目标基因与其耐药功能注释信息结合,获得注释结果。以上基因丰度是用TPM(trans per million),即基因丰度以该基因通过基因长度标准化后的基因丰度在该样本中所有基因长度标准化后的基因丰度总和中占比的百万倍表示。
-
1)微生物群落组成。如图1(a)所示,当在DWDSs的进水中加入磺胺甲恶唑时,由TPM计算的生物膜中门分类水平微生物群落的总丰度仅改变了0.09%。丰度前十的门分别为放线菌门、变形菌门、浮霉菌门、拟杆菌门、酸酐菌门、疣微菌门、蓝藻门、厚壁菌门、芽单胞菌门以及硝化螺旋菌门。在不添加消毒剂的情况下,放线菌门和变形菌门的细菌在条件A1和A2的生物膜中占主导地位,丰度总和占总细菌的比例分别为83.1%和83.7%。许多研究表明,这2个门类是不同管道生物膜中的主要的细菌类型[26-27]。而他们在B1中仅占比24.5%,因此,消毒可以明显降低放线菌门与变形菌门的细菌在细菌群落中的比例。但同时,浮霉菌门细菌无论是占比(58.1%)还是相对丰度(TPM=70 099.6),均远远高于未消毒条件A1和A2,表明浮霉菌门具有较强的抗次氯酸钠消毒能力。厚壁菌门的细菌在3种条件(A1、A2、B1)中的丰度依次为6 839.6、6 607.8和6 802.1 TPM,消毒剂的加入并没有显著影响其丰度变化,因此,厚壁菌门的细菌也具有一定的抗消毒能力,这一结果与之前的研究结果类似[26]。
种水平上未消毒条件下(A1、A2)的优势种中(图1(b)),酸微菌Acidimicrobium sp.BACL27 MAG-120823-bin4在加入磺胺甲恶唑后其相对丰度值由64 879.8 TPM(A1)升高至80 593.3 TPM(A2),相应地其相对丰度占比由7.52%上升至9.34%。因此,微量磺胺甲恶唑对生物膜中该物种丰度产生影响。而消毒条件下(B1),酸微菌Acidimicrobium sp.BACL27 MAG-120823-bin4丰度仅占B1条件生物膜中所有细菌的0.07%,酸微菌属于放线菌门,而该条件下丰度前10的细菌均属于浮霉菌门(图1(c))。因此,消毒可以很好的控制包括酸微菌在内的放线菌门细菌生长,而浮霉菌门的细菌能够更好地适应消毒环境,对消毒表现出一定的抗性。
2)物种的相似性与差异性。A1与A2条件下的群落物种不仅在丰度上差异很小,在物种种类相似程度也很高,相同的物种占到了总物种的97%以上(图2(a))。因此,添加磺胺甲恶唑对微生物群落组成的改变非常小。这一结果与之前的一项研究类似,即磺胺甲恶唑在环境相关质量浓度(< 4 μg·L−1)下没有明显影响河流沉积物中的细菌群落组成[28]。但消毒剂的加入改变了群落的组成,物种数量由12 330种下降至8 374种,因此,次氯酸钠消毒在水处理中确实对微生物生长能够起到一定的控制效果。
如图2(b)所示,在群落的差异性物种中,消毒条件B1与其他2种条件表现出的显著差异性物种均为浮霉菌纲细菌,包括Zavarzinella formosa,Planctomycetaceae bacterium SCGC AG-212-F19,Gemmata_obscuriglobus,Gemmata sp.IIL30,Singulisphaera acidiphila,Gemmata sp.SH-PL17,Schlesneria paludicola,这进一步证实浮霉菌纲细菌抗次氯酸钠消毒能力强。根据研究表明,浮霉菌在面对臭氧的胁迫时也体现了强适应能力,凭借出芽繁殖的方式保障细菌的正常繁殖生长[29]。而其余物种均属于放线菌与变形门细菌,这些物种在A1、A2中生长良好,但一旦长期暴露在次氯酸钠消毒条件中,则会遭到抑制。因此,次氯酸钠消毒对于放线菌与变形门类细菌控制有效[30],但对于浮霉菌几乎没有控制效果。
-
1)群落的抗性基因。与群落组成类似,添加2 μg·L−1磺胺甲恶唑对生物膜上抗性基因丰度影响不明显(图3(a))。在生物膜细菌群落的抗性基因中,Saur_rpoC_DAP、macB、mfd、aminocoumarin resistant alaS、Mtub_katG_INH、mtrA、oleC、Mtub_ropB_RIF、Ecol_ropB_RIF、patA是主要的ARGs类型(图3(a))。在同样添加磺胺甲恶唑的管道中,采取次氯酸钠消毒使得ARGs总量下降了91.9%,这表明消毒对于抗性基因有很好的控制效果。在差异性基因中,消毒条件中丰度占比较高的抗性基因分别是macB、bcrA、patA、mdtB与drrA(图3(b))。而携带抗性基因的细菌主要是Acidimicrobium sp. BACL27 MAG-120823-bin4、Actinobacteria bacterium BACL15 MAG-120619-bin91、alpha proteobacterium HIMB114、Actinobacteria bacterium BACL2 MAG-120802-bin41、actinobacterium SCGC AAA027-L06(图3(c))。这些细菌在各条件中的丰度也处于一个较高的水平,因此,产生抗性基因的一个原因可能是细菌本身能很好的适应生存环境。这一点在携带差异性基因的物种上也有很好的体现,例如未消毒条件中携带差异性抗性基因(macB、patA)的细菌Acidimicrobium sp.BACL27 MAG-120823-bin4和Actinobacteria bacterium BACL15 MAG-120619-bin91在消毒后数量明显下降(图1(b)),该2种抗药基因消毒后同样下降明显。同时,抗性基因与物种的相关性分析也进一步表明物种与抗性基因在不同条件下的变化存在很好的相关性(R2 > 0.8)( 图3(d))。因此,以上结果说明消毒对抗性基因的控制效果很大程度体现在对细菌的控制上。
2)群落功能基因。微生物群落的功能基因包括碳水化合物代谢、氨基酸代谢、能量代谢、辅助因子和维生素的代谢、核苷酸代谢、脂质代谢、信号转导、复制和修复以及翻译等(图4(a))。除信号转导外,部分功能基因的占比在加入磺胺甲恶唑后略有上升。而NaClO消毒使这些功能基因明显减少,这一结果与以前的研究结果一致[31]。
另外,对不同功能基因贡献度靠前的细菌与携带耐药性基因的细菌几乎一致(图4(b))。在A1、A2与B1群落的功能基因组成中,碳水化合物代谢类的功能基因占比分别为13.8%、13.9%和15.8%,在消毒处理后细菌的碳水化合物代谢基因水平出现了上升,说明面对消毒带来的环境胁迫压力时,生物膜中细菌可以通过调整碳水化合物相关过程的代谢来进行响应;与此类似的还有辅助因子与维生素相关的代谢,群落在面临消毒时同样出现了上调的现象,A1、A2和B1条件下该功能基因占比分别为5.47%、5.45%和6.75%。与此同时,功能基因与物种的相关性分析同样说明不同条件下物种丰度与功能基因变化表现出很好的相关性 (R2>0.8)(图4(c))。因此,细菌抗性基因的变化极有可能是与功能基因的变化趋势一致。
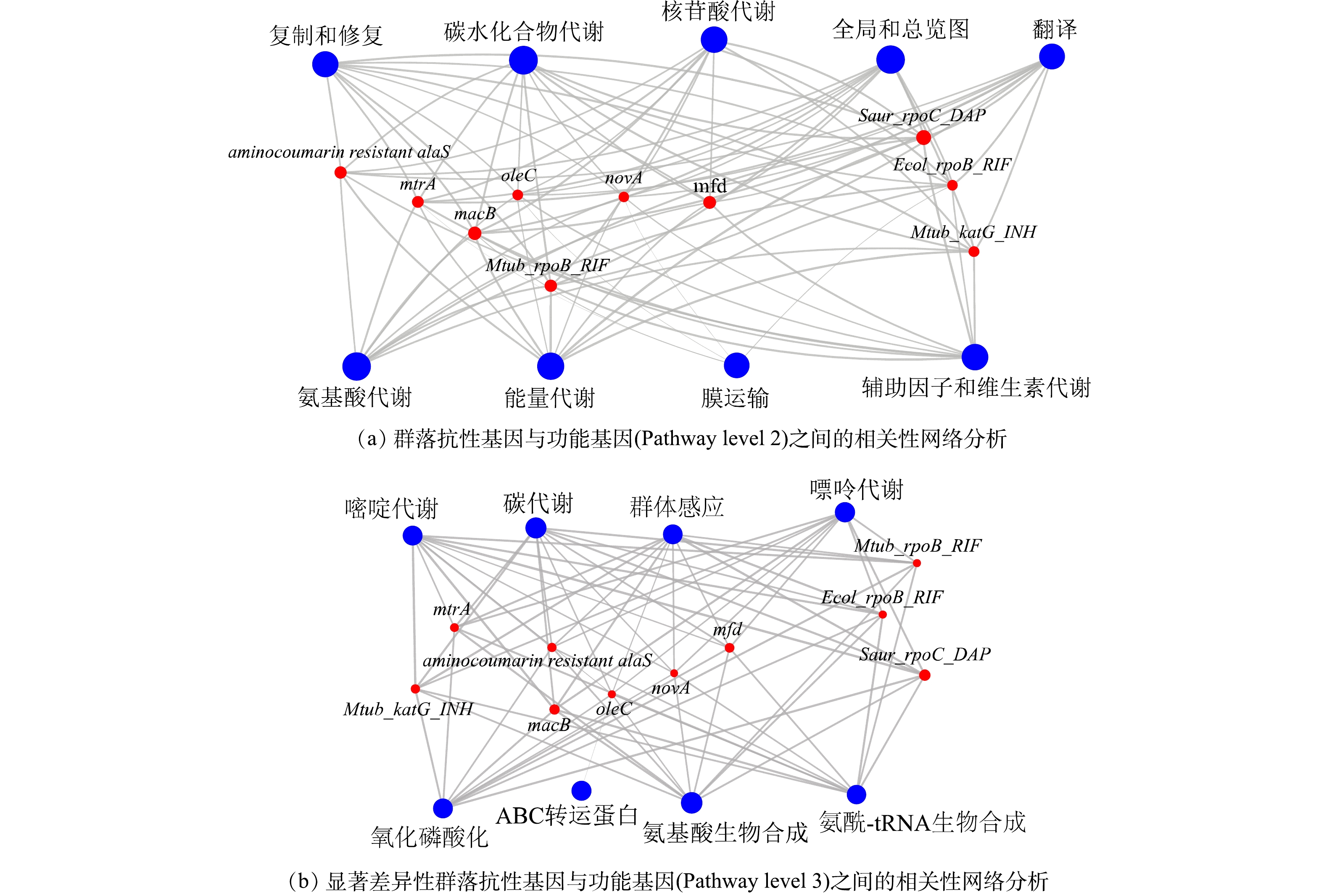
3)群落功能基因与抗性基因之间的关系。对丰度前十的群落功能基因在KEGG二级通路上与抗性基因进行相关性网络分析后,我们发现抗性基因与每一大类的功能基因之间存在不同程度的正相关性(图5(a))。在对显著性差异的ARGs与KEGG三级通路的功能基因进一步分析后,发现存在显著性差异的抗性基因与功能基因((图3(d)与(图4(d))之间仍然存在相似结果,即每一种抗性基因均与各种的功能基因有着不同程度的正相关关系(图5(b))。这一结果表明次氯酸钠是通过改变物种并影响其功能基因进而改变抗性基因的丰度。很多研究中都报道了功能基因对于耐药性基因的形成存在影响,例如中心碳代谢功能影响结核分枝杆菌的耐药性[32],群体感应影响耐药性基因的产生[33]。与此同时,当环境压力改变细菌群落组成时[34],物种功能基因往往也会进行改变[35]。因此,当次氯酸钠消毒显著引起生物膜群落变化时,群落中微生物的功能基因也发生了相应的变化,最终也会影响其耐药性基因的形成。
在图5(b)中,抗性基因oleC是唯一与ABC转运蛋白功能基因呈正相关关系。细菌在某些应激条件下产生细菌素和微毒素会对周围物种产生一定的抗菌活性以便自身生存,而ABC转运蛋白能够帮助细菌通过免疫蛋白或者转运等方式保护自己[36]。在A1、A2中oleC主要存在于放线菌门的细菌中,在添加磺胺后oleC的丰度占比上升了0.14%(图3(b)),但放线菌门的细菌的丰度却出现了下降(图1(a))。因此,磺胺甲恶唑可能作为外源性刺激物质造成ABC转运蛋白功能水平的上升,进而赋予携带oleC抗性基因物种更强的生存优势并最终使得oleC抗性基因保持更高的丰度水平,同时研究证明由oleC的抗生素抗性效果源自ABC-转运体的外排转运系统[37]。但由于放线菌门的细菌并不能适应长期的消毒,导致其细菌丰度下降,进而使得功能基因与抗性基因也出现相同的趋势。因此,oleC抗性基因的产生与细菌功能基因的关系紧密。除oleC外,其他抗性基因均与各类功能基因存在正相关性,包括氨基酸生物合成,碳代谢,嘌呤代谢,嘧啶代谢,氧化磷酸化,群体感应以及氨基酸-tRNA的生物合成等。早先的研究证明了不同的输配水系统条件所施加的选择压力影响群落结构以及功能特性和代谢潜力水平的变化[38],因此,当群落处于长期消毒时,群落的组成发生变化,上述类型的功能基因与抗性基因也随之变化。
-
1) 2 μg·L−1磺胺甲恶唑的添加对生物膜细菌群落结构组成无明显影响,次氯酸钠消毒对不同细菌包括变形菌门和放线菌门有很好的控制效果,但浮霉菌门与厚壁菌门细菌表现出很强的抗次氯酸钠消毒能力。
2)在未消毒条件中丰度前10的抗性基因(包括Saur_rpoC_DAP、macB、mfd、aminocoumarin resistant alaS、Mtub_katG_INH、mtrA、oleC、Mtub_ropB_RIF、Ecol_ropB_RIF、patA)在次氯酸钠消毒后含量下降明显,ARGs总量下降了91.9%,同时携带差异性抗性基因(macB、patA)的细菌在消毒后数量也出现明显下降,因此,次氯酸钠消毒通过控制携带抗性基因物种从而有效控制抗性基因传播。
3)抗性基因的形成与细菌功能代谢变化有关。组间显著性差异的功能基因(包括氨基酸生物合成、碳代谢、嘌呤代谢、嘧啶代谢、氧化磷酸化、群体感应以及氨基酸-tRNA的生物合成)与组间显著性差异的抗性基因存在良好的正相关关系。因此,长期消毒改变了细菌群落组成及其功能,并最终影响抗性基因传播。
微量磺胺甲恶唑对饮用水管网生物膜群落及抗性基因的影响与控制
Influence of micro-sulfamethoxazole on the biofilm bacterial communities and antibiotic resistance genes in drinking water distribution systems and its control
-
摘要: 针对饮用水管网系统可能存在的微生物风险问题,采用模拟不同处理条件下的输配水管道系统,通过宏基因组学分析探究微量磺胺甲恶唑以及次氯酸钠消毒对管道中生物膜与抗性基因组成的影响。结果表明,2 μg·L−1磺胺甲恶唑的添加对微生物群落以及抗性基因组成无明显影响,而浮霉菌门细菌表现出很强的抗次氯酸钠消毒能力。在未消毒条件下丰度前十的抗性基因与携带差异性抗性基因的细菌在消毒后丰度均明显有所下降,次氯酸钠消毒使ARGs总量下降了91.9%,因此,次氯酸钠消毒通过控制携带抗性基因物种从而有效控制群落抗性基因的传播。同时,通过组间显著性差异的功能基因与组间显著性差异的抗性基因相关性分析,功能基因的变化情况与抗性基因变化情况一致,因此,长期消毒改变了细菌群落组成及其功能,并最终影响抗性基因传播。这项研究有助于控制长期运行的饮用水管网输配系统中可能存在的包括抗药基因在内的微生物相关风险问题。Abstract: For the microbial risk in the water of drinking water distribution systems (DWDSs), metagenomic analysis was used to investigate the effects of micro-sulfamethoxazole and NaClO disinfection on biofilm bacterial communities and antibiotic resistance genes (ARGs) in the simulated DWDSs under different treatments. The experimental results showed that the bacterial communities and ARGs did not change significantly with addition of 2 μg-L−1 sulfamethoxazole, while the bacteria of Planctomycetes at phylum level showed a strong resistance to NaClO. The abundance of top ten ARGs and the bacteria carrying differential ARGs detected before disinfection both decreased significantly after disinfection. The total abundance of ARGs decreased by 91.9% due to the influence of NaClO. Therefore, the spread of ARGs was controlled by the NaClO disinfection because of its inhibition on the bacterial species carrying ARGs growth. Meanwhile, the correlation analysis between function genes and ARGs with significant differences among groups showed that the changes in function genes were consistent with changes in ARGs. Therefore, long disinfection changed the bacterial communities and their function, which affected the propagation of different ARGs. This study was contributed to the control of possible microbial risk including the ARGs in the water of long-running DWDSs.
-
微囊藻毒素(microcystins,MCs)是一类由蓝藻产生的环状七肽天然毒素,其中微囊藻毒素-LR(MC-LR)是毒性最强、分布最为广泛的MCs变体之一[1-2]. 夏季富营养化的湖泊容易发生蓝藻暴发从而产生较高浓度的MCs,这会给人体健康和生态环境造成危害. MCs的化学结构稳定,在天然水体中虽然能在光照和微生物的作用下降解,但降解缓慢[3];因此需要采取一些人工手段进行水体治理以去除MCs,减少MCs带来的危害和风险. 常用的MCs去除方法通常应用于水处理厂,如活性炭吸附、混凝沉淀、化学氧化、超滤、生物膜反应器等[3- 4],若在湖泊水体中开展大规模治理,上述方法会有诸多限制. 生物炭由于原料来源广泛、制备方便、成本低廉,有较强的污染物吸附能力,此外还具有固碳减排、改良土壤或底质等多重环境效益,近年来在环境领域受到越来越多的研究和应用[5-6]. 因此,用生物炭来吸附去除水体中MCs是较为可行的方法.
制备生物炭的原料有农林植物废料、畜禽粪便、餐厨垃圾、沼渣污泥等,水生植物残体也是制备生物炭的常见原料[7]. 水生植物是湿地生态系统的重要组成部分,具有分布广、产量大、生长快的特点;但到秋冬季节,水生植物往往会枯萎,残体若不及时处理,会腐烂释放出营养物质造成水体污染[8]. 因此,将水生植物制成生物炭再投放到水体吸附MC-LR,既实现水生植物资源化利用,又能改良水体.
直接热解得到的生物炭的吸附性能通常十分有限,往往不能满足实际工作中去除特定污染物的应用需求[9]. 为了提升其吸附性能,可通过改性来改善其理化性质[10]. 氯化镁由于无毒害、腐蚀性小、成本低,是一种较为理想的改性剂[11]. 将氯化镁用于生物炭改性去除水体中的无机营养盐和重金属有较多的研究,镁改性能在生物炭表面负载含镁矿物,这能增强静电吸引、离子交换、表面络合、化学沉淀从而加强对无机污染物的吸附性能[12-14]. 虽然将镁改性生物炭用于有机污染物的去除研究较少,但Tao等研究认为镁改性可用于提升对可离子化有机污染物的吸附性能[15].
目前,利用水生植物生物炭去除MC-LR的研究较少,而利用镁改性生物炭吸附MC-LR的研究未见相关报道. 本研究在这一方面做新的尝试,利用2种常见的水生植物苦草(Vallisneria natans)和狐尾藻(Myriophyllum verticillatum)制备氯化镁改性生物炭,开展其吸附MC-LR的研究,结合生物炭的表征和吸附特性,探索其改性和吸附机理.
1. 材料与方法(Materials and methods)
1.1 材料和仪器
材料:苦草和狐尾藻采集自江苏省无锡市太湖贡湖湾湿地公园.
试剂:MC-LR(≥95%)购自Taiwan Algal Science Inc.;氯化镁(AR)购自上海凌峰化学试剂有限公司;微囊藻毒素检测试剂盒购自Beacon Analytical Systems Inc.;其他试剂均为分析纯.
仪器:酶标仪(Tecan Infinite 200 PRO)、全自动比表面及孔隙度分析仪(Micromeritics ASAP 2460)、有机元素分析仪(Elementar vario EL Ⅲ)、傅里叶变换红外光谱仪(Nicolet Nexus 870)、X射线光电子能谱仪(Thermo Fisher Nexsa)、X射线衍射仪(Thermo X'TRA)、马弗炉、pH计、恒温摇床等.
1.2 实验方法
1.2.1 生物炭的制备
先将采集到的苦草和狐尾藻清洗,去除表面泥沙,晒干并60 ℃烘干至恒重,粉碎至小于20目. 将植物碎末填充到坩埚中,加入与植物碎末质量(g)等数值体积(mL)的MgCl2溶液浸渍24 h,分别设置0.5、1.0、2.0、4.0 mol·L−1的MgCl2浸渍浓度. 然后将坩埚置于马弗炉中以3 ℃·min−1升温速率加热至500 ℃,维持3 h热解. 热解完成后冷却至室温,用超纯水清洗、烘干后研磨过筛,取100—200目颗粒并再次清洗烘干,得到粒度较为均一的氯化镁改性生物炭. 用K指代苦草,H指代狐尾藻,按照浸渍浓度分别将镁改性苦草生物炭分别标记为KB-0.5Mg、KB-1.0Mg、KB-2.0Mg、KB-4.0Mg;镁改性狐尾藻生物炭分别标记为HB-0.5Mg、HB-1.0Mg、HB-2.0Mg、HB-4.0Mg.
对应地制备500 ℃热解未改性生物炭KB、HB,方法见本课题组先前的研究[16].
1.2.2 生物炭的表征
全自动比表面积孔隙度分析仪测定77 K温度下生物炭对N2的吸附等温线;有机元素分析仪测定生物炭中C、H、N、O含量(其中O元素用氧模式测定);KCl压片法测定生物炭的傅里叶变换红外光谱(FTIR),波数区间为4000—400 cm−1;X射线光电子能谱仪对生物炭进行XPS全谱扫描(结合能1350—0 eV)和特定元素精细谱的扫描;对生物炭进行X射线衍射(XRD),扫描范围5°—90°,速率10(°)·min−1;采用pH漂移法测定生物炭的零电荷点(pHpzc)[17-19].
1.2.3 吸附实验
(1)吸附去除率
为了使体系的pH在吸附过程中维持稳定,以pH 7磷酸缓冲液(使用0.001 mol·L−1 NaH2PO4配制,并用NaOH调节至pH 7)作为背景溶液配制10 μg·L−1的MC-LR溶液. 取2.0 mg生物炭添加到上述50 mL MC-LR溶液中,用螺口玻璃瓶盛装,密封后以180 r·min−1转速25 ℃恒温振荡24 h后取样.
(2)吸附动力学
取4.0 mg生物炭添加到上述100 mL 10 μg·L−1 MC-LR溶液中,密封后以180 r·min−1转速25 ℃恒温振荡,每间隔一段时间进行取样.
(3)吸附等温线
以pH 7磷酸缓冲液为背景溶液配制0—100 μg·L−1不同浓度的MC-LR溶液,取2.0 mg生物炭添加到上述不同初始浓度50 mL MC-LR溶液中,密封后以180 r·min−1转速恒温(分别设置25 ℃和35 ℃)振荡72 h后取样.
(4)pH的影响
配制10 μg·L−1 MC-LR溶液,以0.001 mol·L−1NaH2PO4为背景溶液,并用H2SO4/NaOH调节pH为4、6、8、10. 取2.0 mg生物炭添加到50 mL上述不同pH的MC-LR溶液中,密封后以180 r·min−1转速25 ℃恒温振荡24 h后取样.
(5)溶解性有机质(DOM)的影响
用富里酸(CAS: 479-66-3)、单宁酸(CAS: 1401-55-4)、没食子酸(CAS: 149-91-7)做为模式DOM,探讨不同分子量大小DOM对吸附的影响. 配制10 μg·L−1 MC-LR溶液,以pH 7磷酸缓冲液为背景溶液,分别添加上述DOM使溶液中DOM浓度为2 μmol·L−1. 取2.0 mg生物炭添加到50 mL上述含有不同DOM的MC-LR溶液中,密封后以180 r·min−1转速25 ℃恒温振荡24 h.
上述所有吸附实验均进行平行实验,所有样品取约2 mL用0.45 μm针头滤器过滤,将滤液保存在玻璃小瓶中,测定溶液在吸附前后MC-LR浓度.
1.2.4 MC-LR测定
使用酶联免疫吸附法(ELISA)测定MC-LR,利用Beacon公司生产的ELISA试剂盒进行检测. 若样品浓度较高超过试剂盒量程(2.0 μg·L−1),先对样品进行适当比例稀释,再依据说明书的操作流程进行加样、孵育、洗版、显色,用酶标仪测定450 nm吸光度最终计算MC-LR浓度,检测限为0.1 μg·L−1. 所有样品及平行样品均进行复孔测定.
2. 结果与讨论(Results and discussion)
2.1 生物炭表征
2.1.1 比表面和孔隙度分析
对未改性生物炭KB、HB和镁改性生物炭KB-2.0Mg、HB-2.0Mg进行N2吸附等温线(77 K)的测定,利用BET模型计算生物炭的比表面积SBET,根据等温线P/P0≥0.99处的N2吸附量计算生物炭的总孔孔容Vtot,利用DFT模型计算生物炭的孔径分布. 根据孔径分布计算出微孔(孔径≤2 nm)孔容Vmic和中孔(2 nm<孔径≤50 nm)孔容Vmes. 其中未改性生物炭的部分表征结果引自本课题组先前的研究[16].
从孔径分布图(图1)可以看出,镁改性生物炭在不同孔径下对应的孔容均高于未改性生物炭. 表1 也表明,镁改性生物炭比表面积和孔容明显高于未改性生物炭,且中孔的增加明显,说明氯化镁改性能够促进生物炭孔隙的形成. 研究表明,氯化镁改性能够使生物炭表面产生鳞片状MgO颗粒,该结构有助于提升生物炭的比表面积[20]. 此外,MgCl2在高温下能与H2O反应,形成Mg(OH)2、Mg(OH)Cl、MgO等产物,并释放HCl气体,该过程会改变生物炭的孔隙结构,促进孔容的增加[21-22].
 图 1 苦草(a)和狐尾藻(b)生物炭的孔径分布Figure 1. The pore size distribution of Vallisneria (a) and Myriophyllum (b) biochar表 1 生物炭的比表面和孔径分析Table 1. Specific surface and pore size analysis of biochar
图 1 苦草(a)和狐尾藻(b)生物炭的孔径分布Figure 1. The pore size distribution of Vallisneria (a) and Myriophyllum (b) biochar表 1 生物炭的比表面和孔径分析Table 1. Specific surface and pore size analysis of biochar生物炭Biochar SBET /(m2·g−1) Vtot /(cm3·g−1) Vmic /(cm3·g−1) Vmeso /(cm3·g−1) KB 7.47 2.89×10−2 1.89×10−3 1.36×10−2 HB 7.64 1.93×10−2 2.17×10−3 9.29×10−3 KB-2.0Mg 58.7 1.19×10−1 8.69×10−3 6.05×10−2 HB-2.0Mg 88.7 1.46×10−1 1.81×10−2 6.66×10−2 注:数据保留3位有效数字. 未改性生物炭的SBET、Vmic、Vmes和Vtot数据引自文献[16]. The data retains 3 significant digits. The SBET, Vmic, Vmes and Vtot are quoted from previously published literature. | Show Table DownLoad:
CSV
DownLoad:
CSV
2.1.2 元素分析
生物炭的元素分析结果见表2. 其中C、H、O、N元素含量由元素分析仪测定得到的质量分数,而Mg元素含量由XPS全谱扫描分析得到的原子数占比. 通常,O/C和(N+O)/C可反映生物炭的亲水性和极性,O/C和(N+O)/C的值越大表示亲水性和极性越强;H/C可反映生物炭的芳香性,值越小则芳香性越强[23-24]. 结果显示,氯化镁改性使生物炭的极性和亲水性增加,而芳香性降低. 改性苦草生物炭中Mg元素含量比改性狐尾藻生物炭更高,可见不同植物材料会影响改性过程中Mg元素的负载.
表 2 生物炭元素分析Table 2. Elemental analysis of biochar生物炭Biochar C/% H/% N/% O/% Mg/% O/C (N+O)/C H/C KB 51.92 2.56 2.73 18.24 0.67 0.263 0.309 0.592 HB 65.40 3.12 2.56 16.78 0.97 0.192 0.226 0.572 KB-2.0Mg 39.93 2.71 2.46 15.53 4.30 0.292 0.345 0.814 HB-2.0Mg 58.86 3.10 2.25 17.93 2.01 0.228 0.261 0.632 注:C、H、N、O的含量为质量分数,Mg含量为原子数占比;O/C,(N+O)/C,H/C为原子数比;表中KB和HB的C、H、N、O数据引自先前发表的文献[16]. The content of C, H, N, O is the mass fraction, and the Mg content is the atomic ratio; O/C, (N+O)/C, H/C are the atomic ratio; the C, H, N, O data of KB and HB in the table are quoted from previously published literature. | Show TableDownLoad:
CSV
2.1.3 官能团和元素形态分析
FTIR表征结果见图2. 波数3694—3696 cm−1处尖锐的峰是孤立的—OH伸缩振动引起的,这可能来源于Mg(OH)2中的O—H基团[25-26];3356—3377 cm−1处的宽峰是氢键缔和的—OH伸缩振动[25, 27];1695—1727 cm−1处的峰是酮、醛、羧基或内酯基中C=O伸缩振动[28];1576—1580 cm−1处较强的峰是羧基COO的反对称伸缩振动[29], 1399—1417 cm−1处的弱峰是羧基COO的对称伸缩[29];1034—1035 cm−1处的峰可归因于C—O—C对称伸缩振动[28-29],但也可能是蒙脱石、伊利石等无机硅酸盐类矿物的特征峰[30];镁改性生物炭在467—471 cm−1出现新的峰,可归因于MgO或Mg(OH)2中Mg—O键的伸缩振动[31-33]. 上述FTIR结果表明,生物炭以及改性生物炭表面含有羧基、羟基等多种含氧官能团,且氯化镁改性使生物炭表面负载含镁矿物.

XPS表征结果见图3. 图3 (a, b)是XPS全扫图,可以看出未改性生物炭中除了含有C、N、O元素,还存在Si、Al、Ca、Na等矿物盐类元素;氯化镁改性使生物炭的XPS全谱中出现明显的Mg(1s)谱峰,也带入一定量的Cl元素. 将生物炭的C(1s)精细谱用Avantage软件进行分峰拟合,结果见图3 (c, d). 峰值为284.8 eV的峰代表C—C中的C元素;286.1—286.2 eV处代表C—O;287.5—287.6 eV处代表醛或酮的C=O;288.8 eV处代表羧基或酯基的O—C=O[34-35]. 此外,290.0—290.2 eV处是π−π*峰,是芳环中的离域电子跃迁形成的[34, 36]. 这说明生物炭具有富π电子的表面,能与MC-LR分子中含共轭结构的胍基发生π+−π电子供体-受体(π+−π EDA)相互作用而利于吸附[37]. 氯化镁改性使C—C中C元素占总C元素的比例下降,而C—O和O—C=O占比上升,这表明氯化镁改性使生物炭的含氧官能团增加. 图3 (e, f)是Mg(1s)的分峰拟合谱图,可以看出镁改性生物炭的Mg(1s)谱峰明显强于未改性生物炭,且Mg元素主要以Mg(OH)2和MgO的形式存在,1303.3—1303.4 eV的峰代表Mg(OH)2,1304.0—1304.2 eV的峰代表MgO[38-40].
 图 3 生物炭的XPS谱:XPS全谱(a, b);C(1s)谱及其分峰拟合(c, d);Mg(1s)谱及其分峰拟合(e, f)Figure 3. XPS spectra of biochar: XPS survey (a, b); C(1s) spectra and deconvoluted fitting curves (c, d); Mg(1s) spectra and deconvoluted fitting curves (e, f)
图 3 生物炭的XPS谱:XPS全谱(a, b);C(1s)谱及其分峰拟合(c, d);Mg(1s)谱及其分峰拟合(e, f)Figure 3. XPS spectra of biochar: XPS survey (a, b); C(1s) spectra and deconvoluted fitting curves (c, d); Mg(1s) spectra and deconvoluted fitting curves (e, f)对未改性生物炭KB、HB和镁改性生物炭KB-2.0Mg、HB-2.0Mg进行XRD表征,并通过比对ICDD PDF-2 2004标准谱图进行物相鉴定,结果见图4. 从图4可以看出,镁改性生物炭中含有Mg(OH)2(PDF卡号:44-1482)和MgO(PDF卡号:45-0946)的衍射峰,这说明镁改性生物炭中含有Mg(OH)2和MgO结晶,这也进一步佐证了XPS中的分析结果. 此外,生物炭中还含有CaCO3(PDF卡号:05-0586)和SiO2(PDF卡号:46-1045)等矿物成分.
2.1.4 零电荷点
用pH漂移法测定生物炭的零电荷点(pHpzc),其测定原理是将生物炭投加到不同初始pH的Na2SO4电解质溶液中,当溶液的pH<pHpzc时,生物炭会净吸附H+,使溶液pH升高;当溶液的pH>pHpzc时,生物炭会净吸附OH−,使溶液pH下降;而pH=pHpzc时,则溶液pH不变[41]. 因此,以溶液初始pH(Initial pH)为横坐标、混合24 h之后的pH(Final pH)为纵坐标作图,用曲线连接,曲线上Initial pH = Final pH的点即pHpzc[17-18],结果见图5. 可以看出,未改性生物炭和镁改性生物炭的pHpzc>7,这说明在中性水体中生物炭表面会净吸附H+而带正电[42],而MC-LR分子在中性条件下带负电[43],因此生物炭与MC-LR分子之间存在静电吸引作用. 镁改性生物炭的pHpzc高于未改性生物炭,且苦草和狐尾藻制得的生物炭在镁改性后pHpzc值基本相同. 研究表明Mg(OH)2和MgO有较高的pHpzc,分别在10.8—12和9.8—12之间[44],因此改性负载MgO和Mg(OH)2能提高生物炭的pHpzc.
 图 5 生物炭的零电荷点测定(a)苦草生物炭Vallisneria biochar;(b)狐尾藻生物炭Myriophyllum biocharFigure 5. pHpzc determination of biochar
图 5 生物炭的零电荷点测定(a)苦草生物炭Vallisneria biochar;(b)狐尾藻生物炭Myriophyllum biocharFigure 5. pHpzc determination of biochar2.2 生物炭对MC-LR的吸附去除率
比较未改性生物炭和不同镁浸渍浓度制备的镁改性生物炭对MC-LR的吸附去除性能,投炭量为2.0 mg/50 mL,MC-LR去除率见图6. 从图6可以看出,MgCl2浸渍浓度为2.0 mol·L−1制备得到的镁改性生物炭对MC-LR的吸附性能最佳;与未改性生物炭相比,镁改性生物炭对MC-LR吸附性能有明显的提高. 研究表明,MC-LR的分子尺寸为1.9 nm×1.5 nm×1.1 nm[45],由于体积排阻效应MC-LR分子无法进入微孔,中孔填充是生物炭吸附MC-LR的重要机制[34, 37]. 生物炭的表征结果已表明,镁改性使生物炭的比表面积增大,尤其中孔孔容增加明显,从而提升了生物炭的吸附性能;此外,生物炭表面的含氧官能团能与MC-LR中的氨基、羧基形成氢键[46],而镁改性能使生物炭的含氧官能团增加,有利于对MC-LR的吸附. 镁改性生物炭的pHpzc要高于未改性生物炭(图5),表明镁改性生物炭在溶液中能吸附更多H+而带有更多的正电荷[41],能与带负电的MC-LR分子产生更强的静电吸引力,有利于吸附.
 图 6 未改性生物炭和不同MgCl2浸渍浓度制备的镁改性生物炭对MC-LR的去除率Figure 6. The removal rate of MC-LR of unmodified biochar and Mg-modified biochar prepared with different MgCl2 soaking concentration(投炭量2.0 mg/50 mL,图中K和H分别表示苦草和狐尾藻生物炭,浸渍浓度标记为0的表示未改性生物炭)(carbon addition: 2.0 mg/50 mL. K and H represented Vallisneria and Myriophyllum biochar respectively, and the soaking concentration marked 0 represented the unmodified biochar)
图 6 未改性生物炭和不同MgCl2浸渍浓度制备的镁改性生物炭对MC-LR的去除率Figure 6. The removal rate of MC-LR of unmodified biochar and Mg-modified biochar prepared with different MgCl2 soaking concentration(投炭量2.0 mg/50 mL,图中K和H分别表示苦草和狐尾藻生物炭,浸渍浓度标记为0的表示未改性生物炭)(carbon addition: 2.0 mg/50 mL. K and H represented Vallisneria and Myriophyllum biochar respectively, and the soaking concentration marked 0 represented the unmodified biochar)2.3 吸附动力学
研究未改性生物炭KB、HB和镁改性生物炭KB-2.0 Mg、HB-2.0 Mg对MC-LR的吸附动力学. 用式1计算吸附量,使用OriginPro 2018软件将动力学数据用式(2—5)拟合[47-49]:
吸附量:qt=(C0−Ct)Vm (1) 准一级动力学:qt=qe(1−e−k1t) (2) 准二级动力学:qt=k2qe2t1+k2qet (3) Elovich:qt=1βln(αβt+1) (4) 颗粒内扩散:qt=kit1/2+Ci (5) 式中,t表示吸附时间(h);qt为t时刻的吸附量(μg·g−1);C0和Ct分别为初始MC-LR浓度和t时刻的MC-LR浓度(μg·L−1);V代表溶液体积(L);m代表生物炭的质量(g). qe、k1、k2、α、β、ki、Ci则是各模型拟合计算得到的参数,其中qe代表计算得到的平衡吸附量(μg·g−1).
准一级、准二级动力学和Elovich模型的拟合图形见图7 (a, b),拟合参数见表3. 通过比较拟合优度R2,KB和KB-2.0Mg的吸附过程用准一级动力学拟合效果较好,而HB和HB-2.0Mg的吸附过程则更符合Elovich和准二级动力学模型. 通常,准一级动力学用于描述物理作用主导的吸附,如液膜扩散机制等[50-51];准二级动力学用于描述化学吸附作用,如配位络合、电子的得失和共享、化学键的形成等机制[52];Elovich可反映发生在异质性表面的化学吸附[49, 53].
 图 7 镁改性及未改性生物炭吸附MC-LR的吸附动力学Figure 7. Adsorption kinetics of MC-LR on Mg-modified and unmodified biochar苦草(a)和狐尾藻(b)生物炭准一级、准二级动力学和Elovich模型拟合;苦草(c)和狐尾藻(d)生物炭的颗粒内扩散模型分阶段拟合fitting of pseudo-1st order, pseudo-2nd order, Elovich models of Vallisneria (a) and Myriophyllum (b) biochar; fitting of intra-particle diffusion model of Vallisneria (c) and Myriophyllum (d) biochar in stages表 3 吸附动力学拟合参数Table 3. Fitting parameters of adsorption kinetics
图 7 镁改性及未改性生物炭吸附MC-LR的吸附动力学Figure 7. Adsorption kinetics of MC-LR on Mg-modified and unmodified biochar苦草(a)和狐尾藻(b)生物炭准一级、准二级动力学和Elovich模型拟合;苦草(c)和狐尾藻(d)生物炭的颗粒内扩散模型分阶段拟合fitting of pseudo-1st order, pseudo-2nd order, Elovich models of Vallisneria (a) and Myriophyllum (b) biochar; fitting of intra-particle diffusion model of Vallisneria (c) and Myriophyllum (d) biochar in stages表 3 吸附动力学拟合参数Table 3. Fitting parameters of adsorption kinetics生物炭Biochar Pseudo-1st Pseudo-2nd Elovich k1/h−1 qe/(μg·g−1) R2 k2/(g·μg−1·h−1) qe/(μg·g−1) R2 α/(μg·g−1·h−1) β/(g·μg−1) R2 KB 2.835×10−1 52.75 0.9021 6.530×10−3 56.78 0.8332 51.22 1.018×10−1 0.7084 HB 2.801×10−1 91.78 0.8664 3.760×10−3 101.1 0.9513 111.9 5.777×10−2 0.9827 KB-2.0Mg 8.487×10−2 197.3 0.9779 3.962×10−4 234.1 0.9711 32.95 1.783×10−2 0.9567 HB-2.0Mg 8.281×10−2 203.0 0.9691 3.921×10−4 239.3 0.9776 36.32 1.795×10−2 0.9789 注:数据保留4位有效数字. The data retains 4 significant digits. | Show TableDownLoad:
CSV
吸附过程一般可分为外部液膜扩散阶段、颗粒内扩散阶段、吸附平衡阶段[54]. 从图7(c, d)可以看出,在前期吸附未达到平衡时的阶段,颗粒内扩散模型较好地拟合,由此可见颗粒内扩散也是吸附的重要阶段性机制.
2.4 吸附等温线
测定并计算在不同吸附平衡浓度下生物炭对MC-LR的吸附量(式6),利用Langmuir和Freundlich模型(式7—8)拟合绘制吸附等温线[55-56]:
平衡吸附量:qe=(C0−Ce)Vm (6) Langmuir:qe=qm⋅KL⋅Ce1+KL⋅Ce (7) Freundlich:qe=KF⋅Ce1/n (8) 式中,qe为平衡吸附量(μg·g−1),Ce为平衡浓度(μg·L−1),qm、KL、KF、n均为模型拟合得到的参数,其中qm代表拟合计算得到的最大吸附量(μg·g−1).
等温线拟合参数列于表4,拟合图形见图8. Langmuir模型假设吸附过程是发生在均质表面的单层吸附,且吸附质分子间无相互作用力;Freundlich模型是经验模型,对非均质表面以及多层吸附有较好的拟合效果,对物理吸附和化学吸附均适用[57]. 对于KB和HB,Langmuir模型和Freundlich模型的R2相差不大,说明生物炭吸附MC-LR机制是复杂的,既存在均一的单层吸附,也存在发生在非均质表面上的多层吸附. 35 ℃的qm要高于25 ℃,表明提高温度能提升生物炭对MC-LR的吸附容量.
表 4 吸附等温线拟合参数Table 4. Fitting parameters of adsorption isothermT/℃ Langmuir Freundlich KL/(L·μg−1) qm/(μg·g−1) R2 KF/(μg1−1/n·L 1/n·g−1) n R2 KB 25 5.046×10−2 189.7 0.6500 31.47 2.721 0.6452 35 1.803×10−2 311.2 0.7148 14.59 1.712 0.6899 HB 25 8.469×10−2 300.1 0.6706 67.97 3.089 0.6378 35 3.091×10−2 667.2 0.8925 47.40 1.816 0.8654 KB-2.0Mg 25 4.106×10−2 497.4 0.8846 52.22 2.115 0.9117 35 1.552×10−2 960.7 0.8001 36.23 1.603 0.8455 HB-2.0Mg 25 6.204×10−1 474.5 0.8242 211.1 4.301 0.8946 35 3.488×10−2 1909 0.9875 87.03 1.350 0.9796 注:数据保留4位有效数字.The data retains 4 significant digits. | Show TableDownLoad:
CSV
2.5 吸附影响因素
2.5.1 pH的影响
图9显示了不同pH条件下生物炭对MC-LR的去除率. 由图9可以看出,不同pH会影响去除率的大小,由于MC-LR在强酸碱条件下仍能保持稳定不易分解[58],说明实验结果去除率的差异取决于生物炭吸附的差异. pH升高对未改性生物炭的吸附性能有抑制作用,尤其在pH 10的条件下其对MC-LR的去除率明显下降;而镁改性生物炭受pH的影响较小,仅pH升高到10时其吸附性能略有下降. 研究表明,MC-LR分子中的羧基和氨基在不同pH下会发生电离或离子化,使分子带有不同的净电荷:pH<2.09主要以MCLR+形式存在,pH 2.09—2.19主要是MCLR0,pH 2.19—12.48主要是MCLR−,pH>12.48则主要是MCLR2−[43]. 可见在pH 4—10区间主要以MCLR−存在. 由于镁改性生物炭的pHpzc>10,其在pH 4—10带正电,与MC-LR分子存在静电吸引;未改性生物炭pHpzc 9—10,其在pH 4—8带正电与MC-LR分子存在静电吸引,而在pH 10生物炭几乎不带电或带轻微负电,存在静电排斥. 研究表明,pH的降低能使生物炭表面的净正电荷增加[59],从而增强静电吸引,这可能是较低pH下有着较高的吸附去除率的原因. 此外,pH降低还能使MC-LR分子发生卷曲,使分子体积减小,从而促进吸附[60].
 图 8 吸附MC-LR的吸附等温线Figure 8. Adsorption isotherm of MC-LR adsorption: fitting of Langmuir and Freundlich models of KB (a), HB (b), KB-2.0Mg (c) and HB-2.0Mg (d)KB (a)、HB (b)、KB-2.0Mg (c)和HB-2.0Mg (d)的Langmuir和Freundlich模型拟合
图 8 吸附MC-LR的吸附等温线Figure 8. Adsorption isotherm of MC-LR adsorption: fitting of Langmuir and Freundlich models of KB (a), HB (b), KB-2.0Mg (c) and HB-2.0Mg (d)KB (a)、HB (b)、KB-2.0Mg (c)和HB-2.0Mg (d)的Langmuir和Freundlich模型拟合2.5.2 DOM的影响
由于天然水中存在DOM,为探讨DOM对吸附的影响,选用没食子酸(GA)、富里酸(FA)、单宁酸(TA)做为模式DOM开展研究,结果见图10. 从图10可以看出,添加GA对生物炭吸附MC-LR的几乎没有影响,而添加FA和GA对吸附有明显的抑制作用. MC-LR的分子量为995.2 g·mol−1,分子尺寸为1.9 nm×1.5 nm×1.1 nm[45],如前文所述主要占据生物炭的中孔;而GA分子量(170.12 g·mol−1)和分子尺寸(0.90 nm×0.63 nm×0.28 nm)较小,主要占据微孔[61],与MC-LR的吸附几乎不会发生竞争作用,因此GA的添加对吸附的影响小;FA的分子量(308.24 g·mol−1)大于GA,竞争作用加强,导致生物炭吸附MC-LR减弱;TA分子量(1701.2 g·mol−1)和分子尺寸(1.93 nm×1.73 nm×1.32 nm)较大,依赖中孔填充吸附[61],与MC-LR分子竞争吸附明显,其更大的体积还会阻塞孔道,使MC-LR分子难以进入合适大小的孔隙[62-63],因此TA的添加能产生明显的抑制作用. DOM对未改性生物炭的抑制作用表现为TA>FA>GA;而对于镁改性生物炭,TA与FA的抑制作用相近,这可能是因为氯化镁改性使生物炭的中孔增加,使其不再那么“稀缺”,因而在一定程度缓解了TA和MC-LR对中孔的竞争.
 图 10 DOM对生物炭吸附去除MC-LR的影响Figure 10. The effect of DOM on the removal of MC-LR by biochar
图 10 DOM对生物炭吸附去除MC-LR的影响Figure 10. The effect of DOM on the removal of MC-LR by biochar3. 结论(Conclusions)
(1)用MgCl2溶液浸渍水生植物苦草和狐尾藻,热解制备镁改性生物炭. 氯化镁改性使生物炭的比表面积和孔容增加,尤其是中孔的增加,表面含氧官能团增加. 镁改性生物炭表面负载了MgO和Mg(OH)2,并具有更高的pHpzc值.
(2)镁改性生物炭比未改性生物炭对MC-LR有更强的吸附性能,以2.0 mol·L−1的MgCl2浸渍制备的生物炭对MC-LR有最佳的去除效果. 准一级、准二级动力学、Elovich和颗粒内扩散模型能在不同程度较好地描述吸附过程. 吸附等温线符合Langmuir和Freundlich模型,且较高的温度能提升吸附容量. 较高的pH和较大分子量的DOM会抑制生物炭对MC-LR的吸附,镁改性生物炭会使pH和DOM对吸附的影响减弱.
(3)生物炭对MC-LR的吸附机理是复杂的,可能同时存在物理吸附和化学吸附作用,既存在均一的单层吸附,也可能在非均质表面上发生多层吸附. 此外,颗粒内扩散是吸附的重要阶段性过程. 中孔填充是生物炭吸附MC-LR的重要机制,生物炭和MC-LR分子间可能存在氢键、静电吸引和π+−π EDA相互作用力. 镁改性能增强中孔填充作用,并加强氢键和静电吸引力从而增强吸附.
-
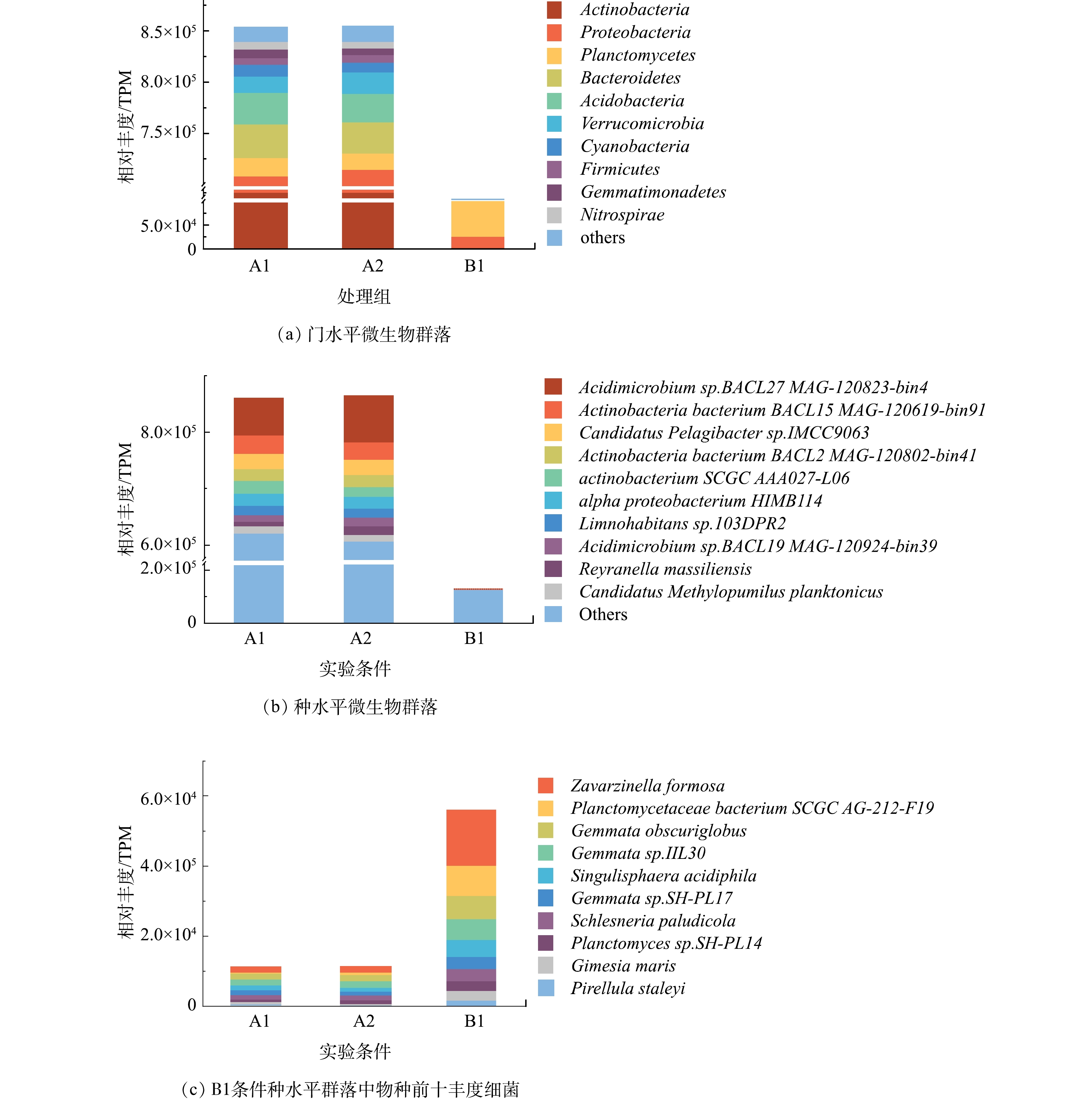
图 1 群落门水平与种水平组成
Figure 1. The relative abundance of total bacterial communities at phylum level and species level.
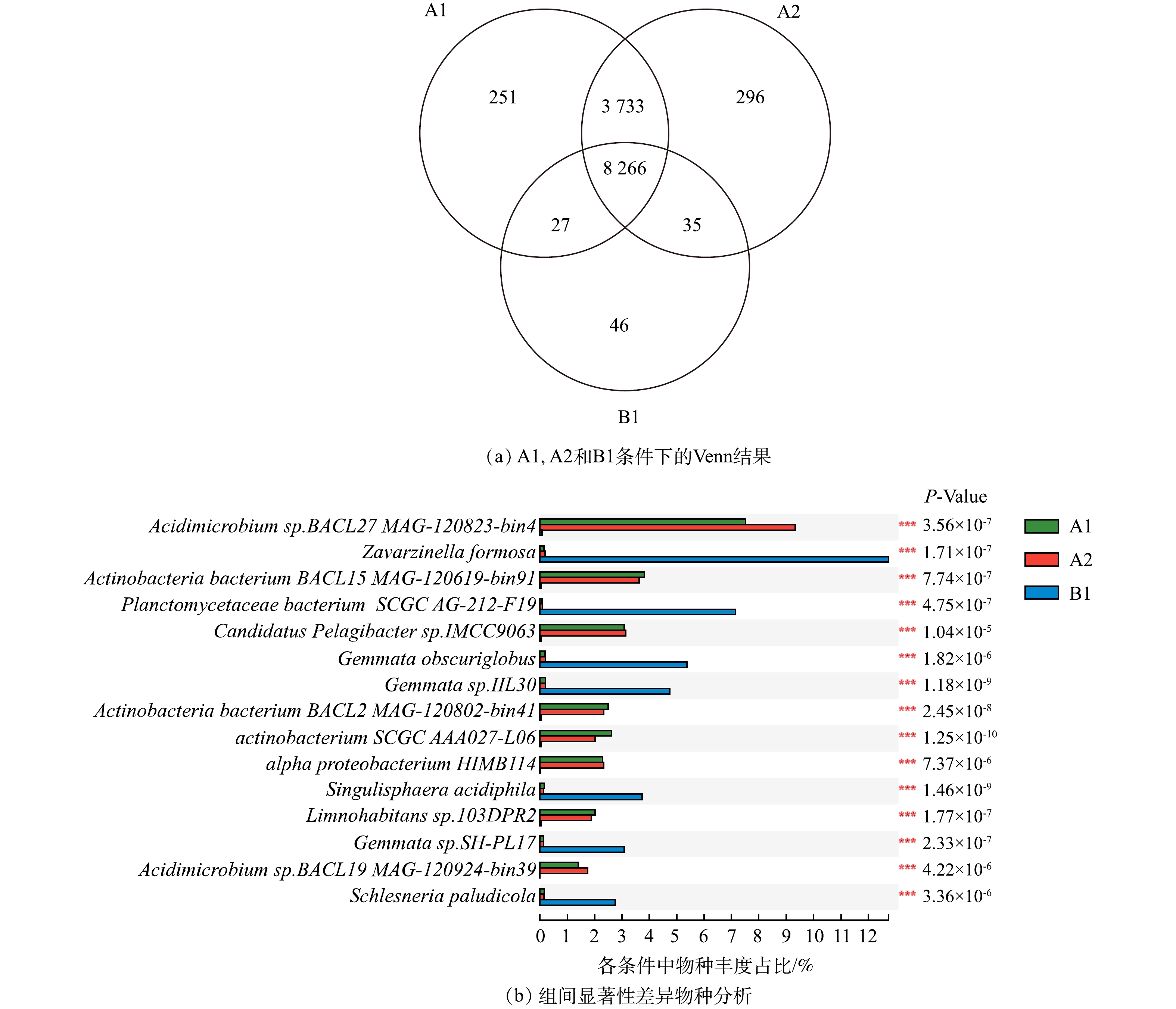
图 2 Venn结果和组间显著性差异物种分析
Figure 2. The Venn results and Metastats analysis at species level
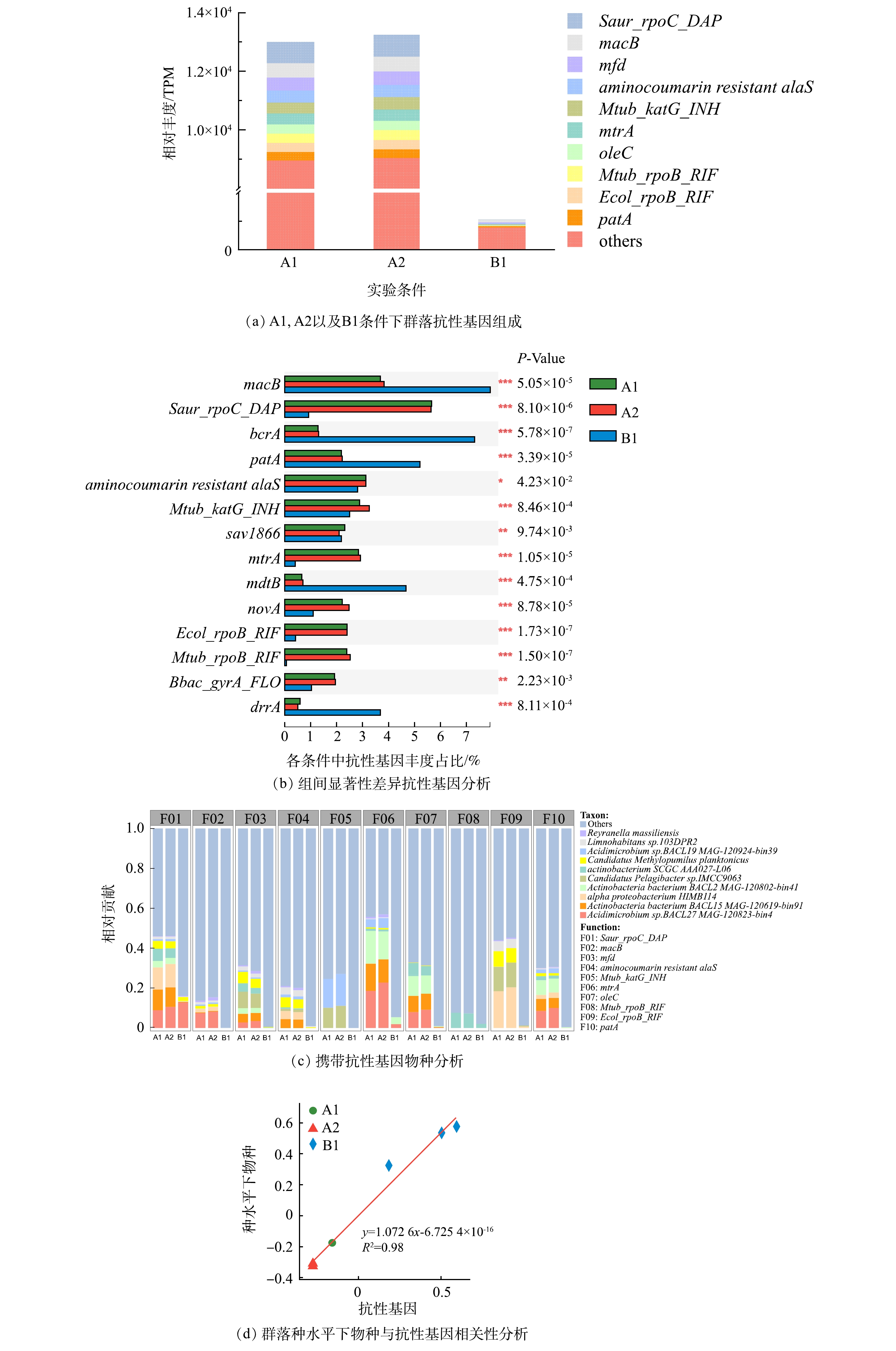
图 3 群落抗性基因组成,差异抗性基因以及携带耐药性基因物种
Figure 3. The relative abundance of antibiotic resistance genes(ARGs) of communities,the Metastats analysis on ARGs and the contribution of each species to the different ARGs.
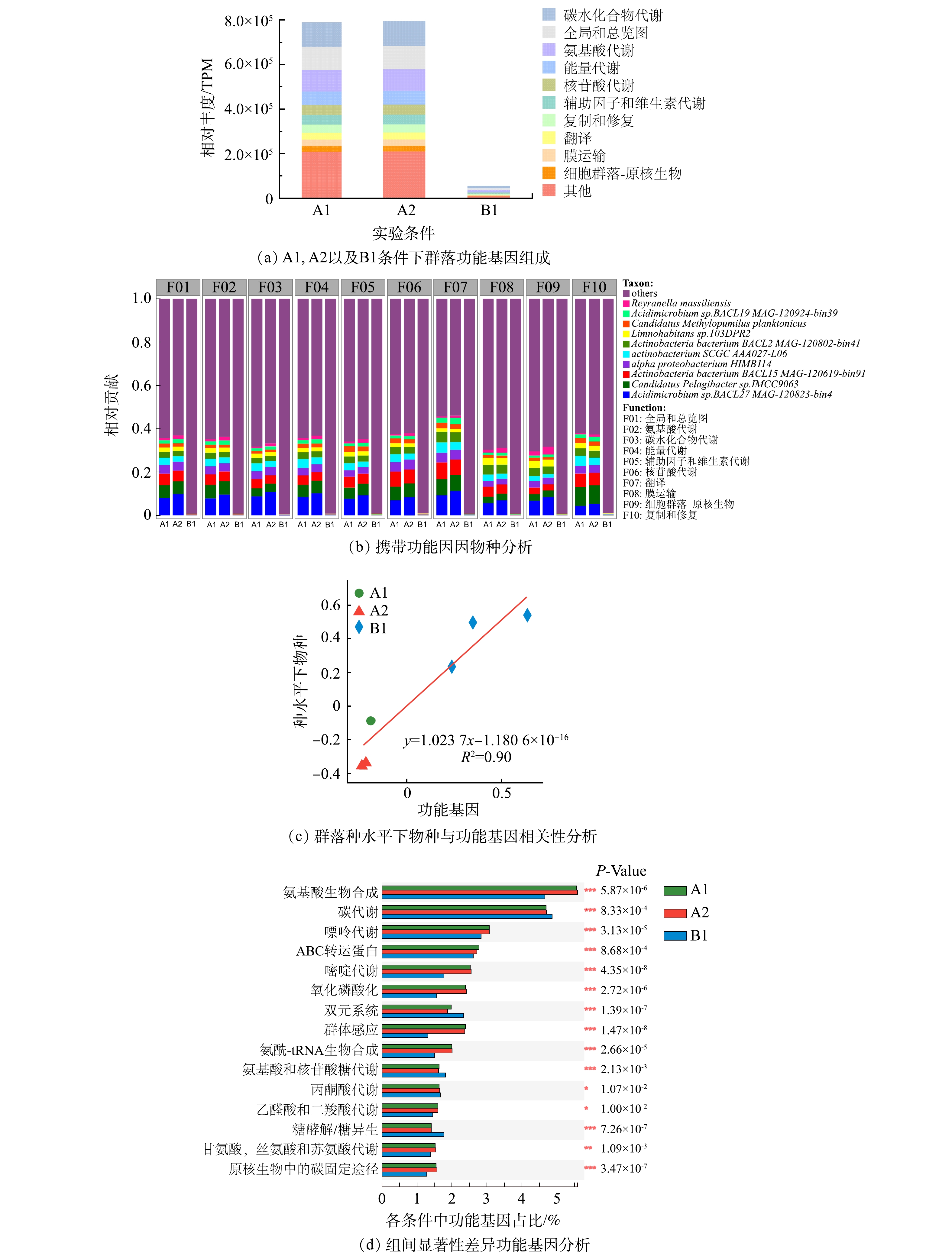
图 4 群落功能基因组成,差异功能基因以及携带功能基因物种
Figure 4. The relative abundance of antibiotic functional genes of communities, the Metastats analysis on KEGG functional genes and the contribution of each species to the different functional genes
-
[1] HOZALSKI R M, LAPARA T M, ZHAO X, et al. Flushing of stagnant premise water systems after the COVID-19 shutdown can reduce infection risk by Legionella and Mycobacterium spp[J]. Environmental Science & Technology, 2020, 54(24): 15914-15924. [2] 张明露, 周贺, 关磊等. 饮用水配水系统中微生物研究方法的进展[J]. 环境与健康杂志, 2015, 32(5): 458-462. doi: 10.16241/j.cnki.1001-5914.2015.05.024 [3] TANG W, LI Q, CHEN L, et al. Biofilm community structures and opportunistic pathogen gene markers in drinking water mains and the role of pipe materials[J]. ACS ES& T. Water, 2021, 1(3): 630-640. [4] 祝泽兵, 裴云燕, 单莉莉等. 供水管网生物膜中微生物种间相互作用及其影响因素综述[J/OL]. 环境工程: 1-15[2023-03-17]. http://kns.cnki.net/kcms/detail/11.2097.X.20221123.0807.002.html. [5] SIDHU J P S, GUPTA V V S R, STANGE C, et al. Prevalence of antibiotic resistance and virulence genes in the biofilms from an aquifer recharged with stormwater[J]. Water Research, 2020, 185: 0043-1354. [6] LIANG J, MAO G, YIN X, et al. Identification and quantification of bacterial genomes carrying antibiotic resistance genes and virulence factor genes for aquatic microbiological risk assessment[J]. Water Research, 2020, 168: 115160. doi: 10.1016/j.watres.2019.115160 [7] CIOFU O, MOSER C, JENSEN P Ø, et al. Tolerance and resistance of microbial biofilms[J]. Nature Reviews Microbiology, 2022, 20: 621-635. doi: 10.1038/s41579-022-00682-4 [8] 钟丹, 周子仪, 马文成等. 供水管网中抗生素抗性基因环境风险浅析[J]. 给水排水, 2020, 56(S2): 59-63. doi: 10.13789/j.cnki.wwe1964.2020.S2.010 [9] LI J, ZHAO L, FENG M, et al. Abiotic transformation and ecotoxicity change of sulfonamide antibiotics in environmental and water treatment processes: A critical review[J]. Water Research, 2021, 202: 117463. doi: 10.1016/j.watres.2021.117463 [10] YIN R, GUO W, REN N, et al. New insight into the substituents affecting the peroxydisulfate nonradical oxidation of sulfonamides in water[J]. Water Research, 2020, 171: 115374. doi: 10.1016/j.watres.2019.115374 [11] ZHANG B, QIN S, GUAN X, et al. Distribution of antibiotic resistance genes in Karst River and its ecological risk[J]. Water Research, 2021, 203: 117507. doi: 10.1016/j.watres.2021.117507 [12] ZHANG Z, WANG Y, CHEN B, et al. Xenobiotic pollution affects transcription of antibiotic resistance and virulence factors in aquatic microcosms[J]. Environmental Pollution, 2022, 306: 119396. doi: 10.1016/j.envpol.2022.119396 [13] LIN Q, LI L, FANG X, et al. Substrate complexity affects the prevalence and interconnections of antibiotic, metal and biocide resistance genes, integron-integrase genes, human pathogens and virulence factors in anaerobic digestion[J]. Journal of Hazardous Materials, 2022, 438: 129441. doi: 10.1016/j.jhazmat.2022.129441 [14] TANG T, CHEN Y, DU Y, et al. Effects of functional modules and bacterial clusters response on transmission performance of antibiotic resistance genes under antibiotic stress during anaerobic digestion of livestock wastewater[J]. Journal of Hazardous Materials, 2023, 441: 129870. doi: 10.1016/j.jhazmat.2022.129870 [15] WANG H, EDWARDS M. A, FALKINHAM J. O 3RD, et al. Probiotic approach to pathogen control in premise plumbing systems? A review[J]. Environmental Science & Technology, 2013, 47(18): 10117-10128. [16] LU Z, SUN W, LI C, et al. Bioremoval of non-steroidal anti-inflammatory drugs by Pseudoxanthomonas sp. DIN-3 isolated from biological activated carbon pro`cess[J]. Water Research, 2019, 161: 459-472. doi: 10.1016/j.watres.2019.05.065 [17] 陈蕾. 污水中抗生素抗性菌及抗性基因的去除技术. 污水中抗生素抗性菌及抗性基因的去除技术[J]. 生态环境学报, 2018, 27(11): 2163-2169. [18] LI B, QIU Y, ZHANG J, et al. Real-time study of rapid spread of antibiotic resistance plasmid in biofilm using microfluidics[J]. Environmental Science & Technology, 2018, 52(19): 11132-11141. [19] FARHAT N, KIM L, MINETA K, et al. Seawater desalination based drinking water: Microbial characterization during distribution with and without residual chlorine[J]. Water Research, 2022, 210: 117975. doi: 10.1016/j.watres.2021.117975 [20] 付树森, 王艺, 王肖霖, 等. 氯和紫外消毒过程中胞外抗性基因的产生特征[J]. 中国环境科学, 2021, 41(10): 4756-4762. doi: 10.3969/j.issn.1000-6923.2021.10.032 [21] LU Z, JING Z, HUANG J, et al. Can we shape microbial communities to enhance biological activated carbon filter performance?[J]. Water Research, 2022, 212: 118104. doi: 10.1016/j.watres.2022.118104 [22] 李晓明, 王飞, 李建勇等. 饮用水中抗生素污染现状及降解技术研究进展[J]. 食品安全导刊, 2016, 144(21): 94-95. doi: 10.16043/j.cnki.cfs.2016.21.069 [23] 钟文辉, 曹一鸣, 肖露等. 自来水厂次氯酸钠消毒技术应用总结[J]. 清洗世界, 2022, 38(6): 93-96. [24] 漆文光. 自来水厂采用次氯酸钠替代液氯消毒效果研究[J]. 供水技术, 2019, 13(3): 43-47. [25] GOMEZ-SMITH C K, LAPARA T M, HOZALSKI R M. Sulfate reducing bacteria and Mycobacteria dominate the biofilm communities in a chloraminated drinking water distribution system[J]. Environmental Science & Technology, 2015, 49(14): 8432-8440. [26] POTGIETER S, DAI Z, HAVENGA M, et al. Reproducible microbial community dynamics of two drinking water systems treating similar source waters[J]. ACS. ES& T. Water, 2021, 1(7): 1617-1627. [27] THOM C, SMITH C J, MOORE G, et al. Microbiomes in drinking water treatment and distribution: A meta-analysis from source to tap[J]. Water Research, 2022, 212: 118106. doi: 10.1016/j.watres.2022.118106 [28] BORSETTO C, RAGUIDEAU S, TRAVIS E, et al. Impact of sulfamethoxazole on a riverine microbiome[J]. Water Research, 2021, 201: 117382. doi: 10.1016/j.watres.2021.117382 [29] CHEN J, YANG Y, LIU Y, et al. Bacterial community shift in response to a deep municipal tail wastewater treatment system[J]. Bioresource Technology, 2019, 281: 195-201. doi: 10.1016/j.biortech.2019.02.099 [30] 韩雪, 孙坚伟, 张力等. 紫外氯胺组合消毒供水系统中病毒微生物的分布特征[J]. 环境科学, 2021, 42(2): 860-866. doi: 10.13227/j.hjkx.202007039 [31] WANG Y H, WU Y H, LUO L W, et al. Metagenomics analysis of the key functional genes related to biofouling aggravation of reverse osmosis membranes after chlorine disinfection[J]. Journal of Hazardous Material, 2021, 410: 124602. doi: 10.1016/j.jhazmat.2020.124602 [32] EOH H, LIU R, LIM J, et al. Microbial characterization during distribution with and without residual chlorine[J]. Frontiers in Cellular and Infection Microbiology, 2022, 12: 958240. doi: 10.3389/fcimb.2022.958240 [33] WANG M, LIAN Y, WANG Y, et al. The role and mechanism of quorum sensing on environmental antimicrobial resistance[J]. Environmental Pollution, 2023, 322: 121238. doi: 10.1016/j.envpol.2023.121238 [34] ZOU S, ZHANG Q, ZHANG X, et al. Environmental factors and pollution stresses select bacterial populations in association with protists[J]. Frontiers in Marine Science, 2020, 7: 659. doi: 10.3389/fmars.2020.00659 [35] MORRISSEY K, IVESA L, DELVA S, et al. Impacts of environmental stress on resistance and resilience of algal-associated bacterial communities[J]. Ecology and Evolution, 2021, 11: 15004-15019. doi: 10.1002/ece3.8184 [36] SMITS S H J, SCHMITT L, BEIS K. Self-immunity to antibacterial peptides by ABC transporters[J]. FEBS Letters, 2020, 594: 3920-3942. doi: 10.1002/1873-3468.13953 [37] AHMED M S, LAUERSEN K J, IKRAM S, et al. Efflux transporters' engineering and their application in microbial production of heterologous metabolites[J]. ACS Synthetic Biology, 2021, 10,4: 646-669. [38] GOMEZ-ALVAREZ, SIPONEN S, KAUPPINEN A, et al. A comparative analysis employing a gene- and genome-centric metagenomic approach reveals changes in composition, function, and activity in waterworks with different treatment processes and source water in Finland[J]. Water Research, 2023, 229: 119495. doi: 10.1016/j.watres.2022.119495 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 3247
- HTML全文浏览数: 3247
- PDF下载数: 119
- 施引文献: 0
