















-
我国分散式农村生活污水[1]、河流湖泊水体[2]等氮污染形势严峻,且存在低碳氮比问题,而国家对环境中的TN要求日益严格。目前污水中氮主要通过生物异养反硝化去除[3],其原理是反硝化菌在缺氧或厌氧环境下以有机物作为电子供体,硝氮 (NO3−-N) 为最终电子受体,通过电子传递链将NO3−依次还原为NO2−、NO、N2O和N2 [4]。碳源与电子供体会影响反硝化效果,碳源缺乏往往导致NO3−去除率低[5]。低碳氮比污水存在碳源不足的问题,致使总氮 (TN) 超标,为提高TN脱除效果,通常需投加额外碳源[6]。固相碳源是一种可替代传统液相碳源的新型碳源[7],包括人工合成聚合物[8]、农业废弃物[5]等,其已被应用于地表水、人工湿地[9]、循环水产养殖[10]等氮的去除中,并取得了一定的效果[11]。植物碳源 (如稻壳、秸秆、木屑、芦苇等[12]) 因其廉价易得逐渐成为研究热点,其主要成分为木质纤维素 (由纤维素、半纤维和木质素组成) ,木质纤维素中的纤维素和半纤维可被微生物降解为小分子有机物,用于反硝化脱氮[3, 5]。一般植物碳源存在碳源释放不稳定、反硝化率低、有机物过量释放等问题[11]。如何在碳源释放稳定的前提下,进一步提高碳源水解能力,是植物碳源应用的关键[11-12]。
我国竹资源丰富,竹林面积约占森林面积的3%[13]。同时竹产业成熟,竹产品获取方便灵活,是一种潜在的廉价碳源。纤维絮状竹刨花是竹加工后的产物,主要成分为木质纤维素[14]。竹刨花不同于粉碎的竹屑,其具有一定强度与韧性[15],纤维交互堆叠并具有丰富孔隙,不易堵塞,表面粗糙,利于微生物附着,可作为污水处理填料和反硝化碳源。与一般禾本科农业植物不同,竹为木质生物质,纤维素和木质素含量更高,纤维结构更加复杂紧密[16],导致微生物的分解相对困难。木质生物质的缺点是反硝化率不高[11, 17],优点是利于长期应用[18]。
零价铁具有化学还原性强、对微生物的生长有益、氧化产物无毒的特点[19],被广泛应用于地下水、废水污染物的去除[20]等方面。生物反应器中的零价铁可刺激微生物进化并增加微生物多样性,释放的铁离子可加强微生物聚集并增加生物量,提高生物降解能力[21]。已有研究[22-23] 将零价铁与厌氧生物处理方法耦合以去除水中硝氮,结果表明,耦合体系具有污染物质去除率高、性能稳定、成本低等特点。纳米铁、铁粉等粒径小的铁虽比表面积大、活性高,但腐蚀过快[24],不利于长期利用。此外,较小的粒径尺寸增加了实际应用难度。已有研究[21]显示,比表面积较低的铁形态在水中的腐蚀相对较慢,在强化生物脱氮的过程中,可减少NH4+的形成,是一种较受青睐的铁形态。铁屑是金属加工后的废料,不仅成本低,而且比表面积也低于纳米铁或铁粉等,利于长期应用,且其具有较好的机械与水力特性,更加符合实际应用需求[25]。目前,将铁屑与竹基固相反硝化系统耦合以强化脱氮的研究鲜见报道。
本研究以纤维絮状竹刨花为固体碳源和生物膜载体,引入铁屑,构成耦合体系,从而高效去除低C/N比废水中TN;通过监测装置进、出水水质,考察体系对TN的去除以及有机物、总铁的释放情况等;采用扫描电镜、高通量测序等表征方法分析微生物群落结构和多样性,旨在为高效、稳定、经济地去除低C/N比废水中TN提供技术参考,同时满足 “十四五”规划中“全面提高资源利用效率”“持续改善环境质量”等的要求。
-
1) 实验材料。竹刨花为毛竹制作竹篾过程中的副产物,刨花丝宽0~4 mm、厚0~1 mm、长1~30 cm,整体呈为团聚状。将竹刨花过60目筛,去除粉屑,40 ℃ 烘干待用,测得其湿体积密度为182 g·L−1。铁屑为车床铁加工后的废料,挑选出宽1.5~3 mm,长2~5 cm的呈螺旋状的铁屑,40 ℃干燥待用,测得其湿体积密度为0.17 mL·g−1。将粒径为1.5~3 cm的鹅卵石与火山岩清洗干燥,用于反硝化装置的承托层与固定层。实验中,投加武汉水之国公司 (www.szghb.com) 生产的复合脱氮菌剂,以确保反硝化菌的挂膜。
2) 实验废水。实验废水采用自来水配置,以KNO3为N源,KH2PO4为P源,加入1 mg·L−1微量元素溶液,补充微生物生长所需微量元素。微量元素溶液中微量元素组成包括120 mg·L−1 ZnSO4·7H2O、20 mg·L−1 MgSO4·7H2O、30 mg·L−1 KI、30 mg·L−1 CoCl2·6 H2O、30 mg·L−1 CaCl2、20 mg·L−1 MnCl2。实验废水中TN为41.62~59.95 mg·L−1,COD为(9.92±3.36) mg·L−1,TP为 (1.14±0.06) mg·L−1,NO2−-N、NH3-N和总铁基本检测不到,pH为 7.29±0.21。
-
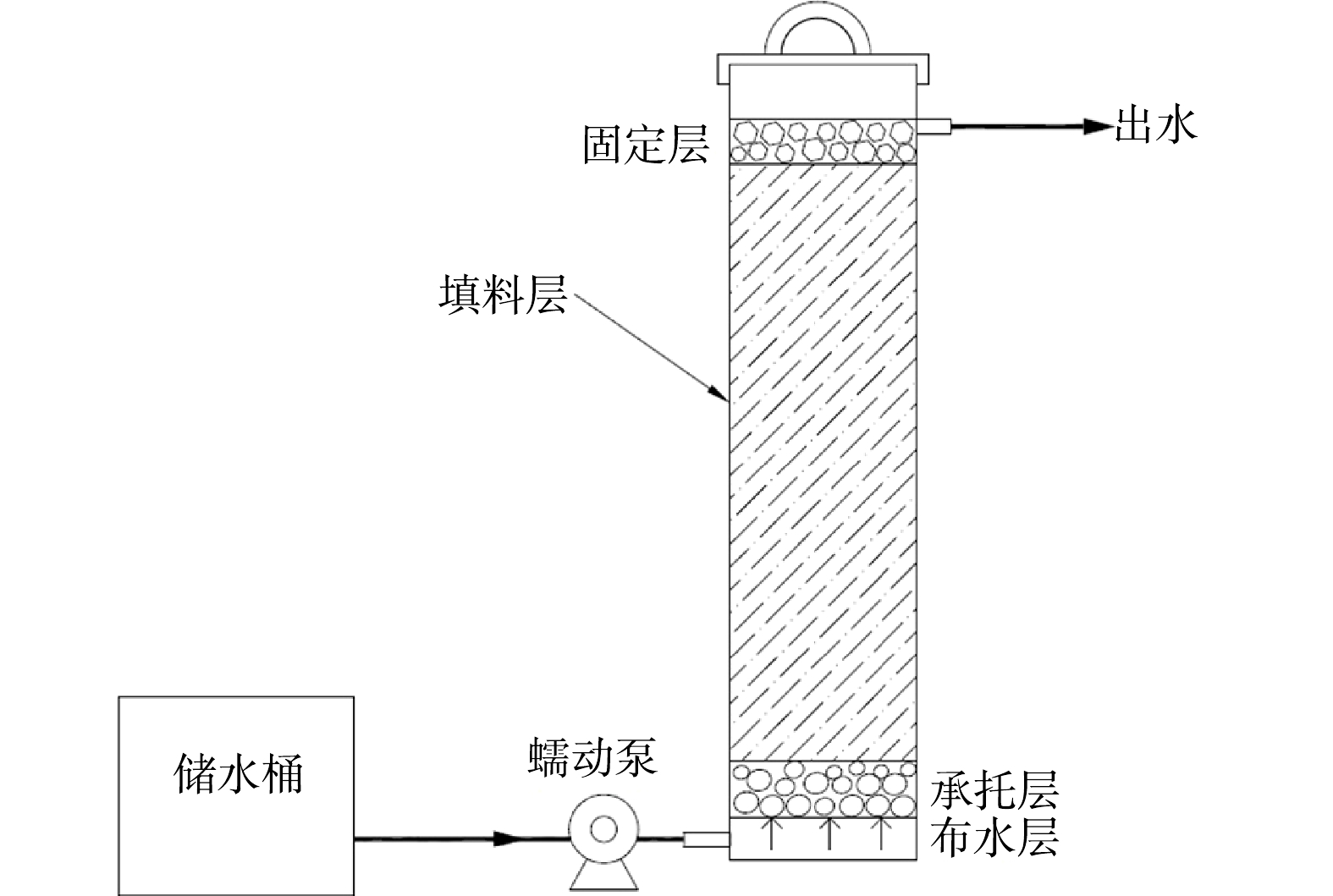
实验采用连续流动态反硝化装置 (图1) ,反应柱为透明有机玻璃制成,内径为11 cm,高为50 cm,从下至上分别为鹅卵石承托层 (约5 cm) 、填料层 (约32 cm) 和火山岩固定层 (约3 cm) 。装置共2组:1号为耦合体系实验组,其填料层为160 g竹刨花、80 g铁屑和1 g菌剂,竹刨花逐层铺设,铁屑分2次铺设于填料层底部,装填时,将1 g菌剂分层铺洒于填料层中;2号为单纯固相反硝化对照组,填料层只有160 g竹刨花,和1 g菌剂,铺设同上,菌剂为武汉水之国复合脱氮菌剂 (主要成分为短小芽孢杆杆菌、腊样芽孢杆菌、苏云金芽孢杆菌属微生物及营养剂等,有效活菌数≥200×108个·g−1) 。实验装置总有效体积约为2.16 L,采用底部连续进水方式,室温运行 105 d (2021年5—9月) 。先从装置底部进水,使填料完全浸没,之后停止进水并浸泡1周,再开始连续进水并计时。进水流速为2 mL·min−1,水力停留时间约为18 h。定期测试进、出水的TN、COD、总铁等指标。0~73 d,进水TN约为40 mg·L−1;74~105 d,进水TN升至60 mg·L−1左右。根据进水TN质量浓度与TN去除效果,运行的0~12 d设定为启动阶段,13~73 d为阶段I,74~105 d为阶段II。
-
1) 水质分析。采用德国耶拿Multi N/C ®2100仪器测定TN,采用重铬酸盐法 (HJ 828-2017) 测定COD,采用邻菲罗啉分光光度法 (HJ/T 345-2007) 测定总铁,采用N-(1-萘基)-乙二胺分光光度法 (GB/T 7493-1987) 测定NO2−-N,采用纳氏试剂分光光度法 (HJ 535-2009) 测定NH3-N,采用钼酸铵分光光度法 (GB 11893-1989) 测定TP,采用电极 (PHB-4) 测定pH,所用药品均为分析纯。
2) 微生物及分子生物学分析。在装置运行的第98天,取装置内部生物膜生长良好的竹刨花进行SEM观察并进行16S rRNA测试。选取2~5 mm竹刨花填料小块,使用2.5%戊二醛 (电镜专用) 使其固定,使用磷酸缓冲液进行清洗,使用乙醇进行梯度脱水,再进行临界点干燥和喷金,然后置于SEM电镜下观察。实验前的竹刨花由于没有生物膜,无需进行生物固定操作,直接干燥喷金观察。取填料层顶部5~7 cm厚湿竹刨花,质量约为100 g,放在装有纯水的烧杯中,用玻璃棒振搅,使其表面生物膜脱落,之后再将竹刨花回填于装置内,剩余生物膜溶液离心浓缩至约5 mL,低温保存,用于高通量测序。高通量测序采用MiseqPE 250测序模式,选择16S rRNA V3~V4区引物338 F和806 R (引物序列ACTCCTACGGGAGGCAGCA和GGACTACHVGGGTWTCTAAT) 对样品进行测试并分析测试结果,得到样品的菌群结构。以上所有表征均由杭州研趣信息技术有限公司完成。
-
有机物是反硝化的主要电子供体与能量来源,但有机物过多会引发二次污染。图2为实验装置进、出水COD变化情况。由图2可知,启动阶段 (0~12 d) ,前5 d,2组出水COD较高 (>100 mg·L−1) ,之后迅速降低,9 d后,2组出水COD低于50 mg·L−1。13~105 d (阶段I~阶段II),2 组出水COD基本稳定,预示挂膜成熟,1号和2号出水COD分别为 (45.97±6.58) mg·L−1和 (32.37±9.05) mg·L−1。启动阶段,竹刨花表层可溶性有机物溶出较多[5],导致前5 d出水COD较高,这确保了生物膜的快速生长。运行约12 d后,竹刨花表层可溶性有机物基本消耗完全,微生物主要通过分解、利用竹纤维素获取能源与电子供体等,但木质纤维素结构复杂稳定,分解较慢。另外,随着菌群的逐渐稳定,微生物对纤维素类物质的分解与利用趋于平衡,随水流出的有机物不多,因此,2组出水COD在阶段I~阶段II基本维持在较低水平。
在阶段I~阶段II,1号出水COD整体较2号高42.04%。这是因为铁促进了微生物的繁殖与酶的分泌,提高了微生物对竹纤维类物质的分解[19, 26-27]。此外,随着TN负荷的增加,在阶段II,1号和2号出水COD较阶段I分别增加9.17%、−16.44%,说明耦合体系有机物供应机制适应性更好。最后,13~105 d,2组出水COD均值均低于一级A限值 (50 mg·L−1,GB 18918-2002) ,未出现碳源过度释放情况。
-
图3为实验装置TN去除及出水COD/N曲线。在启动阶段,2组出水TN先降低再上升,运行12 d后 (阶段I和阶段II) ,2组出水TN基本趋于稳定。由于N是竹基本组分之一[28],竹刨花在初始大量释放有机物,这些有机物中的N经微生物分解后会增加水中TN,因此,第1天,测量2组出水TN均较高 (48.36~52.40 mg·L−1) 。2~7 d,2组出水COD/N均大于4,而满足或完成反硝化所需的COD/TN比为4~15[29],说明此时2组有机物充足,反硝化率高,TN去除率也达到启动阶段的最大值,分别为89.34%和75.98%。这表明2组均形成了较好的脱氮效能,且初期差异不大。8~12 d,2组出水COD/N逐渐降至低点,第12 天,2组出水TN达到最大值,为18.34~31.96 mg·L−1。另外,由图2可知,运行12 d后,2组出水有机物基本稳定,体系内部相对稳定的碳源环境使得反硝化反应趋于平衡,因此,13~105 d ,2组出水TN也相对稳定。
在阶段I,运行61 d,进水TN为 (41.62±1.29) mg·L−1,1号和2号出水TN均值分别为3.03 mg·L−1和21.05 mg·L−1,TN去除率分别为(92.79±9.31)%和(49.52±12.68)%,1号TN去除率比2号高87.36%;且运行50~73 d,1号TN去除率在95%以上,出水TN均值(1.30 mg·L−1)低于地表水IV类标准限制1.50 mg·L−1 (GB 3838-2002) 。在阶段II,运行32 d,进水TN为 (59.95±1.77) mg·L−1,1号和2号出水TN均值分别为16.01 mg·L−1和40.91 mg·L−1,TN去除率分别为70%和30%左右,1号TN去除率比2号高131.09%。总体而言,13~105 d,耦合体系TN去除率较单纯固相反硝化体系高近1倍。
赵文莉等[30]以碱处理玉米芯、零价铁和活性炭构成复合填料处理低碳氮比污水厂尾水,进水NO3−为20~30 mg·L−1,初期有机物多,TN去除率达100%,后有机物降低,运行60 d 时,TN去除率降为60%左右,碳源不足是限制脱氮的主要因素。在本研究的阶段I~阶段II,1号出水COD/N均高于2号,说明1号碳源更丰富,因此1号TN去除率更高。同时,随着TN负荷的增加,2组TN去除率均降低,且出水COD/N均小于4。这是因为竹中有机物释放缓慢,无法完全满足高负荷下的碳源需求。另外,从阶段I至阶段II,1号和2号TN削减量 (式 (1) ) 分别增加了13.89%和−7.44%,这与上文COD变化相似。以上情况说明碳源是影响2组脱氮效能的重要原因。
式中:∆Ci 为TN削减量;C0为进水TN质量浓度;Cei为出水TN质量浓度;i为1号或2号反应器。
纤维素分解后会释放腐殖质和类蛋白、小分子有机酸和糖等[31],而反硝化菌只能利用小分子有机酸脱氮[11]。铁可提高复杂有机物的降解率[19],改善碳源分解,从而提高耦合体系TN去除率。但研究[32]显示,采用植物碳源脱氮时,出水中有机质主要为腐殖质和类蛋白等。这类大分子物质反硝化菌无法直接利用[33],需进一步分解。因此,2组出水始终残留有COD而暂时无法用于脱氮,此时,可后接有机物处理模块,以进一步减少有机物污染。
-
图4 (a) 为实验装置NO2−-N积累情况。反硝化由硝酸盐还原酶 (Nar) 和亚硝酸盐还原酶 (Nir) 等完成。碳源不足,部分NO2−无法还原,造成NO2−积累[4]。碳源充足,Nar和Nir会竞争底物,且NaR竞争力更强[34-35],NO3−会优先还原为NO2−[36],造成NO2−积累[37]。在启动阶段,1~2 d,2组NO2−-N积累较高,由于NiR的合成与诱导时间晚于Nar[36-37],因此,运行初始出现NO2−-N积累,同时也证明2组反硝化菌生长成功。由此可知,在阶段I,1号出水COD/N均值大于4,碳源充足,NO2−-N积累平均质量浓度低于1 mg·L−1;2号出水COD/N均值小于4,碳源不足,24~50 d,NO2−-N积累较多,最大积累量为12.79 mg·L−1,之后逐渐降低并稳定至1 mg·L−1以下。在阶段II,1号出水COD/N降至4以下,碳源不足,78~98 d出现NO2−-N大量积累,最大积累量为11.62 mg·L−1,之后逐渐减少并稳定至2.5 mg·L−1左右;2号出水COD/N依然较低,NO2−-N平均积累质量浓度为1 mg·L−1。碳源不足时,2组均出现NO2−-N大量积累,但大量积累是暂时的,这与钟胜强等[38]和张恒亮等[39]实验情况类似。此外,51~73 d和74~105 d,2号TN削减量均值为23.71和19.04 mg·L−1,变化不显著,因此,51~105 d,2号NO2−-N积累变化不大。总之,耦合体系TN去除过程虽有一定NO2−-N积累,但大量积累时间不长,稳定积累量不高,通过后接曝气设施可减少NO2−-N危害。
植物碳源中的N经生物作用后会释放NH4+[40]。当有机电子供体远高于受体NO3−时,NO3−易异化还原成NH4+ (DNRA)[5, 41]。同时,Fe0生物系统中NO3−可化学还原为NH4+[21, 23]。图4 (b) 为实验装置NH3-N积累情况。可以看出,0~5 d,2组出水NH3-N较高,之后随着有机物释放的减少与稳定,2组出水NH3-N也随之减少和稳定;12 d后,2组出水NH3-N平均质量浓度均低于1 mg·L−1。由此可知,0~5 d,2组出水COD远高于TN,因此,前几天NH3-N较高主要是因为碳源释放与DNRA反应所致。另外,在阶段II (73 d后) ,1号出水NH3-N增加,此时期1号出水COD虽有少量增加,但出水COD/N较低,排除DNRA的影响。已有研究[42]显示,Fe2+易将生物还原型NO2−化学还原为NH4+ (式 (2) ) ,并形成Fe2+-Fe3+水合氧化物,其中的Fe2+会继续与NO3−或NO2−化学反应生成NH4+。在阶段II,NO3−增加,加剧了铁的腐蚀[21, 43],释放更多Fe2+ (式 (3)~式 (5) ) 以及 NH4+ (式 (5) ) 。此时1号碳源缺乏,积累的生物还原型NO2−和残余NO3−增多,导致NH4+进一步增加。由于铁腐蚀有限,故NH4+增加不多,加上材料自身释放的增加,最终导致73 d后1号NH3-N升高。整体而言,耦合体系出水NH3-N虽然略高于单纯固相反硝化体系,但并未出现NH3-N的明显积累。
-
图5为实验装置进出水总铁浓度变化情况。由于进水与2号出水总铁基本低于检测线,故主要讨论1号出水总铁情况。在启动阶段,1号出水总铁先升高后快速降低,运行12 d后,总铁释放稳定,出水总铁为 (0.26±0.09) mg·L−1,整体低于地表水环境质量标准限值0.3 mg·L−1 (GB 3838-2002)。耦合体系总铁的释放主要源于铁屑腐蚀。在启动阶段,铁屑在水中发生氧化、生物腐蚀,生成大量的Fe2+与Fe(OH)2[19, 44],导致前几天出水总铁较高。但铁屑表面的生物膜会减少物质交换而阻碍腐蚀[45],其表面生成的铁氧化物或氢氧化物会使铁钝化而失去活性,一些微生物会利用Fe2+还原NO3−,产生Fe3+,促进铁氧化物沉淀,进一步抑制腐蚀[42]。因此,随着装置的运行,总铁质量浓度迅速减少。然而铁的释放并未停止,有研究[43,46]指出,当水中硝酸盐质量浓度大于10 mg·L−1时,不稳定的腐蚀产物 (β-FeOOH) 容易脱落促进腐蚀。总之,基于各类生物化学活动,铁屑的腐蚀与抑制腐蚀作用处于一定的平衡,使得装置内总铁在运行12 d后基本维持在较低质量浓度范围。铁过高,会对微生物造成损伤[21],较低的铁释放保证了耦合体系强化脱氮的长期作用。
另外,铁型反硝化菌可以Fe2+为电子供体将NO3− /NO2− 还原为N2,但反应Fe/N比为 (3~5) :1[24],而1号总铁均值约为0.3 mg·L−1。铁自养反硝化虽可提高脱氮效能[24, 47],但不是耦合体系TN去除的主要途径。
根据实验结果,铁屑强化反硝化包括3个过程:1) 铁屑腐蚀,生成Fe2+及其氧化物[20] ,反应式见式 (3)~式 (5) ;2) Fe2+氧化,形成Fe(OH)3、FeOOH、Fe2O3、Fe3O4等氧化物[23] ,反应式见式 (2) 、式 (6)~式 (8) ;3) Fe2+、Fe3+及其各类氧化物是提升反硝化的关键,它们可强化微生物繁殖、活性、丰度和多样性等[21],减少有机质流失,提高电子传递效率等[19]。而铁离子与有机物间鳌和/络合反应[48]可促进系统与铁的作用,多种因素综合影响,最终表现为:改善碳源释放,增强脱氮效能。但目前Fe0生物系统强化脱氮机理复杂,尚不明确[21],还有待进一步深入研究。
-
图6为竹刨花被利用前后表面扫描电镜图。被利用前的竹刨花(图6 (a) 和图6 (b) )由于加工原因和竹纤维结构特点,有许多不规则碎片与孔隙,利于微生物附着生长。被利用近100 d后,1号和2号竹刨花均附着有微生物。这些微生物主要为杆状,已有研究[49]显示,固相碳源被利用后,其表面微生物多呈杆状。但1号竹刨花表面(图6 (c) 和图6 (d) )有许多交错黏结、大小不一的纤维絮体,同时微生物与微生物、微生物与纤维絮体之间黏附团聚,形成更为紧密的生物膜。而2号竹刨花(图6 (e) 和图6 (f) )被利用后,表面虽有一些残余纤维絮体,但数量少,表面相对平整,微生物分布也更加稀疏。密实的生物膜利于纤维素分解与污染物的去除,铁一方面刺激了微生物的生长及其胞外物的分泌,强化微生物、纤维絮体等的黏附,同时铁离子及其氢氧化物的絮凝作用进一步将浸泡分解出的竹纤维絮体吸附截留,增加微生物附着点,减少有机质流失,提高有机物的利用率。
-
Alpha多样性指数可表征微生物多样性[4],结果见表1。2组样本的覆盖率指数均大于0.99,表明该测序结果能较好反映菌群的真实情况[8]。Chao1值越大,群落丰度越好[50]。2组样本Chao1指数都大于2 000,说明2体统均具有复杂的群落丰度,但1号群落丰度明显高于2号。Shannon指数、Simpson指数和系统发育树指数反映群落的多样性,受样品群落中物种丰度和物种均匀度的影响。Shannon指数和系统发育树指数越大[51-52],Simpson指数越小[53],则菌群多样性越高。1号的Shannon指数和系统发育树指数均高于2号,而Simpson指数低于2号,说明1号耦合体系内生物多样性高于2号单纯固相反硝化体系。
-
1号与2号竹刨花填料表面微生物门水平群落组成如图7 (a) 所示。2体系细菌门类主要为变形菌门Proteobacteria (49.75%~66.51%) 、放线菌门Actinobacteria (11.39%~12.38%) 、拟杆菌门Bacteroidetes (11.21%~12.93%) ,厚壁菌门Firmicutes (3.27%~4.70%) 和绿弯菌门Chloroflexi (1.65%~3.77%) 等。2体系优势门为变形菌门,但1号变形菌门占比(65.51%)明显高于2号(49.75%)。此外,2号疣微菌门Verrucomicrobia丰度 (8.80%) 远高于1号 (小于1%) 。变形菌门是各类水环境反硝化微生物的主导门类[4, 54-55];拟杆菌门主要由水解菌组成,可降解纤维素在内的大分子有机物[56];放线菌门是丝状菌,具有絮凝作用[57],可降解氨基酸等;厚壁菌门可降解纤维素和进行反硝化[4, 58];绿弯菌门常见于各类污水处理中,是反硝化主要菌种之一[1];疣微菌门在活性污泥中较为常见,其与有机物、氨氮降解有关[59]。总体而言,变形菌门、拟杆菌门、放线菌门、绿弯菌门、厚壁菌门普遍存在于植物碳源反硝化系统中[12, 60],是反硝化主要参与者,2体系这些门类占比均高于80%,说明2体系TN去除主要为生物反硝化。2体系均存在降解有机物的微生物,如拟杆菌门、放线菌门、疣微菌门等。由于2号疣微菌门丰度较大,其有机物降解相关门类 (门水平,丰度>5%) 的总占比高于1号,说明铁屑未促进有机物降解相关门类生长。由于1号菌群总丰度、多样性、生物膜条件比2号更好,说明是各类因素的共同作用增强了耦合体系碳源释放。另外,严子春等[61]发现,富铁生物系统中疣微菌科丰度与空白组相比,显著降低,这与本研究类似,表明铁会抑制疣微菌门/科的生长。2体系门类差异最大的为变形菌门 (1号较高) ,其次为疣微菌门 (1号较低) ,说明铁屑强化了脱氮功能菌的生长。
图7 (b) 为竹刨花填料表面微生物属水平群落组成情况。由于一些序列不能归入已知属,本数据库以Family (科) 后缀-uncultured,-unclassified来表示。变形菌门是2体系占比最大、差异最显著的门类。表2为2体系变形菌门主要属组成 (属,丰度>1%) ,1号和2号变形菌门中优势菌属分别为假单胞菌属Pseudomonas和热单胞菌属Thermomonas。已有研究表明,Pseudomonas[17]、Denitratisoma[54, 62]、 Thermomonas [55]、Xanthobacteraceae_unclassified[61, 63]、Rhodocyclaceae_unclassified[7, 64]和Rhodocyclaceae-uncultured[65-66]、Desulfovibrio[7]、Rhizobiaceae_unclassified[67]、Devosia[55]、Bataproteobacteria_unclassified[67]和Gammaproteobacteria_unclassified[68-69]均是与反硝化有关的微生物。由表2可知:1号变形菌门中反硝化功能菌群共有9个类别 (表2中标记为DNB (denitrifying bacteria) 的菌属) ,总占比为35.75%;2号共有4个类别 (表2中标记为DNB的菌属) ,总占比为23.89%,表明耦合体系具有更多类别的反硝化菌群和占比,这有利于系统的稳定与强化脱氮[23-24]。
另外,与其他脱氮菌不同,1号优势菌属Pseudomonas是一种同步脱氮除磷菌[70]。已有研究[61]显示,富铁环境可明显促进Pseudomonas菌的生长并提升氮磷的去除率。当进水TP为 (1.14±0.06) mg·L−1时,0~105 d,1号和2号TP的平均去除率分别为75%和44%。因铁屑释铁少,故生物同步脱氮除磷是1号高效除磷的重要线索,也表明铁屑在诱导同步脱氮除磷菌方面潜力巨大。
此外,1号变形菌门中硫杆菌Desulforhabdus占比为1.05%,而2号未检测到该菌属。已有研究[24]显示,基于反硝化颗粒污泥的亚铁型厌氧铁氧化系统中的硫杆菌在Fe2+的氧化过程中起着重要作用。这证实耦合体系中存在Fe2+氧化反硝化,其可促进TN的去除,但硫杆菌丰度较小,说明Fe2+氧化反硝化不多,这与耦合体系总铁释放情况基本一致。
-
1) 进水TN为41.62~59.95 mg·L−1、COD/N<0.5、水力停留时间为18 h时,13~105 d,耦合体系碳源较单纯固相反硝化体系丰富,但2体系出水COD均值均低于一级A限值;耦合体系TN平均去除率为73.21%~92.79%,总体较单纯固相反硝化体系高近1倍,出水TN可达到地表水IV类标准;另外,耦合体系中铁释放稳定,出水总铁均值低于地表水环境质量标准限值。
2) SEM表征结果显示,竹刨花表面植物组织碎片与孔隙较多,适于微生物附着生长;被利用近100 d后,耦合体系竹填料表面明显黏附有许多纤维絮体,黏性物质与微生物数量更多,生物膜生长更加密实。
3) 16S rRNA表征结果显示,耦合体系与单纯固相反硝化体系相比,微生物丰度与多样性更高。2体系反硝化脱氮相关门类占主体,优势门类均为变形菌门,但耦合体系变形菌门丰度更高;在变形菌门主要属组成中,耦合体系反硝化菌群 (属水平,丰度﹥1%) 类别和总占比均高于单纯固相反硝化体系。这与耦合体系高效、稳定的脱氮效果基本吻合。
铁屑耦合固相反硝化对低碳氮比废水中总氮的处理
Treatment of total nitrogen in wastewater with low C/N ratio by iron shavings coupled solid-phase nitrification
-
摘要: 为考察脱氮效率并解决碳源不足导致总氮 (TN) 去除率不高的问题,将竹刨花作为固体碳源与挂膜载体,引入铁屑,构成耦合体系,搭建2套反硝化实验装置 (分别为耦合体系1号与单纯固相反硝化体系2号) 并对脱氮率及微生物进行分析。结果表明:进水TN为41.62~59.95 mg·L−1,COD/N<0.5,水力停留时间为18 h; 13~105 d,1号TN平均去除率为73.21%~92.79%,比2号平均去除率高近1倍;1号碳源相对2号更为充足,但2体系出水COD均值都低于一级A;1号总铁释放稳定,出水总铁均值低于0.3 mg·L−1,未出现NH3-N明显积累。SEM表征结果显示,1号竹填料表面黏性物质与微生物数量更多,生物膜更加紧密。16S rRNA表征结果显示:1号具有更高的微生物丰度与多样性;2体系反硝化脱氮相关门类占主体,优势门类均为变形菌门,但1号变形菌门占比高于2号;变形菌门中,1号反硝化菌群 (属水平,丰度>1%) 类别和总占比均高于2号。由此可知,铁屑强化了碳源的分解与利用,促进了脱氮功能菌生长,显著提升了耦合体系脱氮效能,出水TN可达到地表水IV类标准(GB 3838-2002)。竹刨花获取方便灵活,铁屑低廉,两者构成的耦合体系具有巨大的实际应用前景。Abstract: In order to investigate the denitrification efficiency and solve the problem of poor TN removal due to insufficient carbon source, bamboo flakes were used as a solid carbon source and biofilm carrier, and a coupled system was designed with the introduction of iron shavings. Then two sets of denitrification experimental devices were built (coupled system No. 1 and simple solid-phase denitrification system No. 2 , respectively) and their nitrogen removal rates and microorganisms were analyzed. The results showed that when the inlet TN was 41.62~59.95 mg·L−1, COD/N<0.5, and the hydraulic retention time was 18 h, the average TN removal rate of No. 1 was 73.21%~92.79% among 13~105 d, which was nearly 1 time higher than the average removal rate of No. 2; the carbon source of No.1 was more adequate than that of No.2, but the mean effluent COD values of No.2 were lower than that of Class A. The total iron release of No. 1 was stable, the average value of total iron in the effluent was less than 0.3 mg·L−1, and there was no significant accumulation of NH3-N. SEM characterization results showed that there were more sticky material and microorganisms on the surface of the bamboo filler of No. 1, and the biofilm was more compact. 16S rRNA characterization results showed that No. 1 had higher microbial abundance and species diversity; the denitrification-related phylums dominated in both systems, and the dominant microbial phylum were all Proteobacteria, but the abundance of Proteobacteria in No. 1 was higher than that in No. 2; among Proteobacteria, the category and total percentage of denitrifying group (genus level, abundance >1%) in No. 1 were higher than those in No. 2. It can be seen that iron shavings enhanced the decomposition and utilization of carbon source, promoted the growth of nitrogen removal functional bacteria, which significantly improved the efficiency of nitrogen removal in the coupled system, and the effluent TN could reach class IV of the surface water standard (GB 3838-2002). Bamboo flakes are convenient and flexible to obtain, and iron shavings are inexpensive, and their coupled system has great practical application prospects.
-
Key words:
- bamboo flakes /
- iron shavings /
- coupled system /
- denitrification /
- 16S rRNA
-
近年来,土壤污染问题日益严重。我国人口众多、可利用土地资源少,污染土壤修复问题已成为关注重点[1]。有机污染(农药、石油烃和多环芳烃)场地面积约占全国超标土壤的35.49%[2],高于其他类型的土壤污染。由于污染物种类繁多,污染场地土壤类型多样、空间差异大,对土壤修复技术提出了较高要求。现有的土壤修复技术正朝着5大方向发展,即绿色友好的生物修复、联合组配的原位修复、综合修复、基于设备化的快速场地修复和基于环境功能材料的修复[3-4]。原位热修复技术因其适用范围广、环境干扰小、修复效果彻底和可操作性强等优势受到广泛关注[5]。原位热修复技术是少数几种能够去除NAPL的技术之一,并且涉及使用载气或真空系统,该系统将挥发的污染物吹扫到气体处理系统,方便进行二次或异地处置。但是,由于土壤水分和异质性带来的复杂性,使得原位热修复技术大规模应用具有挑战性[6]。
污染土壤修复技术早在二十世纪70年代就已在国外开始应用[4],近年由于国内原位热修复需求日益增多,相关技术被逐步引进。根据加热方式不同,原位热修复技术主要分为蒸汽强化抽提技术(Steam-Enhanced Extraction, SEE)、电阻加热技术(Electrical Resistive Heating, ERH)和热传导加热技术(Thermal Conduction Heating, TCH)3种[7]。对于上述3种原位热修复技术,由于其加热机制不同,所适用的场合也各不相同。如SEE技术,仅适用于低沸点、高渗透率的污染物场地,其修复效率低但修复成本也较低;ERH技术同样仅适用于低沸点污染物,但其升温效率和温度均匀性较好;而TCH技术可适用于含高沸点污染物场地,修复效果彻底、周期短,但其修复方式粗犷,成本较高。表1对比了3种典型原位热修复技术的优缺点和各自适用场合。
表 1 典型原位热修复技术优缺点比较Table 1. Comparison of advantages and disadvantages of typical in-situ thermal repair technology技术手段 修复成本概况 加热方式 温度范围 适用场合 技术优势 技术缺陷 SEE 修复挥发性、半挥发性有机物成本较低[10] 蒸汽、热空气 0~170 ℃[7] 挥发性有机物,高渗透率土壤,地下流速大 去除效率较高、成本低、易操作、可与其他技术灵活组合 受土壤不均质性、渗透率、含水率影响大,处理温度低 ERH 修复成本高[10] 电阻加热 0~100 ℃ 挥发性有机物,中低渗透率土壤,适合质地细密的土壤[11] 对地下非均质性不敏感,加热均匀性强[8],可与生物修复联合使用[9] 不适合干燥的土壤,难以处理高沸点污染物[12] TCH 修复氯化挥发性有机物成本较高[13] 电加热棒、高温烟气 700~800 ℃ 挥发、半挥发、难挥发有机物,中、低渗透率土壤 升温速率快,加热温度高,不受渗透率、土壤质地的影响 能耗高,易受高地下水通量的影响 | Show Table DownLoad:
CSV
DownLoad:
CSV
虽然原位热修复技术的工程应用已较为成熟,积累了丰富的工程经验,但其内在的热质传递机理尚不清晰,由此,导致工程设计往往仅依赖于实践经验而缺乏相关理论指导。本综述将分别从理论、实验、数值研究等方面对原位热修复过程中土壤内热质传递的研究现状与进展进行讨论与分析,以期能够推动土壤原位热修复技术的优化,为工程应用提供参考。
1. 原位热修复过程中热质传递机理及影响因素
1.1 蒸汽强化抽提技术(SEE)
SEE技术由于其具有易操作、成本低等优势,早期被广为使用。 KINGSTON等[14]、康绍果等[15]在研究中提到,自1988年到2007年蒸汽加热技术的应用案例为46例,占总原位加热处理技术的25.2%。SEE技术的工作原理如图1所示,高温蒸汽或热空气通过加热井注射到深层土壤,提高污染区域温度,使得非水相液体污染物(Non-aqueous Phase Liquids, NAPLs)发生解析、溶解和蒸发,在注入气体的推动下移动至抽提区附近进行收集和后处理[16-18]。其热质传递机理为,在高温蒸汽作用下,NAPLs粘度降低、移动性增强;同时,当蒸汽与NAPLs混合时可能发生正共沸,从而降低了沸点。
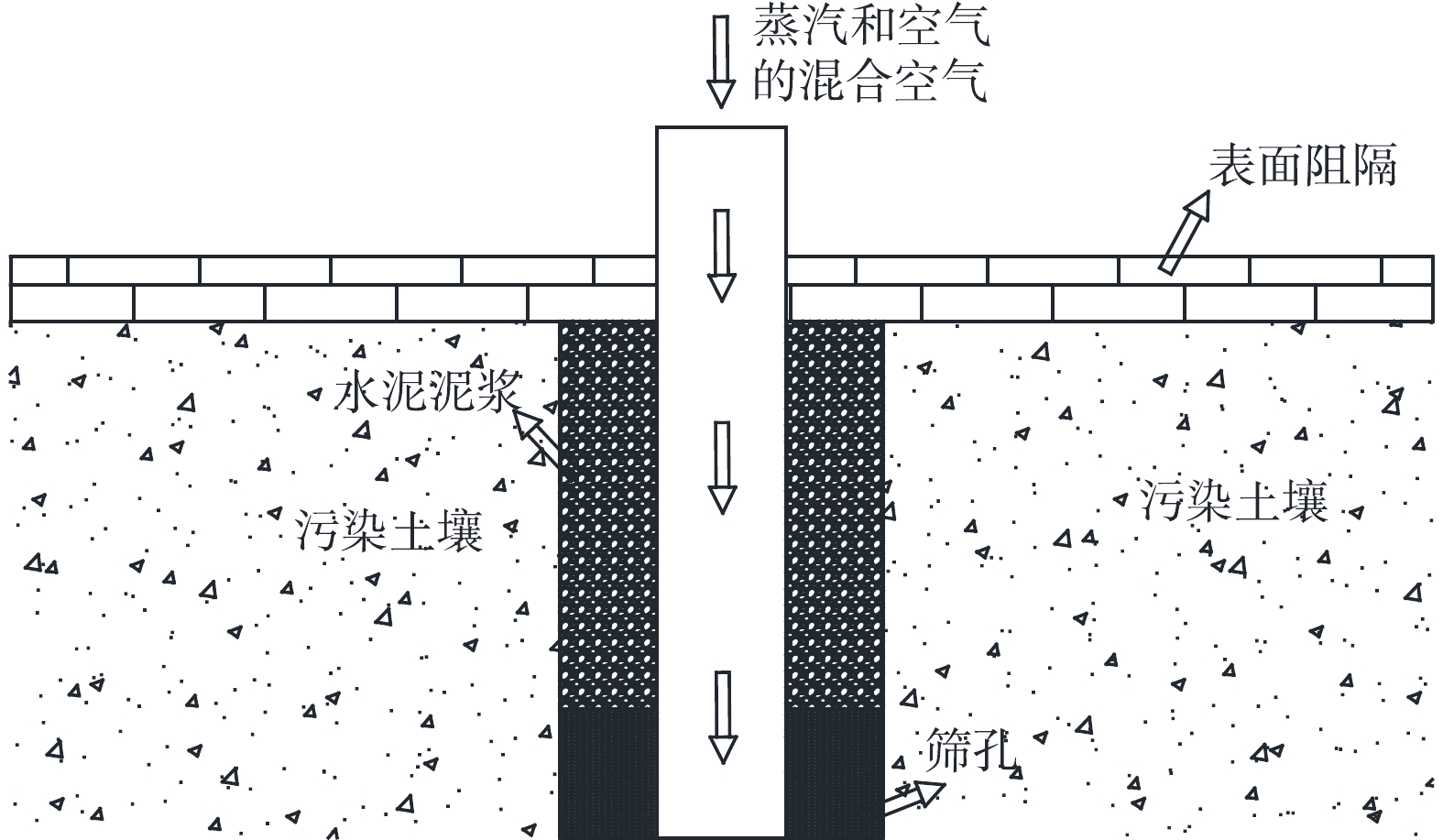 图 1 SEE技术工作原理示意图Figure 1. Schematic diagram of the working principle of SEE technology
图 1 SEE技术工作原理示意图Figure 1. Schematic diagram of the working principle of SEE technologySEE技术修复非水相液体污染物NAPLs的效果受其性质影响[19],如黏度和挥发性。当污染土壤内通入高温蒸汽,在毛细管压力和重力的共同作用下,驱动NAPLs、水和蒸汽的共同流动。SEE技术和轻油采集技术等都是通过注入高温蒸汽增强剩余非水相液体到蒸汽相的传质的方法,使得进入蒸汽中的NAPLs被运输到运移的前端并凝结,从而降低原NAPLs的黏度,并且高温蒸汽还增加特定区域的蒸汽压和分子扩散系数,其传热效果使NAPLs热膨胀,相对渗透率增大,这些机理的综合作用导致了NAPLs性质改变,蒸汽驱动效率高。
非均质性是SEE技术重要影响因素[20]。作为典型多孔介质的土壤,它的非均质性主要影响了修复区域的气体通透性,较容易形成优势通道,使得气体流经修复区域时沿着优势通道运移,因而出现“绕流”现象,减少了NAPLs随气流的去除量。土壤的非均质性还影响了热量的传递,使热在修复区域前进不均匀,这将导致去除效率的降低。土壤的非均质性会造成吸附性不一致,吸附性越大,去除效果越小。其他如通风流量、土壤含水率、污染物的饱和蒸汽压等也对该技术的修复有一定影响[21-23]。
1.2 电阻加热技术(ERH)
ERH技术是一种基于焦耳定律发热原理对土壤进行加热的修复方式[15],其原理图如图2所示。ERH的实现方法是将异相电极装在污染场地的不同位置,当通入电流时,土壤、电极和地下水形成回路并产生热量,从而加热土壤。当土壤温度接近沸点时,水分快速蒸发,此时水蒸汽携带污染物蒸汽向上运动;同时,由于水分的挥发,导电性降低,因此,在修复过程中需不断补充水分。有研究表明,电阻加热技术能使土壤快速达到100 ℃[9],但由于水分和土壤孔隙结构的限制,最高达到的温度约120 ℃。ERH技术并不是直接去除地下污染物,而是在物理、化学和生物条件方面产生变化,促进污染物从地下清除或就地转化为潜在的无毒物质[24]。
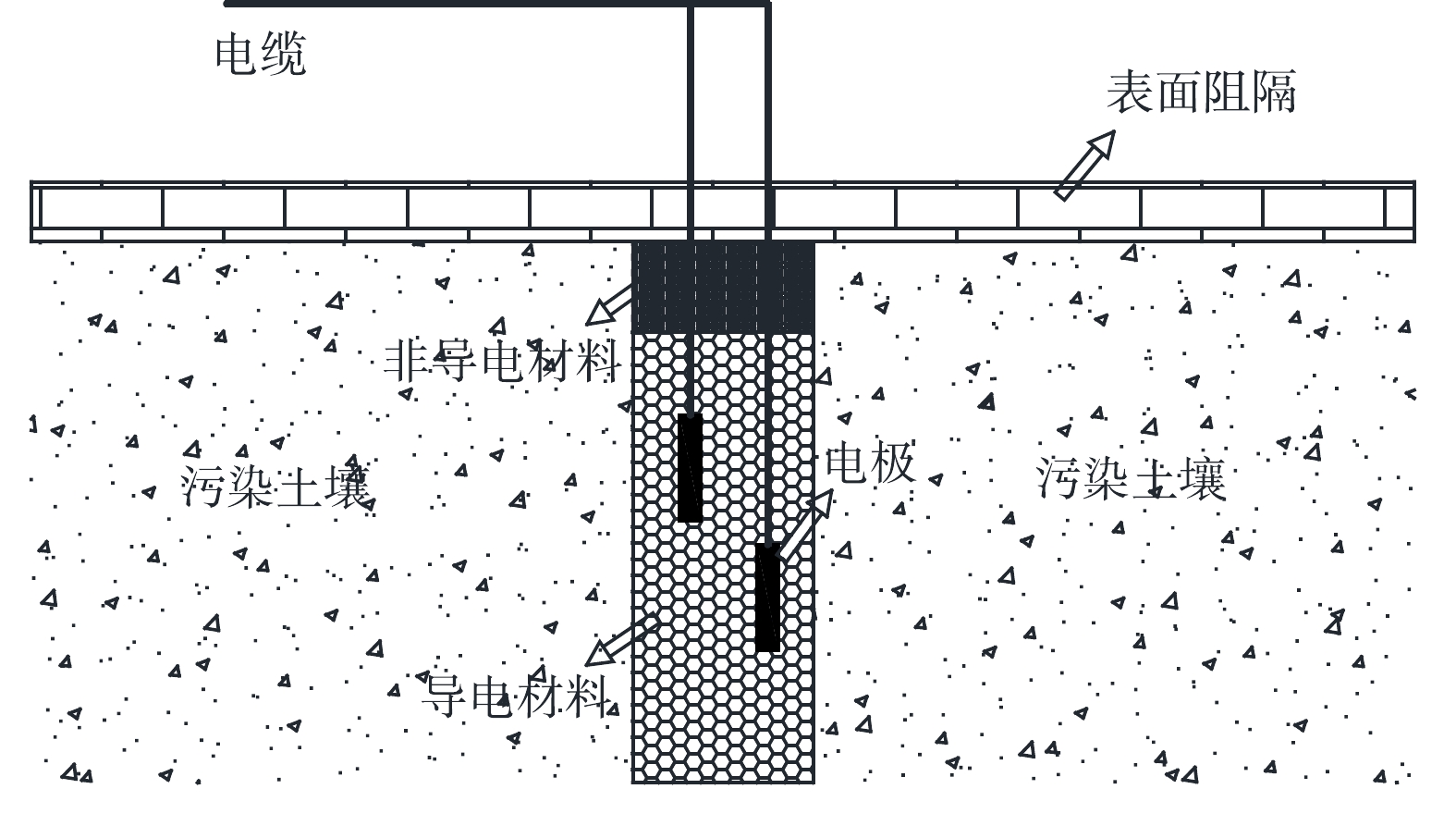 图 2 ERH技术工作原理示意图Figure 2. Schematic diagram of the working principle of ERH technology
图 2 ERH技术工作原理示意图Figure 2. Schematic diagram of the working principle of ERH technology在ERH技术修复过程中,含水率的影响至关重要[25]。有实验证明了含水量较高的土壤,其升温速率也高于干燥土壤[26]。此外,水分还能与特定污染物发生正共沸,从而有利于污染物的解析和挥发。HAN等[27]利用实验室规模ERH设备研究了多环芳烃去除效率的影响因素,水与多环芳烃发生正共沸被证实是ERH去除多环芳烃的主要机理。
电相分布是影响ERH技术修复效果的另一因素[28-30]。近年来,三相系统和六相系统已经发展成为加热低渗透层的可靠技术[31-33]。相比三相系统,六相系统加热均匀性更强,可有效防止加热区域中出现未升温区域,但是由三相电转化为六相电的成本较高[12],大规模应用受限,六相系统适用于对加热均匀性有硬性要求的实验或工程实践。目前,六相系统多应用于小试研究,而三相系统多用于现场[8]。有研究表明,电加热引起的热力学变化可以有效去除淤泥和粘土的氯化溶液[33-34]。电相之间的变换有利于加热区域的均匀性分布,六相系统加热土壤能提高污染物的去除。
此外,电极布置方式、污染物性质、土壤类型等也会对ERH修复技术产生影响[35]。最佳的电极间距不受土壤类型、水饱和度和地下电导率的影响。电极间距越近,温升越快,修复周期越短,但成本较高。由于土壤类型的不同,其导电系数相差较大,最大可达几百倍,因此修复的难易程度及修复效果也相差较大[36]。另外,电阻加热会降低NAPLs的黏度和与水的界面张力,增强溶解性,使去除速率加快。上述因素都会不同程度地影响ERH技术的修复效果。
1.3 热传导加热技术(TCH)
TCH技术利用电加热棒或高温烟气对污染土壤进行加热,热量通过热传导和热对流方式在污染地块中进行传递,土壤内的水分和污染物受热蒸发,由抽提井捕集并输送到地面进行处理,其原理示意图如图3所示。TCH修复过程中涉及了热量传递、水分迁移、蒸汽扩散、多相流动、气液相变和污染物脱附及迁移、热解等机制。BAKER等[36]和LACHANCE等[37]阐述了饱和多孔介质中挥发性有机物TCH修复的相关机制,其中包含了蒸发、蒸馏、沸腾、水解、氧化和热解,并认为最重要的去除机制是蒸发。
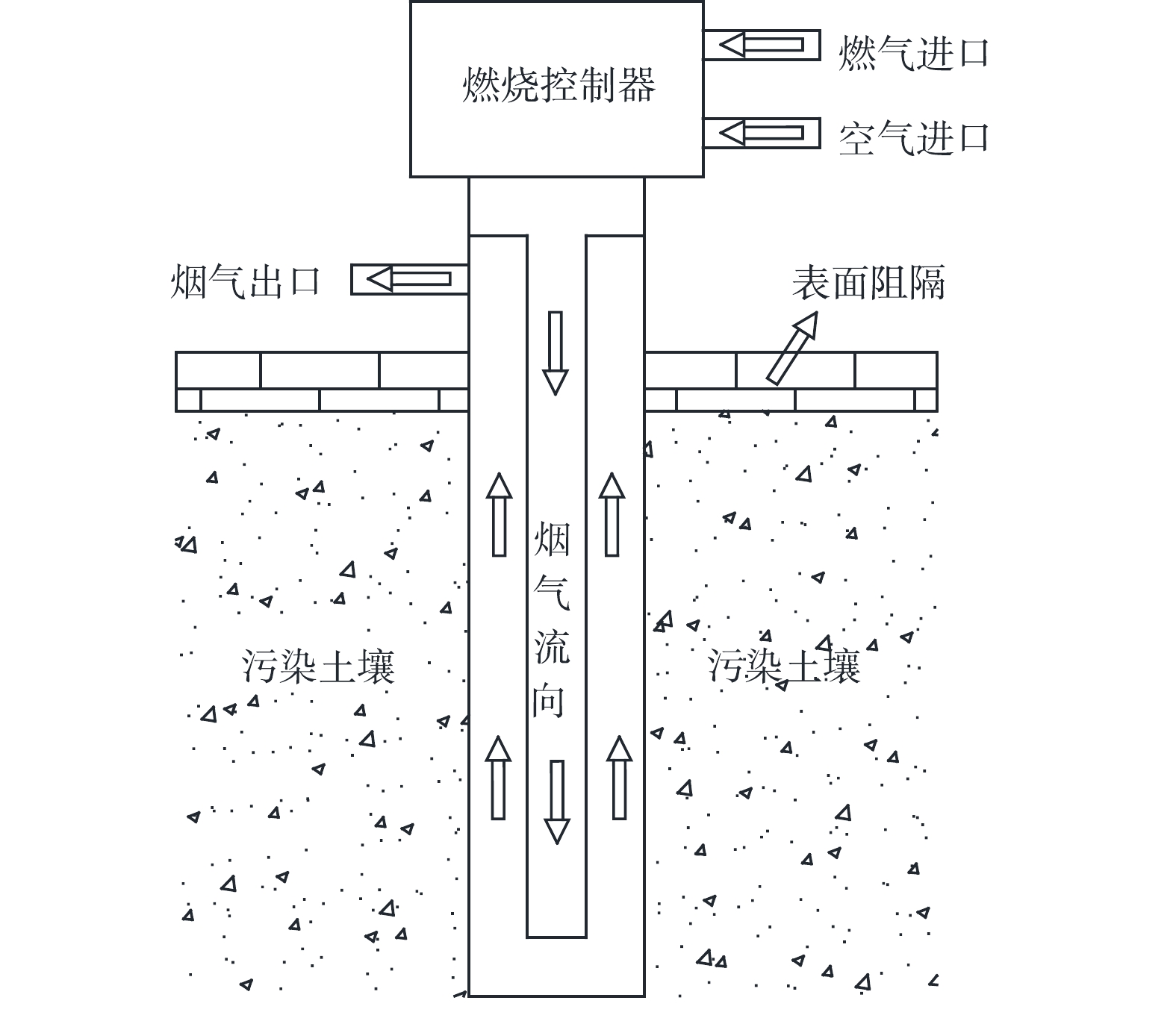 图 3 TCH技术(燃气加热)工作原理示意图Figure 3. Schematic diagram of the working principle of TCH technology (gas heating)
图 3 TCH技术(燃气加热)工作原理示意图Figure 3. Schematic diagram of the working principle of TCH technology (gas heating)加热温度是影响TCH修复过程的主要因素[38-41]。温度越高,修复周期越短,但能耗和成本也较高,因此,需要进行平衡,一般选取加热温度为500 ℃[42]。研究表明[43],当加热温度达到一定程度后,升温速率放缓,此时提升温度将导致能量大幅损失。加热温度过高对于环境修复并不可取,在较低温度下,通过延长加热时间也可达到较好的修复效果。
此外,升温速率也是影响TCH热脱附的重要因素。加热速率能够直接控制土壤和载气之间的传热速率以及解吸和降解速率,从而影响去除过程的效率[44]。白四红等[45]对多氯联苯污染土壤热脱附的实验研究表明,升温速率增大,降解速率升高,脱附效率下降。其主要原因是,在低温区间主要发生脱附反应,升温速率越慢,脱附反应的时间越长。
2. 原位热修复过程中热质传递机制研究进展
土壤作为典型的多孔介质,它包含了土壤固体颗粒、土壤溶液和气体等,因此,在研究土壤热质传递时,利用多孔介质现有的研究成果不失为一种好的方法。土壤中湿分的存在状态对土壤的热质传递特性影响很大,因此,依据土壤中水分含量将土壤的形态大概分为3类:湿饱和土壤(空隙内充满液态水)、非饱和土壤(空隙内同时存在气态和液态水)、干饱和土壤(空隙内只存在气态水)。在自然状态下,土壤大多为包含固体颗粒、水分、蒸汽和空气的非饱和多相系统。在对污染土壤进行原位热修复过程中,科研工作者应充分考虑大气环境等外部因素和土壤物理结构等内部因素对湿分场、温度场及污染物场的影响。然而,这一过程很复杂,难以简单地利用实验方法进行分析,下面将从理论研究、实验研究及数值研究3方面进行阐述。
2.1 理论研究
由于土壤特性和热质传递过程的复杂性,在进行理论研究之前,需要对研究对象进行一定的假设处理。如将土壤视为均质各向同性的不可压缩介质;液相和气相均为连续,忽略气体吸附效应;土壤内空气和水蒸气为理想气体;各相处于局部热平衡;忽略土壤内的热辐射、流体压缩功和粘性耗散;忽略有机污染物相变潜热;忽略重力等。土壤多孔介质主要热质传递机理如图4所示,将迁移机制归纳为热量迁移、湿分迁移、污染物迁移以及耦合迁移4类。
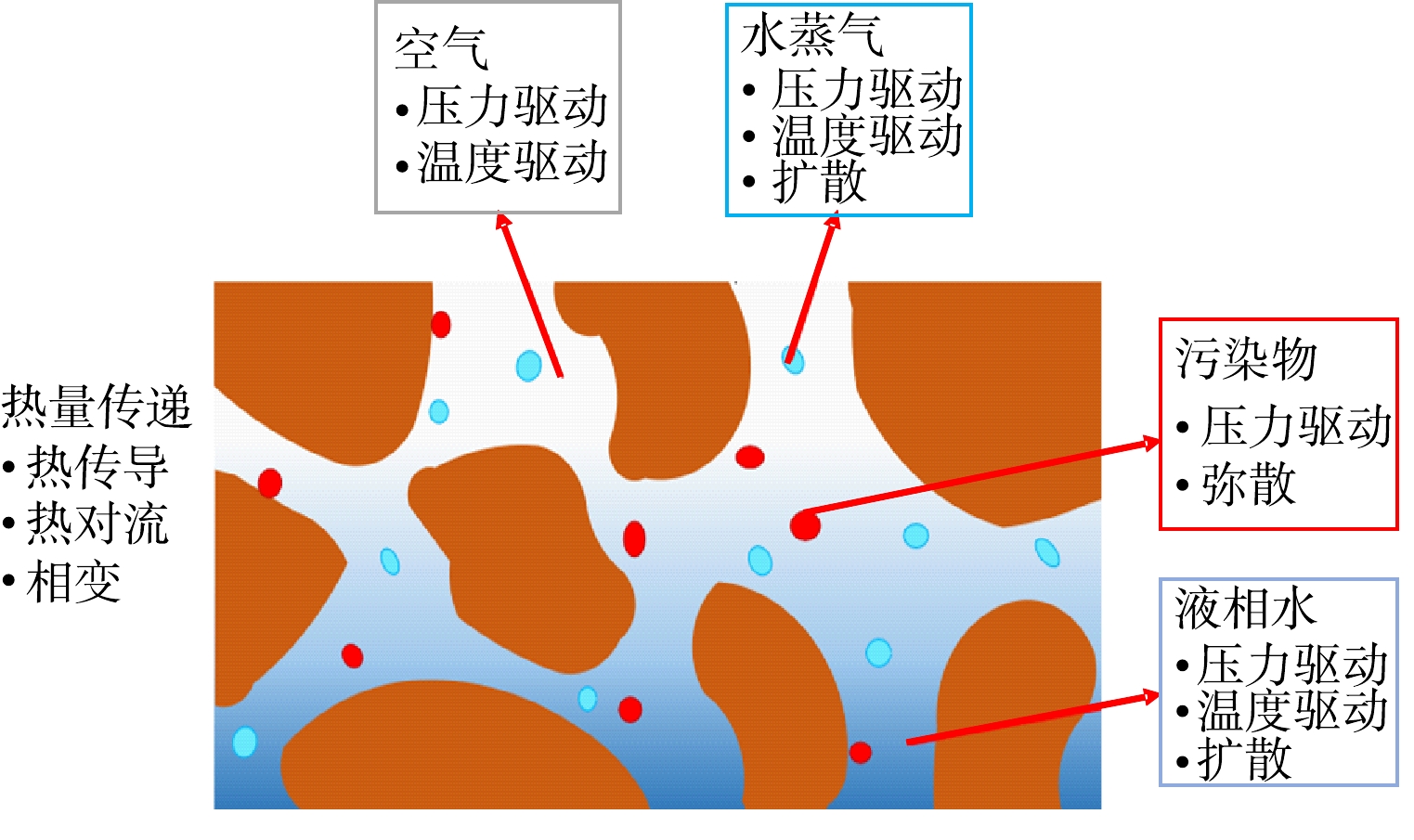 图 4 土壤内热质迁移机制示意图Figure 4. Schematic diagram of heat and mass transfer mechanism in soil
图 4 土壤内热质迁移机制示意图Figure 4. Schematic diagram of heat and mass transfer mechanism in soil土壤内湿分迁移包括液相水和水蒸汽。液相水的迁移机制研究从Darcy提出著名的达西定律开始[46-47]。此后,众多研究者提出了相关理论。SHERWOOD和COMINGS[48]在固体干燥研究过程中发现了液态水的运动是在重力、外加外力或毛细作用引起的压力差的影响下发生,且认为在一般干燥过程中液体运动是由于毛细作用引起。DE VRIES和PHILIP[49-50]以土壤为研究对象,考虑到之前温度梯度下简单的水汽扩散理论,忽略了水蒸气、液相和固相的相互作用以及土壤整体温度梯度的差异等关键问题,故在多孔介质领域首次提出以温度梯度和湿分梯度为推动势的双场热-湿耦合传递数学模型。其中,液相迁移基于非饱和Darcy定律而蒸汽迁移基于Stefan扩散定律。该模型被广泛应用于地下储能和地源热泵(地埋热管换热器)的研究[51-53]。LIU等[54]对上述模型进行了改进,采用连续介质模型和体积平均法,对自然条件下干湿分层土壤内热湿迁移发展了相关理论模型;同时,考虑了液态水、水蒸气和不凝性空气的迁移以及气液相变对热质传递的影响,该模型具有较强的普适性,但方程过于复杂,不利于数值求解。到目前为止,湿分迁移机制主要有压力梯度引起的渗流(基于非饱和Darcy定律),分子扩散(基于Fick扩散定律),温度梯度引起的扩散(Soret效应),以及由对流、扩散和相变引起的水蒸气迁移等。
土壤内热量传递主要包括热传导、对流和相变换热。其中,热传导发生于固体骨架、液体和气体之间,由傅里叶导热定律确定;热对流包括液相和气相的对流和扩散,相变潜热主要为水的气液相变。LARBI[55]研究中阐述了LUIKOV将不可逆热力学应用到多孔介质热质传递的研究,在1954年建立了以温度和湿度为参变量的热质耦合双参数理论模型,1964年提出了包含温度场、湿度场和应力场的偏微分控制方程组,期望能够描述热质传递过程中各驱动力与因变量的关系;但方程组中的唯象因子不全都具有实际的物理意义,在计算过程中需要大量的假设和经验参数,缺乏科学的理论依据。在传热过程中,热导率和比热容是关键参数。籍龙杰等[56]指出,土壤热导率受温度和含水率的影响,随含水率的增大,土壤热导率缓慢增加,最后趋于平稳;而比热容随温度先增大后减小。
土壤内污染物的迁移方式包括随液相水的运移以及在气相中的流动。在液相水中的迁移机制包括对流、分子扩散、机械弥散以及挥发,其中,附着于固体表面和游离在液相水中的污染物通过物理方式进入气相。NIELSON等[57]结合溶质迁移基本原理和连续性方程,首次提出了对流-弥散方程,这使得研究热-湿-污染物的相互作用成为一种可能。NASSAR[58]在研究封闭土柱的热质传递问题过程中发现,土壤冻融循环过程的热质传递受多种相互作用机理的影响,其中土壤含水量和土壤溶液浓度对结冻深度有较大影响。CLASS等[59]和CLASS等[60]提出水-气-NAPL模型,用于描述低压(大气压力数量级)下的非等温水-气-NAPL系统,利用该模型验证了热管效应等,缺点是忽略了固体基质中污染物的吸附作用。曾敏等[61]采用曲线坐标系下压力与速度耦合的 SIMPLEC算法,模拟复杂多孔介质腔体内的自然对流换热问题,讨论了Darcy数和孔隙率等对自然对流换热的影响。杨通等[62]利用有限元分析软件ADINA对多孔介质热流固耦合进行了分析,以热力采油为例,分析了注蒸汽以及流体流动等因素对采油的影响。
耦合迁移主要包括热-湿迁移和热-湿-污染物迁移,其中热-湿迁移研究机理较深入。WANG等[63]提出了应用于多孔介质的两相输运模型,此模型为理解多孔介质复杂的两相流动开辟了前景,但未包括惯性流动,不适用于全部非达西定律流动。JIN等[64]建立非了饱和土温度和水分时空分布数值模型,但该模型仅考虑热传导对温度和水分分布的作用,未考虑水分运移的影响。GAO等[65]建立了温度梯度和湿度梯度双驱动的一维非饱和土热湿传递模型,研究热效应下的非饱和土热湿耦合作用,经实验和数值模拟得出土壤水分传递通量与热源温度呈正相关。该项研究还发现,在相同的热源温度下,水分含量低的土壤具有较强的水分传递,水分含量高的土壤具有较强的传热能力,这为研究热湿传递理论奠定良好基础。
为了研究多相传热传质过程,HU等[66]提出了一种基于新能量方程的多孔介质流动沸腾修正多相混合模型。该模型利用动力学焓对多相混合模型的能量方程进行重构,以提高模型的收敛性和精度,消除了非物理跳跃,保持单相和两相区域之间的平稳过渡,并通过蒸腾冷却实例,研究了低压对温度分布的影响,揭示了动态响应延迟和蒸汽堵塞效应的潜在风险。WHITAKER[67]做了必要的假设对表征体元采用体积平均理论和引入连续介质力学相关理论,建立了基于连续介质模型的热质传递模型,将热质传递过程从微观水平提升到了宏观水平。这为进一步的理论工作和复杂多相过程模型的建立提供良好的基础,该方法最大缺陷是拥有大量的待定传输系数。
此外,一些研究者将热质传递对象的某些结构和参数也纳入考虑范围。白苗苗[68]认为,目前所建立的传统模型没有充分考虑土壤物理结构的影响,只是将其假设为连续性介质。在土壤热质传输过程中,不能真实反映多孔介质内部复杂的拓扑结构,也不能合理解释一些如优势流现象等,故提出将孔道网络理论引用到土壤热质传递研究,并通过相关模拟和试验的对比分析得出所建立的模型能够较好的描述实际土壤的热质传递过程。沈妍等[69]通过双向热流固耦合研究,分析了压降和骨架微观变形机制,并进行了多孔介质水力性能测试。LIU等[54]认为,湿分扩散系数、水力传导系数和有效导热系数是影响热湿迁移的关键参数,这些参数受温度和饱和度(含水率)的共同影响,并提出了它们之间的函数关系。多场耦合的迁移机制本质上是各种参数之间的内在耦合关系,且各因素相互牵连,十分复杂。图5表现了有机污染土壤内典型传质过程。
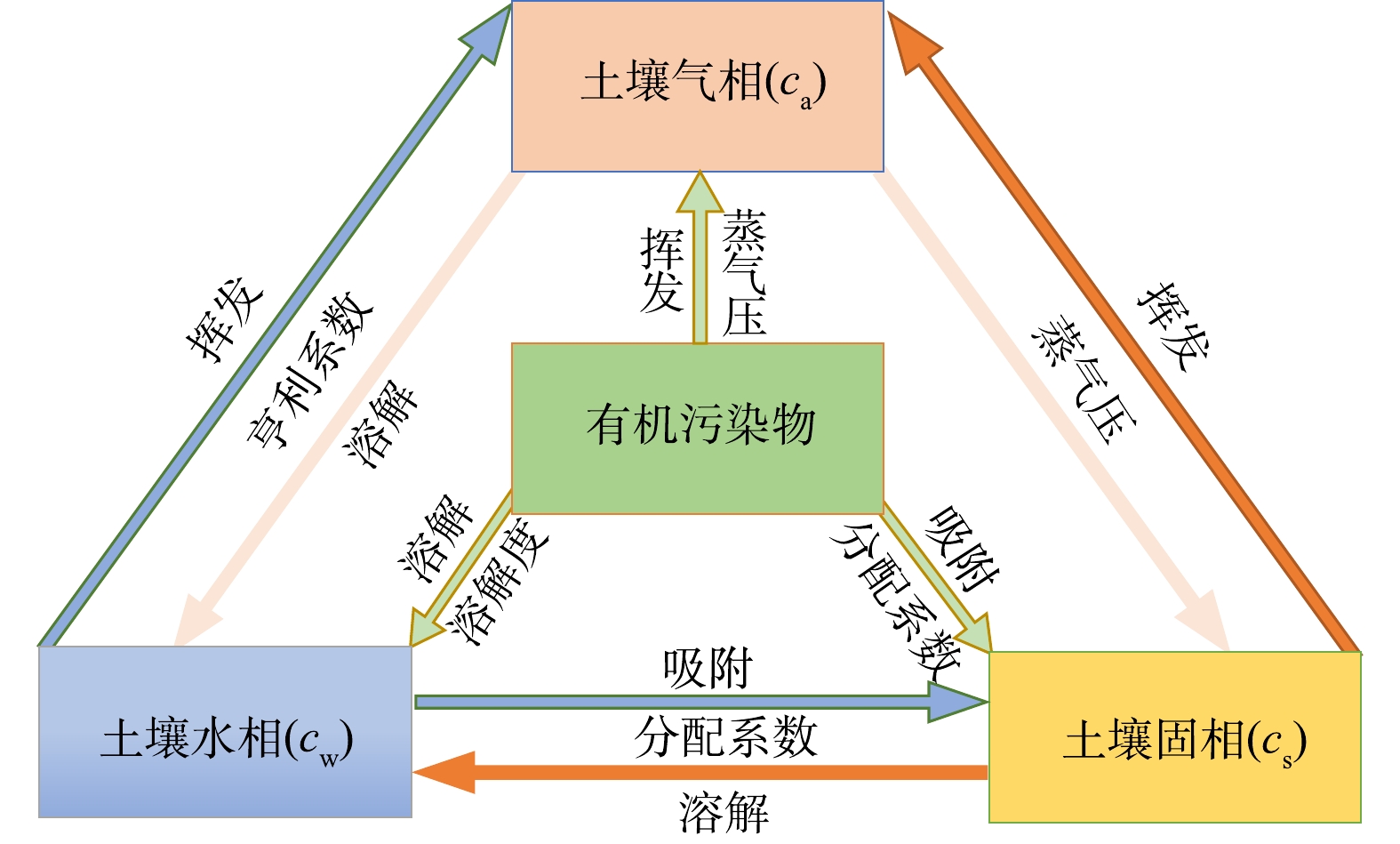 图 5 有机污染土壤内物质输运过程示意图Figure 5. Schematic diagram of material transport process in organic contaminated soil
图 5 有机污染土壤内物质输运过程示意图Figure 5. Schematic diagram of material transport process in organic contaminated soil2.2 实验研究
实验研究是理论研究的前提,理论研究为实验研究的结果,且实验研究还可以验证数值研究,故实验研究的重要性不言而喻。王华军等[70]建立了土壤高温储热热湿迁移实验台,研究80 ℃下热湿耦合迁移规律,发现高温储热条件下,温度场和湿度场之间具有较强的关联性。CHEN等[71]依托地源热泵课题搭建一维水平简化土柱实验平台,研究土壤充热过程热湿传递的耦合作用,分析了热源温度和初始湿度对土壤中热量和水分传递的影响。FORSYTH[72]以控制容积有限元法为基础建立了DNAPL污染现场蒸汽冲洗数值模型,并利用非线性磁通限制器求解多相、多组分地下流动方程,分析得出蒸汽注入与空气注入相比,蒸汽注入对注入井周围土壤的非均质性并不敏感,这为SEE技术的应用提供了理论指导;但该方法具有一定的缺陷:一是过渡使用磁通限制器会导致少量NAPL的虚假外观,二是算法的收敛性在某些方面会很差。CHERATI等[53]对单根储热桩周围的土壤热湿迁移进行了研究,利用格林函数将Philip提出的热-湿耦合理论模型的偏微分方程组转化为常微分方程组,获得的解析解与实验数据吻合较好。表2为几种典型原位热修复过程多场耦合案例。
表 2 原位热修复多场耦合研究现状Table 2. Research status of in-situ thermal repair multi-field coupling污染物 处理方式 实验装置 研究变量 参考文献 三氯乙烯 ERH 沙箱 TCE浓度、亨利定律常数 [34] 燃油 ERH 管式炉 土壤、温度、含水率 [73] 四氯化乙烯 ERH 沙箱 温度、硫酸盐浓度 [74] 多环芳烃 ERH 坩埚+管式炉 水分、盐分、电强度 [75] 三氯乙烷、氯仿 ERH 沙箱 温度、电流、地下水流 [76] Cr污染 ERH 沙箱 柠檬酸电解液浓度 [77] 笨和多环芳烃 TCH 燃烧器 温度、压力 [78] 非水相液体、四氯乙烯 TCH 二维水槽 温度、时间 [36] | Show TableDownLoad:
CSV
2.3 数值研究
数值研究来源于理论研究,数值研究以控制方程为核心,以相关软件为手段,以建立相应几何模型,做出合理假设以及设立相应边界条件为具体措施,扩展研究原位热修复技术的影响因子,从而达到优化实施方案,降低能源损耗的目的。其中,边界条件主要包括流动边界条件和温度边界条件。针对土壤水分运动情况设置流动边界条件,针对热量交换情况设置温度边界条件。控制方程主要包括质量迁移方程和热量迁移方程。
液相水迁移控制方程(包括压力梯度引起的渗流、分子扩散和热扩散项、相变源项)如式(1)所示。
∂(ρlθl)∂t+∇(ρlθl→Vl)=∇(ρl(DT,l∇T+Dθ,l∇θl))−˙m (1) 式中:ρl为土壤内液相密度,kg·m−3;θl为土壤内液相体积分数,该参数与孔隙度直接相关,可影响沸腾阶段维持时间;
→Vl ˙m 水蒸气迁移控制方程(包括压力梯度引起的对流项、扩散项、相变源项)如式(2)所示。
∂(ρvθg)∂t+∇(ρvθg→Vg)=∇(Dv∇ρv)+˙m (2) 式中:ρv为土壤内水蒸气密度,kg·m−3;θg为土壤内气相体积分数;
→Vg ˙m 气相(包含水蒸气、不凝性空气、污染物蒸汽)整体迁移方程(包括对流项和相变源项)如式(3)所示。
∂(ρgθg)∂t+∇(ρgθg→Vg)=˙m (3) 式中:ρg为土壤内气相密度,kg·m−3;θg为土壤内气相体积分数;
→Vg ˙m 液相污染物迁移方程(包括对流项、弥散项和热脱附源项)如式(4)所示。
∂(θlCl)∂t=−∇(→VlCl−θlDl∇Cl)−˙W (4) 式中:θl为土壤内液相体积分数;
→Vl ˙W 气相污染物迁移方程(包括对流项、弥散项和热脱附源项)如式(5)所示。
∂(θgCg)∂t=−∇(→VgCg−θgDg∇Cg)+˙W (5) 式中:θg为土壤内气相体积分数;
→Vg ˙W 热量迁移方程(包括固、液、气的热传导,气、液的扩散和对流换热,水分蒸发冷凝的相变潜热)如式(6)所示。
∂∂t[(ρc)mT]+cp,lT[→Vl∇(ρlθl)]+cp,gT[→Vg∇(ρgθg)]+cp,lθl[→Vl∇(ρlT)]+cp,gθg[→Vg∇(ρgT)]=∇(λm∇T) - γ˙m (6) 式中:ρl、ρg分别为土壤内液相和气相密度,kg·m−3;θl、θg分别为土壤内液相和气相体积分数;
→Vl →Vg ˙m 在选定控制方程的基础上,借助仿真软件对选定研究对象进行计算,从而得到目标结果。WANG等[79]就现场热修复过程中的传热性能,提出了原位土壤热修复的质量模型和热模型,利用COMSOL数值仿真软件对土壤加热过程中的传热特性进行了研究,分析得出地下水渗流对土壤温度和含水量的分布有显著影响。龙回龙等[80]利用计算流体力学CFD方法建立了耦合低温水气变换反应的多孔介质二维拟均相反应器模型,对反应器内组分浓度、速度场及黏度进行了可视化分析。李绪萍等[81]采用数值计算方法研究了多孔介质内自然对流时驱动力与流动阻力的关系,进而探讨了多孔介质中物质和能量的输运问题。ZHU等[82]利用CMG’S STARS软件研究了ERH、VAPEX混合修复中电极布置、电压、井距和非均匀性效应等因素对混合修复过程的影响,发现混合过程横向格局表现良好。宋明启[83]基于粗宏观表征体元RMV科学提出了多孔介质保温材料的热质传递“三箱”模型,但该模型只涉及导热过程,没有对流和辐射传热过程。LIM等[84]发现,过去对土壤蒸汽提取效率的研究,主要应用一维和二维模型,但这不能完全模拟非均质性土壤中复杂流体流动和运移过程,故采用COMSOL软件去识别质量传递值,预测了实验室中污染土壤去除NAPL的情况。该仿真结果表明,瞬态传质系数对SVE“拖尾”效应具有重要意义。赵静[85]就淋洗修复、表面活性剂强化修复、原位生物修复等问题,采用FLUENT软件模拟了石油污染物在流体及土壤相互作用下的迁移及浓度变化,其模拟结果与实验结果总体趋势一致,吻合度较好,对现场修复具有一定的指导意义。BORTONE等[86]利用COMSOL软件模拟了地下水和污染物流100年内的变化,提出了渗透性吸附屏障(PABS)模型,数值模拟再现了在含水层和阻挡层内部发生的运移和吸附现象,现场数据验证了模型污染物拦截和降低浓度的有效性。
3. 问题与展望
3.1 热质传递机理研究
原位热修复热质传递是一个多物理场动态耦合问题,目前没有合适的模型能涵盖所有问题[87],对热质传递机理认识仍不够充分,有必要对其理论模型开展更为深入的研究。主要原因为:1)非饱和流动和饱和流动的差异性,使现有饱和流动模型无法应用到非饱和流动研究;2)多孔介质热质传递的复杂性,流体在微小的骨架孔隙空间流动时,各组分含量不断变化,使得物性也不断变化;3)非饱和多孔介质中物质间相互作用复杂,包括毛细力、固体颗粒对液体的Darcy阻力、气体与液膜之间的相互作用、液膜运动的惯性力及自身的重力等;4)多孔介质的换热机制复杂,包括固体骨架、流体和气体的导热、蒸发-冷凝交换的潜热换热、蒸汽和液体的扩散、对流和辐射换热等;5)目前有许多测定液体扩散系数、水分、水分传递势和比的实验方法,这些复杂的物理值称为土壤的传质特性,但这些参数的测定仍存在一定误差,这增加了传质机理预测的难度。
现有热质传递机理研究集中在单场单组分和双场耦合研究,随着研究的深入,多相多组分多场耦合研究也进入研究者们的视线。国内外已经提出了一些适用于多相耦合的热质传递模型,但该模型往往只是在个别实验中适用,并不具有广泛适用性。未来的研究方向将会向多组分多场耦合热质传递模型发展,期望发展一个通用的数学模型。
3.2 热质传递数值研究
目前,国内外多孔介质多相多组分的热质传递研究主要以模型与机理研究、应用研究和实验研究为主,由于包含多相流动并伴随流体相变的多孔介质内部的流动及各相含量的变化都难以通过实验测量,故模型与机理研究相对较多。其中,以数值研究为主导的模型与机理研究较为薄弱,多数研究者主要采用“实验为主、数值为辅”的研究思路。导致这种情况的主要原因为:1)参数化问题,目前的模型很多是由经验公式获得的,但迄今为止的水热参数经验公式还不完善,没有完全解决其机理性问题[88];2)现有科研工作者往往采用一种软件去验证自己提出的模型或者结论,但由于不同软件的侧重方向不同,这将导致模拟结果不同;3)因土壤实际条件十分复杂,同样的模型在不同的条件下(土壤质地、含水量、土壤温度等)其模拟效果不同。
有关热质传递数值研究,未来的主要发展方向可能基于热质传递数值模拟方法,将数值模拟的结果与大数据结合,利用统计方法确定模拟关键参数,分析对污染物时空分布规律的影响,这有助于进一步明确耦合作用关系,快速分析不同情况下的修复效果从而指导工程实践。
-
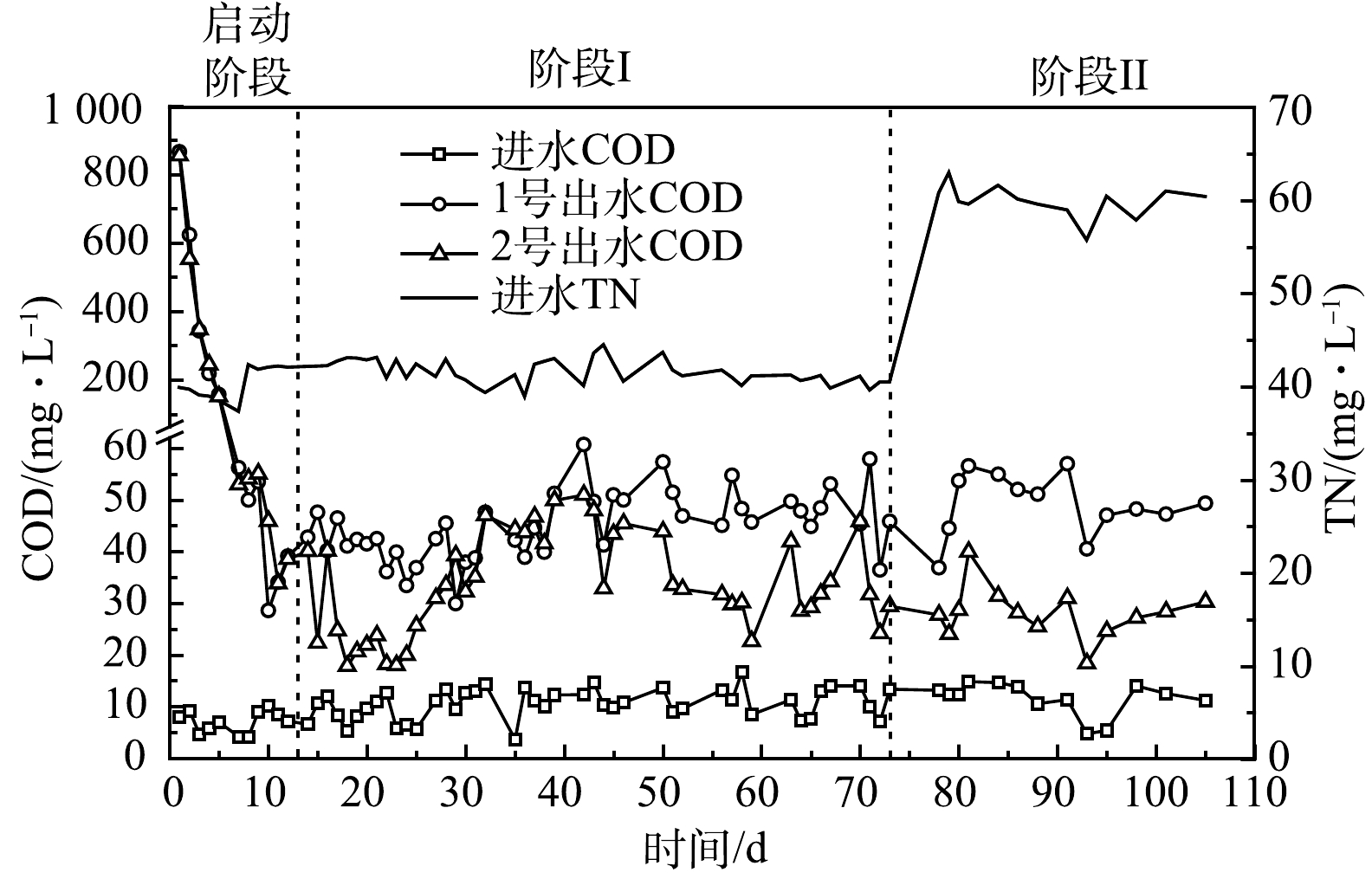
图 2 实验装置进、出水COD变化
Figure 2. Variation of COD in the influent and effluent of the experimental devices
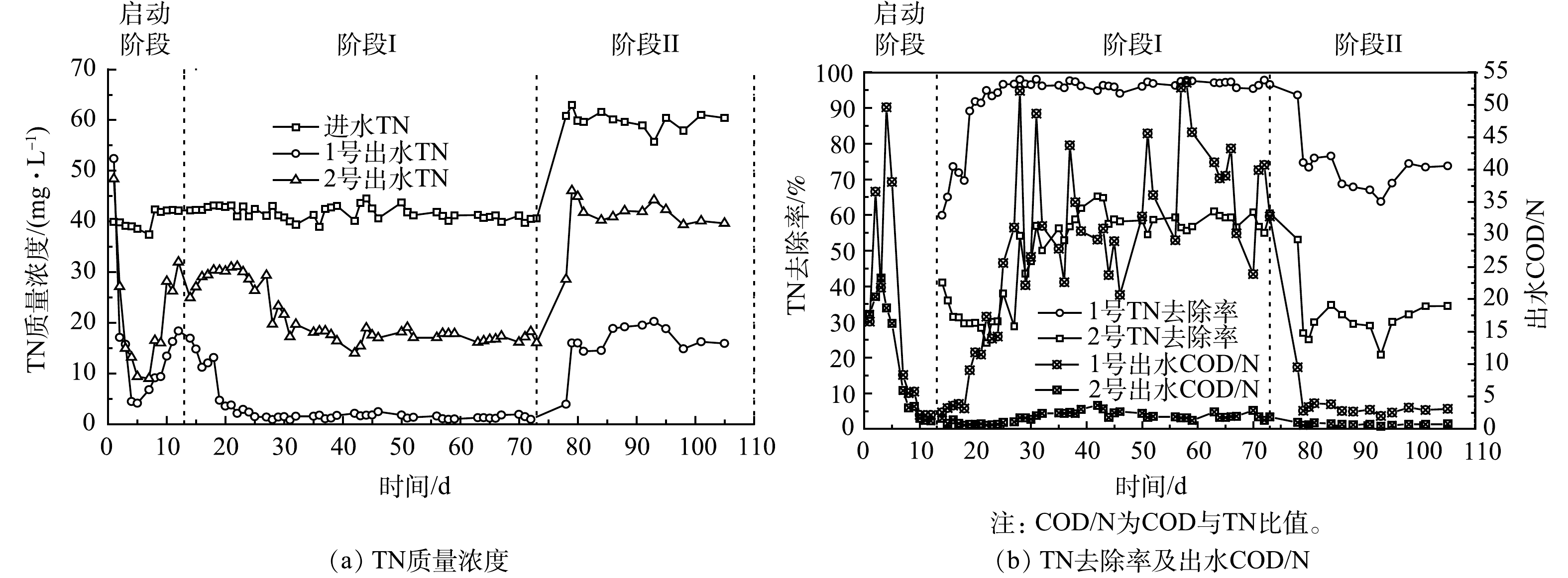
图 3 实验装置TN去除及出水COD/N曲线
Figure 3. TN removal and the effluent COD/N variation curve of the experimental devices
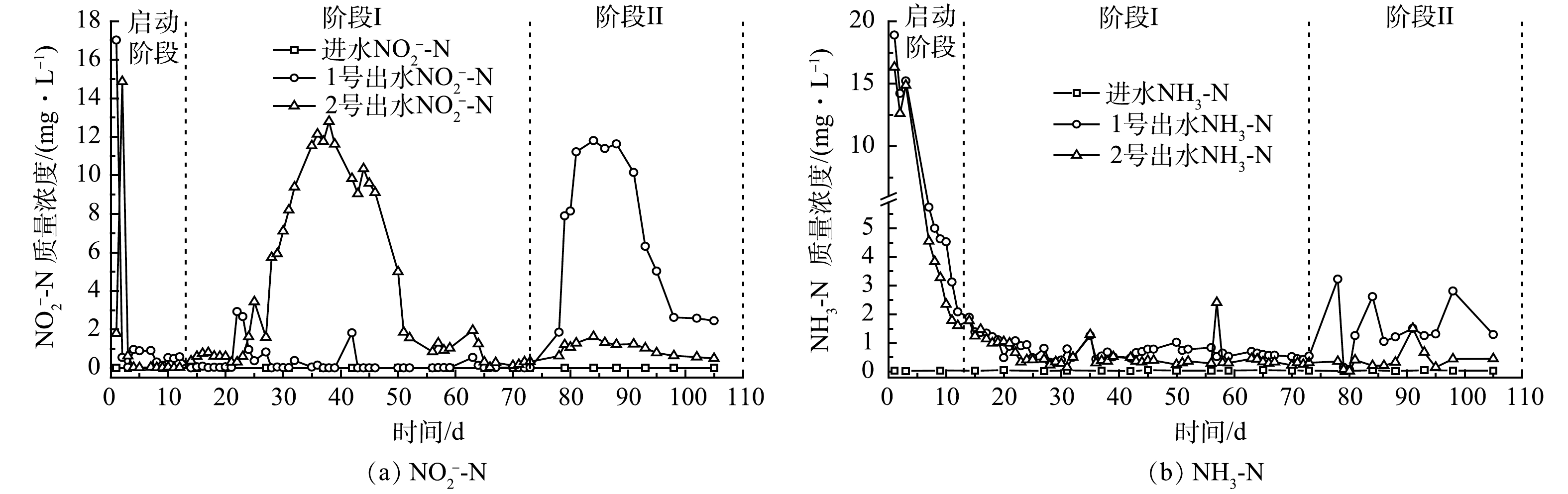
图 4 实验装置NO2--N与NH3-N积累情况
Figure 4. Accumulation of NO2--N and NH3-N in the experimental devices
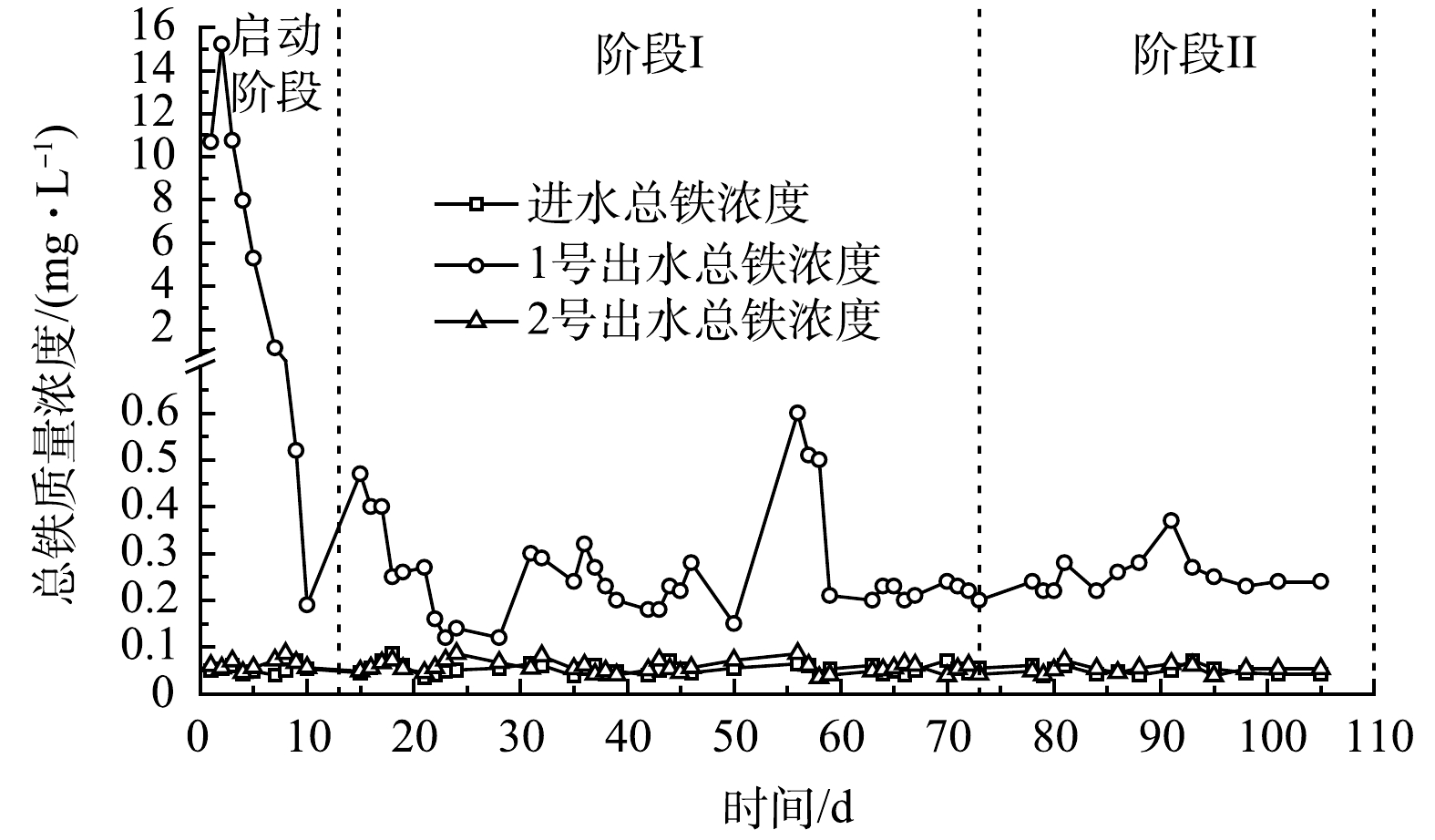
图 5 实验装置进出水总铁浓度变化情况
Figure 5. Variation of total iron concentration in the inlet and outlet water of the experimental devices

图 6 竹刨花填料被利用前后表面扫描电镜
Figure 6. SEM images of the surface of fresh and used bamboo flakes

图 7 填料表面微生物门、属水平群落组成
Figure 7. Analysis of the community composition of microbial phylum and genus levels on different filler surfaces
表 1 微生物Alpha多样性指数
Table 1. Microbial Alpha diversity index
样品 Chao1指数 Simpson指数 Shannon指数 观察到的物种指数 系统发育树指数 覆盖率指数 1号 3 238.29 0.012 6 8.975 7 3 230.1 126.485 0.996 0 2号 2 276.47 0.025 5 7.772 2 2 265.2 105.928 0.996 2
下载: 导出CSV
表 2 变形菌门主要属组成 (属,丰度大于1%)
Table 2. Major genus composition of Proteobacteria (genera, abundance >1%)
1号属名称 占比/% 2号属名称 占比/% Denitratisoma (DNB) 4.46 Thermomonas (DNB) 11.44 ANP-Rhizobium 6.91 Denitratisoma (DNB) 7.31 Pseudomonas (DNB) 8.66 ANP-Rhizobium 3.16 Rhodocyclaceae_uncultured (DNB) 8.01 Burkholderiaceae_unclassified 3.34 Burkholderiaceae_unclassified 3.95 Xanthobacteraceae_unclassified (DNB) 3.71 Xanthobacteraceae_unclassified (DNB) 3.25 Sphingobium 2.31 Rhodocyclaceae_unclassified (DNB) 4.01 Pleomorphomonas 1.27 Desulfovibrio (DNB) 3.61 Afipia 1.16 Pleomorphomonas 1.57 Devosia (DNB) 1.47 Burkholderiaceae_uncultured 1.63 — — Afipia 1.07 — — Rhizobiaceae_unclassified (DNB) 1.32 — — Gammaproteobacteria_unclassified (DNB) 1.40 — — Betaproteobacteriales_unclassified (DNB) 1.03 — — Desulforhabdus 1.05 — — 合计 51.93 合计 35.17 注:DNB为反硝化菌(denitrifying bacteria)。
下载: 导出CSV
-
[1] 赵远哲, 杨永哲, 王海燕, 等. 新型填料A/O生物滤池处理低碳氮比农村污水脱氮[J]. 环境科学, 2020, 41(5): 2329-2338. doi: 10.13227/j.hjkx.201910118 [2] ASHOK V, HAIT S. Remediation of nitrate-contaminated water by solid-phase denitrification process: A review[J]. Environmental Science and Pollution Research, 2015, 22(11): 8075-8093. doi: 10.1007/s11356-015-4334-9 [3] SÁNCHEZ M P, SULBARÁN-RANGEL B C, TEJEDA A, et al. Evaluation of three lignocellulosic wastes as a source of biodegradable carbon for denitrification in treatment wetlands[J]. International Journal of Environmental Science and Technology, 2020, 17(12): 4679-4692. doi: 10.1007/s13762-020-02815-9 [4] HAO S Z, SHAN S X, HUI W Y, et al. Intensified heterotrophic denitrification in constructed wetlands using four solid carbon sources: Denitrification efficiency and bacterial community structure[J]. Bioresource Technology, 2018, 267: 416-425. doi: 10.1016/j.biortech.2018.07.029 [5] YU L, YAN G K, WANG H Y, et al. Release mechanism, secondary pollutants and denitrification performance comparison of six kinds of agricultural wastes as solid carbon sources for nitrate removal[J]. International Journal of Environmental Research and Public Health, 2021, 18(3): 1232. doi: 10.3390/ijerph18031232 [6] GUAN X X, JI G X, XU S Y, et al. Selection of agricultural straws as sustained-release carbon source for denitrification in a drawer-type biological filter[J]. Water, Air, & Soil Pollution, 2019, 230(18): 1-11. [7] SUN H, ZHOU Q, ZHAO L, et al. Enhanced simultaneous removal of nitrate and phosphate using novel solid carbon source/zero-valent iron composite[J]. Journal of Cleaner Production, 2020, 289: 125757. [8] YANG Z, YANG L, WEI C, et al. Enhanced nitrogen removal using solid carbon source in constructed wetland with limited aeration[J]. Bioresource Technology, 2018, 248: 98-103. doi: 10.1016/j.biortech.2017.07.188 [9] LI Y Y, WANG S, LI Y, et al. Corn straw as a solid carbon source for the treatment of agricultural drainage water in horizontal subsurface flow constructed wetlands[J]. Water, 2018, 10(4): 511. doi: 10.3390/w10040511 [10] SHEN Z Q, ZHOU Y X, HU J, et al. Denitrification performance and microbial diversity in a packed-bed bioreactor using biodegradable polymer as carbon source and biofilm support[J]. Journal of Hazardous Materials, 2013, 250-251: 431-438. doi: 10.1016/j.jhazmat.2013.02.026 [11] ZHANG F F, MA C J, HUANG X F, et al. Research progress in solid carbon source-based denitrification technologies for different target water bodies[J]. Science of the Total Environment, 2021, 782: 1466669. [12] ZHOU B B, DUAN J J, XUE L H, et al. Effect of plant-based carbon source supplements on denitrification of synthetic wastewater: Focus on the microbiology[J]. Environmental Science and Pollution Research International, 2019, 26(24): 24683-24694. doi: 10.1007/s11356-019-05454-x [13] 李玉敏, 冯鹏飞. 基于第九次全国森林资源清查的中国竹资源分析[J]. 世界竹藤通讯, 2019, 17(6): 45-48. [14] 郑龙, 吴义强, 左迎峰. 竹剩余物资源化利用研究现状与展望[J]. 世界林业研究, 2021, 34(3): 82-88. [15] 江泽慧, 王戈, 费本华, 等. 竹木复合材料的研究及发展[J]. 林业科学研究, 2002(6): 712-718. doi: 10.3321/j.issn:1001-1498.2002.06.013 [16] 彭洋洋. 预处理对竹子组织结构和纤维的影响及高得率纳米纤维素制备的研究[D]. 广州: 华南理工大学, 2019. [17] WANG J, CHU L. Biological nitrate removal from water and wastewater by solid-phase denitrification process[J]. Biotechnology Advances, 2016, 34(6): 1103-1112. doi: 10.1016/j.biotechadv.2016.07.001 [18] HARTZ T K, SMITH R, CAHN M D, et al. Wood chip denitrification bioreactors can reduce nitrate in tile drainage[J]. California Agriculture, 2017, 71: 41-47. doi: 10.3733/ca.2017a0007 [19] XU Y, WANG C, HOU J, et al. Application of zero valent iron coupling with biological process for wastewater treatment: A review[J]. Reviews in Environmental Science and Biotechnology, 2017, 16: 667-693. doi: 10.1007/s11157-017-9445-y [20] FU F, DIONYSIOU D D, LIU H. The use of zero-valent iron for groundwater remediation and wastewater treatment: A review[J]. Journal of Hazardous Materials, 2014, 267: 194-205. doi: 10.1016/j.jhazmat.2013.12.062 [21] YOU G, WANG P, HOU J, et al. The use of zero-valent iron (ZVI)-microbe technology for wastewater treatment with special attention to the factors influencing performance: A critical review[J]. Critical Reviews in Environmental Science and Technology, 2017, 47(10): 877-907. doi: 10.1080/10643389.2017.1334457 [22] 杨燕, 朱静平. 添加零价铁的反硝化系统中发生的主要反应[J]. 工业水处理, 2021, 41(3): 77-82. [23] WANG C, XU Y, HOU J, et al. Zero valent iron supported biological denitrification for farmland drainage treatments with low organic carbon: Performance and potential mechanisms[J]. Science of the Total Environment, 2019, 689: 1044-1053. doi: 10.1016/j.scitotenv.2019.06.488 [24] 于妍, 刘宁, 廖祖刚, 等. 铁型反硝化脱氮技术研究进展[J]. 中国环境科学, 2022, 42(1): 83-91. doi: 10.3969/j.issn.1000-6923.2022.01.010 [25] OU C, SHEN J, ZHANG S, et al. Coupling of iron shavings into the anaerobic system for enhanced 2, 4-dinitroanisole reduction in wastewater[J]. Water Research, 2016, 101: 457-466. doi: 10.1016/j.watres.2016.06.002 [26] 童海航, 石德智, 刘嘉宇, 等. 金属纳米颗粒辅助木质纤维素暗发酵生物制氢的研究进展[J]. 化工学报, 2022, 73(4): 1417-1435. doi: 10.11949/0438-1157.20211412 [27] 颜涌捷, 任铮伟. 纤维素连续催化水解研究[J]. 太阳能学报, 1999, 20(1): 56-59. doi: 10.3321/j.issn:0254-0096.1999.01.010 [28] 杨清培, 欧阳明, 杨光耀, 等. 竹子生态化学计量学研究: 从生物学基础到竹林培育学应用[J]. 植物生态学报, 2016, 40(3): 264-278. doi: 10.17521/cjpe.2015.0298 [29] 韦琦, 罗方周, 徐相龙, 等. A2/O工艺处理低温低碳氮比生活污水的脱氮效率及反应动力学[J]. 环境工程学报, 2021, 15(4): 1367-1376. doi: 10.12030/j.cjee.202010126 [30] 赵文莉, 郝瑞霞, 王润众, 等. 复合碳源填料反硝化脱氮及微生物群落特性[J]. 中国环境科学, 2015, 35(10): 3003-3009. doi: 10.3969/j.issn.1000-6923.2015.10.017 [31] 许兵, 张旭, 刘佳, 等. 植物碳源对人工湿地脱氮过程的影响[J]. 工业水处理, 2021, 41(12): 89-94. doi: 10.19965/j.cnki.iwt.2021-0201 [32] ZHENG Y, CAO T, ZHANG Y, et al. Characterization of dissolved organic matter and carbon release from wetland plants for enhanced nitrogen removal in constructed wetlands for low C-N wastewater treatment[J]. Chemosphere, 2021, 273: 129630. doi: 10.1016/j.chemosphere.2021.129630 [33] 陈广林, 胡荣庭, 祖凌鑫, 等. 木质碳源释碳能力的优化及其对地下水生物反硝化的强化效果[J]. 环境工程学报, 2022, 16(1): 154-163. doi: 10.12030/j.cjee.202110085 [34] 李卫芬, 郑佳佳, 张小平, 等. 反硝化酶及其环境影响因子的研究进展[J]. 水生生物学报, 2014, 38(1): 166-170. [35] 赵文莉, 郝瑞霞, 李斌, 等. 预处理方法对玉米芯作为反硝化固体碳源的影响[J]. 环境科学, 2014, 35(3): 987-994. doi: 10.13227/j.hjkx.2014.03.024 [36] 董晓莹, 彭党聪. 不同碳氮比下污水反硝化过程中亚硝氮积累的特性研究[J]. 环境科学学报, 2017, 37(9): 3349-3355. [37] GE S, PENG Y, WANG S, et al. Nitrite accumulation under constant temperature in anoxic denitrification process: The effects of carbon sources and COD/NO3-N[J]. Bioresource Technology, 2012, 114: 137-143. doi: 10.1016/j.biortech.2012.03.016 [38] 钟胜强, 杨扬, 陶然, 等. 5种植物材料的水解释碳性能及反硝化效率[J]. 环境工程学报, 2014, 8(5): 1817-1824. [39] 张恒亮, 朱铁群, 王海燕, 等. 芦苇碳源投加量对表面流人工湿地中试系统强化脱氮启动的影响[J]. 环境工程技术学报, 2017, 7(3): 332-339. doi: 10.3969/j.issn.1674-991X.2017.03.047 [40] 彭锦玉, 张克峰, 王全勇, 等. 以4种天然植物材料为碳源的固相反硝化研究[J]. 工业水处理, 2021, 41(10): 1-9. [41] 万雨轩, 王鑫. 废水处理中异化硝酸盐还原为铵的研究进展[J]. 土木与环境工程学报, 2021, 43(6): 134-144. [42] ETIQUE M, JORAND F P, ZEGEYE A, et al. Abiotic process for Fe(II) oxidation and green rust mineralization driven by a heterotrophic nitrate reducing bacteria (Klebsiella mobilis)[J]. Environmental Science & Technology, 2014, 48(7): 3742-3751. [43] 张雅君, 吕静静, 孙丽华, 等. 不同硝酸盐浓度对再生水管网腐蚀状况的影响[J]. 腐蚀科学与防护技术, 2018, 30(3): 259-265. doi: 10.11903/1002.6495.2017.229 [44] ZHU L, GAO K, JIN J, et al. Analysis of ZVI corrosion products and their functions in the combined ZVI and anaerobic sludge system[J]. Environmental Science and Pollution Research International, 2014, 21(22): 12747-12756. doi: 10.1007/s11356-014-3215-y [45] 李园怡, 李杰. Fe0-生物铁法中铁的微生物腐蚀机理研究进展[J]. 绿色科技, 2019(8): 50-52. [46] SUN H, SHI B, YANG F, et al. Effects of sulfate on heavy metal release from iron corrosion scales in drinking water distribution system[J]. Water Research, 2017, 114: 69-77. doi: 10.1016/j.watres.2017.02.021 [47] WANG R, YANG C, WANG W Y, et al. An efficient way to achieve stable and high-rate ferrous ion-dependent nitrate removal (FeNiR): Batch sludge replacement[J]. Science of the Total Environment, 2020, 738: 139396. doi: 10.1016/j.scitotenv.2020.139396 [48] 李伟, 李贤军, 王望, 等. 木材腐朽机理研究现状及展望[J]. 世界林业研究, 2022, 35(2): 64-69. [49] 方晶晶, 马传明, 刘存富. 反硝化细菌研究进展[J]. 环境科学与技术, 2010, 33(S1): 206-210. [50] 严子春, 唐瑞祥, 吴大冰. 有机物对厌氧氨氧化生物膜反应器脱氮效能及微生物群落的影响[J]. 环境科学学报, 2021, 41(4): 1303-1308. [51] 刘亚妮, 朱宏伟, 黄荣新, 等. 曝气生态滤池中微生物群落组成及物种多样性[J]. 中国环境科学, 2020, 40(3): 1075-1080. doi: 10.3969/j.issn.1000-6923.2020.03.017 [52] 万琼, 吴仪, 王信, 等. BAF中不同高度海绵铁填料表面物种多样性分析[J]. 环境工程技术学报, 2018, 8(2): 161-168. doi: 10.3969/j.issn.1674-991X.2018.02.022 [53] 彭帅, 陈晓国, 李晓光, 等. 不同填埋龄垃圾腐殖土中细菌群落结构特征[J]. 环境工程技术学报, 2021, 11(5): 879-887. doi: 10.12153/j.issn.1674-991X.20210147 [54] ZHANG M, DARAZ U, SUN Q, et al. Denitrifier abundance and community composition linked to denitrification potential in river sediments[J]. Environmental Science and Pollution Research International, 2021, 28(37): 51928-51939. doi: 10.1007/s11356-021-14348-w [55] 张兰河, 左正艳, 王旭明. 固相反硝化系统中微生物群落结构的研究进展[J]. 生物技术通报, 2015, 31(1): 39-45. [56] 杨腾腾, 周宏, 王霞, 等. 微生物降解纤维素的新机制[J]. 微生物学通报, 2015, 42(5): 928-935. [57] 赵彬, 丁雪松, 吴丹青, 等. 高负荷条件下好氧颗粒污泥同步脱氮除碳特性及微生物群落结构分析[J]. 环境工程学报, 2020, 14(2): 295-304. doi: 10.12030/j.cjee.201903202 [58] EICHORST S A, VARANASI P, STAVILA V, et al. Community dynamics of cellulose-adapted thermophilic bacterial consortia[J]. Environmental Microbiology, 2013, 15(9): 2573-2587. doi: 10.1111/1462-2920.12159 [59] 林芳妃. 活性污泥法去除城市径流雨水有机污染物研究[D]. 兰州: 兰州理工大学, 2020. [60] HU R T, ZHENG X, ZHENG T Y, et al. Effects of carbon availability in a woody carbon source on its nitrate removal behavior in solid-phase denitrification[J]. Journal of Environmental Management, 2019, 246: 832-839. [61] 严子春, 史登峰, NOSAKHARE I. 富铁填料强化A/O-曝气生物滤池工艺的脱氮除磷效果[J]. 环境污染与防治, 2017, 39(11): 1186-1188. [62] XU D, YING S, WANG Y H, et al. A novel SAD process: Match of anammox and denitrification[J]. Water Research, 2021, 193: 116874. doi: 10.1016/j.watres.2021.116874 [63] OSVALDO D F, GUILLERMO Q, REBECA P, et al. Simultaneous biological nitrous oxide abatement and wastewater treatment in a denitrifying off-gas bioscrubber[J]. Chemical Engineering Journal, 2016, 288: 28-37. doi: 10.1016/j.cej.2015.11.088 [64] 蔺凌云, 尹文林, 潘晓艺, 等. 自然微生物挂膜处理水产养殖废水的效果及微生物群落分析[J]. 水生生物学报, 2017, 41(6): 1327-1335. doi: 10.7541/2017.164 [65] 苑宏英, 王雪, 李原玲, 等. 碳氮比对低温投加介体生物反硝化脱氮的影响[J]. 环境工程学报, 2020, 14(1): 60-67. doi: 10.12030/j.cjee.201902025 [66] CHU L B, WANG J L. Denitrification performance and biofilmcharacteristics using biodegradable polymers PCL ascarriers and carbon source[J]. Chemosphere, 2013, 91: 1310-1316. doi: 10.1016/j.chemosphere.2013.02.064 [67] WANG X Y, LI Y J, CIAMPITTI I A, et al. Response of soil denitrification potential and community composition of denitrifying bacterial to different rates of straw return in north-central China[J]. Applied Soil Ecology, 2022, 170: 104312. doi: 10.1016/j.apsoil.2021.104312 [68] LV B, ZHANG D, CHEN Q, et al. Effects of earthworms on nitrogen transformation and the correspond genes (amoA and nirS) in vermicomposting of sewage sludge and rice straw[J]. Bioresource Technology, 2019, 287: 121428. doi: 10.1016/j.biortech.2019.121428 [69] KANG Z, ZOU J, HUANG Y, et al. Tuber melanosporum shapes nirS-type denitrifying and ammonia-oxidizing bacterial communities in Carya illinoinensis ectomycorrhizosphere soils[J]. PeerJ, 2020, 8: e9457. doi: 10.7717/peerj.9457 [70] 王雨阳, 李茹莹. 同步脱氮除磷复配菌剂对河流水质净化效果研究[J]. 环境科学学报, 2022, 42(5): 187-194. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 5551
- HTML全文浏览数: 5551
- PDF下载数: 50
- 施引文献: 0
