




-
《城镇污水处理厂污染物排放标准》(GB18918-2002)规定了TN的一级A排放标准限值为15 mg·L−1 ,
${\rm{NH}}_4^ +$ -N的一级A排放标准限值为5 mg·L−1 [1]。2016年以来,各地相继发布了城镇污水处理厂的污染物地方排放标准,其中的出水氮指标愈加严格[2]。例如,北京市的《城镇污水处理厂水污染物排放标准》(DB11/890-2012)中TN和${\rm{NH}}_4^ +$ -N的一级A标准限值分别调至10 mg·L−1 和1.0 (1.5) mg·L−1 [3] 。更严格的排放标准也给污水处理厂的生物脱氮工艺提出了更高要求。以前置缺氧反硝化(缺氧-好氧,anoxic-oxic,AO)为代表的生物脱氮工艺是主流的生活污水脱氮技术[4-6]。在相关的工程应用中,一般采取增大内回流比的方式来提高工艺的脱氮效率。然而,内回流液来自于曝气区,其溶解氧含量较高,导致缺氧区不能保持理想的缺氧状态,从而限制了脱氮效率的提升,且很难实现深度脱氮。若在好氧段的后端增加缺氧段形成后置缺氧段,即构成了缺氧-好氧-缺氧(anoxic-oxic-anoxic,AOA)工艺,便可充分利用微生物胞内糖原或聚羟基脂肪酸酯驱动反硝化来进行脱氮[7-8]。该工艺不仅脱氮效果好,还可利用内碳源以节省费用,并在一定程度上减少剩余污泥的产量、降低污泥处置费用,是一种有发展潜力的技术[4]。然而,由于微生物自身存储碳源不足,在长时间的好氧反应后,细胞内的碳源大多已被氧化,此时细胞处于饥饿状态,导致后置缺氧区的内源反硝化效率较低,故仍需采用提高污泥浓度、促进短程硝化反硝化等措施以提高后置缺氧区的反硝化效率[9-13]。
本研究在中试规模装置的后置缺氧区添加生物填料,以提高系统内污泥浓度,并通过添加羟胺调控不同硝化细菌以实现短程硝化反硝化,再优化外回流比以提升整体脱氮,以期为污水深度脱氮工艺的进一步工程化应用提供参考。
-
实验用水取自北京市某再生水厂曝气沉砂池,具有低C/N的特点。该水样平均COD为172 mg·L−1,平均BOD5为95 mg·L−1,平均TP为3.2 mg·L−1,平均碱度为725 mg·L−1,平均
${\rm{NH}}_4^ + $ -N为42.8 mg·L−1,平均${\rm{NO}}_{\rm{2}}^{\rm{ - }} $ -N和${\rm{NO}}_{\rm{3}}^{\rm{ - }} $ -N均小于1.0 mg·L−1。装置中接种的污泥取自该厂多级AO生化池,其MLSS为5 610 mg·L−1、MLVSS为4 160 mg·L−1。 -
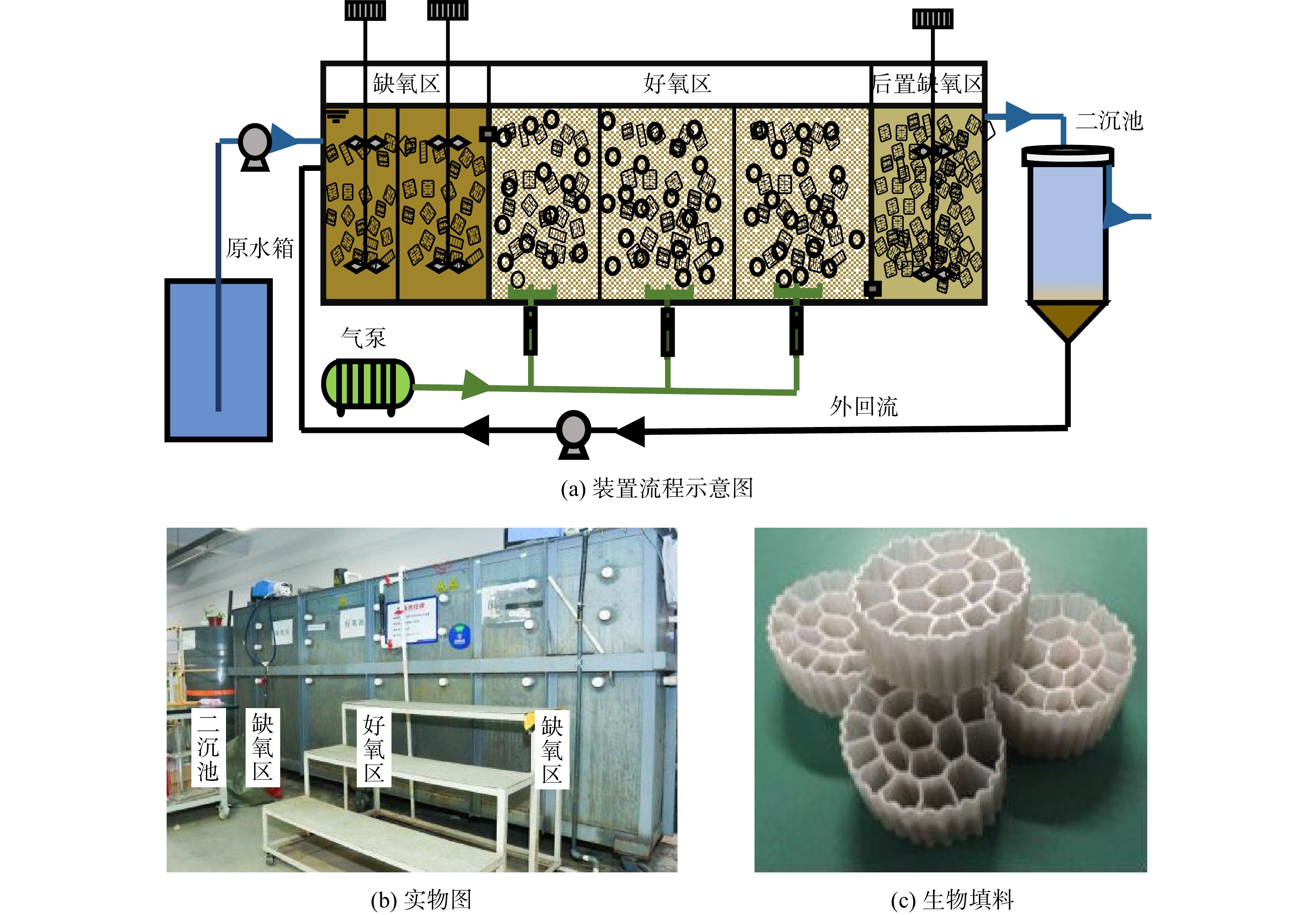
中试装置含进水箱、进水管路、提升泵、反应器、气泵、二沉池、回流管路及排水管路等部件,其流程见图1。进水从进水箱通过提升泵输送至反应器。反应器总容积3 m3,划分为6格。其中,缺氧区分为2格(A1、A2),好氧区分为3格(O1、O2、O3),而后置缺氧区仅有1格(A3)。好氧区通过气泵进行供气。3个区域内均填充有聚乙烯生物填料,填充比例分别为10% (缺氧区)、10% (好氧区)、50% (后置缺氧区)。反应器出水进入二沉池后进行沉淀分离,二沉池的容积为0.785 m3,回流管路设置于底部锥形漏斗。部分沉淀污泥经过外回流管路通过提升泵回到缺氧区,出水及剩余污泥则外排。
-
该中试装置的设计处理水量为4.41 m3·d−1,总水力停留时间(HRT)为16.2 h。缺氧区、好氧区和后置缺氧区的HRT分别为3.7、8.8和3.7 h。在运行过程中,通过控制排泥速度将系统中污泥质量浓度控制在3 500~5 500 mg·L−1。在运行末期,3个区域内生物膜对应的污泥质量浓度均低于1 g·L−1。初始污泥外回流比设置为100%。好氧区溶解氧(DO)维持在1~4 mg·L−1。在装置运行过程中,投加至好氧区的羟胺质量浓度为5 mg·L−1。
在挂膜结束后,将运行过程分为3个阶段,相应的运行时间及控制条件如表1所示。阶段1为对照阶段,用以考察该工艺在常规运行条件下的脱氮效果;阶段2为强化脱氮阶段,即在阶段1基础上,通过在好氧区投加羟胺以积累
${\rm{NO}}_2^{\rm{ - }} $ -N进而促进短程反硝化,用以考察系统的强化脱氮效果;阶段3为深度强化脱氮阶段,即在阶段2的基础上,通过提高外回流比,用以进一步考察系统的强化脱氮潜力。 -
每天进行水质监测,主要监测指标有
${\rm{NH}}_4^ + $ -N、TN、${\rm{NO}}_{\rm{2}}^{\rm{ - }} $ -N和${\rm{NO}}_{\rm{3}}^{\rm{ - }}$ -N。进出水样品分别来自进水箱和二沉池,其余各处理段水样采自各池体的出水口。将水样采用0.45 µm中速滤纸过滤后方可进行分析。所有指标均采用标准方法测定。温度、DO采用WTW340i测定。微生物样品采自接种污泥,以及缺氧区、好氧区和后置缺氧区的填料。利用十二烷基苯磺酸钠(SDS)法获得菌体DNA,用琼脂糖凝胶电泳检测DNA纯度和浓度。以稀释的基因组DNA为模板,使用带 Barcode 的特异引物和高效保真酶进行PCR。PCR产物用2%琼脂糖凝胶电泳检测,纯化后在48 RXNS文库构建,在Ion S5TMXL进行上机测序。序列在Illumina HiSeq 4000平台上进行分类分析。
-
为充分了解短程硝化反硝化的程度,采用式(1)计算好氧区出水中的亚硝酸盐氮积累率( nitrite accumulation rates,NAR )。
式中:C(
${\rm{NO}}_2^{\rm{ - }} $ -N)和C(${\rm{NO}}_3^{\rm{ - }} $ -N)分别代表好氧区出水中${\rm{NO}}_2^{\rm{ - }} $ -N和${\rm{NO}}_3^{\rm{ - }} $ -N的质量浓度,mg·L−1。 -
在挂膜驯化完成后,生物膜耦合AOA系统对
${\rm{NH}}_4^ + $ -N和TN的去除效果如图2所示。在3个不同运行阶段,系统均可稳定去除${\rm{NH}}_4^ + $ -N。在进水${\rm{NH}}_4^ + $ -N平均质量浓度分别为38.5、34.3 和38.5 mg·L−1 的条件下,出水${\rm{NH}}_4^ + $ -N质量浓度均可低于1.0 mg·L−1 ,其平均去除率分别为97.67%、98.40%和97.41%。结果表明,羟胺的投加基本不会影响系统对${\rm{NH}}_4^ + $ -N的去除。在此基础上增大外回流比后,系统对TN的去除效率明显提升。在对照阶段(阶段1),系统进水TN保持在约39.4 mg·L−1,出水平均TN为20.7 mg·L−1,平均去除率仅为47.1%。而在好氧区投加羟胺之后(阶段2),进水TN虽稍有降低(均值为35.3 mg·L−1),但其平均值降至16.0 mg·L−1,平均TN去除率则增至54.7%。当外回流比调整为140% (阶段3)时,系统的进水平均TN为39.2 mg·L−1,而出水平均TN却降至11.3 mg·L−1,平均去除率增至71.3%。在阶段3的前期,因疫情导致系统重启动的影响,使得系统出水平均TN有所升高,但到后期,出水TN仍稳定低于10.0 mg·L−1。该中试装置所在的北京市某再生水厂的处理工艺包含多级AO、双层矩形平流沉淀池、 超滤深度处理等工艺段,其出水TN低于15 mg·L−1,可满足北京市地方标准《城镇污水处理厂水污染物排放标准》( DB 11 /890—2012) B级标准。而本装置的出水TN持续低于10 mg·L−1,说明中试研究取得了较好效果。
-
由于系统对
${\rm{NH}}_4^ + $ -N的去除主要发生在好氧区内,且3个阶段的去除效率高度一致,故本节仅分析TN的去除情况。计算3个阶段内各典型周期的缺氧、好氧和后置缺氧区脱氮贡献情况,结果如图3所示。在阶段1,TN的去除集中发生在缺氧区A1池 (33%)和后置缺氧区A3池(12%);在阶段2,投加羟胺后TN的去除集中发生在缺氧区(36%)、后置缺氧区(9%)和二沉池(8%);在阶段3,随着外回流比的增大,在系统的3个区域中均出现了TN的去除现象。其中,缺氧区、好氧区、后置缺氧区的TN去除贡献率分别为39%、13%和34%。
在阶段1,TN的去除仅发生在缺氧区A1池,说明A2池已无可利用外碳源,故推测A1池中已出现碳源不足,后置缺氧区对TN的去除为内源反硝化过程的结果。在阶段2投加羟胺后,缺氧区与后置缺氧区的TN去除效率与阶段1接近,而在阶段2的二沉池中出现了TN被去除的现象,说明在二沉池中污泥发生了内源反硝化。在阶段3,缺氧区TN去除率较阶段1提升了6%,后置缺氧区较阶段1提高了22%。不仅如此,好氧区也表现出对TN的去除作用,且去除率超过10%。该结果表明在好氧区内存在同步硝化反硝化过程。综上所述,在于好氧区内投加羟胺的条件下,生物膜耦合AOA系统对TN的去除效率得以大幅提升,且后置缺氧区的去除贡献最大。
-
监测了好氧区的出水指标
${\rm{NO}}_3^{\rm{ - }} $ -N及${\rm{NO}}_2^{\rm{ - }} $ -N,并根据数据计算了亚硝酸盐氮积累率,结果如图4所示。在阶段1,好氧区的出水中未检测到${\rm{NO}}_2^{\rm{ - }} $ -N,说明系统在此阶段全程进行硝化反应;在阶段2中投加羟胺后的1周内,NAR一直较低(<5%),但从第2周开始,NAR迅速上升,并在5 d内达到47.4%;在阶段3时继续投加羟胺、增大外回流比的条件下,NAR持续升高,最高达到70%。受新冠疫情影响,该装置停运了一段时间。系统重新启动后,受新加污泥的影响,前期短程硝化进程被破坏。随着羟胺的持续投加,
${\rm{NO}}_2^{\rm{ - }} $ -N得以快速积累,最终在羟胺投加第13 d时积累率升至75%。之后,在进水TN保持稳定的条件下,${\rm{NO}}_2^{\rm{ - }} $ -N积累率继续上升并稳定保持在约90%,最高积累率为97%。在实验进行末期,进水TN出现较大波动,而出水${\rm{NO}}_3^{\rm{ - }} $ -N有所提升,但NAR仍保持在约70%。以上分析表明,通过在好氧区投加质量浓度为5 mg·L−1的羟胺可抑制NOB活性,使硝化停留在
${\rm{NO}}_2^{\rm{ - }} $ -N阶段,从而实现了短程硝化。该过程与已有研究[14-17]结果基本一致。XU等[18]发现:在投加羟胺的质量浓度为10 mg·L−1时,SBR中会出现${\rm{NO}}_2^{\rm{ - }} $ -N快速积累的现象,平均积累率达到了99.8%。通过FISH检测还证明,AOB的丰度远远高于NOB。HAO等[19]在完全硝化滤池中添加质量浓度(以氮元素的质量浓度计)为2.5 mg·L−1的羟胺,通过监测${\rm{NO}}_2^{\rm{ - }} $ -N的积累量,证明羟胺对NOB有抑制作用,且恢复完全硝化需要至少30 d,说明添加羟胺能促进短程硝化的实现。本研究与BRACKLOW等[20]和VOCKS等[10]的研究结果一致。在未添加外部碳源时,后置缺氧段微生物利用胞内糖原或PHA驱动反硝化脱氮,促进了TN的去除。还有研究表明[21],当内碳源被作为反硝化碳源时,
${\rm{NO}}_2^{\rm{ - }} $ -N的还原速率为${\rm{NO}}_3^{\rm{ - }} $ -N的1.6倍。这与本研究的结果基本一致,即在相同HRT条件下,系统在阶段3实现了稳定的${\rm{NO}}_2^{\rm{ - }} $ -N积累,使得在阶段3系统的TN去除率高于阶段1,且${\rm{NO}}_2^{\rm{ - }} $ -N积累得越多,TN去除率增得更大。综上所述,在阶段1,缺氧区及后置缺氧区对TN的去除路径为全程反硝化;在阶段2,由于刚开始投加羟胺,系统短程硝化效果尚未稳定,此时
${\rm{NO}}_{\rm{2}}^{\rm{ - }} $ -N与${\rm{NO}}_3^{\rm{ - }} $ -N共同存在,缺氧区和后置缺氧区对TN的去除路径为短程反硝化和全程反硝化;在阶段3,系统已进入稳定期,主要生成${\rm{NO}}_{\rm{2}}^{\rm{ - }} $ -N并伴有少量${\rm{NO}}_3^{\rm{ - }} $ -N,此时的缺氧区和后置缺氧区中TN去除路径主要为短程反硝化。由于短程反硝化比全程反硝化速度快、碳源消耗少,故在阶段3后置缺氧区的脱氮效果得到了大幅提升。又由于缺氧区内消耗的碳源量降低,使得未消耗的碳源进入了好氧区,促使同步硝化反硝化发生,故进一步提升了好氧区内的脱氮效果。 -
微生物的群落组成可在一定程度上反映环境条件。取实验末期(第140 天)各区域填料上生物膜中的微生物进行高通量测序,并与接种污泥中的微生物群落进行了对比。图5为各样品微生物在属水平上的构成。
3个区域中主要优势菌均包含Acinetobacter (不动杆菌属)。由于Acinetobacter属细菌既可参与硝化反应又可参与反硝化反应,故该菌属已发展成为脱氮系统的主要优势菌[22]。该菌在后置缺氧区内丰度最高,说明其可能是进行内源反硝化的主要微生物。另外,在后置缺氧区,反硝化菌Bacillus (芽孢杆菌属)和Thermomonas (嗜热单胞菌属)的富集现象说明该区域发生的反硝化可能还与这2种菌有关[23]。
Dok59为缺氧区的主要优势菌属,属于变形菌门,可参与异养反硝化,亦为厌氧氨氧化反应器的优势菌属之一[22-25]。Dok59在缺氧区中丰度最高,说明其既可利用碳源进行反硝化,又可参与厌氧氨氧化。厌氧氨氧化菌Candidatus Brocadia在接种污泥中占比几乎为0,而在实验末期缺氧区的填料生物膜中明显增加,占比为0.7%。故推测,可能是来水中的氨氮及回流带来的
${\rm{NO}}_{\rm{2}}^{\rm{ - }} $ -N为该菌的生长提供了必要基质。分析缺氧区进出水中的${\rm{NH}}_4^ + $ -N监测数据后发现,缺氧区存在对${\rm{NH}}_4^ + $ -N的去除现象,去除率最高为9.4%。因此,以上2种菌在缺氧区的富集共同促进了在低C/N进水条件下$ {\rm{NH}}_4^ + $ -N和TN的去除[26-27]。Caldilinea为好氧区的第2优势菌属。该菌可消耗有机物,并将
${\rm{NO}}_3^{\rm{ - }} $ -N和${\rm{NO}}_2^{\rm{ - }} $ -N作为电子受体,故在好氧区内实现同步硝化反硝化可能与该菌的存在有关[28]。Nitrospira (硝化螺菌属)可将
${\rm{NO}}_2^{\rm{ - }} $ -N氧化成${\rm{NO}}_3^{\rm{ - }} $ -N,是污水处理系统中常见的NOB[29]。Nitrospira在接种污泥里占比1.3%。经过140 d的运行,其在缺氧区、后置缺氧区和好氧区填料中占比分别降至0.1%、0.3%和0.2%,从而印证了该系统已实现了短程硝化反硝化。微生物群落结构分析结果表明,通过在中试规模AOA系统中添加羟胺可有效抑制NOB生长,实现较稳定的
${\rm{NO}}_2^{\rm{ - }} $ -N积累,并在强化后置缺氧区脱氮的同时提升了系统的脱氮效果。因此,羟胺可应用于短程硝化的实际工程中,本结果可为实现城市污水主流工艺的短程硝化反硝化深度脱氮提供参考。 -
实现短程硝化是生物膜耦合AOA工艺提升TN去除率的重要途径,而羟胺的投加是亦非常关键。由于运行成本是城镇污水处理厂选择药剂的重要指标,故通过对比筛选污水处理厂常用药剂,以确定最佳投放碳源种类。
表2为投加羟胺或其他常用碳源的费用对比。由于投加的羟胺质量浓度较低,在处理相同规模的污水时,每日投加量远低于乙酸钠和葡萄糖的投加量。尽管羟胺单价稍高,但处理单位体积污水增加成本仅为投加乙酸钠时的60%、投加葡萄糖时的80%,可降低运行成本。此外,羟胺仅需在好氧池投加,降低了操作的运维难度;而通过实现短程硝化,可降低系统的污泥排放量,从而省下部分污泥处理处置费用。
-
1) 通过投加羟胺和优化系统外回流比,实现了生物膜耦合AOA系统的强化深度脱氮,出水平均TN由20.7 mg·L−1降至11.3 mg·L−1,后期出水稳定低于10.0 mg·L−1。在羟胺投加质量浓度5 mg·L−1、外回流比140%的条件下,后置缺氧区的TN去除率提升了22%、缺氧区提升了6%、好氧区提升了12%。
2) 系统通过投加羟胺实现了好氧区短程硝化、缺氧及后置缺氧区短程反硝化。当系统稳定运行时,好氧区亚硝酸盐氮积累率维持在70%以上,最高可达97%。同时,由于后置缺氧区短程反硝化消耗碳源降低,提升了好氧区同步硝化反硝化与缺氧区内源反硝化的脱氮能力。
3) Acinetobacter为系统中的优势菌属,其可在好氧区进行硝化、在缺氧及后置缺氧区进行反硝化。Caldilinea为好氧区反硝化的承担者。Dok59为缺氧区主要反硝化菌,可联合Candidatus Brocadia进行厌氧氨氧化脱氮。Bacillus和Thermomonas在后置缺氧区的富集有助于该区域内的TN去除。羟胺的投加导致亚硝酸盐氧化菌Nitrospira生长受到明显抑制,从而实现了短程硝化反硝化。
致谢 感谢国家水技术创新社会化服务平台在装置运行、分析测试方面给予的支持。感谢杨南阳、徐汉文在装置搭建、运行和文章初稿撰写中给予的帮助。
基于生物膜耦合AOA的城镇生活污水深度脱氮工艺中试研究
A pilot-scale study of hybrid system of biofilm and anoxic-oxic-anoxic process for enhanced nitrogen removal of municipal wastewater
-
摘要: 针对缺氧-好氧-缺氧(AOA)工艺中后置缺氧区效率偏低的问题,通过耦合生物膜、投加羟胺及优化外回流比等方式,在不外加碳源的条件下开展了中试规模的低C/N污水深度脱氮实验,考察了后置缺氧区对强化脱氮的贡献,并分析了系统强化脱氮的实现途径。结果表明:在投加的羟胺质量浓度为5 mg·L−1、外回流比为140%的条件下,系统脱氮效率可提升40%;其中,后置缺氧区提升22%,后期出水TN稳定低于10.0 mg·L−1;脱氮途径由全程硝化反硝化转变为短程硝化反硝化,稳定期系统亚硝酸盐氮积累率达90%以上。微生物群落结构分析结果表明,Acinetobacter为优势菌属参与了系统硝化反硝化,优势菌属Caldilinea 和Dok59及明显富集菌属Candidatus Brocadia、Bacillus和Thermomonas均对脱氮有促进作用。以上结果可为该工艺的进一步工程应用提供参考。Abstract: In view of the low efficiency of post-anoxic zone in the process of anoxic-oxic-anoxic (AOA), a pilot-scale experiment of deep nitrogen removal from low C/N wastewater was carried out without adding carbon sources, by coupling biofilms, adding hydroxylamine and optimizing external reflux ratio. The contribution of post-anoxic zone to enhanced nitrogen removal was explored and ways for enhanced nitrogen removal were analyzed. The results showed that the nitrogen removal efficiency increased by 40% under the conditions of 5 mg·L-1 hydroxylamine and a reflux ratio at 140%. The efficiency of the post-anoxic zone increased by 22%, and the total nitrogen (TN) in the effluent was lower than 10 mg·L-1. The denitrification pathway changed to partial nitrification and denitrification from complete nitrification and denitrification. The nitrite accumulation rate reached more than 90% during the stable operation. The analysis of microbial community structure showed that Acinetobacter was the dominant bacterium involved in systematic nitrification and denitrification. The dominant bacteria Caldilinea and Dok59 promoted the nitrogen removal, as well as the obvious enrichment bacteria Candidatus Brocadia、 Bacillus and Thermomonas. These results can provide reference for further engineering application of the process.
-
Key words:
- biofilm /
- hydroxylamine /
- partial nitrification /
- endogenous denitrification /
- pilot study
-

-

图 3 在不同运行阶段各工艺段中的TN去除贡献
Figure 3. TN removal contributionin each process stage at different running stages

图 4 好氧区出水中
${\rm{NO}}_3^{\rm{ - }} $ -N、${\rm{NO}}_2^{\rm{ - }} $ -N的质量浓度及亚硝酸盐氮的积累率Figure 4. The mass concentrations of NO3--N and NO2--N and accumulation rate of NO2--N in effluent of oxic area
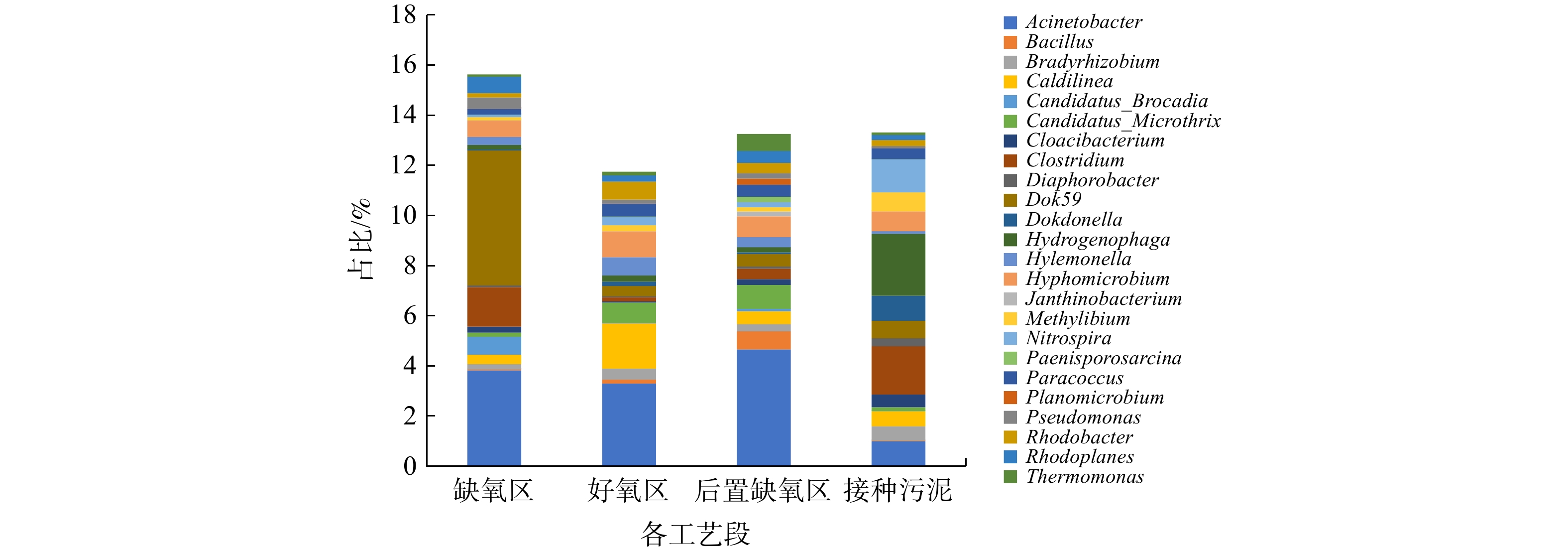
图 5 AOA系统各区段填料生物膜及接种污泥中的微生物群落结构对比
Figure 5. Comparison of microbial community structure of anoxic, oxic, post-anoxic and original sludge in AOA system
表 1 AOA系统的运行工况
Table 1. Operating conditions of AOA system
运行阶段 运行时长
/d羟胺质量浓度
/ (mg·L−1)外回流比
/%温度
/℃阶段1 0~37 0 100 18~20 阶段2 38~51 5 100 18~20 阶段3 52~145 5 140 18~25  下载: 导出CSV
下载: 导出CSV
表 2 投加羟胺与其他常用碳源的经济性对比分析
Table 2. Comparative analysis of economy for hydroxylamine and other common carbon sources dose
碳源
种类投加位置 投加的质量浓度
/(mg·L−1)试剂单价
/(元·kg−1)单位水量增加成本
/(元 ·104 m−3)羟胺 好氧池 5 30 1 086 乙酸钠 厌氧/缺氧池 36 (按C/N为4) 5 1 800 葡萄糖 厌氧/缺氧池 45 (按C/N为5) 3 1 350 注:假设处理水量为1.0 × 104 m3·d−1,出水平均TN由19 mg·L−1
降至10 mg·L−1。
下载: 导出CSV
-
[1] 国家环境保护总局. 城镇污水处理厂污染物排放标准 GB 18918-2002[S]. 北京: 中国标准出版社, 2003. [2] 陈秀成. 长三角地区污水处理厂排放标准解析及提标改造对策思考[J]. 给水排水, 2019, 45(11): 29-32. [3] 北京市环境保护局. 城镇污水处理厂水污染物排放标准DB11/890-2012[S/OL]. 2012. http://sthjj.beijing.gov.cn/bjhrb/resource/cms/article/bjhrb_810268/502337/2019122315153572044.pdf. [4] 王慰, 王淑莹, 张琼, 等. 后置缺氧UCT分段进水工艺处理低C/N城市污水[J]. 中国环境科学, 2016, 36(7): 1997-2005. doi: 10.3969/j.issn.1000-6923.2016.07.014 [5] 张勇, 王淑莹, 赵伟华, 等. 低温对中试AAO-BAF双污泥脱氮除磷系统的影响[J]. 中国环境科学, 2016, 36(1): 56-65. doi: 10.3969/j.issn.1000-6923.2016.01.010 [6] 彭永臻, 王建华, 陈永志. A2O-BAF联合工艺处理低碳氮比生活污水[J]. 北京工业大学学报, 2012, 38(4): 590-595. [7] COATS E R, MOCKOS A, LOGE F J. Post-anoxic denitrification driven by PHA and glycogen within enhanced biological phosphorus removal[J]. Bioresource Technology, 2011, 102(2): 1019-1027. doi: 10.1016/j.biortech.2010.09.104 [8] WINKLER M, COATS E R, BRINKMAN C K. Advancing post-anoxic denitrification for biological nutrient removal[J]. Water Research, 2011, 45(18): 6119-6130. doi: 10.1016/j.watres.2011.09.006 [9] MIELCAREK A, RODZIEWICZ J, JANCZUKOWICZ W, et al. The impact of biodegradable carbon sources on nutrients removal in post-denitrification biofilm reactors[J]. Science of the Total Environment, 2020, 720: 137377. doi: 10.1016/j.scitotenv.2020.137377 [10] VOCKS M, ADAM C, LESJEAN B, et al. Enhanced post-denitrification without addition of an external carbon source in membrane bioreactors[J]. Water Resource, 2005, 39(14): 3360-3368. DOI:10.1016/j.watres.2005.05.049. [11] 黄霞, 文剑平, 俞开昌, 等. 强化内源反硝化的膜-生物反应器脱氮除磷工艺及装置: 中国, 200810097427.2[P]. 2008-10-08. [12] 邓仁健, 张金松, 曲志军. 污泥浓度对双重后置反硝化工艺脱氮除磷的影响[J]. 环境科学研究, 2014, 27(7): 97-803. [13] 巩秀珍, 于德爽, 袁梦飞, 等. 后置短程反硝化AOA-SBR工艺实现低C/N城市污水的脱氮除磷[J]. 环境科学, 2019, 40(1): 360-368. [14] 马斌. 城市污水连续流短程硝化厌氧氨氧化脱氮工艺与技术[D]. 哈尔滨: 哈尔滨工业大学, 2012. [15] 张金铭, 王宇佳, 胡雪松. 强化厌氧氨氧化工艺的研究进展[J]. 辽宁化工, 2021, 50(6): 822-827. doi: 10.3969/j.issn.1004-0935.2021.06.020 [16] LI J, ZHANG Q, LI X, et al. Rapid start-up and stable maintenance of domestic wastewater nitritation through short-term hydroxylamine addition[J]. Bioresource Technology, 2019, 278: 468-472. doi: 10.1016/j.biortech.2019.01.056 [17] WANG Y, WANG H, ZHANG J, et al. Deciphering the evolution of the functional genes and microbial community of the combined partial nitritation-anammox process with nitrate build-up and its in situ restoration[J]. RSC Advances, 2016, 6(113): 111702-111712. doi: 10.1039/C6RA23865C [18] XU G, XU X, YANG F, et al. Partial nitrification adjusted by hydroxylamine in aerobic granules under high DO and ambient temperature and subsequent anammox for low C/N wastewater treatment[J]. Chemical Engineering Journal, 2012, 213: 338-345. doi: 10.1016/j.cej.2012.10.014 [19] HAO O J, CHEN J M. Factors Affecting nitrite buildup in submerged filter system[J]. Journal of Environmental Engineering, 1994, 120(5): 1298-1307. doi: 10.1061/(ASCE)0733-9372(1994)120:5(1298) [20] BRACKLOW U, DREWS A, GNIRSS R, et al. Influence of sludge loadings and types of substrates on nutrients removal in MBRs[J]. Desalination, 2010, 250(2): 734-739. doi: 10.1016/j.desal.2008.11.032 [21] 王少坡, 彭永臻, 于德爽, 等. 常温短程内源反硝化生物脱氮[J]. 北京工业大学学报, 2005, 31(3): 298-302. [22] 刘泽英. Acinetobacter sp. Y1的氨氮去除性能及其关键酶的研究[D]. 太原: 太原理工大学. 2016. [23] KUYPERS M M M, MARCHANT H K, KARTAL B. The microbial nitrogen-cycling network[J]. Nature Reviews Microbiology, 2018, 16(5): 263-276. doi: 10.1038/nrmicro.2018.9 [24] 许明熠. 耦合厌氧氨氧化技术的生物电化学系统同步脱氮产电机理研究及功能菌群落分析[D]. 广州: 华南理工大学. 2017 [25] 陈重军, 张海芹, 汪瑶琪, 等. 基于高通量测序的ABR厌氧氨氧化反应器各隔室细菌群落特征分析[J]. 环境科学, 2016, 37(7): 2652-2658. [26] HU A, ZHENG P, MAHMOOD Q, et al. Characteristics of nitrogenous substrate conversion by anammox enrichment[J]. Bioresource Technology, 2011, 102(2): 536-542. doi: 10.1016/j.biortech.2010.07.015 [27] ZEKKER I, KROON K, RIKMANN E, et al. Accelerating effect of hydroxylamine and hydrazine on nitrogen removal rate in moving bed biofilm reactor[J]. Biodegradation, 2012, 23(5): 739-749. doi: 10.1007/s10532-012-9549-6 [28] 邢金良, 张岩, 陈昌明, 等. CEM-UF组合膜-硝化/反硝化系统处理低C/N废水及种群结构分析[J]. 环境科学, 2018, 39(3): 1342-1349. [29] STACKEBRANDT E, GOEBEL B M. Taxonomic note a place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology[J]. Internation Journal of Systermatic Bacteriology, 1994, 44(4): 846-849. -
 点击查看大图
点击查看大图
计量
- 文章访问数: 5929
- HTML全文浏览数: 5929
- PDF下载数: 211
- 施引文献: 0













































