

 下载:
下载:




-
非点源污染日益成为流域水环境质量提升的主要限制因素[1-3]。非点源污染导致大量氮磷等营养元素进入水体,从而加速水体富营养化进程[4]。已有的研究表明, 全球至少有30%~50%地表水受到非点源污染影响[5-6]。我国非点源水污染问题也很严重[7-8],氮、磷污染物中分别有46.52%和67.21%来自于农业源[9]。
受人类生产和生活活动干扰剧烈的小流域往往同时存在农业非点源和村镇非点源[10-11]。当前对两类非点源污染过程的研究主要考虑降雨强度和下垫面类型对氮、磷等污染物输出的影响[12-16],以及非点源污染与颗粒物流失的关系[17-18]。然而,小流域非点源综合治理需要全面了解各下垫面上污染物的空间分布、污染过程、组分形态及负荷等特征,以便在非点源污染面广量大的特征下对重点下垫面或土地利用类型采取针对性管控措施。两类非点源污染在空间分布、污染过程、污染类型及负荷大小上存在明显差异,但目前大多数研究只关注单一下垫面或下垫面的某类性质,鲜见在同一个小流域内同步对村镇非点源和农业非点源展开研究,致使小流域非点源污染管控措施的针对性不强。
我国南方红壤丘陵地区小流域人多地少,人口与环境间的矛盾比较尖锐。流域内毁林垦荒和过度施肥现象严重,农业非点源对小流域水体污染贡献突出[19-20]。此外,该地区人口密集、村镇分布集中,小流域内的村镇非点源污染同样不可忽视[21-22]。因此,在我国南方红壤丘陵地区小流域开展农业非点源和村镇非点源研究将对水环境改善具有重要意义。目前,在该地区的非点源污染有关研究多集中在水土流失和土壤侵蚀等方面[23-24],对比观测小流域内村镇非点源和农业非点源过程从而揭示两者污染特征差异的研究还较少。
本研究以浙江省中部红壤丘陵区域的岩下小流域为例,在2020年4—9月,对流域内林地、农业种植用地和村镇道路等主要下垫面的径流出口进行监测,揭示不同下垫面非点源磷污染物的输出过程与规律,多角度比较不同下垫面非点源磷污染物的输出特征,并讨论磷输出过程与降雨过程、流量和颗粒物流失之间的关系,以期为区域非点源污染的源头控制和水环境综合治理提供参考。
全文HTML
-
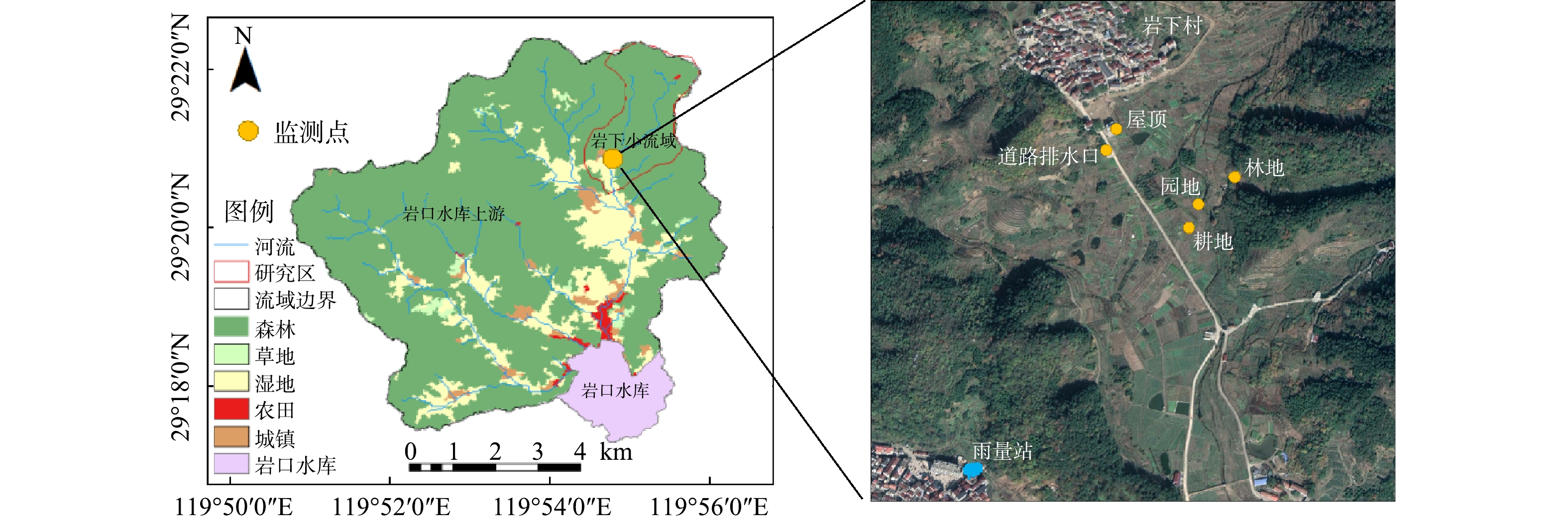
岩下小流域位于钱塘江流域义乌江支流航慈溪上游岩口水库流域内,地形以低山丘陵为主,岗地平地均有分布,三面环山,集水面积约为4.52 km2,属于典型的南方丘陵山地流域。当地日照充足,雨量丰沛,多年平均降雨1 400 mm,且降水量时空分布不均,主要集中在4 月中旬至8月中旬。台风暴雨和梅雨是形成洪水的主要因素。研究区内的土壤主要是红壤和改良的水稻土。红壤肥力差,易发生水土流失。研究区内农业用地分布集中,主要包括园地和耕地,分别占总面积的6.8%和1.9%。其中,园地以桃树种植为主,耕地以蔬菜种植为主;林地面积约占总面积的87.4%;建设用地约占总面积的0.7%,包括村镇道路和村民居住点等。当地已对生活污水进行了截污纳管,流域污染以非点源污染为主。研究区地理位置见图1。
-
为减少监测过程中的干扰,选择具有独立径流出口的林地、园地、耕地、村镇道路和屋顶进行监测(见图1)。在监测过程中,挡水墙和围堰等措施被用来保证监测小区不与周边地块发生径流交换,即实测地块集水面积。其中,林地面积约500 m2,为针阔混交林,采样时树木生长多年,地表杂草覆盖均匀。园地面积为152 m2,采样时为桃树果实生长期和成熟期。耕地面积为100 m2,主要作物为豆角和芋头,采样时为作物生长期。村镇道路排水口收集村镇地表排水,道路材质为水泥路面,集水面积约为1 000 m2。居民住宅屋顶长度为5 m,侧宽度为2.5 m,具有独立排水管。雨量计站点位于岩下村1.5 km处的黄山四村村民楼顶,型号为RG3-M型翻斗式雨量计(Onset Computer Corporation,美国 HOBO)。
监测时间从2020年4月初开始,到9月末结束。共监测到15次降雨,其中4次采集到完整径流过程,共采集样品122个,降雨信息见表1。在各地块排水口用带有刻度的集流桶收集径流,记录集流桶内径流体积和收集时间以计算采样时流量。采样频率参考以往的研究[25-26],初期每5 min采集1次,累计采集3次,中后期每10~30 min采集1次。用径流润洗后的聚乙烯采样瓶保存样品,样品采集后即送回实验室于5 ℃冰柜中冷藏,并于48 h内完成检测。
-
岩口水库曾发生过水华现象[27],监测结果表明部分区域水体总磷超标。根据以往研究发现,磷污染物输出可能与水土流失过程存在一定的关系,故本研究测定样品中的总磷(TP)、总溶解态磷(TDP)、总悬浮颗粒物(TSS)。原样品摇匀后采用0.45 μm滤膜过滤得到样品的TDP。样品采用过硫酸钾消解后,用钼酸盐分光光度法 (GB 11893-1989) 测定TP和TDP。总颗粒态磷(TPP)即为TP与TDP之间的浓度差值。采用烘干法(见《水和废水监测分析方法(第4版)》)测定TSS。
质量控制方法:采用标准样品和实际样品同步测量的方法,检验从制样到样品分析测定的整个分析过程的污染程度;采用标准药品和规范操作,并对实际样品进行平行样品测定。结果表明,标准物质的回收率为95%~105%,分析过程污染较小不影响试验结果;各平行样品的平行性均在10%以内;标准曲线的线性相关系数在0.992以上。
-
采用Excel进行数据统计计算,利用SPSS 24进行相关性分析,利用Origin2017进行图表绘制。分析过程中用到次降雨事件污染物平均浓度(event mean concentration, EMC)代表次降雨下垫面污染物的平均输出水平[28-29],其计算方法见式(1)。
式中:EMC为次径流污染物的平均浓度,mg·L−1;M为整个径流过程中污染物的质量,mg;V为径流总体积,L;Ct为t时刻污染物的浓度,mg·L−1;Qt为t时刻径流流量,L·min−1;Δt为采样间隔时间,min;t为降雨径流持续时间,min。
利用次降雨过程中污染物总质量,计算下垫面单位面积污染物输出强度[30],计算方法如式(2)所示。
式中:P为次径流污染的输出强度,kg·hm−2;M为整个径流过程中污染物的质量,kg;S为下垫面面积,hm−2;Ct为t时刻污染物的浓度,kg·L−1;Qt为t时刻径流流量,L·min−1;Δt为采样间隔时间,min;t为降雨径流持续时间,min。
以1 min内降雨量代表瞬时降雨强度,其数值通过雨量计监测得到。此外,用下垫面单位面积输出负荷乘以相应的下垫面面积得到次降雨的污染负荷。
1.1. 研究区概况
1.2. 样品采集及现场观测
1.3. 样品处理与质量控制
1.4. 数据分析
-
下垫面的差异对污染物输出过程有着重要影响。图2为一场典型降雨事件(8月4日,降雨量59.8 mm,图2(a)~(e))中流域主要下垫面污染物输出情况。由图2可知,5种主要下垫面的产流先后顺序最早为村镇道路和屋顶,其次是园地和耕地,最晚为林地(见图2(f)~(j))。在产流开始前30 min,园地、耕地和村镇道路输出的TSS浓度较高。产流初期三者输出的TSS浓度随着雨峰出现的时间产生波动性的变化,之后随着降雨量增加逐渐减小,这与以往研究结果较为一致[31-32](图2(g)~((i))。林地的产流时间晚于30 min,且在整场降雨过程中TSS的浓度变化趋于平缓,浓度值为156~261 mg·L−1(见图2(f))。这是因为,监测点的林地植被覆盖面广,地表裸露土地少,降雨初期可以拦截携带污染物的侵蚀颗粒被径流冲刷和传输。屋顶产流时间较快,但其TSS浓度为115 ~161 mg·L−1,变幅最小,整体较稳定(见图2(j))。
从磷素的输出过程来看,各下垫面的磷污染物浓度在产流过程中整体呈现出先增加后减少的趋势(图2(k)~(o))。对照《地表水环境质量标准》(GB3838-2002)比较下垫面径流水质发现,园地、耕地在降雨前期和中期TP浓度均超出《地表水环境质量标准》中Ⅴ类水标准,但在降雨后期污染物输出浓度介于Ⅲ类至Ⅴ类之间(见图2(l)~(m))。以上结果说明,过度施肥导致了这两类下垫面土壤本底磷素较高,应开展测土配方施肥以减少磷素积累。村镇道路在降雨前30 min内TP超出Ⅲ类水标准,最大值超过Ⅴ类水标准,但中后期浓度介于Ⅲ类和Ⅴ类之间(见图2(m))。相比于上述各下垫面,林地的TP输出较低,整体处于Ⅲ类水到Ⅴ类水范围(见图2(k))。而同属于建设用地的屋顶,TP输出最少,总体处于Ⅱ类水平(见图2(o))。由此可见,施肥及其他生产活动导致园地、耕地和村镇道路的TP总体超过Ⅴ类水标准,林地的TP总体在Ⅲ类水上下波动,而屋顶的TP总体则在Ⅲ类水以下。此外,下垫面的TP与TSS存在正相关关系。
-
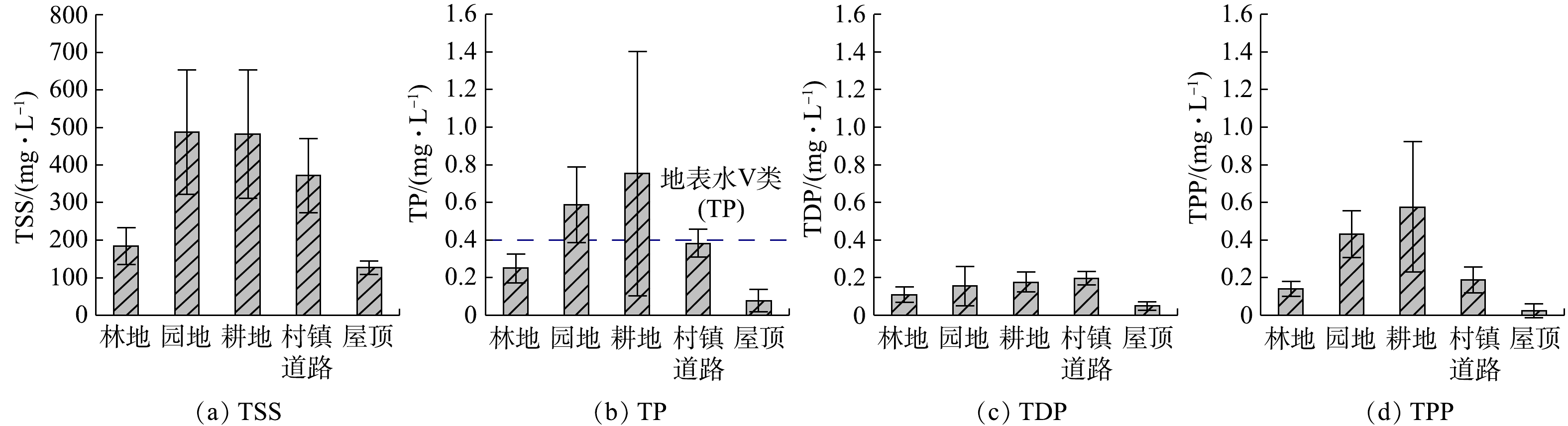
下垫面特征除了影响污染物输出过程外,还会影响次降雨污染物的平均输出水平(EMC)。为减少单次降雨污染物浓度的不确定性[33],采用多次降雨事件(6月19日、8月4日、8月7日和8月27日)EMC的平均值表征监测期间各下垫面的径流水质。下垫面的差异对TSS及磷污染物EMC特征同样有着重要影响(见图3(a)~(d))。5种主要下垫面输出TSS的EMC从大到小顺序为:园地(488 mg·L−1)>耕地(482 mg·L−1)>村镇道路(372 mg·L−1)>林地(184 mg·L−1)>屋顶(126 mg·L−1)(见图3(a))。这说明,随着降雨冲刷过程的发生,耕地和园地的土壤侵蚀较为严重。此外,5种主要下垫面上TP的EMC从大到小顺序为:耕地(0.75 mg·L−1)>园地(0.59 mg·L−1)>地表Ⅴ类水标准(0.40 mg·L−1)>村镇道路(0.38 mg·L−1)>林地(0.25 mg·L−1)>地表Ⅴ类水标准(0.20 mg·L−1)>屋顶(0.08 mg·L−1)(见图3(b))。考虑到园地和耕地输出TP的EMC同样最大,其污染河湖水质的风险较高,为控制小流域的磷输出,应优先治理农业种植用地。
进一步分析不同形态磷对TP的贡献,可发现建设用地的TP输出形态以溶解态TDP为主,村镇道路和屋顶的TDP对TP分别贡献了51.33%和70.71%;而其他3种下垫面的TP则以颗粒态TPP为主,林地的TPP对TP的贡献率为56.20%,园地和耕地则分别达到了73.63%和76.60%。由于建设用地的不透水面的比例比其他3种下垫面大得多,说明下垫面的类型会导致磷输出组成的差异。这主要是因为建设用地受居民生活习惯影响,更易累积散排的生活污水、生活垃圾等含TDP较多的污染物。为控制污染输出,应对居民倾倒污水和垃圾的习惯进行规范,减少直排散排。
-
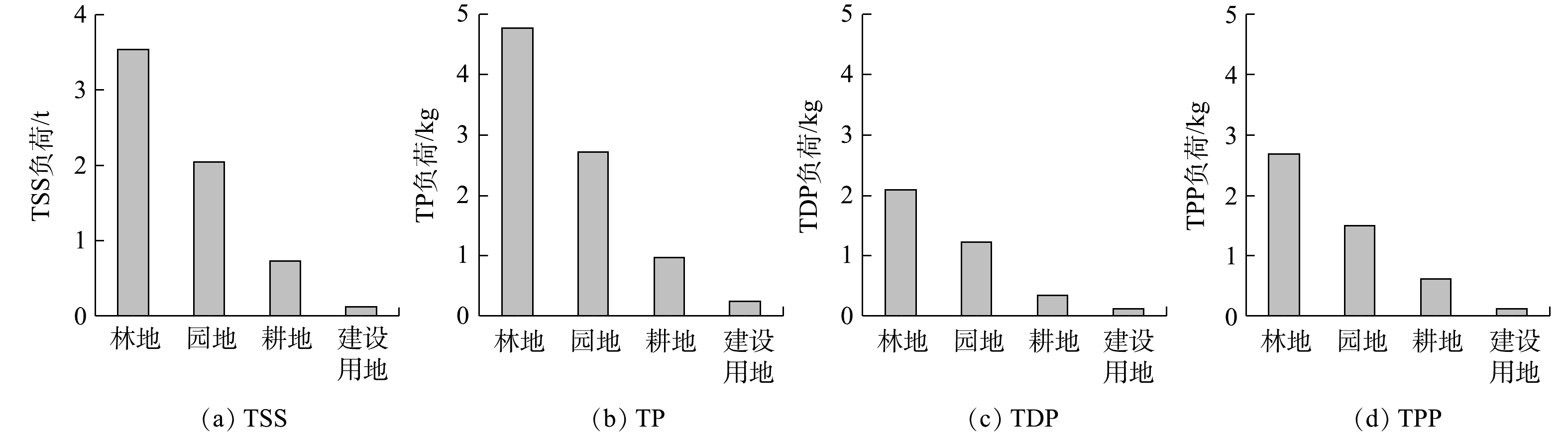
利用次降雨(8月4日)污染物的浓度、流量及监测点面积等数据,对下垫面的单位面积输出负荷进行了分析。总的来说,不同下垫面单位面积污染物输出负荷的大小顺序与污染物EMC的大小顺序明显不同(图4(a)~(d))。TSS的单位面积输出负荷高低顺序为:村镇道路(72.14 kg·hm−2)>耕地(41.88 kg·hm−2)>屋顶(33.52 kg·hm−2)>园地(33.07 kg·hm−2)>林地(14.28 kg·hm−2)(见图4(a))。而5种主要下垫面上TP污染物的单位面积污染负荷从大到小依次为:村镇道路(0.07 kg·hm−2)>耕地(0.06 kg·hm−2)>园地(0.04 kg·hm−2)>屋顶(0.021 kg·hm−2)>林地(0.019 kg·hm−2)(见图4(b))。这说明,相比于污染物EMC大小的排序,村镇道路和屋顶单位面积输出负荷的排序明显提高,这仍是因为下垫面透水/不透水特征的差异。村镇道路和屋顶的不透水面的比例较大,二者的单位面积径流量远大于园地和耕地等强透水下垫面,导致其单位面积输出强度有了较大提升。
因此,根据单位面积输出的TP强度,应重点关注村镇道路和农业种植用地对整个岩口水库流域总磷负荷的影响。首先,须对以园地和耕地为主的农业种植用地进行水土流失治理和源头控制;同时应加强流域内村镇道路初期雨水的截留,尤其注意在小到中雨情况下,村镇道路是流域主要产生径流污染的下垫面,其污染物具有直接入河的风险。此外,还应保护好林地,发挥其在水土保持、净化水质等方面的积极作用。
-
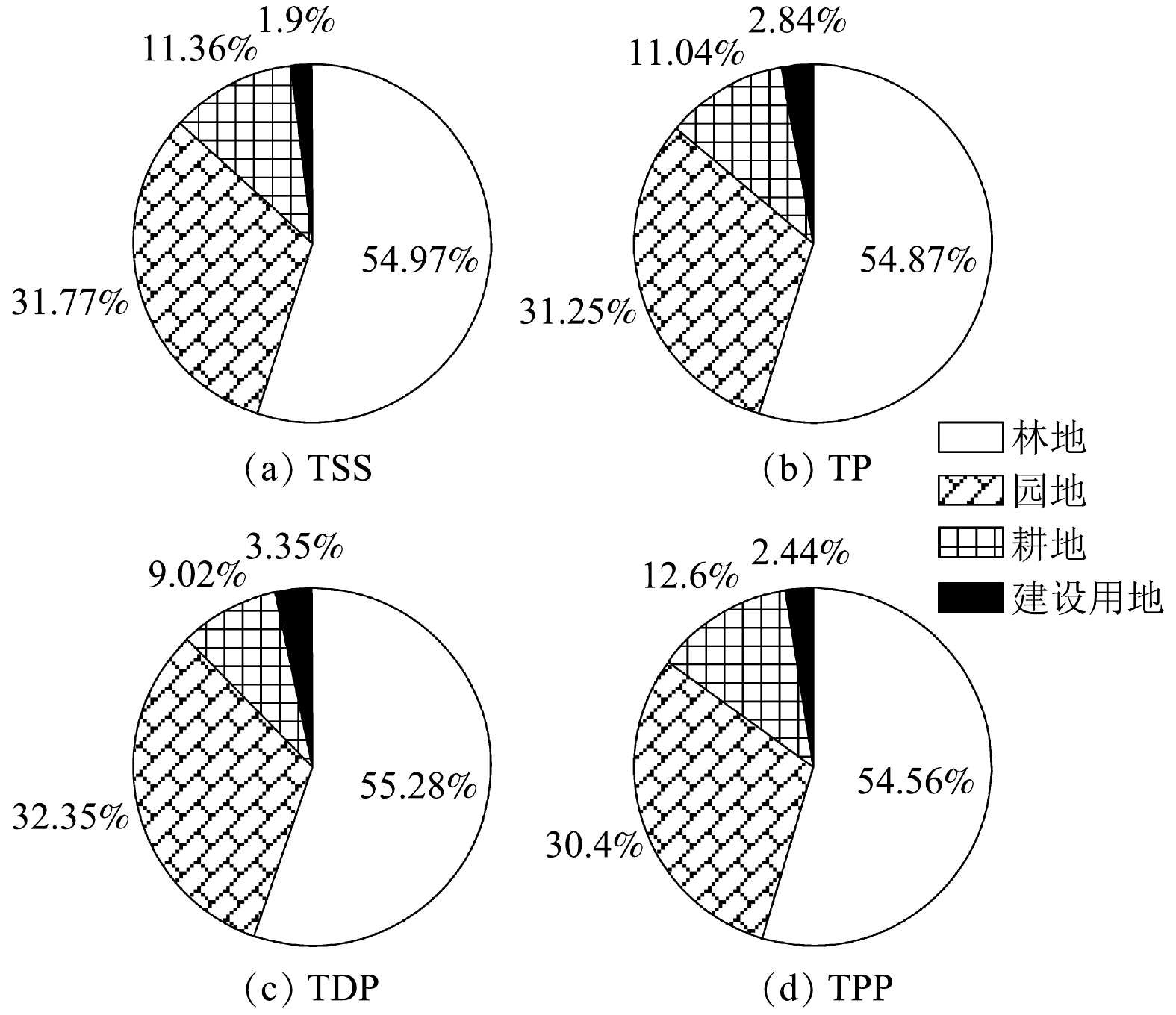
分析各主要下垫面对次降雨污染物的负荷及贡献比例,可以进一步为小流域综合治理提供决策依据。通过下垫面单位面积磷输出强度和小流域内各下垫面面积及作物种植比例,可计算出典型的次降雨尺度上(8月4日降雨,降雨量59.8 mm)岩下小流域内(4.52 km2)的非点源污染负荷,结果如图5所示。其中,建设用地的污染负荷包括了村镇道路和屋顶2种下垫面的负荷。在次降雨条件下,岩下小流域共计输出泥沙6.42 t,TP 8.7 kg。对输出TSS贡献最多的为林地,园地次之,两者的贡献比分别是54.97%和31.77%;对TP、TDP、TPP 3种负荷的前两位贡献顺序同样是林地和园地,其中,林地对TP、TDP和TPP负荷的贡献率达到54.87%、55.28%和54.56%,园地则为31.25%、32.35%和30.4%(见图6)。上述结果表明,尽管林地单位面积污染强度小,但其面积较大,仍然是污染负荷的主要贡献者。因此,应在重点治理园地和耕地水土流失的基础上,进一步加大对林地的保护,尤其是对林地中的裸露地表,应增加植被覆盖,并对水土保持措施进行维护。
2.1. 不同下垫面磷污染物的输出过程及规律
2.2. 不同下垫面磷污染物的次降雨平均浓度特征
2.3. 不同下垫面单位面积输出磷污染物负荷的特征
2.4. 次降雨事件不同下垫面的磷污染负荷估算
-
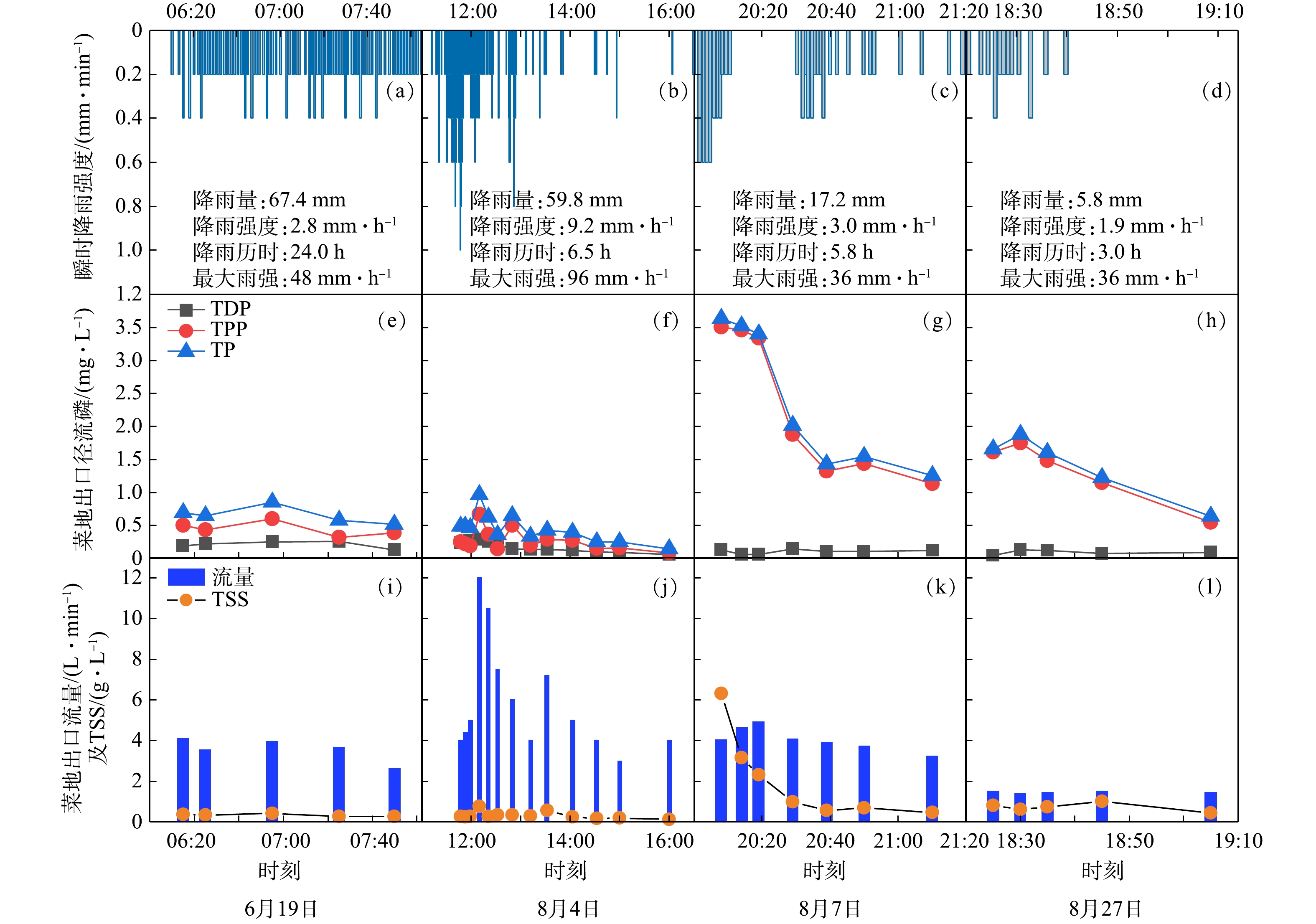
当降雨流量小于10 mm或者平均降雨强度小于1.8 mm时,园地和耕地等农业种植地块产流的概率较小(见表1),故降雨特征是磷污染物输出过程的潜在影响因子。为探究降雨事件的降雨强度、流量等对地块磷输出的影响,以耕地为例,分析了4场降雨条件下(6月19日、8月4日、8月7日和8月27日)其瞬时雨量、磷输出浓度、流量及TSS流失随时间的变化规律,结果如图7所示。由图7可知,降雨强度(见图7(a)~(d))对地块磷污染物输出有明显影响(见图7(e)~(h))。当瞬时降雨强度相对稳定时,磷浓度表现的相对稳定(见图7(a)和见(e));当瞬时降雨强度增加或减小时,磷浓度也会随之增加或减小(见图7(b)和(f))。
从TP、流量和TSS三者关系来看,2次大雨条件下(6月19日和8月4日),三者的变化趋势较一致(见图7(e)、(i)、(f)和(j))。然而,在中雨和小雨条件下情况则出现了变化。8月7日(中雨)降雨过程中,TP与TSS变化过程较为一致,而与流量变化明显不同(见图7(g)和(k));8月27日(小雨)降雨过程中,流量和TSS几乎不变,而TP则出现了先上升后下降的趋势(见图7(h)和(l))。这说明降雨类型影响了下垫面TP、流量和TSS之间的关系[34],可能是径流的冲刷侵蚀强度随降雨条件出现了差异,影响了TP的输出,进而导致TP、流量和TSS之间的关系发生变化。同时,这也说明TP输出特征可能受到流量和TSS流失过程的影响。以上结果可能对不同降雨条件下非点源污染控制措施的效率研究提供参考。
-
以8月4日降雨为例,对不同下垫面径流中磷输出浓度与TSS做相关性检验,得到如表2所示结果。该结果表明,监测过程中5种主要下垫面输出的TP、TPP与TSS均显著相关,TP、TPP与TSS的相关系数最高分别达到0.993和0.990,说明TP和TPP的输出过程与TSS的流失过程关系密切。这可能是由于降雨冲刷造成了土壤侵蚀,而土壤侵蚀发生后导致颗粒物和磷的流失[35-36],其中TPP流失与颗粒物释放的同步性更强。该结果表明,为控制小流域的磷输出,应减少各下垫面的TSS流失。比如,园地和耕地的径流出口增加水土保持措施,在村镇道路排水渠周围设置一定的低影响开发措施(如植草沟等)[37-39]。
为进一步探究磷输出浓度与流量的关系,接着对不同降雨条件下(6月19日、8月4日和8月7日)园地和耕地2种下垫面的磷输出浓度与流量进行皮尔逊相关性检验。结果表明,在8月4日和8月7日的2次降雨过程中,耕地的TP与流量显著相关,如表3所示。而园地的TP与流量则一直未显著相关(表3中未列出)。以上结果表明,地块的磷输出可能与流量有关系,但这种关系因地块类型而存在差异。其中,以菜地为代表的耕地土质松软,径流冲刷导致磷污染物输出效果明显,从而磷输出与流量关系密切;而以桃园为代表的园地情况则相反,其下垫面相对坚硬,径流变化与磷输出存在一定的时间间隔,致使磷输出与流量相关性较差[40]。上述结果进一步表明,不同下垫面的磷输出具有不同特征。因此,建议在实施治理措施控制非点源污染时,注意对不同土地类型进行精准配置以提升控制效率。此外,未来可结合人工模拟等方法验证磷输出与流量的关系,以及不同水土保持措施下非点源污染物的输出特征。
3.1. 降雨特征对磷污染物输出过程的影响
3.2. 磷输出特征与TSS流失和流量的关系
-
1)受污染物累积过程和径流冲刷过程的影响,林地、园地、耕地、村镇道路和屋顶5种主要下垫面的污染物输出过程、EMC特征、磷输出组成和单位面积输出负荷,存在明显差异。
2)从污染物输出过程来看,各下垫面产流过程中受瞬时强降雨影响,均存在产流初期TP较高的现象,其中园地、耕地及村镇道路的TP总体超过V类水标准。
3)从污染物输出结果来看,次降雨情况下农业种植用地输出的TP降雨平均值最高,村镇道路输出的单位面积TP负荷最高,而林地输出的TP对小流域污染总量贡献最多。在小流域治理中,应综合考虑上述结果,因地制宜制定污染控制措施。
4)除了下垫面外,降雨强度、降雨量、流量和水土流失过程等均会影响磷污染物输出特征。而通过皮尔逊相关性检验还发现,次降雨条件下TP和TPP的输出与TSS的流失呈正相关关系,可通过减少各下垫面TSS的流失控制小流域的磷输出。
