

 DownLoad:
DownLoad:



-
我国畜禽养殖业发展迅速,规模化养猪场的比例逐年增加,但畜禽粪便的资源化利用率较低,从而导致了较严重的环境污染问题,畜禽粪便已成为农业面源污染的主要来源[1]。干清粪收集工艺大大提高了粪便含固率,为开展农业废弃物厌氧干发酵技术创造了有利条件;同时,通过干发酵可获得丰富的清洁能源沼气,有利于我国农业的绿色可持续发展[2]。
厌氧干发酵处理能力大、用水量小、容积产气率高。而且,发酵后的沼渣含水率低,能够直接生产有机肥,有助于构建种养循环体系[3]。但是,由于农业废弃物组分结构复杂、难降解转化,易产生酸积累,会产生启动慢、传质效率低和发酵过程稳定性差等问题[4]。为有效缓解厌氧干发酵的酸抑制、提高其甲烷产率,相关学者开展了大量的研究工作。如齐利格娃等[5]研究了猪粪与稻草的不同原料配比对厌氧发酵性能的影响,当猪粪与稻草配比为2∶1时,丙酸含量较纯猪粪组减少了71.8%,累积VS甲烷产率较纯猪粪组相比提高了13.0%。于佳动等[2]研究了喷淋频率对玉米秸秆和牛粪的混合厌氧干发酵影响,当接种量为30%时,喷淋频率间隔8 h的有机酸浓度相较于间隔2 h增加了46.3%,累积甲烷产量也相应降低。刘春软等[6]研究了不同添加剂对猪粪厌氧干发酵特性的影响,添加2.5%的生物炭处理组的总有机酸(TVFAs)峰值较纯猪粪组减少了4.8%,平均甲烷含量提高了4.4%。
目前,关于厌氧干发酵酸积累的研究主要集中在发酵工艺改进和影响因素筛选及优化等方面,但由于喷淋和外源添加剂在实际应用中成本高且不易开展规模化应用,因此对避免和缓解干发酵中酸抑制仍需开展更深入的研究。本研究以猪粪为主要发酵底物,通过中温批式实验研究底物非混合接种对厌氧干发酵过程有机酸积累和产甲烷特性的影响,并采用修正的Gompertz动力学模型模拟产气过程,进而评估非混合接种对猪粪厌氧干发酵的促进作用。
全文HTML
-
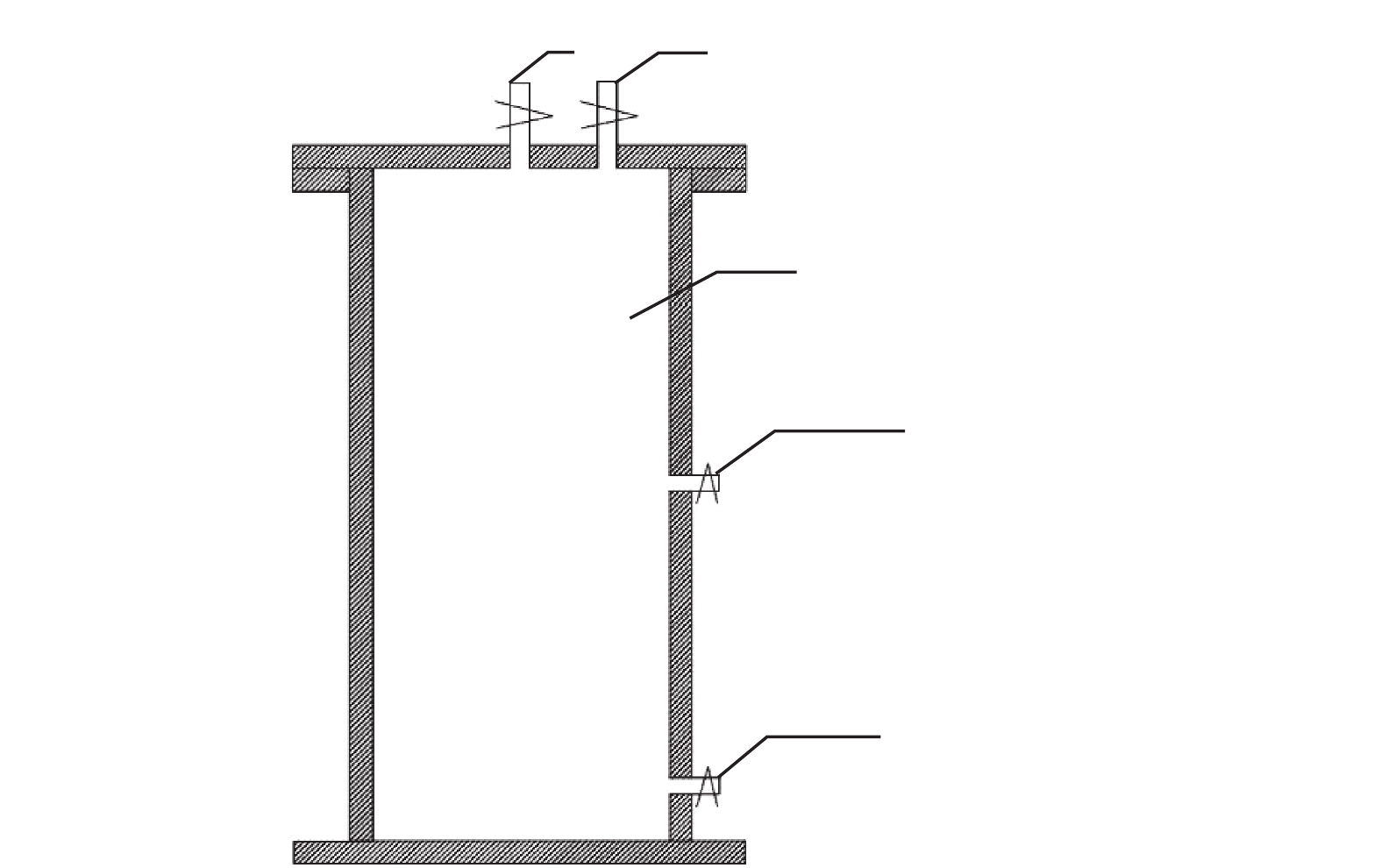
实验用猪粪为天津市西青区某养殖场的鲜猪粪,取回后储存于(4±1) ℃的冰箱。玉米秸秆收获后进行粉碎,粒径为1.0~3.0 mm,存放于室温阴凉通风处。接种物取自实验室自产的猪粪厌氧干发酵残余物,已不产气。底物与接种物的理化指标见表1。实验装置为自制批式厌氧干发酵反应器(见图1)为有机玻璃材质,内径90 mm,高170 mm,总容积为1.1 L。
-
按不同的发酵底物和接种方式设4组实验,每组实验3个重复。具体实验设计见表2。其中,非混合接种(P-CL组和P-ML组)实验组中,采取的进料方式为分层进料:首先分别将接种物和底物按质量均分3份,然后取1份接种物平铺至反应器底部,再将1份底物均匀铺至接种物层上方(不搅拌),重复上述操作共3次,最后形成接种物位于底物下方共3层。各反应器的总进料量为600 g(TS=20%),接种比为25%(按VS计),装料后将反应器加盖密封,连接3 L集气袋,置于(37±1) ℃恒温水浴锅中发酵。
根据发酵产气量,每1~3 d测量沼气产量,并测定气体组分。每3 d从发酵罐侧面取样口采集发酵固体样品(其中,非混合接种的实验组从发酵罐侧面上层和下层取样口进行取样后,再将其混合均匀),测量pH、溶解性化学需氧量(SCOD)、挥发性有机酸(VFAs)和氨氮的理化指标。
-
总固体含量(TS)、挥发性固体含量(VS)、总氨氮(TAN)和pH采用《水和废水监测分析方法》[7]分析测定;C、N含量采用Vario EL cube元素分析仪测定。
总有机酸(TVFAs)测定:固体样品经去离子水稀释10倍(质量计)后加稀硫酸调节pH<3.0,离心10 min后取上清液与丙酮按比例混合,混合液经0.45 µm滤膜过滤后采用Thermo-trace-1300气相色谱仪测定VFAs(乙酸、丙酸、丁酸和异丁酸)质量浓度,气相色谱配毛细管柱(TR-FFAP,30 m×0.53 mm×1 µm),氦气为载气(流速8.00 mL·min−1),柱箱温度为90 ℃,进样口、FID检测器温度分别为200和220 ℃。VFAs总质量浓度为各种VFA浓度之和(以乙酸计)[8]。
SCOD采用哈希快速消解方法[9]测定。游离氨(FAN)的质量浓度[10]通过公式(1)计算。
式中:CFAN为游离氨质量浓度,mg·g−1;CTAN为总氨氮质量浓度,mg·g−1;t为厌氧发酵温度,℃;pH为发酵固体样品的pH。
沼气产量用湿式气体流量计配蠕动泵进行测量,沼气中CH4和CO2体积分数采用Thermo-trace-1300气相色谱仪测定(Porapak Q色谱柱(2 m×φ2 mm)、氦气为载气(压力75 kPa)、柱箱温度40 ℃、进样口和热导检测器(TCD)温度均为200 ℃)。
-
采用修正的Gompertz模型模拟实验过程中的累积VS产甲烷量[11]。模型方程见式(2)。
式中:P为t时刻对应的累积VS产甲烷量,mL·g−1;Pmax为最终累积VS甲烷产率,mL·g−1;Rmax为最大VS产甲烷速率,mL·(g·d)−1;λ为迟滞期,d;t为时间,d;e为exp(1)=2.7183。根据模型拟合结果推算发酵过程中最大VS产甲烷速率Rmax和迟滞期λ。
1.1. 实验原料与装置
1.2. 实验设计
1.3. 分析方法
1.4. 动力学模型
-
图2为各处理发酵过程中TVFAs、乙酸、正丁酸、丙酸和异丁酸的质量浓度变化曲线。由图2(a)可知,发酵的前7 d各处理组的TVFAs质量浓度不断上升。这是因为,在反应初期,有机物被水解产酸细菌分解成VFAs,由于水解产酸细菌的生长速度快于产甲烷菌,使得产生的VFAs不能被及时分解,各组的TVFAs质量浓度不断上升[12],第7天达到第一个质量浓度峰值。其中,P-C组达到30.1 mg·g−1,较P-M组、P-ML组和P-CL组分别高出了11.0%、18.1%和22.8%。P-C组在第37天时的TVFAs质量浓度达到第2个峰值24.6 mg·g−1,其后质量浓度明显降低并在46 d后趋于平稳,至结束时(第63天),TVFAs质量浓度仍维持在15.2 mg·g−1附近。P-M组的TVFAs在30 d后呈现明显下降趋势,在发酵第40天时的TVFAs质量浓度为9.6 mg·g−1,之后缓慢降低,至实验结束时为3.6 mg·g−1。P-CL组和P-ML组的TVFAs质量浓度则在21 d后急剧下降。其中,P-CL组在25 d后又经历了1个先升高再下降的过程,实验结束时为2.4 mg·g−1,略低于P-M组。可以看出,P-C组和P-CL组在20~25 d后均有1个明显的TVFAs质量浓度先升高后降低的过程;而P-M组和P-ML组此过程不明显或者不存在。其主要原因是,猪粪中不同有机质组分的降解难易程度不同,当单独猪粪为发酵底物时,需要较长时间才能充分将有机酸消耗利用;而且,当玉米秸秆与猪粪两种底物发酵,体系为微生物提供更均衡的营养,发酵前期加快有机物水解酸化进程,从而减少发酵体系内有机酸的积累。P-ML组TVFAs质量浓度在第25天时降低至2.6 mg·g−1,并在33 d后稳定在0.5~0.8 mg·g−1,实验结束时为0.7 mg·g−1,较P-C组、P-M组和P-CL组分别减少了95.3%、80.2%和70.7%。TVFAs质量浓度降低主要是因为随着发酵的进行,产甲烷菌的适应性逐渐增强,能及时将产生的VFA分解转化为甲烷,但不同处理组之间TVFAs质量浓度发生明显下降的时间及降低幅度差别明显。与混合接种(P-C组与P-M组)相比,非混合接种(P-CL与P-ML)具有更低的VFAs质量浓度,且出现明显下降的时间提前。
图2(b)为发酵过程中乙酸质量浓度的变化曲线。可以看出,P-CL组在发酵的第40天出现第2个明显的乙酸峰值,达到4.6 mg·g−1,相较于P-C组和P-M 组分别降低了42.6%和41.2%。P-ML组的乙酸质量浓度只有1个峰值,在21 d后,TVFAs中乙酸含量不断增加,并在第33天达到76.3%后小幅度波动。
图2(c)中,正丁酸质量浓度整体呈先升高后降低的变化趋势,在发酵结束时,各组的正丁酸基本消耗殆尽。在发酵的第7天,各组表现出1个明显的峰值,P-CL组和P-ML组分别为6.7和8.1 mg·g−1,较P-C组(8.6 mg·g−1)和P-M组(9.2 mg·g−1)分别降低了22.0%和12.1%,且在第21天时正丁酸质量浓度减少到1.0 mg·g−1以下。以上结果表明,非混合接种能够促进乙酸和正丁酸的分解转化,降低质量浓度,有利于产甲烷发酵,而且猪粪玉米秸秆混合物非混合接种(P-ML组)效果较明显。这是由于非混合接种方式使局部产甲烷微生物占主导优势,可迅速消耗发酵体系内产生的有机酸。同时,以玉米秸秆和猪粪为底物进行非混合厌氧发酵时,一方面两种底物平衡了发酵体系的C/N比;另一方面玉米秸秆疏松多孔的结构,有利于气体的释放,提高了微生物对原料的利用效率,从而有效促进该发酵体系内的有机酸分解转化。
从图2(d)中可以看出,除P-C组的丙酸质量浓度呈升高的趋势外,其余实验组均呈先升高后降低的总体趋势。在发酵的第21天,P-CL组和P-ML组表现出1个明显的丙酸质量浓度峰值,分别为8.6和11.4 mg·g−1;随后,P-CL组和P-ML组的丙酸含量和占比不断减少,在发酵结束时丙酸基本被完全消耗。混合接种的实验组(P-C组和P-M组)在前49 d丙酸质量浓度增加。其中,P-M组在30 d后TVFAs中丙酸含量超过P-C组,在第49天后丙酸质量浓度快速下降,发酵结束时为2.5 mg·g−1,较P-C组减少了70.1%。这表明,在混合接种发酵组中,底物为猪粪玉米秸秆混合发酵组(P-M组)的乙酸消耗较快,使TVFAs中丙酸含量不断提高。WANG等[13]研究不同VFAs对厌氧发酵的影响时发现,当丙酸质量浓度达900 mg·L−1时,发酵体系内的产甲烷微生物受到明显抑制。在本实验中,非混合接种发酵组的甲烷产量和甲烷体积分数均未出现明显的下降趋势(图3)。有研究[14]指出,丙酸积累不利于甲烷生产,当体系以乙酸和丁酸为主要有机酸时,有利于提高体系产气量。本实验中的P-C组随着发酵的进行,丙酸质量浓度不断增加,发酵结束时的丙酸质量浓度积累至8.3 mg·g−1,该浓度远超过相关研究[13]的抑制浓度(900 mg·L−1)。P-C组丙酸质量浓度积累形成了酸抑制现象,从而影响了该组的产气效果,这也与图3(a)的结果一致。各组异丁酸的质量浓度变化(图2(e))基本一致,呈先升高后减少的趋势。第25天时,P-C组、P-M组、P-CL组和P-ML组异丁酸质量浓度分别为2.2、1.1、0.3和0.1 mg·g−1。其中,P-CL组和P-ML组分别在第56天和第37天将发酵体系内的异丁酸全部转化完,这说明非混合接种发酵体系更容易利用异丁酸。
-
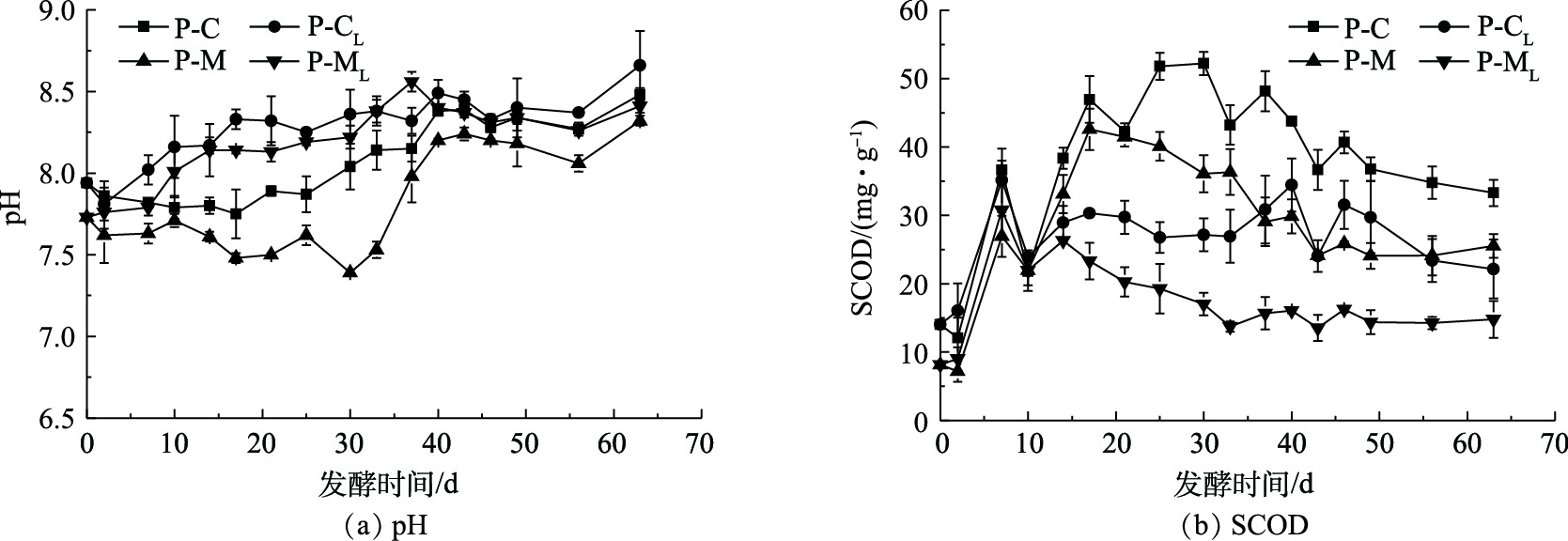
图4为实验过程中各处理组的pH和SCOD随时间变化曲线。由图4(a)可以看出,各发酵组pH大小基本呈现P-CL组>P-ML组>P-C组>P-M组的规律,其中非混合接种高于混合接种。在发酵的前30 d,P-M组的pH总体呈下降趋势。这可能是因为,发酵原料中玉米秸秆的纤维素和半纤维素的水解产酸速率较快[15],但产甲烷菌消耗VFAs的速率相对较低导致了有机酸不断积累。同样,底物原料含有玉米秸秆的P-ML组的pH则呈缓慢升高趋势。这是因为,非混合接种改变了有机酸和微生物的接触方式,使底物层和接种物层附近区域的F/I比降低、产甲烷微生物发挥出群体优势,这表明非混合接种能够加快有机酸转化甲烷效率。P-M组和P-ML组与P-C组和P-CL组相比,具有较低的TVFAs质量浓度(图2(a)),但pH仍低于P-C组和P-CL组。这主要是因为,P-M组和P-ML组处理中添加了玉米秸秆,降低了体系中TAN质量浓度(图5),并导致该发酵体系的缓冲能力有所下降。P-ML组和P-CL组具有相对较高的pH,这表明非混合接种的发酵方式可以促进产甲烷微生物对VFAs的消耗,进而避免VFAs积累。
SCOD反映了发酵样品中可溶性有机物的含量,这是衡量酸化过程中,水解酸化能力的重要指标,稳定的SCOD可为微生物厌氧发酵提供稳定的物质基础[16-17]。由图4(b)可以看出,4个实验组的SCOD呈先升高后降低再升高最后降低趋于稳定的总体变化趋势,且与TVFAs(图2(a))变化规律相似。在4个发酵组中,猪粪接种物全混合发酵组(P-C组)的SCOD值最高,且变化幅度最大,在发酵的第7天和第30天,SCOD值分别达到36.6和52.2 mg·g−1这2个峰值。发酵结束时,P-C组的SCOD值为33.3 mg·g−1,这分别是P-CL组、P-M组和P-ML组的1.5、1.3和2.3倍。P-CL组和P-ML组的第2个SCOD峰值分别为34.4和26.3 mg·g−1,相较于P-C组和P-M组分别降低了34.1%和38.3%。P-CL组和P-ML组在发酵10 d后的SCOD相对更稳定,且明显低于P-C组和P-M组,其中P-ML组的SCOD值最小。这说明,非混合接种能够维持发酵体系SCOD的稳定同时促进发酵体系的水解,提高甲烷产率。其中,猪粪玉米秸秆非混合接种发酵组更能促进体系内的大部分的可溶性有机物被微生物水解利用转化为甲烷。
-
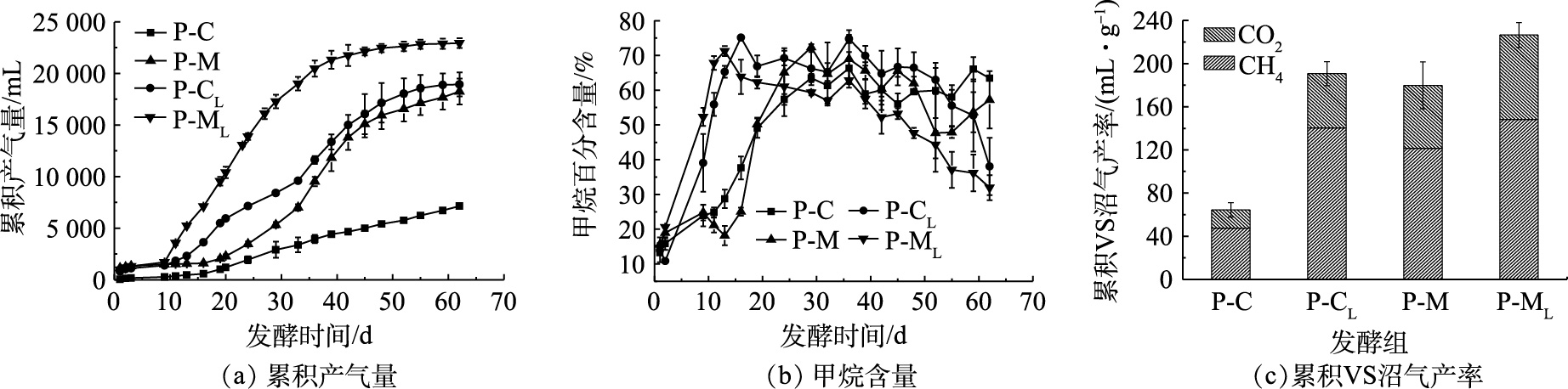
图3为各处理组发酵过程中累积产气量、甲烷含量和累积VS沼气产率。结合图3(a)和表3可以看出,各实验组经历了11~19 d的迟滞期后进入快速产气阶段,然后产气才渐趋于平稳。发酵结束时,P-CL组和P-ML组的累积产气量分别为18.9和22.9 L,较相同底物全混合接种的发酵组分别提高了164.9%和25.8%。由前期研究[18]可知,猪粪玉米秸秆混合原料非混合接种未出现迟滞期。本实验非混合接种发酵组均出现迟滞期的主要原因可能是,接种物在发酵实验前未驯化培养,因此产甲烷菌发酵前期活性较低,产气量较少。据图3(a)显示,在10~33 d内,P-CL组和P-C组的产气速率较其余组减慢,结合图4(b)可知,P-CL组和P-C组的SCOD质量浓度在发酵的10~40 d和10~37 d分别增加了30.5%和54.8%,而其余发酵组的SCOD质量浓度是不断降低的。这说明,在底物为猪粪的发酵组中,有机物的水解酸化可正常进行,但对氨氮浓度较为敏感的产甲烷菌活性可能受到抑制,从而导致产气速率减慢。综合图3(a)和图3(c)可以看出,P-CL组较P-M组相比,产气效果较好,集中在发酵前期(前33 d)且发酵启动最迅速;而在后期P-CL组的日产气速率逐渐减缓,在发酵结束时,2组的累积VS沼气产率差异不明显。其可能的原因是,发酵体系内的微生物在前期更容易利用猪粪中的有机物进行繁殖,从而提高累积产气量。随着发酵的进行,猪粪玉米秸秆混合原料的P-M组体系内碳源和氮源充足,产气未受影响,P-CL组则出现氮源过多,氨氮含量升高进而对产甲烷菌活性有抑制作用,这也与图5(a)的结果一致。
沼气中甲烷含量能够反映厌氧发酵体系碳素的转化效率[19]。尽管在不同接种方式下,各实验组在产气高峰阶段的甲烷含量差异不大,均在70%左右,但各组从发酵启动阶段到沼气中甲烷含量相对稳定(70%)所经历的时间有明显的差别,这段时间可以反映底物被产甲烷菌群利用的难易程度。在非混合接种实验组(P-CL组和P-ML组)中,甲烷浓度在13~16 d达到峰值(75.1%和71.3%),较P-C组和P-M组有明显的提前,这表明非混合接种发酵能够促进微生物可利用物质的甲烷化,与图4(b)的结果一致。
由各处理累积VS沼气产率的结果(图3(c))可知,P-ML组和P-CL组的累积VS甲烷产率分别为148.2和140.4 mL·g−1,分别是P-M组(121.5 mL·g−1)和 P-C组(47.5 mL·g−1)的1.2倍和3.0倍,其值略高于猪粪与香蕉秸秆混合厌氧干发酵[20](128.0 mL·g−1)和添加20%生物炭的猪粪厌氧干发酵[21](126.2 mL·g−1)的累积VS甲烷产率。为更好地比较不同接种方式对缓解厌氧干发酵酸积累的作用,本研究的接种比仅为25%,低于齐利格娃等[5]研究中的40%,从而导致P-C组的累积VS甲烷产率相对较低(188.8 mL·g−1)。以上结果表明,非混合接种对猪粪单一原料和猪粪玉米秸秆混合原料发酵均具有明显的产甲烷促进作用。其原因可能是,底物和接种物分层进料,使主要的产甲烷作用不在VFAs积累的区域进行,避免了有机酸对产甲烷菌的抑制,从而提高了甲烷产率[20]。同时可知,混合原料发酵的产甲烷效果优于猪粪单独发酵,这一方面是因为混合原料平衡了发酵底物的碳氮比;另一方面是因为猪粪玉米秸秆混合增加介质传导[22],进而提高产气效率。
用修正的Gompertz模型对干发酵累积VS甲烷产率进行了拟合,结果表明(表3),不同处理的拟合曲线呈现较高的拟合度。猪粪玉米秸秆混合原料非混合接种发酵组(P-ML组)的迟滞期(λ=10.6)最短,猪粪非混合接种发酵组(P-CL组)次之(λ=12.4),而猪粪接种物全混合发酵(P-C)的迟滞期最长(λ=18.5)。其中,非混合接种实验组(P-CL组和P-ML组)迟滞期的时长较其余组减少了24.6%~42.6%。在发酵结束的第62天,各实验组中只有P-CL组和P-ML组已进入产气平稳阶段。这表明,非混合接种方式不仅可实现猪粪厌氧干发酵的快速启动,还能够减少有效发酵时间。在产甲烷速率方面,P-C组最大VS产甲烷速率为0.98 mL·g−1,其值为各处理组最小。P-M组、P-CL组和P-ML组的最大VS产甲烷速率分别为3.6、4.8和6.2 mL·g−1,是P-C组的2.7、3.9和5.4倍。由此可见,非混合接种能够有效缩短迟滞期、提高产甲烷速率。
-
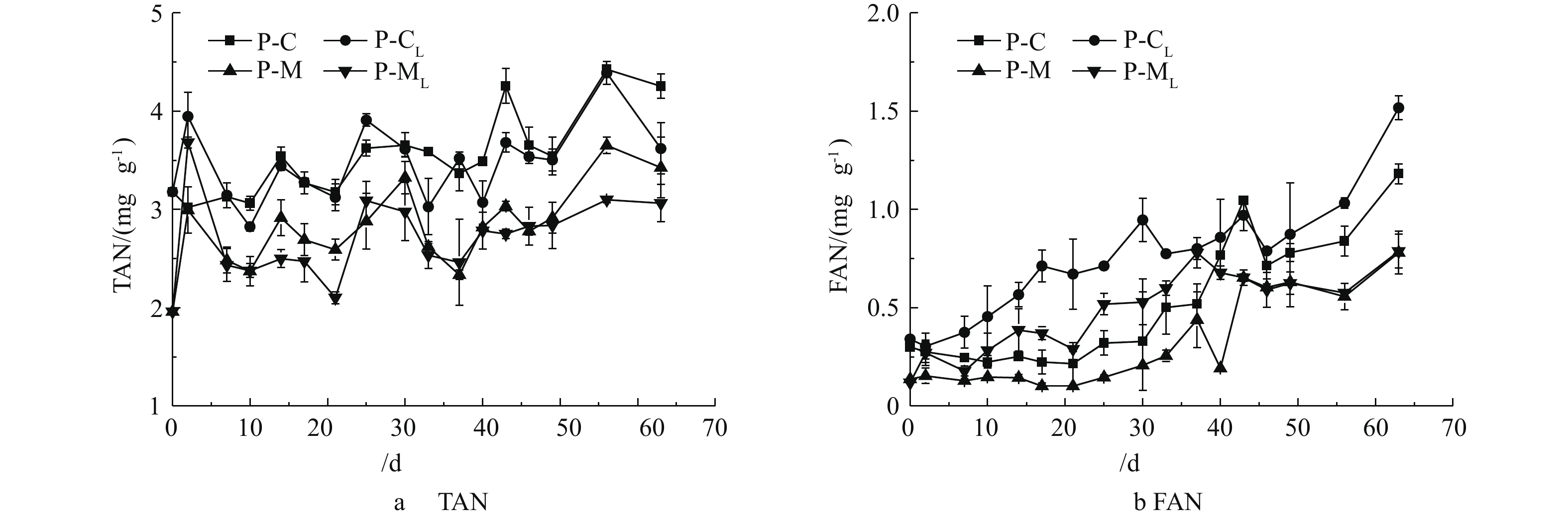
氨氮是厌氧发酵需要关注的重要指标之一,质量浓度过高会抑制微生物的产甲烷作用。图5为发酵过程中TAN和FAN的质量浓度变化曲线。据图5(a)显示,随着发酵的进行,TAN质量浓度呈逐渐上升的趋势[23],但不同实验组的TAN质量浓度差异明显。在发酵7 d后,以猪粪玉米秸秆为原料的P-M组和P-ML组TAN质量浓度范围在2.1~3.7 mg·g−1内波动,总体低于猪粪单一原料的P-C和P-CL组(2.8~4.4 mg·g−1)。P-M组与P-ML组、P-C组与P-CL组因为发酵原料相同,TAN质量浓度差别不明显,但P-ML组和P-CL组的累积VS甲烷产率较P-M组和P-C组却分别增加了22.0%(121.5 mL·g−1、148.2 mL·g−1)和195.6%(47.5 mL·g−1、140.4 mL·g−1),这与前期研究[18]结果一致。陈闯等[24]的猪粪连续干式发酵的实验结果表明,当氨氮浓度从2 250 mg·L−1(TS=20%)增加到3 800 mg·L−1(TS=35%),VS平均产气率减少了74.2%,从660.0 mL·g−1下降到170.0 mL·g−1。在本研究中,P-ML组和P-CL组的TAN质量浓度在第56天分别达到4.4和3.1 mg·g−1后,累积VS甲烷产率依然明显增加。这可能是因为,一定范围内的TAN质量浓度增加,有利于发酵体系的缓冲能力提高,加速中间产物如有机酸的转化,为产甲烷菌提供生长繁殖所需的营养物质[25]。以上结果表明,影响产气性能的主要原因是有机酸的积累而并非高质量浓度的氨氮的抑制作用;同时,非混合接种有助于提高发酵体系对氨氮的耐受浓度。
据图5(b)中显示,FAN质量浓度由于TAN质量浓度及pH的不同有较大的差异,但各组的总体变化趋势是逐渐升高。反应开始后,除P-M组外,其余组的FAN质量浓度随pH的增大迅速增加。HANSEN等[26]认为,当FAN质量浓度超过1 100 mg·L−1时,产甲烷菌的活性会受到抑制进而影响产气效率。在本研究中,P-CL组和P-C组在第30天和第43天时FAN质量浓度分别为1.0和1.1 mg·g−1,随后呈升高趋势,此阶段的FAN质量浓度均接近或超过已有研究[26]中报道的抑制浓度(1 100 mg·L−1)。发酵结束时,P-CL组的FAN质量浓度1.5 mg·g−1仍高于抑制浓度,是P-C组的1.3倍,但累积产气量是P-C组的2.6倍。这表明,非混合接种发酵可有效减轻高氨氮对产甲烷微生物的抑制作用,从而提高产气量。
2.1. 厌氧干发酵过程中的VFAs变化
2.2. 厌氧干发酵体系的pH和SCOD变化
2.3. 厌氧干发酵产甲烷性能及动力学分析
2.4. TAN和FAN质量浓度变化
-
1)猪粪玉米秸秆混合原料非混合接种组的TVFAs质量浓度在实验结束时为0.7 mg·g−1,较其余组减少了70.7%~95.3%,这表明非混合接种的方式可有效降低有机酸的积累浓度。
2)非混合接种发酵中,猪粪玉米秸秆混合原料非混合接种产气效果最佳,累积VS甲烷产率达148.16 mL·g−1。猪粪非混合接种发酵的沼气中甲烷含量最高为75.1%。
3)猪粪玉米秸秆混合原料非混合接种组和猪粪非混合接种组的氨氮质量浓度在第56天分别达到4.4和3.1 mg·g−1时,产气未受明显抑制。这主要是因为底物与接种物非混合发酵有助于提高发酵体系对氨氮的耐受浓度。
4)底物与接种物非混合发酵能够缓解有机酸积累、缩短厌氧发酵的启动时间、促进甲烷化过程,从而提高产气量。
