










-
废水生物脱氮、防治水体富营养化一直都是水处理方向面临的主要问题之一[1]。传统的生物脱氮方法主要为全程硝化反硝化脱氮。短程硝化作为一种新型生物脱氮工艺,是将硝化反应控制在氨氧化阶段,将亚硝酸盐氧化菌(nitrite oxidizing bacteria,NOB)分离出反应器并将氨氧化菌(ammania oxidizing bacteria,AOB)保留在反应器中,实现氨氮氧化产物为亚硝酸盐的过程[2]。相比传统的全程硝化,短程硝化可以节省25%曝气量和30%的反应时间[3],节约碳源40%[4],具有较低的污泥产量[5]。目前提出的控制短程硝化的影响因素包括游离氨(free ammonia,FA)、游离亚硝酸(free nitrite acid,FNA)、溶解氧(dissolved oxygen,DO)、温度、pH等[6-9]。此外,曝气时间也是实现和维持稳定短程硝化的主要控制因素,曝气时间短导致反应不完全,出水不达标[10];曝气时间过长,短程硝化会转化为全程硝化[11]。因此,维持稳定长久的短程硝化需要综合考虑各种控制因素,以实现对反应过程的综合控制。
除上述影响因素外,盐度也会对生物脱氮处理过程产生影响。有研究表明,低浓度的盐可以促进微生物的生长,盐度过高则会影响细胞的渗透压,导致微生物的活性受到抑制[12]。盐度对微生物虽有毒害作用,但可通过逐步提高盐度或投加耐盐污泥,使微生物适应盐环境,减轻盐度对微生物的抑制作用[13]。HAMODA等[14]研究发现,采用活性污泥法处理高盐废水时,系统生物活性和有机物去除率均有提高;ASIAN等[15]研究表明,一定的盐度对微生物活性有促进作用;徐洁等[16]采用序批式曝气生物滤池工艺处理含海水的污水,发现AOB的耐盐能力高于NOB。因此,可通过添加盐度驯化的方式,抑制NOB的活性,实现短程硝化。短程硝化技术应用前景广泛,但通过活性污泥法进行短程硝化,稳定性较难控制,且硝化细菌本身作为自养菌生长缓慢,非常容易从反应器中流失[17],这都增加了短程硝化工艺运行的困难。
包埋固定化技术通过包埋材料(人工或天然高分子材料)将游离细胞或者酶定位于限定区域,使其保持活性并可反复利用,是一种有效的防止菌体流失、提高反应器内生物量的生物截留手段[18-20]。与活性污泥法相比,包埋固定化技术具有处理效率高、反应易于控制、菌种高纯高效、生物浓度高、固液分离效果好等优点[21]。因此,本研究采用水性聚氨酯(waterborne polyurethane,WPU)为包埋材料制作硝化包埋颗粒,研究盐度及曝气时间对包埋颗粒短程硝化效果的影响,并以盐度为控制因素探究其对连续流短程硝化反应器启动及运行的影响,以期为包埋固定化技术与短程硝化工艺的耦合脱氮提供参考,并为含盐废水的处理提供参考。
-
实验所用硝化污泥取自青岛市某污水处理厂消化池,具有良好的硝化性能。选取一定量的污泥,采用0.1 mol·L−1盐酸进行酸化处理,之后采用去离子水和PBS缓冲溶液各洗3次,以去除污泥中残留的
NH+4 -N,得到水泥复合物。以质量分数计,将10%的WPU与90%的水泥复合物混匀,依次加入10%的四甲基乙二胺,4%的过硫酸钾溶液(KPS)和3%的粉末活性炭,迅速搅拌均匀,约30 min后,混合液凝胶成固态,将固态胶体切割成3 mm×3 mm×3 mm的立方体,得到包埋固定化颗粒。制得的包埋颗粒为黑色,在水中呈均相,具有良好的物理强度和化学稳定性。包埋剂(WPU)和交联剂/引发剂(四甲基乙二胺、KPS)均为分析纯。图1为包埋颗粒的数码照片。 -
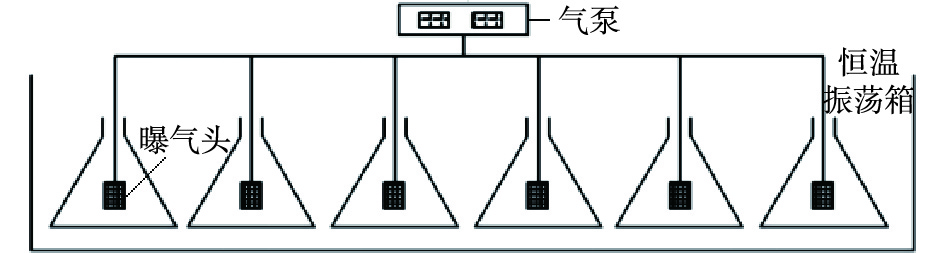
为研究不同盐度对包埋颗粒短程硝化反应过程的影响,采用6个250 mL的锥形瓶作为SBR进行批次实验,图2为实验反应装置。包埋颗粒投加率为10%,初始
NH+4 -N浓度为100 mg·L−1。反应器进水NaCl浓度分别为0、5、10、20、30、50 g·L−1,运行周期为12 h,包括进水5 min,出水5 min。以曝气砂头曝气,曝气量维持为0.1 L·min−1;通过NaHCO3调节pH至7.5~8.0;通过恒温振荡箱,将反应器温度控制在30 ℃。实验采用人工配水。周期结束后,检测出水ρ(NH+4 -N)、ρ(NO−2 -N)和ρ(NO−3 -N),分析氨氮去除率(ηA)、NO−2 -N积累率(RNAR)和硝酸盐积累率(R硝酸盐积累率)的变化情况,以判断不同盐度对短程硝化反应的影响。在SBR反应系统中,继续进行不同曝气时间下的批次实验,运行条件与不同盐度下的批次实验条件相同。每隔2 h取水样,测定一个SBR周期(12 h)内各形态氮(
NH+4 -N、NO−2 -N、NO−3 -N)含量的变化情况,并计算ηA、RNAR和R硝酸盐积累率。ηA、RNAR和R硝酸盐积累率分别按式(1)~式(3)计算。式中:ηA为氨氮去除率;ρ进水(
NH+4 -N)、ρ出水(NH+4 -N)分别为进、出水NH+4 -N浓度,mg·L−1;RNAR为亚硝态氮积累率;ρ出水(NO−2 -N)为出水NO2−-N浓度,mg·L−1;R硝酸盐积累率为硝酸盐积累率;ρ进水(NO−3 -N)、ρ出水(NO−3 -N)分别为进、出水NO3−-N浓度,mg·L−1。 -
实验采用连续流的升流式厌氧污泥床(up-flow anaerobic sludge bed,UASB)反应器,实验装置见图3。反应器由有机玻璃制成,有效容积为5 L,采用下部进水,上部出水的方式。包埋颗粒投加率为10%。反应器上部设有溢流堰,防止包埋颗粒随出水流出。以曝气砂头曝气,维持曝气量为0.1 L·min−1;通过加热棒维持反应器内温度为30 ℃,通过NaHCO3维持pH至7.5~8.0,HRT为8 h。周期结束后,检测出水ρ(
NH+4 -N)、ρ(NO−2 -N)和ρ(NO−3 -N)。 -
实验采用人工模拟废水,
NH+4 -N由NH4Cl提供。主要成分如下:NH4Cl 305 mg·L−1,NaCl 20.5 mg·L−1,KCl 9.6 mg·L−1,NaHCO3 468 mg·L−1,CaCl2·2H2O 9.6 mg·L−1,MgSO4·7H2O 33.6 mg·L−1,NaH2PO4·12H2O 46.4 mg·L−1。微量元素[22]添加量为1 mL·L−1,主要成分下:FeSO4·7H2O 1 g·L−1,MnCl2·4H2O 0.2 g·L−1,CuSO4·5H2O 0.1 g·L−1,CoCl2·6H2O 0.3 g·L−1,H3BO3 0.2 g·L−1,ZnSO4·7H2O 0.2 g·L−1,NiCl2·6H2O 0.2 g·L−1。 -
NH+4 -N:纳氏试剂分光光度法;NO−2 -N:N-(1-萘基)-乙二胺分光光度法;NO−3 -N:麝香草酚分光光度法[23];DO:JPBJ-608便携式溶解氧测定仪;pH:PHS-3C pH计。 -
取25 mL成功启动短程硝化反应器的包埋颗粒于250 mL锥形瓶中,加入模拟废水。以NH4Cl配制不同浓度的
NH+4 -N溶液,其余组分同表1模拟废水成分。将锥形瓶放入30 ℃恒温振荡培养箱中,120 r·min−1振荡培养。pH为7.5~8.0,维持曝气量为0.1 L·min−1,间隔8 h,取样测定水中NH+4 -N浓度,对包埋颗粒的基质动力学进行分析。 -
不同盐度对包埋颗粒短程硝化的影响。以硝化污泥制作包埋颗粒,按填充率10%投入反应器中。通过批次实验研究不同盐度对包埋颗粒短程硝化反应的影响,反应器进水
NH+4 -N浓度为100 mg·L−1,NO−2 -N和NO−3 -N浓度均为0 mg·L−1。图4为不同盐度系统中ηA、RNAR和R硝酸盐积累率的变化。向锥形瓶中投加NaCl后,ηA及RNAR均受到不同程度的影响。由图4(a)可知,由于NaCl的突然加入,系统内的微生物活性出现了短暂的下降,分析认为,可能是微生物不能马上适应在盐环境下生长,活性受到抑制。此时ρ(NaCl)为0 g·L−1的系统中ηA最高。运行一段时间后,包埋颗粒系统中的微生物开始适应盐环境。这可能是由于在长时间的选择与淘汰中,包埋颗粒系统中不能适应有盐环境的微生物受到抑制,能适应环境的菌群通过不断调节已经从被迫改变到积极适应[24],成为优势菌群[25],使处理效果有了进一步提高。从第4天开始,ρ(NaCl)为5 g·L−1和10 g·L−1的系统中,ηA开始呈现上升趋势,并分别在第9天和第8天超过ρ(NaCl)为0 g·L−1的系统;随着反应器的持续运行,ηA逐渐趋于稳定。其中ρ(NaCl)为0 g·L−1时,ηA始终维持在45%以上;ρ(NaCl)为5 g·L−1系统中,ηA在第9天后稳定在50%左右,ρ(NaCl)为10 g·L−1系统中,ηA在稳定运行后维持在55%以上;ρ(NaCl)为20 g·L−1的系统中,ηA也存在上升趋势,但低于0 g·L−1的系统;而ρ(NaCl)为30 g·L−1和50 g·L−1系统中,高盐环境使AOB和NOB的活性被抑制,氨氮去除受到影响,稳定运行后ηA仅为20%左右。
由图4(b)和图4(c)可知,ρ(NaCl)为0 g·L−1和5 g·L−1系统的RNAR基本为0%,而R硝酸盐积累率可达到40%以上,系统中的
NH+4 -N基本都经过硝化反应由NO−2 -N转化为NO−3 -N,为全程硝化反应。ρ(NaCl)为10 g·L−1系统中,RNAR呈现逐步上升的趋势,经过6 d的运行之后,RNAR达到90%,而R硝酸盐积累率趋于0%。ρ(NaCl)为20 g·L−1的系统中,RNAR稳定后可达到80%以上。ρ(NaCl)为30 g·L−1和50 g·L−1系统中,RNAR较低,且出水NO−3 -N浓度基本为0 g·L−1,表明系统内NOB的活性受到了抑制,不能完成将NO−2 -N氧化为NO−3 -N的过程。由此可见,盐度可作为脱氮系统中脱氮种群的抑制剂,在长期运行条件下,NOB菌群的活性被抑制,实现AOB菌群的优化,使短程硝化过程得以实现;但是盐度过高也会抑制AOB的活性。在不同的盐度条件下,AOB和NOB受到的抑制程度不同;在相同的盐度条件下,AOB比NOB有更强的耐盐性。叶柳[26]通过微生物计数实验(MPN法)发现,虽然盐度对AOB和NOB都有抑制作用,但2种菌群对盐度的适应能力不同,AOB的耐盐能力要强于NOB;雷中方[27]通过实验证明,废水中含盐量的增加,可能导致微生物的正常代谢功能遭到破坏;CUI等[28]的研究表明,无机盐对NOB有较强的抑制能力,可以通过向普通废水中投加无机盐实现短程硝化。通过分析可知,当盐度超过10 g·L−1时,NOB的活性受到抑制;超过20 g·L−1时,盐度的升高使细胞渗透压增大[29],不利于细胞内底物和氧的传递,导致AOB的活性也受到抑制。因此,10 g·L−1 NaCl浓度下短程硝化效果较好,可作为包埋系统短程硝化反应器的启动盐度。
曝气时间对包埋颗粒短程硝化稳定性的影响。通过控制盐度运行短程硝化反应器后,继续进行批次实验,研究曝气时间对包埋颗粒短程硝化稳定性的影响。测得系统中不同曝气时间下各系统出水三氮浓度、ηA和RNAR的变化,如图5所示。实验进水 NH+4 -N浓度为100 mg·L−1,曝气量为0.1 L·min−1,系统内DO浓度小于1 mg·L−1。由分析可知,ρ(NaCl)超过20 g·L−1时AOB和NOB活性均受到抑制,系统短程硝化效果受到影响,因此,ρ(NaCl)为30 g·L−1和50 g·L−1系统不再进行稳定性实验。由图5可见,ρ(NaCl)为0 g·L−1的系统中,ηA呈现先上升后下降的趋势,曝气时间为8 h时达到最大值47%,随后开始下降;RNAR随曝气时间的增加而下降,基本趋于0%。ρ(NaCl)为5 g·L−1时,ηA在8 h时达到最大值41%,而RNAR仅为3.04%。ρ(NaCl)为10 g·L−1,曝气时间为8 h时系统的短程硝化效果最好,此时系统出水ρ(
NH+4 -N)为42 mg·L−1,ηA可达到56%;出水ρ(NO−2 -N)为50 mg·L−1,RNAR达到96%。ρ(NaCl)为20 g·L−1时,ηA和RNAR分别在曝气时间为8 h时达到50%和90%,短程硝化效果低于ρ(NaCl)为10 g·L−1的系统。由图5可知,各系统在不同曝气时间下包埋颗粒短程硝化的效果是不同的。进水ρ(NaCl)为10 g·L−1,曝气时间为8 h时RNAR最高,短程硝化稳定性最好。虽然在低DO环境下,AOB对氧的亲和力大于NOB,但当曝气时间超过8 h时,NOB会逐渐适应低氧环境,且大量
NO−2 -N的存在为NOB提供了基质[30]。如不控制曝气时间,NO−2 -N会在NOB的作用下向NO−3 -N转化,导致RNAR下降,短程硝化转化为全程硝化。因此,维持短程硝化反应需要保持一定的曝气时间,以实现NO−2 -N的积累。 -
由批次实验可知,10 g·L−1 NaCl浓度可用于启动包埋颗粒短程硝化反应器。图6为连续流UASB反应器运行期间进、出水中
NH+4 -N、NO−2 -N、NO−3 -N的浓度变化。进水NH+4 -N浓度为100 mg·L−1,NO−2 -N和NO−3 -N浓度均为0 mg·L−1。可以看出,包埋颗粒从第1天开始就出现NH+4 -N的去除,ηA为34.82%,表现出良好的性能。但系统的短程硝化效果不稳定,第11天时,ηA仅达到11.70%,原因可能是,包埋颗粒在反应器内分布不均匀,导致出水水质受到影响。第15天向系统内投加10 g·L−1 NaCl,经过一段时间的盐度驯化后,包埋颗粒开始适应盐环境。第20天开始,出水中ρ(NH+4 -N)明显降低,并逐渐趋于稳定,ηA可稳定在50%以上。出水中ρ(NO−2 -N)从第20天开始逐渐升高,RNAR由第11天的10.25%增加到第50天的81.03%。由此可见,NaCl的加入抑制了NOB的活性,使NO−2 -N转化为NO−3 -N的过程受到影响,出水中ρ(NO−2 -N)升高而ρ(NO−3 -N)逐渐降低,RNAR增大。由分析可知,10 g·L−1 NaCl的加入初步实现了连续流UASB包埋颗粒短程硝化反应器的启动。盐度对短程硝化反应的影响在目前研究中的结果并不一致,就现有的研究[31]看,不同的工艺类型以及接种的生物来源(盐度驯化种群、非盐度驯化种群、或者耐盐种群)和处理废水类型(在大多数研究中采用人工配水)可能是造成目前研究很不一致的重要因素。王淑莹等[25]采用全程硝化反硝化研究SBR工艺的脱氮效果发现,7.5 g·L−1的NaCl浓度可以作为活性污泥系统短程硝化反硝化的启动盐度;一定浓度的NaCl盐度可以作为一种化学抑制剂实现对硝化菌的选择性抑制。崔有为等[32]的实验结果表明,采用MUCT工艺处理低含盐量的生活污水,在不超过8 g·L−1的盐度环境内,生物可以实现良好的处理效率,亚硝酸盐积累明显;叶柳等[24]在使用MUCT工艺处理含盐生活污水时,稳定盐度在10 g·L−1后,在好氧段实现了高的亚硝酸盐积累,从而通过回流至缺氧段实现了短程脱氮。由此可见,在不同的运行工艺条件下,盐度对短程硝化反应会造成不同的影响。此外,有研究[32]表明,不同功能菌群对盐度耐受能力与盐度实施的选择作用有关,不同种属的微生物或不同功能的微生物耐盐能力有差异,这种差异表现在应对盐度抑制时不同的调节能力上。
-
选用成功启动短程硝化反应器的包埋颗粒,在10 g·L−1盐度、8 h的曝气时间下进行动力学实验,以测定包埋颗粒的基质动力学特性。包埋颗粒的短程硝化动力学过程受
NH+4 -N浓度的限制,在处理过程中,高氨氮环境会形成较高浓度的游离氨(FA)[33]。FA是AOB的生长基质,高浓度FA会对AOB的氨氧化过程产生抑制;且NOB对FA有潜在的适应性,在反应器运行后期,FA对NOB的抑制会大幅降低[34]。因此,研究NH+4 -N对包埋颗粒短程硝化的基质抑制特性有利于进一步强化短程硝化效果。假设在实验过程中氨氧化菌细胞数量不发生变化,且反应器内始终处于完全混合状态,那么可用Haldane模型[35]对包埋颗粒短程硝化的动力学特性进行描述,其计算公式见式(4)。式中:v为反应速率,kg·(kg·d)−1;vmax为最大反应速率,kg·(kg·d)−1;S为基质浓度,mmol·L−1;Km为半速率常数,mmol·L−1;Ki为半抑制常数,mmol·L−1。实验测得,包埋颗粒短程硝化速率与基质浓度的对应关系见表1。
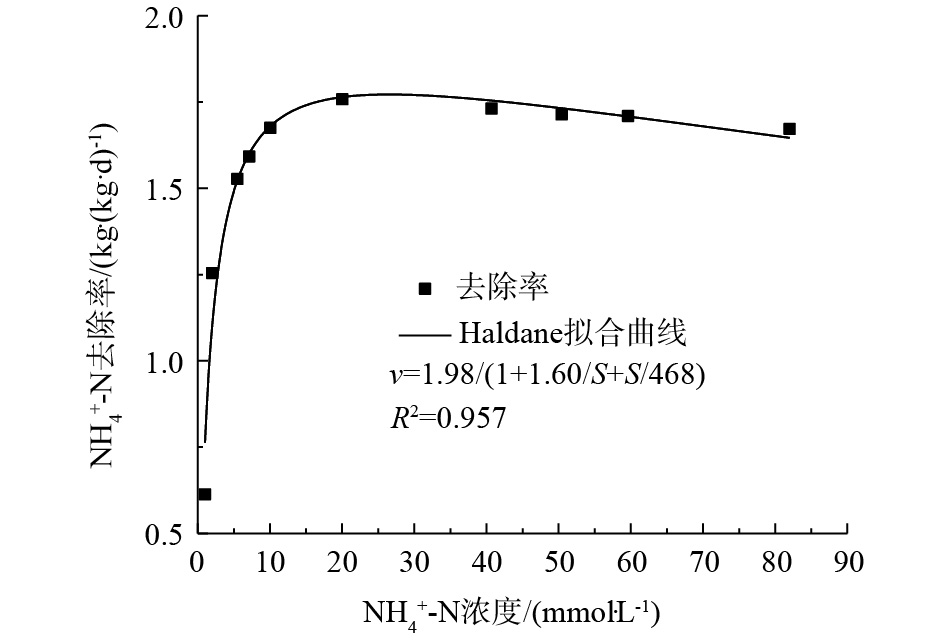
通过origin软件,以Haldane模型对表1中实验数据进行非线性拟合,得到包埋颗粒对NH4+-N的动力学方程。由基质浓度降解曲线,计算出包埋颗粒的最大氨反应速率(SAA-
NH+4 -N)[36]。由图7可知,包埋颗粒最大氨反应速率(SAA-NH+4 -N)为1.98 kg·(kg·d)−1,对氨的半速率常数为1.60 mmol·L−1,氨的半抑制常数为468 mmol·L−1,R2为0.957,表现出良好的相关性。较高的反应速率可以促进基质的转化,而包埋颗粒对氨的半速率常数较小,表明其对基质的亲和力较大,利于短程硝化反应的进行[18]。因此,10 g·L−1盐度系统中的包埋颗粒具有优良的动力学特性,有助于短程硝化反应器的高效运行。 -
1)研究表明,盐度对包埋颗粒短程硝化反应器的启动存在一定的影响。在NaCl浓度为10 g·L−1的盐度系统中,硝化污泥包埋颗粒的ηA和RNAR最高,分别可达到55%和90%,成功实现了包埋颗粒的短程硝化反应;控制曝气时间可以保证包埋颗粒短程硝化过程的稳定运行,进水ρ(NaCl)为10 g·L−1,曝气时间为8 h时系统的短程硝化效果最好,ηA可达到56%,RNAR达到96%。
2)在NaCl浓度为10 g·L−1,曝气时间为8 h的条件下可实现包埋颗粒UASB反应器的初步启动,ηA和RNAR分别可达到50%和80%以上;基质动力学研究表明,包埋颗粒对氨氮的动力学特性符合Haldane基质抑制动力学模型,包埋颗粒具有优良的动力学特性。此项研究可为含盐废水的氨氮处理提供参考。
盐度和曝气时间对包埋颗粒短程硝化启动的影响及其动力学分析
Effect of salinity and aeration time on the start-up of partial nitrification of immobilized particles and its kinetics analysis
-
摘要: 针对短程硝化反应器启动时间长、效果不稳定等问题,使用水性聚氨酯(WPU)制作硝化污泥包埋颗粒,利用SBR设置盐度梯度以及不同曝气时间进行批次实验,启动短程硝化;通过控制最佳反应条件启动UASB短程硝化反应器,同时进行动力学分析。结果表明:在批次实验过程中,随着盐度的增加,氨氮去除率(ηA)及
NO−2 -N积累率(RNAR)先上升后下降。当NaCl浓度为10 g·L−1时,短程硝化效果最佳,ηA为55%,RNAR为90%;不同曝气时间对短程硝化的稳定性有较大的影响,曝气时间为8 h时短程硝化效果最稳定,ηA和RNAR分别达到56%和96%。控制NaCl浓度为10 g·L−1,HRT为8 h,成功实现了UASB短程硝化反应器的启动;包埋颗粒对氨氮的动力学特性符合Haldane基质抑制动力学模型,具有优良的动力学特性。研究可为包埋颗粒与短程硝化工艺的耦合脱氮提供参考,并为含盐废水的处理提供技术支持。Abstract: In view of the long start-up time and unstable effect of partial nitrification reactor, waterborne polyurethane (WPU) was used to prepare the immobilized particles of nitrification sludge. The partial nitrification was started up through batch tests in the SBR reactor at different salinity gradients and different aeration times. The UASB partial nitrification reactor was started up by controlling the optimum conditions, and the kinetic analysis was performed simultaneously. The results showed that the ammonia nitrogen removal rate (ηA) andNO−2 -N accumulation rate (RNAR) increased firstly and then gradually decreased as the salinity increased in batch tests. The partial nitrification effect reached the best when the NaCl concentration was 10 g·L−1, and ηA and RNAR were 55% and 90%, respectively. Different aeration time had a great influence on the stability of partial nitrification. The partial nitrification effect was the most stable when the aeration time was 8 h, and ηA and RNAR reached 56% and 96%, respectively. The UASB reactor was successfully started up by controlling the NaCl concentration at 10 g·L−1 and HRT at 8 h. The kinetics of the immobilized granules on ammonia nitrogen was in accordance with the Haldane matrix inhibition kinetics model, which indicated that the immobilized granules had excellent kinetic characteristics. The study can provide a reference for the coupling denitrification of immobilized granules and partial nitrification processes, and provide technical support for saline wastewater treatment.-
Key words:
- immobilized granules /
- partial nitrification /
- salinity /
- start-up /
- aeration time /
- kinetic analysis
-
在国务院“水十条”对矿井水的最新定位下,煤炭开采形成的矿井水资源保护与综合利用成为实现我国煤炭工业绿色发展和生态文明建设中的重大关键问题[1]. 据统计我国吨煤开采产生矿井水为1.87 m3,矿井水产出量为每年6.88×109 m3,平均利用率约为35%[2-3],主要原因是矿井水水质差,导致不能直接利用,需要进行不同程度的预处理或深度处理,而高额的处理成本限制了综合利用[4-7].
近年来,国内外学者在地下水、地表水的水化学特征方面取得了一系列成果[8-12], 如探讨了神东矿区的高氟矿井水分布特征及形成机制[13];研究了辛置井田地下水的水化学特征和水-岩作用机理[14];系统分析了伊敏矿区地下水的水化学特征与其控制因素[15];对郭家湾煤矿井田内不同区域水化学特征进行差异性分析和水质综合评价[16];研究了高铁锰矿井水的水化学特征与其净化机制[17]. 研究主要侧重矿区水文地球化学特征及形成机制,而对干旱区矿井水综合利用于农田灌溉的相关评价相对较少.
我国西部煤炭资源丰富,占全国总量的70%以上,水资源匮乏,生态环境脆弱[18-20]. 在国家“以水定产,以环境承载力定产”煤炭工业发展理念下,矿井水综合利用成为西部矿区高质量快速发展的卡脖子问题. 因此,在地处干旱半干旱区的煤矿矿井水综合利用相关研究显得尤为重要. 新上海一号煤矿地处毛乌素沙漠边缘,属半干旱半沙漠大陆性气候,干旱少雨[21]. 矿井水若能有效的综合利用于农田灌溉,可有效降低矿井水综合利用成本,实现社会-经济-环境的高质量协同发展.
本研究拟对以新上海一号井田为例,针对各含水层矿井水形成作用进行研究,并对各含水层矿井水的灌溉适宜性进行评价,可为干旱半干旱区矿井水综合利用工作提供依据.
1. 材料与方法(Materials and methods)
1.1 研究区概况
研究区为新上海庙一号矿井田,其自上而下地层主要有:新生界第四系(Q)和古近系(E);白垩系志丹群(K1zd);侏罗系直罗组(J2z);含煤岩系侏罗系延安组(J2y);三叠系延长组(T3y),其赋存的含水层依次命名为:新生界、白垩系、直罗组、煤系间、宝塔山、三叠系含水层[22]. 研究区内有21条断层,其中F2、FD5和DF20是导水断层.
研究区内井筒掘进和工作面推进过程中的直接充水水源为直罗组、煤系间、延安组和三叠系含水层,间接充水水源为新生界含水层. 为实现矿井水防控,在采掘活动前需要对相应充水含水层进行疏放工程,因此以上述充水含水层为对象进行水化学特征和矿井水综合利用模式研究.
1.2 样品采集与测试
采样分为4个阶段:2006年煤层顶板含水层勘探阶段;2012年近煤层含水层水文地质补充勘探阶段;2016年和2019年各个含水层水文地质补充勘探阶段. 各主要含水层共采集55组样品,检测项目包括K++Na+、Ca2+、Mg2+、Cl−、SO42−、HCO3−、CO32−、pH和TDS,取样点信息详见表1.
表 1 研究区采样点信息表Table 1. The mine water sampling points of the study area采样时间Sampling time 采样含水层Sampling aquifer 采样钻孔编号Sampling borehole number 样品组数Number of samples 2006 新生界 W1、W3、W7、W11、W17、W21、W22、W25、W28、W32 10 白垩系 1604、1202 2 煤系间 2403(上段)、1602(上段)、1202、2403(下段)、1602(下段) 5 2012 白垩系 Z1、Z8 2 直罗组 Z1、Z2、Z3、Z8、Z10 5 煤系间 Z4、Z5、Z6、Z7、Z12、Z13、Z14、Z16 8 2016 白垩系 B3、B5、B9 3 直罗组 B10 1 煤系间 B1、B13、B35、B38 4 宝塔山 B2、B4、B6、B7、B8、B12、B14、 7 三叠系 B36 1 2019 宝塔山 B15、B37、B44、B45、B47 5 三叠系 B39、B46 2 合计 55 | Show Table DownLoad:
CSV
DownLoad:
CSV
1.3 数据分析方法
利用Matlab对矿井水水化学数据进行整理统计和计算,利用Piper图分析地下水水化学类型,利用SPSS软件进行各离子与TDS的相关性分析,通过Gibbs图、氯碱指数图、脱硫酸系数图、离子比值图、水力联系度及矿井水盐碱害分类探讨矿井水水化学特征成因及灌溉适宜性.
2. 结果与讨论(Results and discussion)
2.1 水化学分析
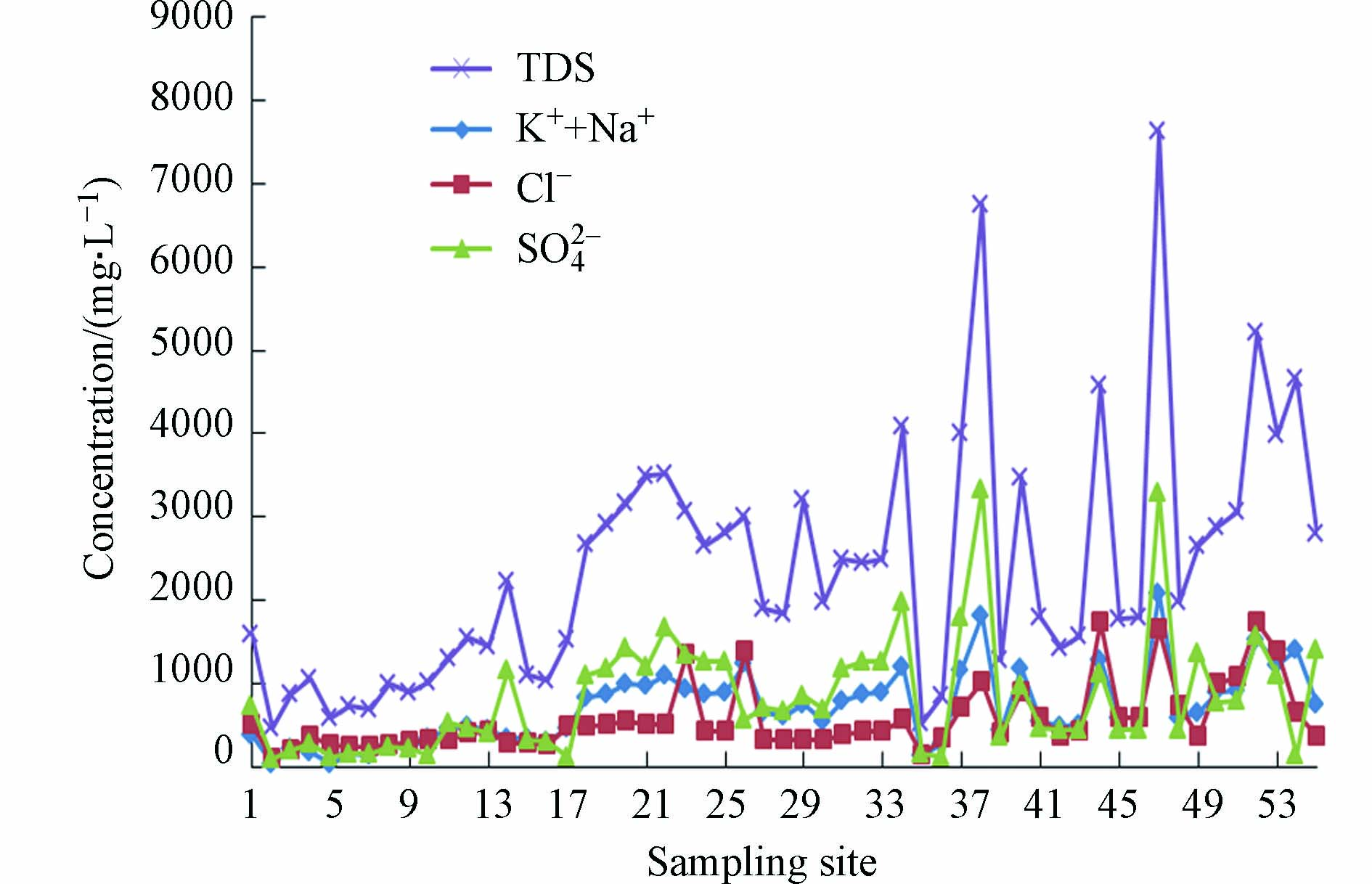
根据研究区内所采集的矿井水样检测结果分析可得,各含水层矿井水的pH值为7.58—12.06,均呈碱性. TDS为466.00—7624.90 mg·L−1,除了新生界,其余均为高矿化度矿井水,而且从新生界到三叠系,埋深越大,越不易接受第四系含水层的补给,因此TDS逐渐增大. 水中主要离子组成变化及不同水样化学组成类型特征采用Piper三线图(图1)分析[23],K++Na+、SO42−和Cl−明显占优势. 由白垩系含水层至三叠系含水层矿井水化学类型由SO4·Cl-Na逐渐过渡为Cl SO4-Na,新生界含水层水化学类型较为多样.
2.2 相关性分析
TDS可以来描述离子特征,利用SPSS软件计算得到Pearson相关性系数矩阵(见表2),可以看出除pH外,TDS与各离子均呈显著相关性[24]. 根据相关性分析结果,TDS与K++Na+、SO42−和Cl−呈显著正相关(P<0.01),相关系数分别为0.973、0.868和0.757,从图2可知,TDS、K++Na+、SO42−和Cl−变化趋势相同,说明K++Na+、SO42−和Cl−是TDS的主要来源.
表 2 TDS与各离子之间的相关系数矩阵Table 2. Correlation coefficient matrix among TDS and ions项目 Item K++Na+ Ca2+ Mg2+ Cl− SO42− HCO3− CO32− pH TDS K++Na+ 1.000 Ca2+ 0.481 1.000 Mg2+ 0.353 0.178 1.000 Cl− 0.778** 0.433 0.555* 1.000 SO42− 0.820** 0.322 0.433 0.526* 1.000 HCO3− 0.292 −0.080 0.075 0.331 0.082 1.000 CO32− 0.246 0.619 −0.083 0.073 −0.146 0.070 1.000 pH −0.184 −0.057 −0.177 −0.207 −0.261 −0.187 0.078 1.000 TDS 0.973** 0.580* 0.389 0.757** 0.868** 0.174 0.213 −0.216 1.000 *表示在置信度0.05时相关性显著;**表示在置信度0.01时相关性显著. | Show TableDownLoad:
CSV
2.3 水化学形成作用
2.3.1 水化学演化特征
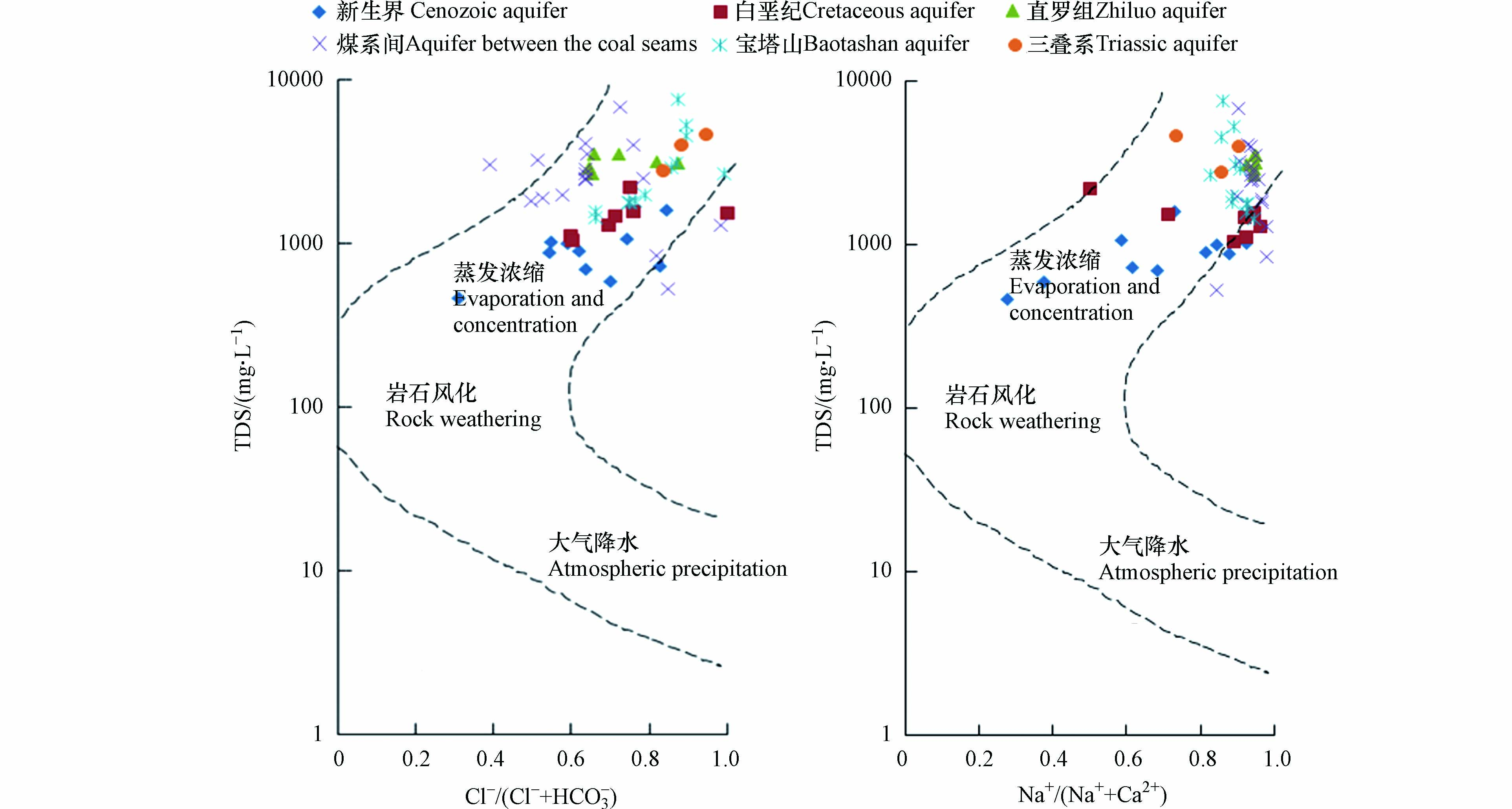
Gibbs可有效揭示水化学组分的形成机制,在水化学分析中得到广泛应用[25]. Gibbs图横坐标为阴离子或阳离子质量浓度比,纵坐标为TDS质量浓度,判别方法见表3.
表 3 基于Gibbs图的水化学组成形成作用判别方法[26]Table 3. Determination method of formation of water chemical composition based on Gibbs diagram[26]判别指标 Discriminative index 判别依据 Discrimination on the basis of 阴离子质量浓度比值Cl−/(Cl−+HCO3−) 0.5—1 <0.5 0.5—1 阳离子质量浓度比值Na+/(Na++Ca2+) 0.5—1 <0.5 0.5—1 TDS 较高(>1000) 中等(75—1000) 较低(<75) 主要形成作用 蒸发浓缩作用 岩石风化作用 大气降水作用 | Show TableDownLoad:
CSV
从图3可知,研究区内绝大部分水样分布在Gibbs图右上部,即TDS在1000—7625 mg·L−1之间,Cl−/(Cl−+HCO3−)或Na+/(Na++Ca2+)接近1,说明矿井水的水化学组分主要受蒸发浓缩影响.
2.3.2 阳离子交替吸附作用
阳离子交替吸附作用是由于Ca2+、Mg2+吸附能力较大,可置换岩土颗粒表面的Na+,使其在水中的含量降低[27]. 为了定量分析其作用的方向与强度,可以绘制研究区矿井水氯碱指数图,其中横坐标为CAI1,纵坐标为CAI2,计算公式如下:
CAI1=c(Cl−)−c(Na++K+)c(Cl−) (1) CAI2=c(Cl−)−c(Na++K+)2×c(SO42−)+c(HCO3−) (2) 式中,c表示毫摩尔浓度,单位为mmol·L−1. 若CAI1和CAI2均<0,则阳离子交替吸附作用较高;反之,若CAI1和CAI2均>0,则阳离子交替吸附作用较差[27].
从图4可知,各含水层矿井水均存在不同程度的阳离子吸附作用,由新生界至三叠系含水层,氯碱指数逐渐减小,随着阳离子交替吸附作用不断加强,Ca2+、Mg2+含量不断下降.
2.3.3 脱硫酸作用
脱硫酸作用指的是水中的SO42−被还原,导致水中的SO42−含量下降,同时HCO3−含量升高的过程,其主要化学方程式为:
SO42−+2C+2H2O→H2S+2HCO3− 脱硫酸作用强度的主要评价指标是脱硫酸系数c(SO42−)/c(Cl−)[28],脱硫酸系数图中横坐标为SO42−与Cl−的摩尔浓度比,纵坐标为HCO3−的摩尔浓度. 从图5可知,研究区各含水层矿井水c(SO42−)/c(Cl−)数值较小,一般小于0.5,各个含水层存在不同程度的脱硫酸作用. 其中煤系间和直罗组含水层c(SO42−)/c(Cl−)一般为0.5—1.5,其受脱硫酸作用的影响较小. HCO3−在图中相关性不明显,说明HCO3−的唯一来源并不是脱硫酸作用.
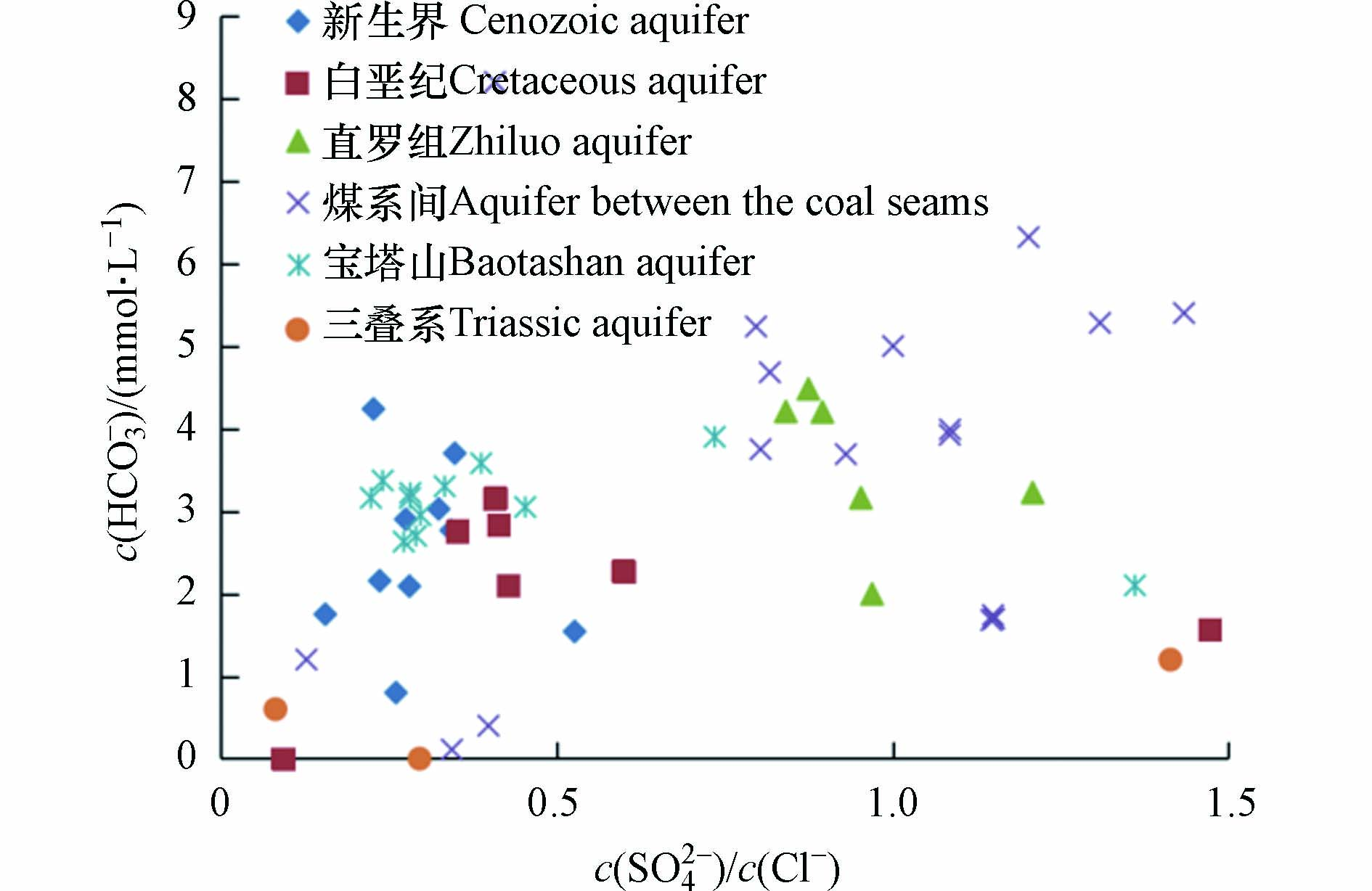 图 5 各含水层矿井水的脱硫酸系数图Figure 5. Desulphidation coefficient of mine water from each aquifer
图 5 各含水层矿井水的脱硫酸系数图Figure 5. Desulphidation coefficient of mine water from each aquifer2.3.4 离子比值分析
离子比值分析是用于揭示水化学组成及离子来源常用的方法[29]. 岩盐溶解产生的Na+和Cl−物质量的比值为1,所以c(Na+)/c(Cl−)常被用来解释水中Na+的来源[30].
从图6可知,研究区新生界含水层水样的c(Na+)/c(Cl−)等于1,其他含水层水样的比值大于1. 以上情况说明,新生界含水层水样中Na+的唯一来源是钠盐溶解;其他含水层水样中相对Cl−偏高的Na+可能与硅酸盐的风化有关(2NaAlSi3O8+2CO2+11H2O→Al2Si2O5(OH)4+2HCO3−+2Na++4H4SiO4),同时存在其他含钠矿物的溶解[31]. 井田内含水层岩性以砂岩为主,矿物成分多为长石等硅酸盐类矿物,Na+偏高的原因可能是硅酸盐类矿物的溶解作用和阳离子交替吸附作用,使水中的Ca2+和Mg2+被吸附,释放的Na+也是导致Na+含量升高的一个原因. 当水中c(Ca2++Mg2+)/c(SO42−+HCO3−)=1时,代表Ca2+和Mg2+主要来源于碳酸盐和硫酸盐的溶解[32-35].
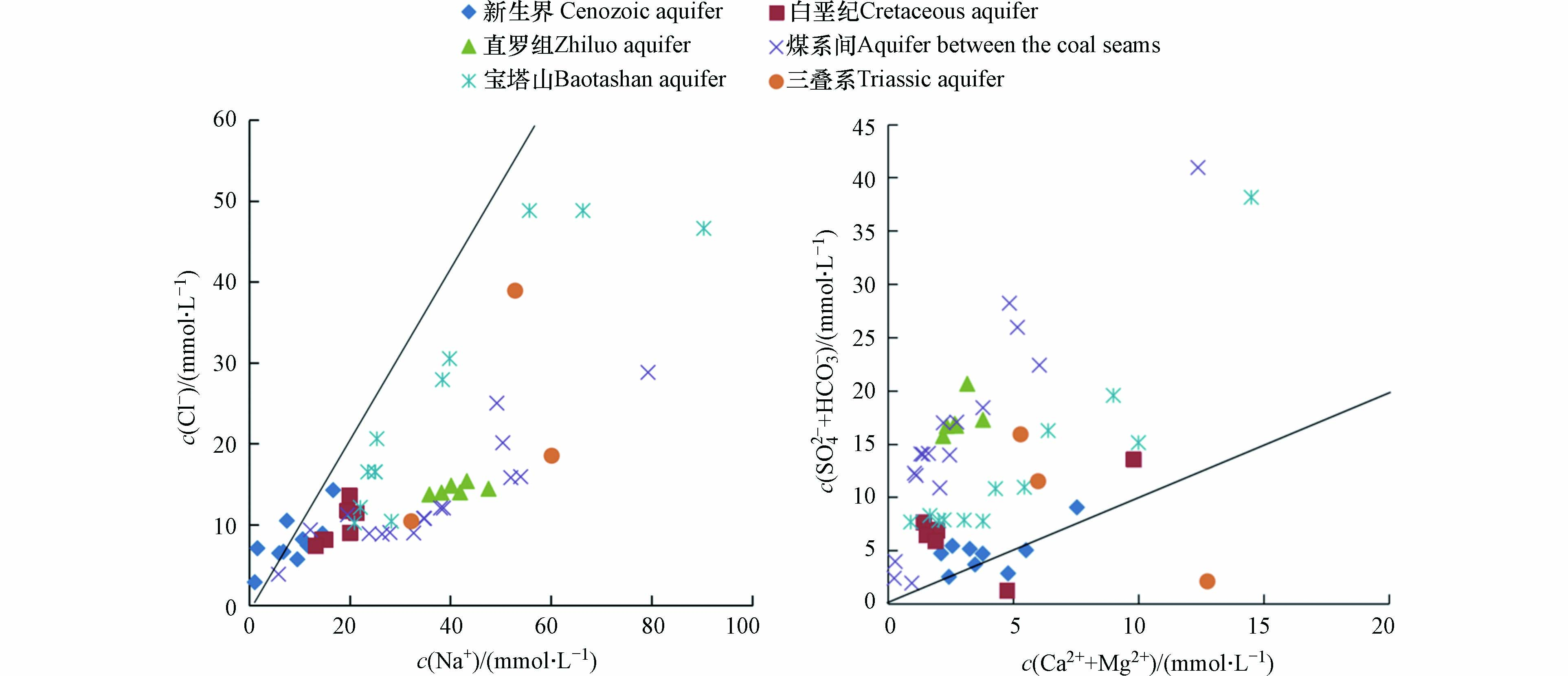 图 6 各含水层矿井水的Cl−-Na+和(Ca2++Mg2+)-(SO42−+HCO3−)比值图Figure 6. Cl−-Na+ ratios and (Ca2++Mg2+)-(SO42−+HCO3−)ratios of mine water from each aquifer
图 6 各含水层矿井水的Cl−-Na+和(Ca2++Mg2+)-(SO42−+HCO3−)比值图Figure 6. Cl−-Na+ ratios and (Ca2++Mg2+)-(SO42−+HCO3−)ratios of mine water from each aquifer从图6可知,研究区内仅有新生界含水层水样c(Ca2++Mg2+)/c(SO42−+HCO3−)基本等于1,其他含水层水样小于1. 说明新生界含水层水样中的Ca2+和Mg2+来自于碳酸盐及其溶解作用,其他含水层水样中的Ca2+和Mg2+来源有限(含水层多为砂岩),煤系间及其顶底板含水层存在大量的S2−氧化也会导致SO42−增加.
相关研究表明,研究区内除新生界之外的深部含水层相对封闭,径流缓慢,水岩作用较弱,其离子含量较高,矿化度较大[36]. 矿井水的水化学组分主要受蒸发浓缩作用影响,同时阳离子交替吸附作用随着埋深增大不断加强,Ca2+、Mg2+含量不断下降,并受到不同程度脱硫酸作用的影响.
2.3.5 含水层之间水力联系
从前面的分析可知Cl−是研究区各含水层中含量较高的离子,由于其浓度一般仅受到含水层的影响,受其他因素影响较小,故可以利用各含水层的Cl−浓度来判断含水层之间的水力联系紧密程度. 含水层间水力联系度K计算公式为:
K=0.5×Cl−1−Cl−2Cl−1+Cl−2 (3) 式中,Cl−1和Cl−2分别代表不同含水层中Cl−的平均质量浓度.
当K<0.2,表示两个含水层之间水力联系强;当0.2<K<0.4,说明两个含水层之间水力联系中等;当K>0.4,说明两个含水层之间水力联系弱. 根据研究区水化学数据,分别计算了6个含水层之间的水力联系度. 由表4可知,研究区个含水层之间水力联系度K均小于0.2,说明各含水层之间均存在紧密的水力联系,这与研究区东西两侧的F2和DF20断层具有良好的导水性有关.
表 4 各含水层之间水力联系程度Table 4. Hydraulic connection between each aquifer含水层Aquifer Cl−平均浓度/(mg·L−1)Mean concentration of Cl− 水力联系度KDegree of hydraulic connection K 水力联系程度Degree of hydraulic connection 新生界 275.76 — — 白垩系 351.36 0.06(与新生界) 强 直罗组 564.66 0.12(与白垩系) 强 煤系间 517.10 0.02(与直罗组) 强 宝塔山 902.28 0.14(与煤系间) 强 三叠系 801.18 0.03(与宝塔山) 强 | Show TableDownLoad:
CSV
相关研究表明,研究区内断层附近的各含水层之间存在水力联系[36],这也导致其水文地球化学特征与周边同层含水层具有显著差异.
2.4 灌溉适宜性评价
新上海一号井田地处干旱半干旱地区,水资源相当匮乏. 通过上述综合分析可知,由于矿井水的矿化度高,深度处理成本高,不适宜综合利用于生活饮用水. 随着矿井水综合利用于农田灌溉的需求越来越大,迫切需要开展矿井水灌溉适宜性评价方法研究.
灌溉水源直接影响农作物生长的评价指标是盐碱害[37]. 盐害常用电导率(EC)表征[38],碱害常用钠吸附比(SAR)和钠百分含量(SC)表征(表5),其计算公式(4)—(5)[39],公式中离子浓度单位均为mmol·L−1.
参数Parameter 分级标准Classification standard 参数Parameter 分级标准Classification standard 参数Parameter 分级标准Classification standard EC/(μs·cm−1) <250 适宜(C1) SAR/(mmol·L−1)1/2 <10 非常适宜(S1) SC/% <20 非常适宜 250—750 允许的(C2) 10—18 适宜(S2) 20—40 适宜 750—2250 不确定(C3) 18—26 允许的(S3) 40—60 允许的 >2250 不适宜(C4) >26 不适宜(S4) 60—80 不确定 >80 不适宜 注:C代表盐化级别,S代表碱化级别,1、2、3、4分别代表低、中、高、极高四个等级. | Show TableDownLoad:
CSV
SAR=c(Na+)+c(K+)√c(Ca2+)+c(Mg2+) (4) SC=c(Na+)+c(K+)c(Na+)+c(K+)+2×c(Ca2+)+2×c(Mg2+)×100 (5) 根据表5灌溉水水质分类标准可以进行矿井水灌溉的适宜性评价. 从图7(a)可知,研究区内矿井水的EC值和SAR值普遍较大,45%的矿井水EC值大于2250,位于盐害极高(C4)范围内;70%的矿井水SAR值大于18,位于碱害极高(S3)范围内;10%的矿井水SAR值大于26,位于碱害极高(S4)范围内;即研究区内除新生界和白垩系矿井水的盐碱害较严重(C4S3或C4S4区域),未经深度处理不适宜用于农田灌溉. 从图7(b)可知,73%的矿井水中SC值大于80,不适宜直接用于农田灌溉.
从图7可知,整体而言自上而下含水层的矿井水灌溉适宜性逐渐降低,新生界和白垩系含水层的矿井水基本适宜农田灌溉;而直罗组、宝塔山和三叠系含水层的矿井水适宜性一般为不确定-不适宜,不适宜直接用于农田灌溉,若达到农田灌溉水质要求,其矿井水深度处理成本相对较高. 因此,针对井田内矿井水的综合利用应采取“分层分质综合利用”的原则,可有效降低矿井水综合利用成本,实现社会-经济-环境的高质量协同发展.
3. 结论(Conclusion)
(1)研究区阳离子以K++Na+为主,阴离子以SO42−和Cl−为主,新生界含水层水化学类型较为多样,由白垩系含水层至三叠系含水层矿井水化学类型由SO4·Cl-Na逐渐过渡为Cl·SO4-Na;TDS与K++Na+、SO42−和Cl−相关性最为显著,相关系数分别为0.973、0.868和0.757.
(2)研究区矿井水的水化学组分主要受蒸发浓缩影响,各含水层矿井水均存在不同程度的阳离子吸附作用,由新生界至三叠系含水层,阳离子交替吸附作用不断加强;新生界、白垩系、宝塔山和三叠系含水层受到不同程度脱硫酸作用影响,煤系间和直罗组含水层矿井水受脱硫酸作用影响较小.
(3)新生界含水层矿井水中Na+的唯一来源是岩盐溶解;其他含水层矿井水中相对Cl−偏高的Na+可能受硅酸盐的风化和其他含钠矿物的溶解影响;新生界含水层矿井水中的Ca2+和Mg2+来自于碳酸盐和硫酸盐的溶解,其他含水层矿井水中的Ca2+和Mg2+来源有限,煤系间及其顶底板含水层矿井水存在大量的S2−氧化也会导致SO42−增加;各含水层之间存在紧密的水力联系,这与断层具有导水性有关.
(4)研究区内45%的矿井水EC值大于2250,70%的矿井水SAR值大于18,73%的矿井水中SC值大于80,大部分矿井水不适宜直接用于农田灌溉. 新生界和白垩系含水层的矿井水基本适宜农田灌溉,而直罗组、宝塔山和三叠系含水层的矿井水不适宜用于农田灌溉,因此研究区内矿井水宜实施“分层分质综合利用”.
-
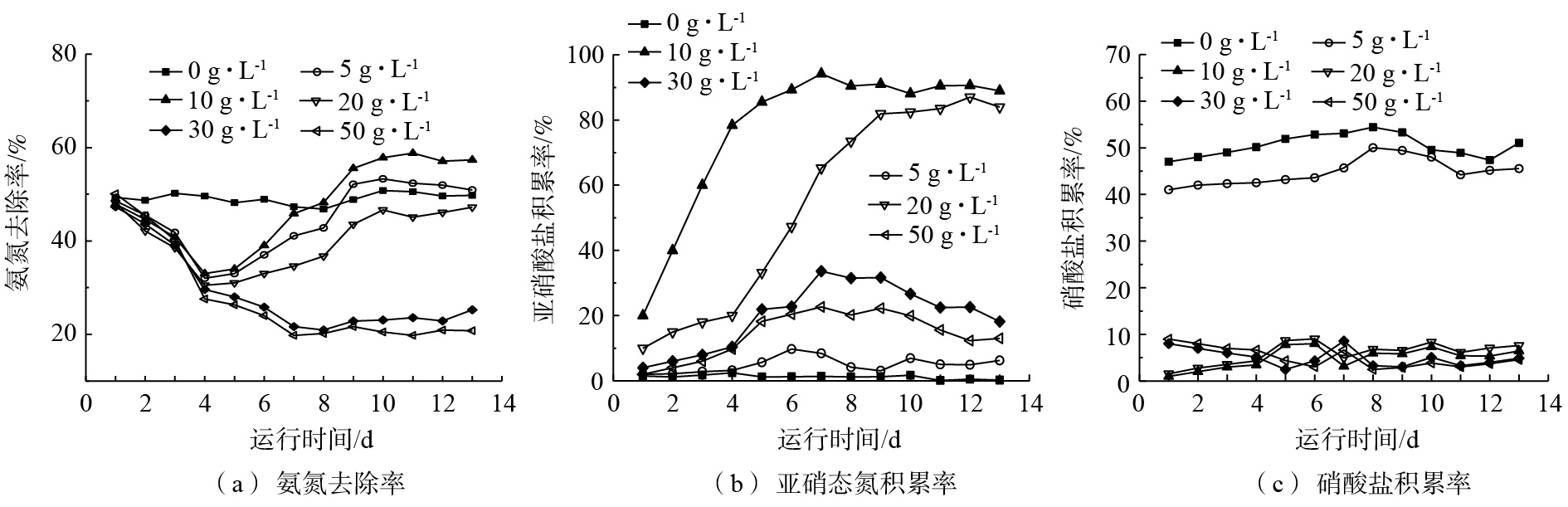
图 4 不同盐度下氨氮去除率、亚硝态氮积累率和硝酸盐积累率的变化
Figure 4. Effect of different salinity on nitrogen removal rate, nitrite accumulation rate and nitrate accumulation rate
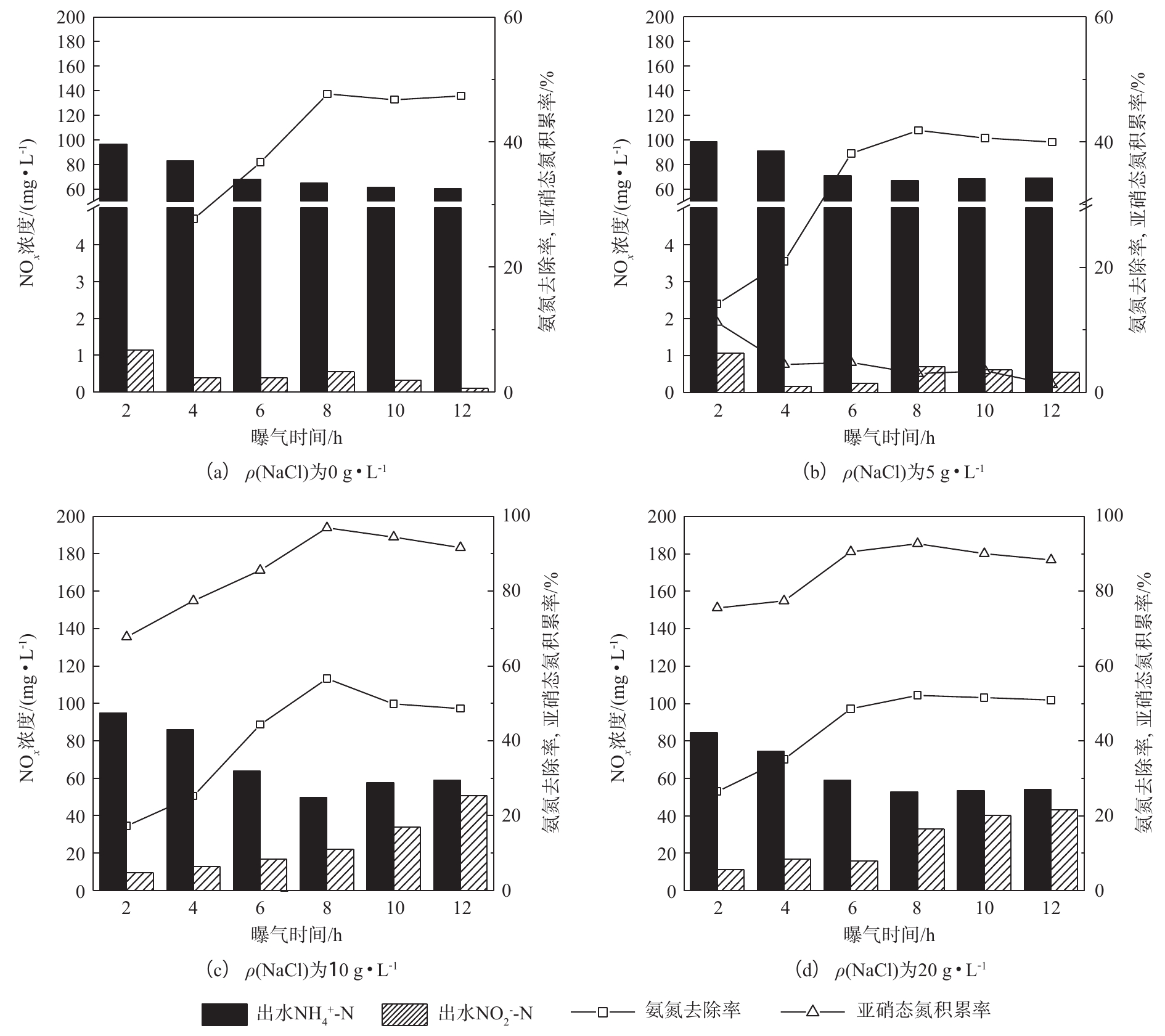
图 5 不同ρ(NaCl)下曝气时间对氨氮去除率和亚硝态氮积累率的影响
Figure 5. Effect of aeration time on the nitrogen removal rate and nitrite accumulation rate at different ρ(NaCl)
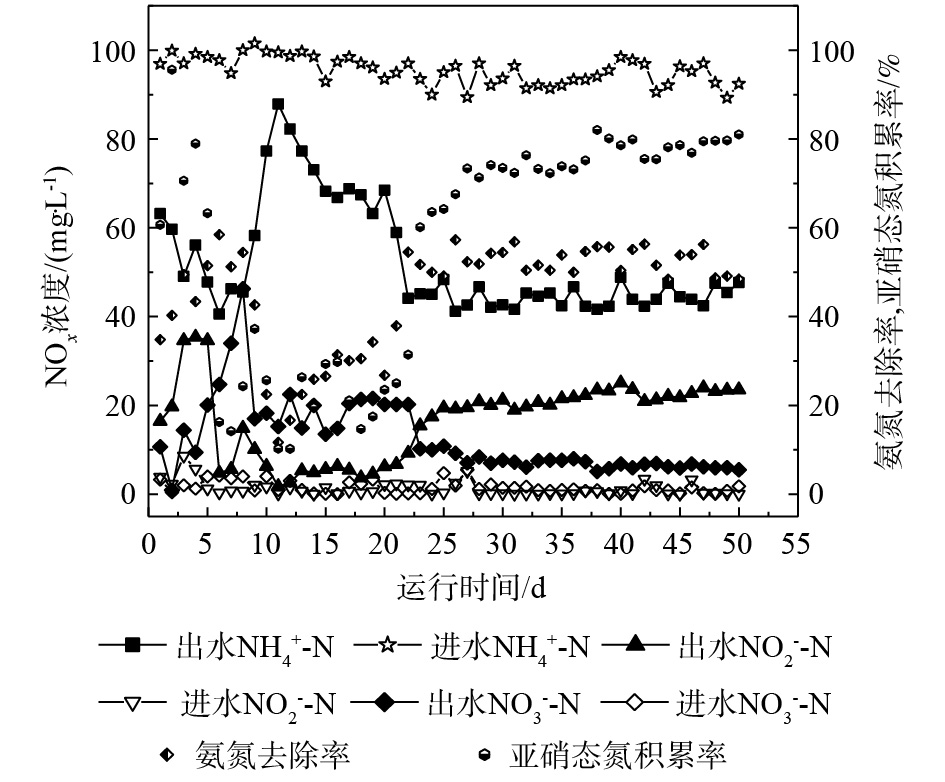
图 6 UASB反应器运行期间
NH+4 -N、NO−2 -N、NO−3 -N浓度变化Figure 6. Concentration change of
NH+4 -N,NO−2 -N,NO−3 -N in UASB reactor during operation表 1 基质浓度对短程硝化反应的影响
Table 1. Effect of substrate concentrationon partial nitrification
水样序号 NH+4 -N浓度 /(mmol·L−1)</td> <td class="table_top_border" align="center" valign="middle"><inline-formula>${\rm{NH}}_4^{+} $<alternatives><img class="graphic" src="201902020liuchengcheng_Z-20190624170635.jpg"><img class="graphic" src="201902020liuchengcheng_Z-20190624170635.png"></alternatives></inline-formula>-N去除/(kg·(kg·d)<sup>−1</sup>)</td> </tr> </thead>
<tbody> <tr> <td class="table_top_border2" align="center" valign="middle">1</td> <td class="table_top_border2" align="center" valign="middle">1</td> <td class="table_top_border2" align="center" valign="middle">0.613</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">2</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">2</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.254</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">3</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">5.5</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.527</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">4</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">7.14</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.592</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">5</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">10.07</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.675</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">6</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">20.04</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.758</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">7</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">40.71</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.731</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">8</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">50.42</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.714</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">9</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">59.64</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.709</td> </tr> <tr> <td class="table_bottom_border" align="center" valign="middle">10</td> <td class="table_bottom_border" align="center" valign="middle">82</td> <td class="table_bottom_border" align="center" valign="middle">1.672</td> </tr> </tbody>
</table></div></foreignObject></svg>"></inline-formula>-N浓度 /(mmol·L<sup>−1</sup>)</td> <td class="table_top_border" align="center" valign="middle"><inline-formula>${\rm{NH}}_4^{+} $<img class="inline-formula" src="data:image/svg+xml,<svg xmlns='http://www.w3.org/2000/svg' width='350' height='600'><foreignObject width='2000' height='100%'><div xmlns='http://www.w3.org/1999/xhtml' style='font-size:16px;'><table>
<thead> <tr> <td class="table_top_border" align="center" valign="middle">水样序号</td> <td class="table_top_border" align="center" valign="middle"><inline-formula>${\rm{NH}}_4^{+} $<alternatives><img class="graphic" src="201902020liuchengcheng_Z-20190624170626.jpg"><img class="graphic" src="201902020liuchengcheng_Z-20190624170626.png"></alternatives></inline-formula>-N浓度 /(mmol·L<sup>−1</sup>)</td> <td class="table_top_border" align="center" valign="middle"><inline-formula>${\rm{NH}}_4^{+} $<alternatives><img class="graphic" src="201902020liuchengcheng_Z-20190624170635.jpg"><img class="graphic" src="201902020liuchengcheng_Z-20190624170635.png"></alternatives></inline-formula>-N去除/(kg·(kg·d)<sup>−1</sup>)</td> </tr> </thead>
<tbody> <tr> <td class="table_top_border2" align="center" valign="middle">1</td> <td class="table_top_border2" align="center" valign="middle">1</td> <td class="table_top_border2" align="center" valign="middle">0.613</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">2</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">2</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.254</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">3</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">5.5</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.527</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">4</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">7.14</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.592</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">5</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">10.07</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.675</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">6</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">20.04</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.758</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">7</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">40.71</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.731</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">8</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">50.42</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.714</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">9</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">59.64</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.709</td> </tr> <tr> <td class="table_bottom_border" align="center" valign="middle">10</td> <td class="table_bottom_border" align="center" valign="middle">82</td> <td class="table_bottom_border" align="center" valign="middle">1.672</td> </tr> </tbody>
</table></div></foreignObject></svg>"></inline-formula>-N去除/(kg·(kg·d)<sup>−1</sup>)</td> </tr> </thead>
<tbody> <tr> <td class="table_top_border2" align="center" valign="middle">1</td> <td class="table_top_border2" align="center" valign="middle">1</td> <td class="table_top_border2" align="center" valign="middle">0.613</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">2</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">2</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.254</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">3</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">5.5</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.527</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">4</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">7.14</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.592</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">5</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">10.07</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.675</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">6</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">20.04</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.758</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">7</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">40.71</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.731</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">8</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">50.42</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.714</td> </tr> <tr> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">9</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">59.64</td> <td align="center" valign="middle" style="height:13pt;padding-top:1.5pt;">1.709</td> </tr> <tr> <td class="table_bottom_border" align="center" valign="middle">10</td> <td class="table_bottom_border" align="center" valign="middle">82</td> <td class="table_bottom_border" align="center" valign="middle">1.672</td> </tr> </tbody>
</table></div></foreignObject></svg>)
NH+4 -N去除/(kg·(kg·d)−1)1 1 0.613 2 2 1.254 3 5.5 1.527 4 7.14 1.592 5 10.07 1.675 6 20.04 1.758 7 40.71 1.731 8 50.42 1.714 9 59.64 1.709 10 82 1.672
下载: 导出CSV
-
[1] 李凌云, 彭永臻, 吴蕾, 等. 短程硝化颗粒污泥的培养与特性分析[J]. 土木建筑与环境工程, 2010, 32(3): 119-123. doi: 10.11835/j.issn.1674-4764.2010.03.021 [2] 赵梦月, 彭永臻, 王博, 等. SBR工艺实现长期稳定的部分短程硝化[J]. 化工学报, 2016, 67(6): 2525-2532. [3] ISANTA E, REINO C, CARRERA J, et al. Stable partial nitritation for low-strength wastewater at low temperature in an aerobic granular reactor[J]. Water Research, 2015, 80: 149-158. doi: 10.1016/j.watres.2015.04.028 [4] GE S J, WANG S Y, YANG X, et al. Detection of nitrifiers and evaluation of partial nitrification for wastewater treatment: A review[J]. Chemosphere, 2015, 140: 85-98. doi: 10.1016/j.chemosphere.2015.02.004 [5] 侯爱月, 李军, 卞伟, 等. 不同短程硝化系统中微生物群落结构的对比分析[J]. 中国环境科学, 2016, 36(2): 428-436. doi: 10.3969/j.issn.1000-6923.2016.02.019 [6] GARRIDO J M, VANBENTHUM W A, VANLOOSDRECHT M C, et al. Influence of dissolved oxygen concentration on nitrite accumulation in a biofilm airlift suspension reactor[J]. Biotechnology and Bioengineering, 1997, 53(2): 168-178. doi: 10.1002/(ISSN)1097-0290 [7] CIUDAD G, GONZALEZ R, BORNHARDT C, et al. Modes of operation and pH control as enhancement factors for partial nitrification with oxygen transport limitation[J]. Water Research, 2007, 41(20): 4621-4629. doi: 10.1016/j.watres.2007.06.036 [8] 张小玲, 彭党聪, 王志盈. 传统与短程反硝化的影响因素及特性研究[J]. 中国给水排水, 2002, 18(9): 1-3. doi: 10.3321/j.issn:1000-4602.2002.09.001 [9] GUO J H, PENG Y Z, HUANG H J, et al. Short- and long-term effects of temperature on partial nitrification in a sequencing batch reactor treating domestic wastewater[J]. Journal of Hazardous Materials, 2010, 179(1/2/3): 471-479. [10] 彭永臻, 杨岸明, 李凌云, 等. 短程硝化最优曝气时间控制与硝化种群调控[J]. 哈尔滨工业大学学报, 2013, 45(2): 101-105. doi: 10.11918/j.issn.0367-6234.2013.02.018 [11] 吴煌州, 邱凌峰, 陈益明, 等. 不同曝气量对短程硝化特性的影响[J]. 福州大学学报(自然科学版), 2014, 42(1): 161-166. doi: 10.7631/issn.1000-2243.2014.01.0161 [12] LI X, YUAN Y, YUAN Y, et al. Effects of salinity on the denitrification efficiency and community structure of a combined partial nitritation- anaerobic ammonium oxidation process[J]. Bioresource Technology, 2018, 249: 550-556. doi: 10.1016/j.biortech.2017.10.037 [13] 张晓玲. 盐度对SBR和SBBR工艺短程硝化反硝化的影响[D]. 青岛: 中国海洋大学, 2015. [14] HAMODA M F, AL-ATTAR M S. Effects of high sodium chloride concentrations on activated sludge treatment[J]. Water Science and Technology, 1995, 31(9): 61-72. doi: 10.2166/wst.1995.0345 [15] ASLAN S, SIMSEK E. Influence of salinity on partial nitrification in a submerged biofilter[J]. Bioresource Technology, 2012, 80: 24-29. [16] 徐洁, 熊小京, 郑天凌, 等. 序批式曝气生物滤池处理含海水污水中的硝化性能[J]. 环境工程学报, 2015, 9(3): 1062-1066. doi: 10.12030/j.cjee.20150312 [17] 陈光辉, 李军, 邓海亮, 等. 热冲击法实现包埋活性污泥稳定亚硝化[J]. 四川大学学报(工程科学版), 2015, 47(5): 196-202. [18] 陈光辉, 李军, 邓海亮, 等. 包埋菌启动厌氧氨氧化反应器及其动力学性能[J]. 化工学报, 2015, 66(4): 1459-1466. [19] CHEN Y, LIU Q G, ZHOU T, et al. Ethanol production by repeated batch and continuous fermentations by saccharomyces cerevisiae immobilized in a fibrous bed bioreactor[J]. Journal of Microbiology and Biotechnology, 2013, 23(4): 511-517. doi: 10.4014/jmb [20] DONG Y M, ZHANG Z J, JIN Y W, et al. Nitrification characteristics of nitrobacteria immobilized in waterborne polyurethane in wastewater of corn-based ethanol fuel production[J]. Journal of Environmental Sciences, 2012, 24(6): 999-1005. doi: 10.1016/S1001-0742(11)60893-0 [21] 李尧, 张振家, 方海军. 包埋固定化微生物工艺技术处理高氨氮化工废水[J]. 净水技术, 2012, 31(3): 32-35. doi: 10.3969/j.issn.1009-0177.2012.03.008 [22] STROUS M, GERVEN E V, ZHENG P, et al. Ammonium removal from concentrated waste streams with the anaerobic ammonium oxidation (anammox) process in different reactor configurations[J]. Water Research, 1997, 31(8): 1955-1962. doi: 10.1016/S0043-1354(97)00055-9 [23] 张倩, 王淑莹, 苗圆圆, 等. 间歇低氧曝气下CANON工艺处理生活污水的启动[J]. 化工学报, 2017, 68(1): 289-296. [24] 叶柳, 彭永臻, 崔有为, 等. 革新MUCT工艺处理含盐污水[J]. 北京工业大学学报, 2006, 32(12): 1087-1092. doi: 10.11936/bjutxb2006121087 [25] 王淑莹, 唐冰, 叶柳, 等. NaCl盐度对活性污泥系统脱氮性能的影响[J]. 北京工业大学学报, 2008, 34(6): 631-635. doi: 10.11936/bjutxb2008060631 [26] 叶柳. 含盐生活污水脱氮除磷的特性及优化控制[D]. 北京: 北京工业大学, 2010. [27] 雷中方. 高浓度钠盐对废水生物处理系统的失稳影响综述[J]. 工业水处理, 2000, 20(4): 6-9. doi: 10.3969/j.issn.1005-829X.2000.04.002 [28] CUI Y W, PENG Y Z, GAN X Q, et al. Achieving and maintaining biological nitrogen removal via nitrite under normal conditions[J]. Journal of Environmental Sciences, 2005, 17(5): 794-797. [29] SANCHEZ O, MARTI M C, ASPE E, et al. Nitrification rates in a saline medium at different dissolved oxygen concentrations[J]. Biotechnology Letters, 2001, 23(19): 1597-1602. doi: 10.1023/A:1011977629398 [30] 张昭, 李冬, 仲航, 等. 常温下低氨氮污水的短程硝化启动研究[J]. 中国给水排水, 2012, 28(9): 35-39. doi: 10.3969/j.issn.1000-4602.2012.09.009 [31] YE L, PENG C Y, TANG B, et al. Determination effect of influent salinity and inhibition time on partial nitrification in a sequencing batch reactor treating saline sewage[J]. Desalination, 2009, 246(1/2/3): 556-566. [32] 崔有为, 彭永臻, 李晶. 盐度抑制下的MUCT处理效能及其微生物种群变化[J]. 环境科学, 2009, 30(2): 488-492. doi: 10.3321/j.issn:0250-3301.2009.02.030 [33] 李芸, 熊向阳, 李军, 等. 膜生物反应器处理晚期垃圾渗滤液亚硝化性能及其抑制动力学分析[J]. 中国环境科学, 2016, 36(2): 419-427. doi: 10.3969/j.issn.1000-6923.2016.02.018 [34] 胡晓娜, 戴晓虎, 曹达文, 等. 限氧条件下亚硝化的稳定运行及动力学[J]. 化工进展, 2015, 34(12): 4198-4202. [35] 许晓毅, 尤晓露, 吕晨培, 等. 包埋固定化活性污泥脱氮特性与微生物群落分析[J]. 环境科学, 2017, 38(5): 2052-2058. [36] 唐崇俭, 熊蕾, 王云燕, 等. 高效厌氧氨氧化颗粒污泥的动力学特性[J]. 环境科学, 2013, 34(9): 3544-3551. 期刊类型引用(3)
1. 夏兴良,梁志超,陈书科,潘振. 短程硝化的影响因素及其强化方法的研究进展. 广东化工. 2024(09): 116-118 .  百度学术
百度学术
2. 张馨文,王荣震,冯成业,张文智,徐征和. 生活污水短程硝化脱氮工艺的快速启动及稳定性研究. 环境工程. 2022(10): 9-14 . 百度学术
3. 高慧娟,向斯,程凯. 适用于低氨污水的高效氨氧化菌的分离筛选及其氨氧化特性. 环境科学与技术. 2020(01): 150-157 . 百度学术
其他类型引用(6)
-

 点击查看大图
点击查看大图

计量
- 文章访问数: 4730
- HTML全文浏览数: 4730
- PDF下载数: 53
- 施引文献: 9
