


 DownLoad:
DownLoad:

 百度学术
百度学术


-
氨氮是藻类生长所需的营养物质之一,然而氨氮过量会导致藻类数量激增,破坏水体平衡,藻类生长也会造成水中溶解氧急剧下降,进而严重影响水生生物的生存。高浓度氨氮废水因排放量大、成分复杂、毒性强、对生态环境危害性大,而受到人们广泛关注,现在已有成熟的处理方法,但研究者往往易忽略低浓度氨氮废水的处理[1]。有研究[2]表明,低浓度氨氮对大多数鱼类依然具有毒性。因此,探究实用且有效的低浓度氨氮废水处理方法迫在眉睫。
常用去除氨氮的方法可分为生物法和物理化学法2大类[3]。生物法主要包括厌氧氨氧化[4]、硝化反硝化[5]和藻类培养[6]等。但生物法启动慢,且易受生存环境影响,如温度、pH和进水水质等均可能影响生物法的处理效率[7]。物理化学法种类较多,主要包括吹脱法[8]、化学沉淀法[9]、电渗析法[10]和离子交换法[11-12]等。吹脱法与化学沉淀法都比较适合高浓度氨氮去除,不适用于低浓度氨氮的处理[8]。电渗析法同样仅适合在高浓度条件下进行,且能量效率较低。而离子交换法适用于低浓度条件下氨氮的去除,且离子交换剂吸附饱和后可进行再生,其再生液可进行氨氮的回收,具有能耗低、占地小、操作简单等优点。
常用的离子交换剂有天然沸石、改性沸石、离子交换树脂等[13]。现有研究一般采用对氨氮具有高选择性的沸石为离子交换剂[14]。但沸石的吸附容量较离子交换树脂低,且再生过程会对其吸附性能造成损害[15-16]。刘宝敏等[17]研究了强酸型离子交换树脂对氨氮的吸附,吸附容量约可达13.30 mg·g−1,且再生后仍然具有良好的吸附性能。吴达兵[18]研究了自制离子交换树脂对氨氮的吸附,吸附容量约为30.34 mg·g−1。相较于离子交换树脂,粉末树脂常用于火电厂凝结水处理系统,技术比较成熟,其主要作用为过滤除铁及化学除盐[19]。粉末树脂具有离子交换作用,较小的粒径(100 μm)增加了其与污水接触面积,吸附容量更高,可减少再生次数[20]。然而目前极少有研究者利用粉末树脂去除生活污水中的氨氮,故研究Na型粉末树脂具有极其重要的意义。
本研究通过静态批式实验分析预处理、pH及粉末树脂投加量对粉末树脂回收低浓度氨氮的影响。在此基础上,进一步研究了水中钙离子和镁离子对离子交换过程的影响,并采用吸附等温线以及吸附动力学对数据进行了拟合。在静态实验的基础上,进一步开展动态实验,以探究运行方式对Na型粉末树脂吸附低浓度氨氮可行性的影响。本研究进一步验证了粉末树脂用于低浓度氨氮废水回收的可能性。
全文HTML
-
实验材料:H型粉末树脂、Na型粉末树脂、强酸型离子交换树脂。
实验试剂:氯化铵、氯化钙、氯化钾、氯化镁与去离子水配制溶液作为模拟废水,所用试剂均为分析纯;膜浓缩实际废水。
实验仪器:恒温振荡摇床、分析天平、紫外-可见光分光光度计和pH计。
-
1) 粉末树脂的预处理。将H型粉末树脂用饱和NaCl溶液冲洗至中性,再用去离子水清洗,去除粉末树脂中的NaCl。将清洗好的粉末树脂(Na型粉末树脂)置于60 ℃的烘箱中烘干,备用。
2) 静态吸附。将树脂投加到100 mL模拟氨氮废水(20、100、300 mg·L−1)溶液中,置于恒温振荡摇床中,反应温度稳定为25 ℃,150 r·min−1转速下振荡,分别在5、10、30、60 min进行取样,考察预处理、pH、粉末树脂投加量、共存阳离子、再生等因素对吸附效果的影响,其控制变量见表1。
3) 吸附等温线。将100 mL不同浓度(20、100、300 mg·L−1)的氨氮溶液置于150 mL锥形瓶中,分别加入0.1、0.2、0.4、0.6 g Na型粉末树脂,保持反应温度为25 ℃,150 r·min−1下振荡1 h,取上清液测定氨氮浓度。
4) 粉末树脂与离子交换树脂动态实验对比分析。粉末树脂与离子交换树脂密度不同,离子交换柱(高25 cm,内径2.6 cm)体积为133 mL,故取填充度为50%(70 mL)和相同质量(20 g)的树脂分别进行实验。废水自交换柱下端流入,上端流出,设定流速为6.7 mL·min−1,利用实际膜浓缩出水进行实验。每1 h取1次出水,绘制吸附曲线。
-
在本研究中,氨氮去除率、吸附速率、吸附容量及电子摩尔数依次按式(1)、式(2)、式(3)和式(4)计算。
式中:R为氨氮去除率;C0为离子初始质量浓度,mg·L−1;Ce为吸附平衡时溶液中离子质量浓度,mg·L−1;Ct为t时刻溶液中离子质量浓度,mg·L−1;m为树脂投加量,g;t为反应时间,min;M为阳离子摩尔质量,g·mol−1;n为离子带电子数。
1.1. 实验材料、试剂与仪器
1.2. 实验方法
1.3. 计算公式
-
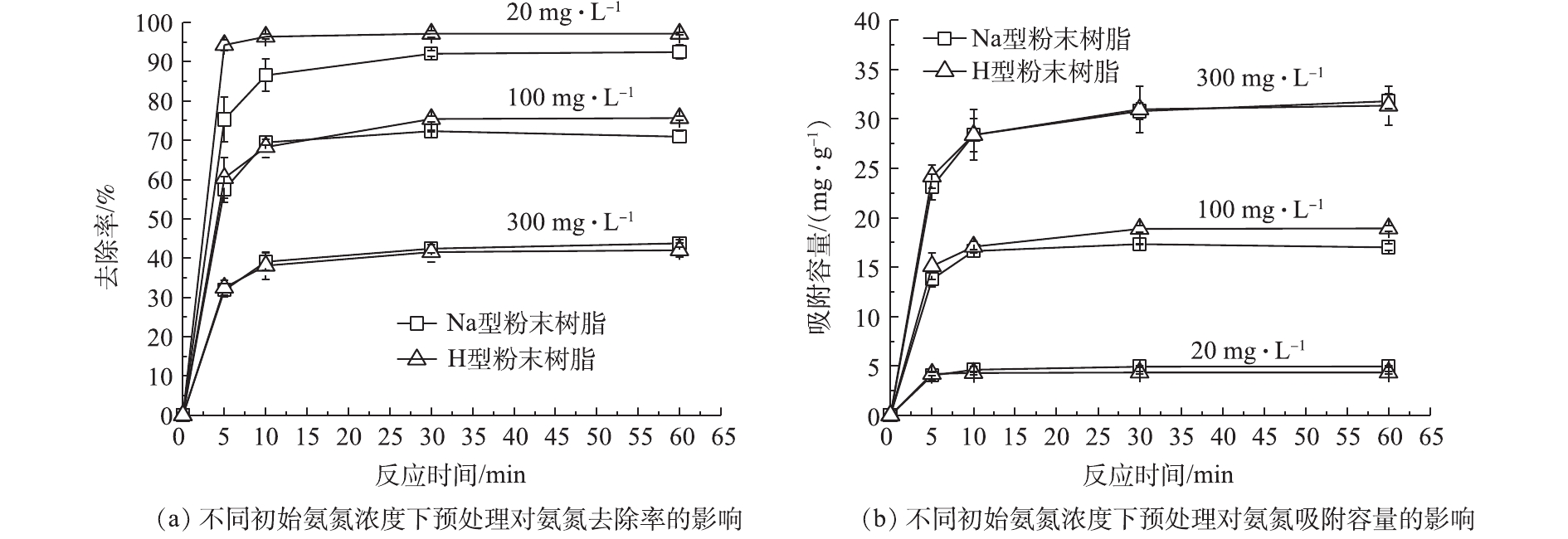
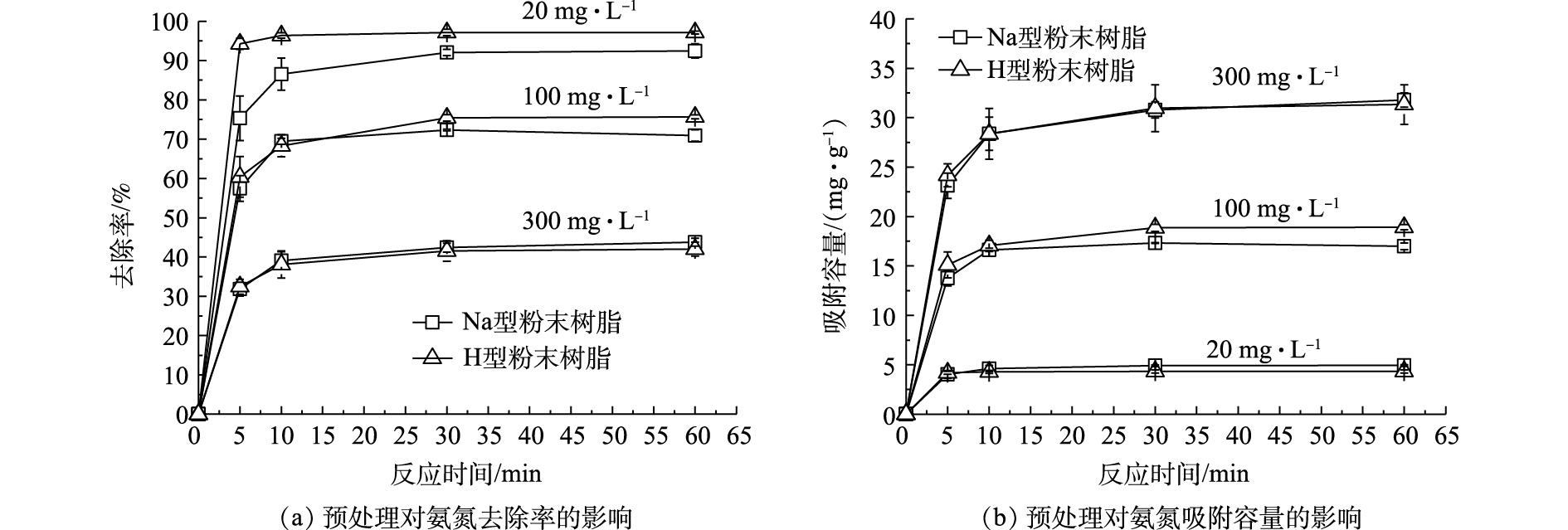
在不同氨氮浓度(20、100、300 mg·L−1)条件下,预处理对粉末树脂的氨氮去除率及吸附容量影响结果如图1所示。可以看出,进行预处理对粉末树脂的氨氮去除率和吸附容量均造成了一定程度的下降。这可能是由于在离子交换过程中,H型粉末树脂中H+与
NH+4 交换后,pH降低使更多的氨氮以NH+4 形式存在,更有利于离子交换,如式(5)所示。在预处理后,粉末树脂对氨氮的去除率和吸附容量仅降低了约5%。由此表明,预处理对粉末树脂吸附氨氮的能力影响较小。预处理后的Na型粉末树脂不仅具有较高的吸附氨氮能力,而且交换后的出水呈中性,故不影响后续处理。
-
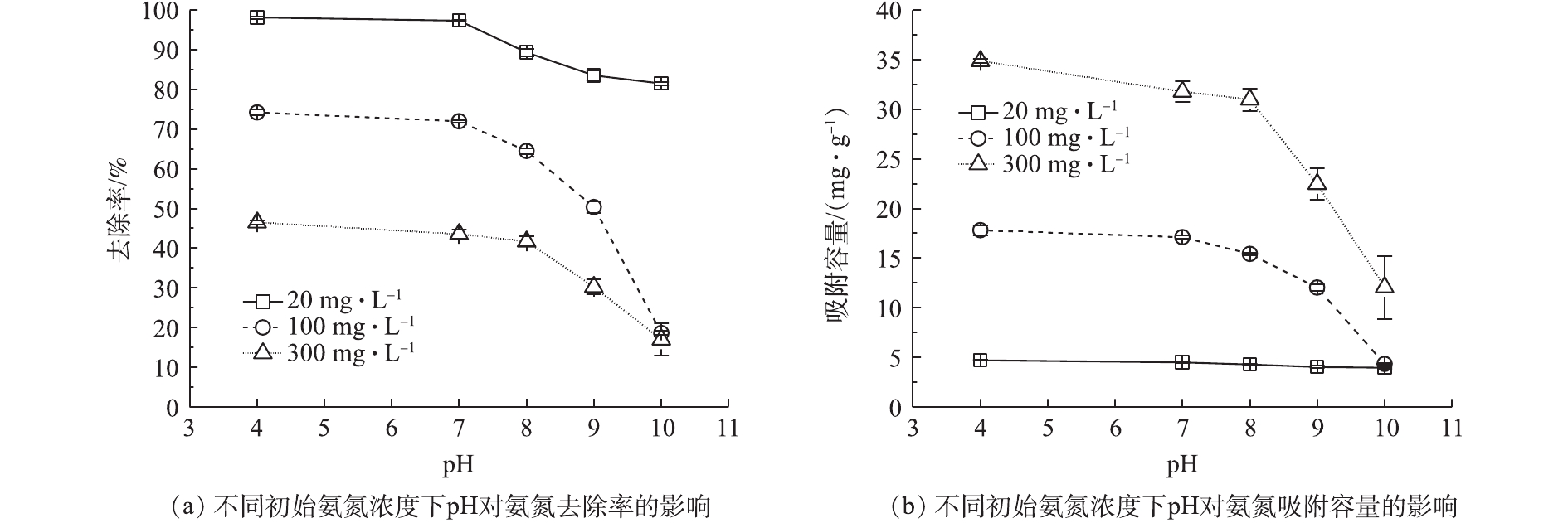
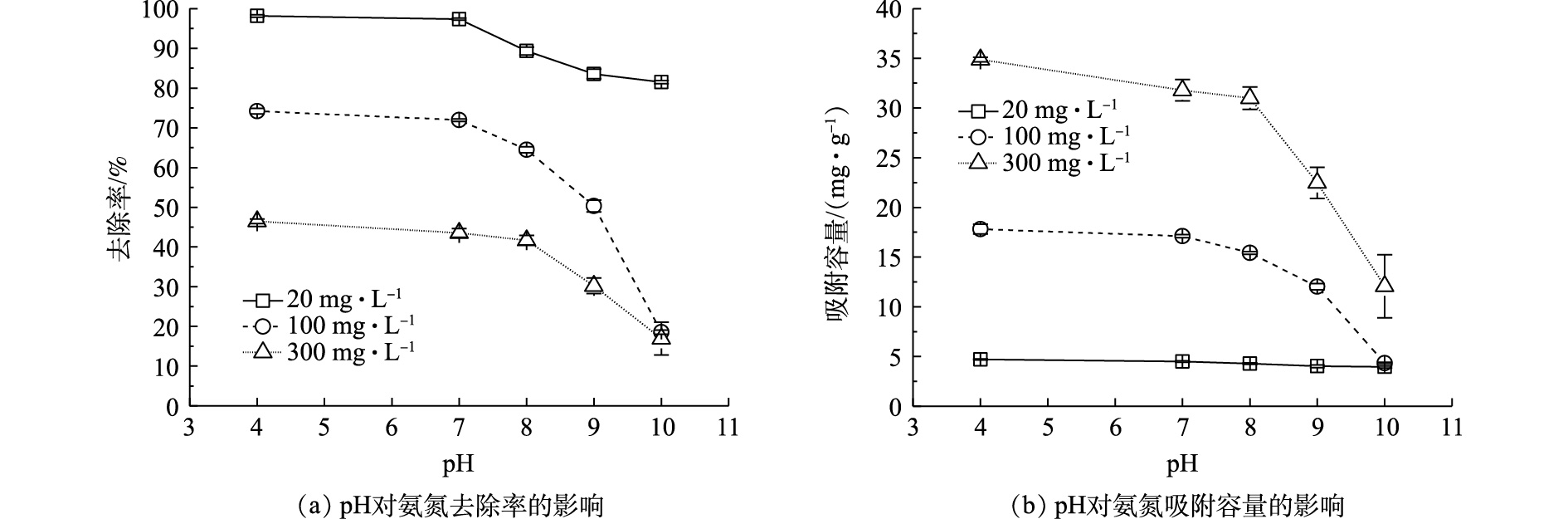
在不同氨氮浓度(20、100、300 mg·L−1)条件下,pH对Na型粉末树脂去除氨氮的去除率及吸附容量的影响如图2所示。由图2(a)可知,pH从4逐渐升高到10时,在不同氨氮浓度下,氨氮去除率与吸附容量均呈现不同程度的下降,与pH=4时相比,在pH=10时,氨氮去除率下降了17%~56%,吸附容量下降了0.8~22 mg·g−1。其原因可能为:pH≤8时,溶液中氨氮主要以离子态(
NH+4 )形式存在;pH>8时,平衡移向非离子态(NH3)形式,如式(5)所示。由于pH=7时的氨氮的去除率及吸附容量与pH=4时相近,且pH=7更接近于原废水pH,故选用pH=7进行后续的实验。 -
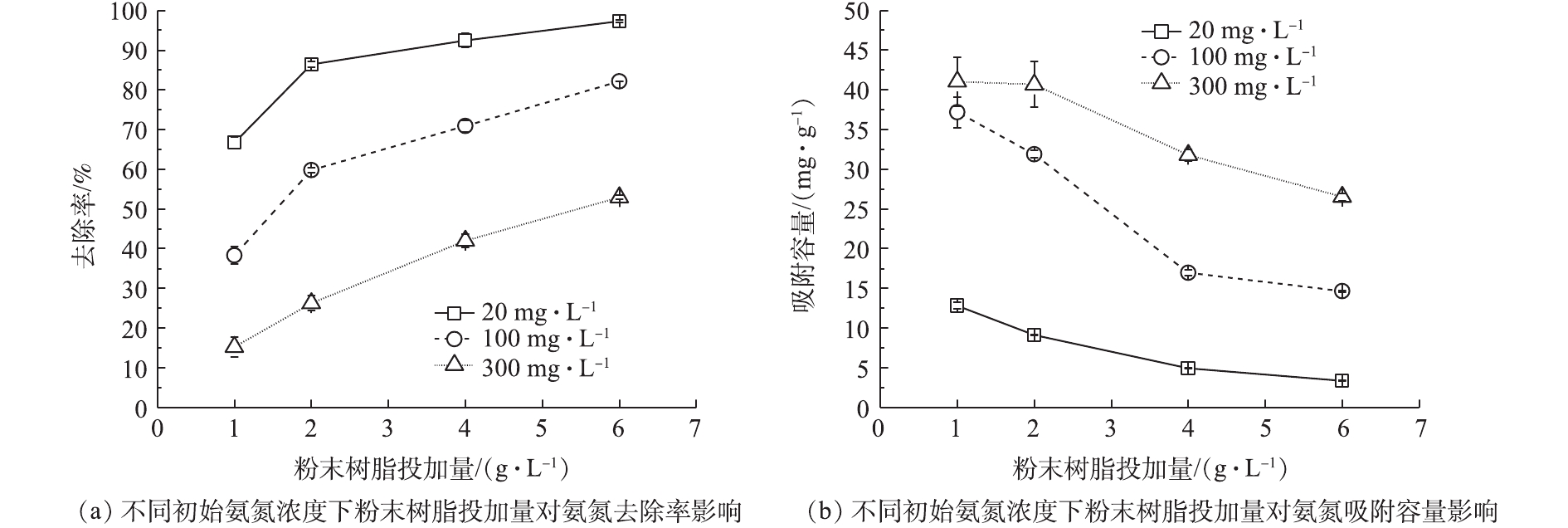
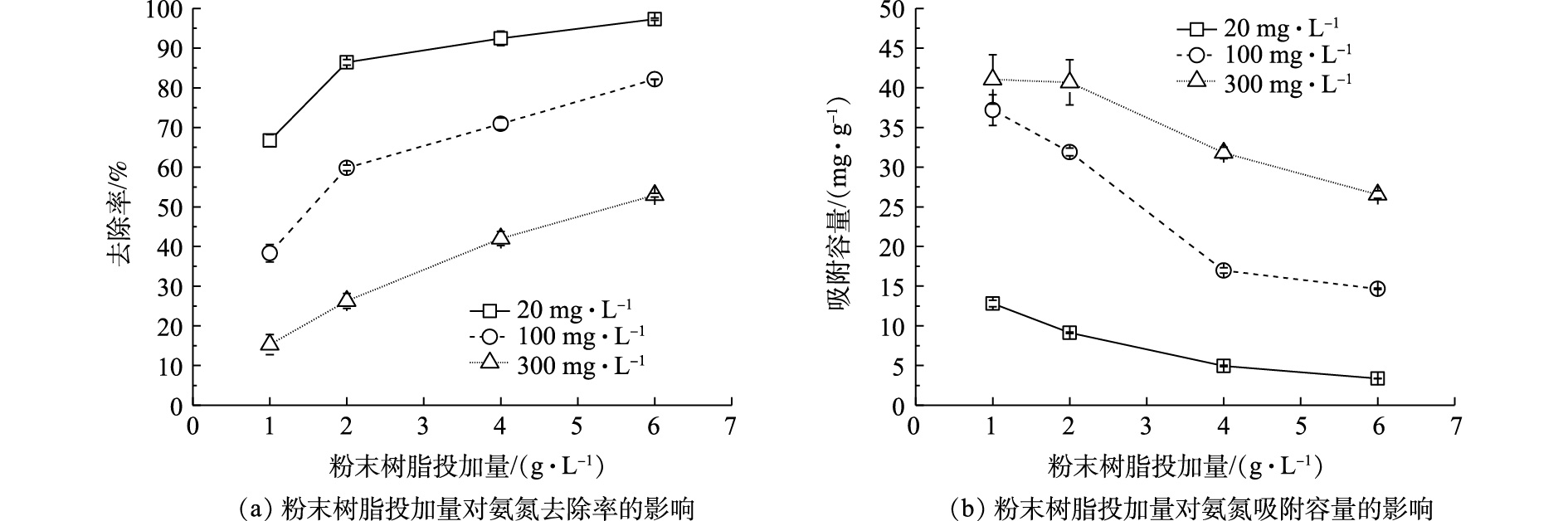
不同氨氮浓度(20、100、300 mg·L−1)条件下,Na型粉末树脂投加量对氨氮的去除率及吸附容量的影响如图3所示。由图3可知,增加Na型粉末树脂投加量,会提升氨氮的去除率,但会降低其吸附容量。这可能是因为树脂投加量越大,树脂表面积就越大,树脂吸收位点增加,相应的氨氮的去除率越高;Na型粉末树脂与氨氮浓度差越小,相应的吸附容量越低。Na型粉末树脂投加量为4 g·L−1时,不同初始氨氮浓度下氨氮吸附容量(4.85、16.93、31.33 mg·g−1)与粉末树脂投加量为6 g·L−1时氨氮吸附容量(3.36、14.65、26.55 mg·g−1)相比,吸附容量减少,但氨氮去除率并未有显著上升,因此,从经济成本考虑,选用粉末树脂投加量为4 g·L−1进行阳离子共存影响实验。
表2列出了不同初始氨氮浓度下不同粉末树脂投加量对应吸附速率随反应时间的变化情况。随反应时间的不断推进,吸附速率逐渐下降,在约30 min时,反应达到平衡。这是由于最初Na型粉末树脂与模拟废水中氨氮浓度相差大,反应驱动力大,但随着反应时间的增加,氨氮浓度下降,反应驱动力逐渐减小。
初始氨氮浓度为300 mg·L−1,粉末树脂投加量为2 g·L−1,反应为5 min时,得到最大吸附速率6.73 mg·(g·min)−1。这是由于粉末树脂投加量越低,与溶液中氨氮浓度差越大,反应驱动力越大,吸附速率也越高。
-
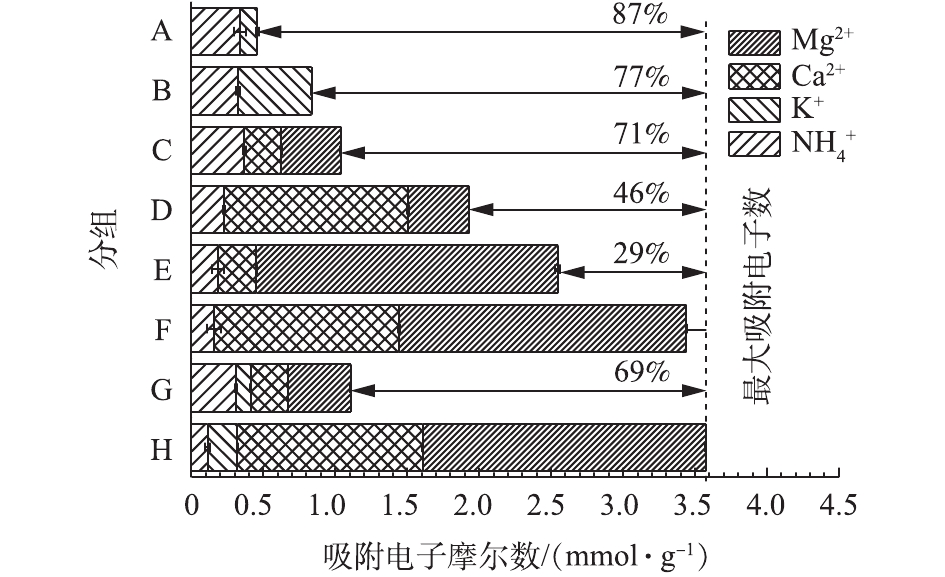
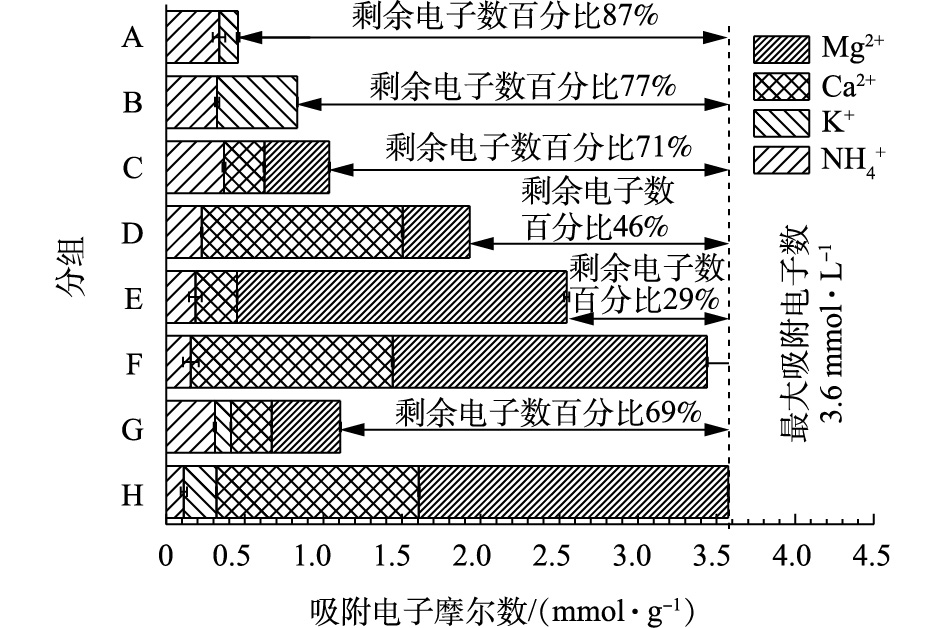
单位质量的Na型粉末树脂吸附各阳离子的电子分布情况如图4所示。表3为不同分组中各阳离子初始浓度。比较A组、B组、C组可知,K+对
NH+4 的吸附产生的抑制作用较小,这可能是由于K+与NH+4 价位相同,且水合半径相近,所以抑制作用较小;当二价阳离子与氨氮浓度均为20 mg·L−1时,共存阳离子对NH+4 的吸附无抑制作用,这可能是由于Na型粉末树脂中有足够的吸收位点,可以完全吸附低浓度(20 mg·L−1)的共存阳离子。比较D组、E组、F组可知,当二价阳离子浓度上升时,二价阳离子对
NH+4 的吸附具有抑制作用,这可能是由于Na型粉末树脂对离子的去除主要原理为离子交换,故离子价态越高,越容易被交换,从而会相应减少树脂上的吸收位点。由图4可知,Na型粉末树脂最大吸附电子数为3.6 mmol·L−1左右,且对NH+4 不具有选择性。比较G组、H组可知,当一价和二价阳离子共存时,在初始阳离子浓度均为低浓度条件下,Na型粉末树脂将阳离子全部吸附还剩余69%的吸附电子容量;将K+、Mg2+、Ca2+浓度提高到100 mg·L−1时,Na型粉末树脂的吸附达到饱和,最大吸附电子数约为3.6 mmol·g−1。二价阳离子对一价离子的吸附产生抑制作用。
-
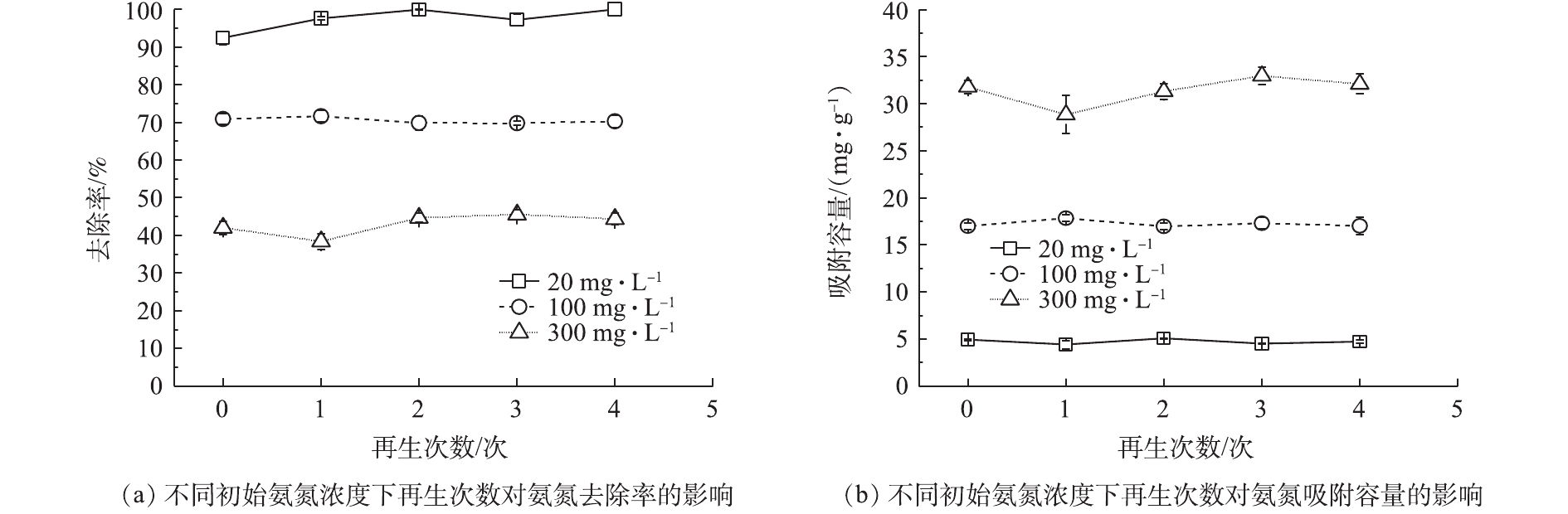
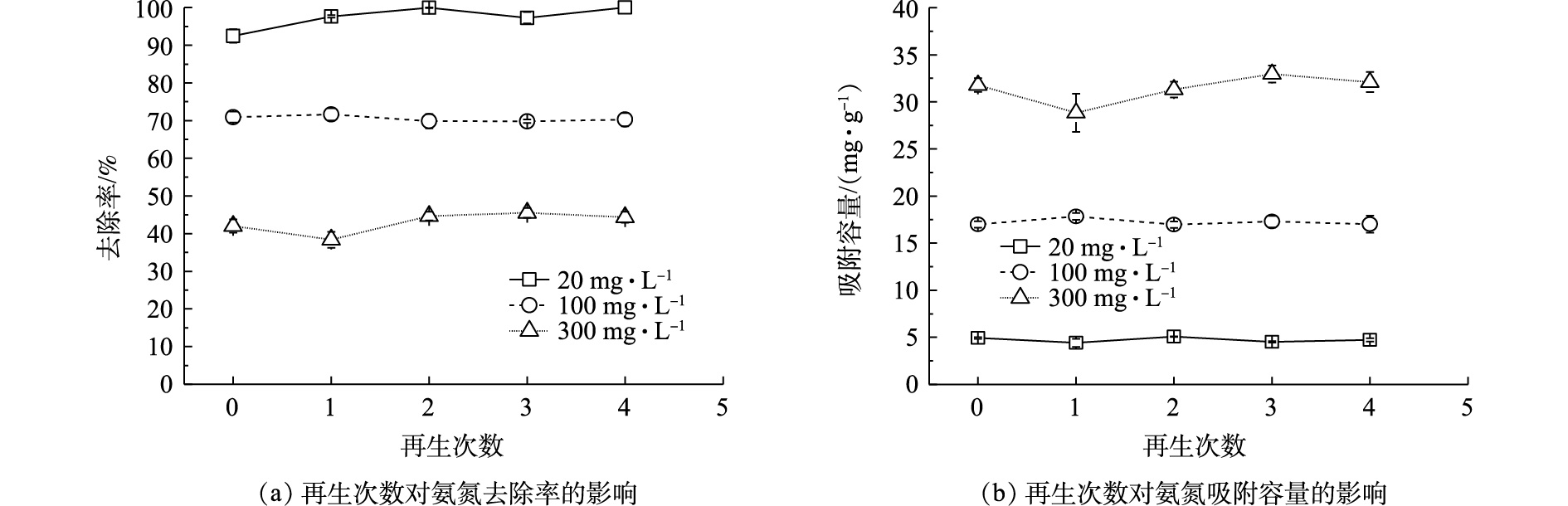
图5为Na型粉末树脂多次再生后对氨氮的吸附容量和去除率的影响。由图5可知,在不同初始氨氮浓度下,与新鲜树脂相比,再生4次后的粉末树脂对氨氮吸附容量下降了0.1~0.3 mg·g−1,其对氨氮的吸附性能未发生显著变化。这表明在粉末树脂失效后,经再生后可以反复使用,且再生后的再生液可以进行氨氮富集利用。因此,粉末树脂用在市政污水去除氨氮是可行的。
-
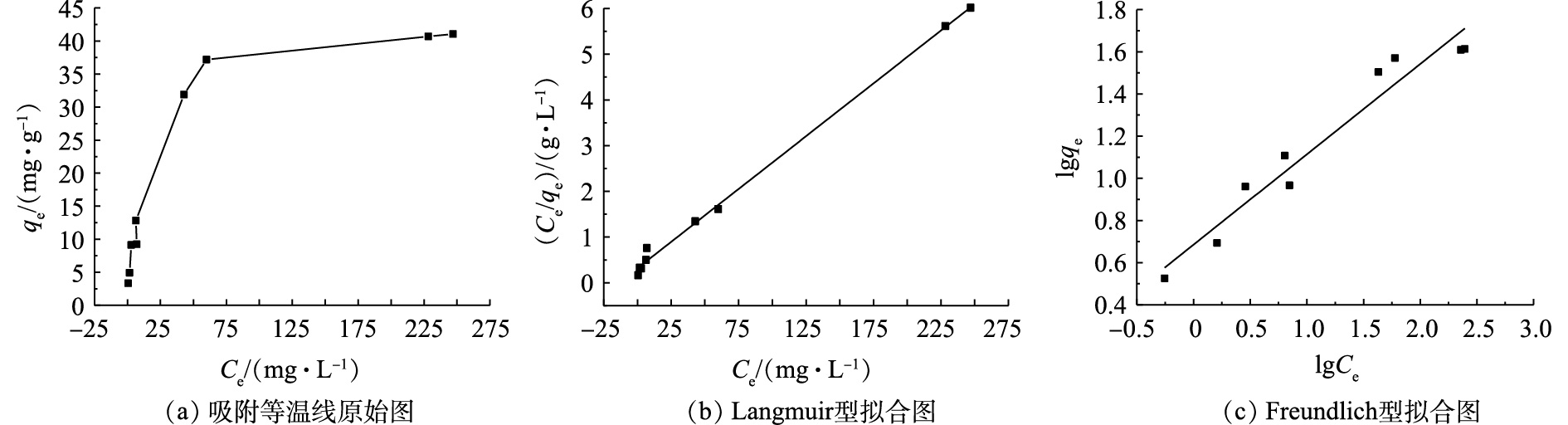
在25 ℃下,研究Na型粉末树脂吸附模拟废水中
NH4+ 的过程。通过Langmuir与Freundlich模型拟合吸附等温线,得到吸附材料的最大吸附容量、吸附相关系数等,拟合结果如图6所示。Langmuir和Freundlich吸附等温线方程如式(6)和式(7)所示。式中:qe为吸附平衡时的吸附容量,mg·g−1;qmax为理论饱和吸附容量,mg·g−1;KL为Langmuir吸附常数;n为经验常数;KF为Freundlich吸附常数。
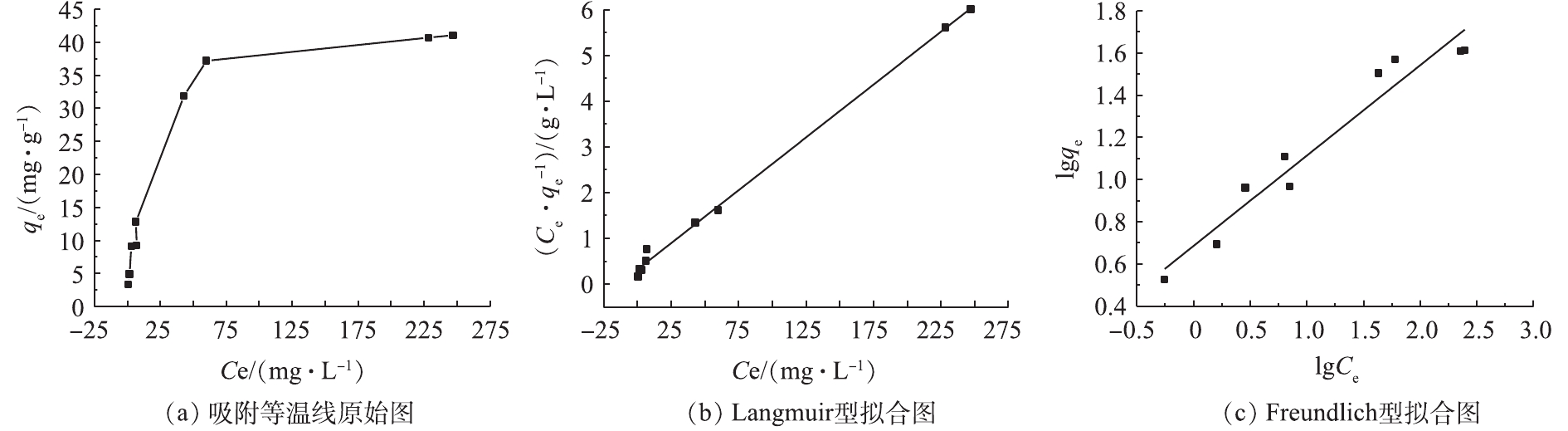
实验结果表明,Langmuir吸附等温线拟合系数R2=0.981 4,而Freundlich吸附等温线拟合系数R2=0.894 0,故更符合Langmuir吸附等温线拟合系数。这表明Na型粉末树脂对
NH4+ 离子的吸附以单分子层吸附为主。通过理论计算,得出最大吸附容量为44.39 mg·g−1,符合实验结果。其他拟合系数见表4。表5列出了4 g·L−1 Na型粉末树脂对20 mg·L−1氨氮的极限吸附容量。由表5可知,吸附氨氮21次,总的吸附容量达42.2 mg·g−1,与上述实验的最大吸附容量相符。这表明,Na型粉末树脂对低浓度氨氮的最大吸附容量不受氨氮浓度的限制,因此,可用于回收低浓度氨氮废水中的氨氮。
-
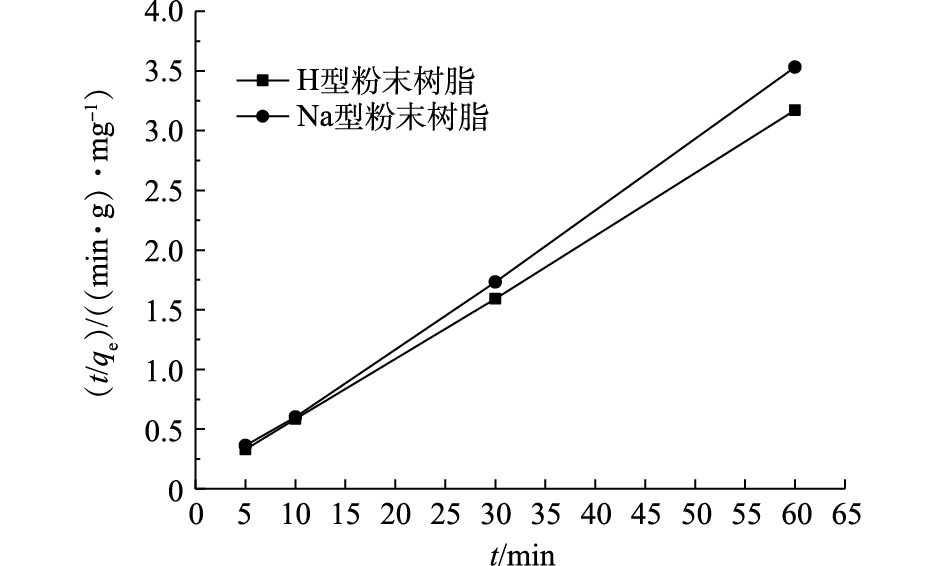
在Na型粉末树脂吸附氨氮时,粉末树脂内外的传质过程均符合线性推动力模型,故孔表面的反应步骤起决定作用。拟合采用的准二级反应动力学方程如式(8)所示。
式中:qt为t时刻Na型粉末树脂对氨氮的吸附容量,mg·g−1;k2为准二级反应动力学常数。
温度为25 ℃时,粉末树脂吸附氨氮拟合曲线如图7所示。由图7可知,H型和Na型粉末树脂对氨氮的吸附均符合准二级动力学模型。经过计算,H型粉末树脂k2为0.000 173,Na型粉末树脂k2为0.000 124。H型粉末树脂反应速率常数略大于Na型粉末树脂,说明H型粉末树脂能更快达到平衡。这是由于H+的水合半径大于Na+水合半径,当离子所带电荷量相同时,H+更易与
NH4+ 进行离子交换,其反应速率与Na+反应速率相比较快,这与上述实验结果相符。 -
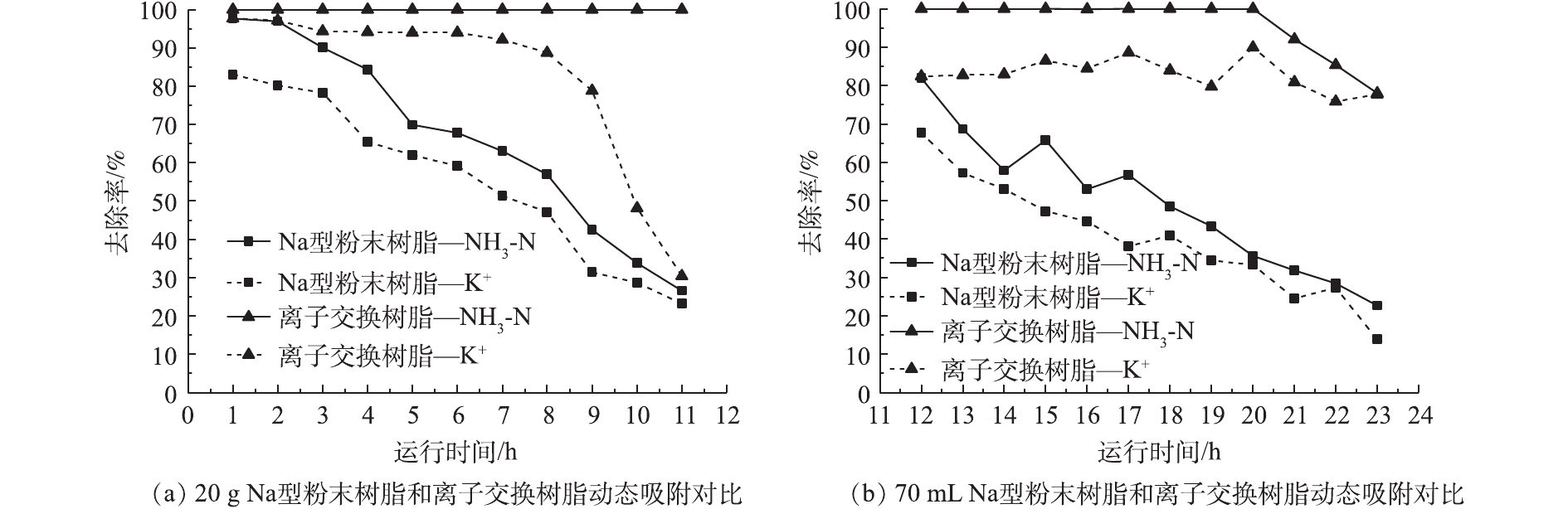
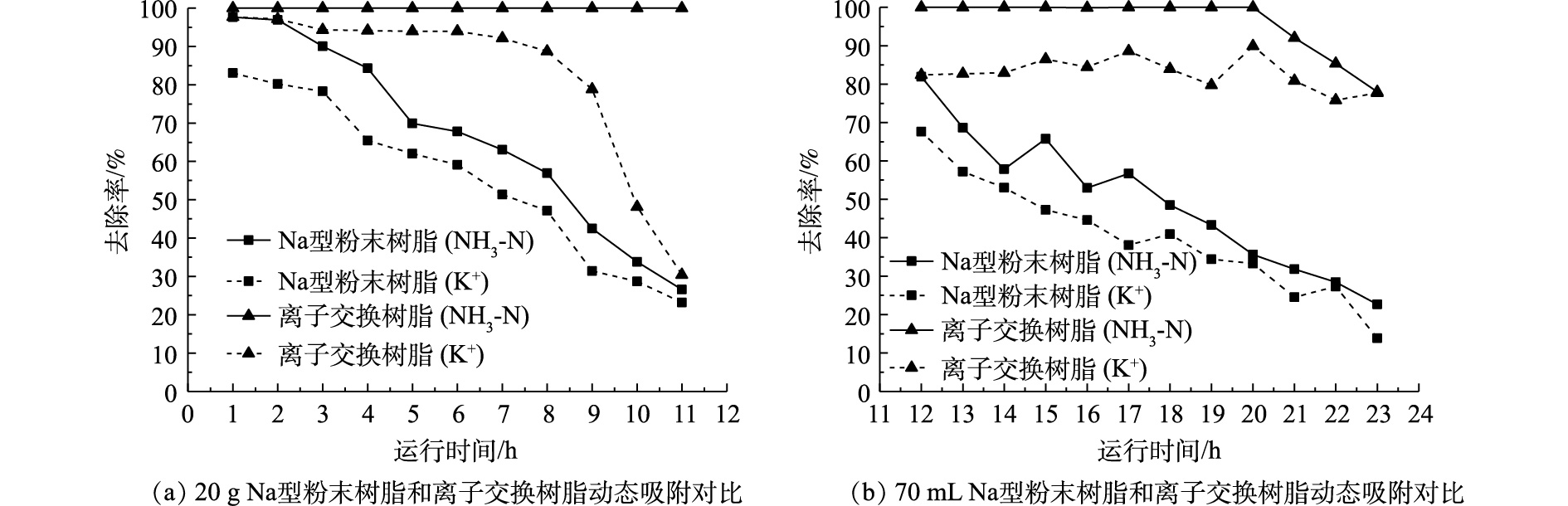
本研究选用膜浓缩后的出水作为实际污水进行离子交换树脂和Na型粉末树脂的对比分析,其水质参数如下:COD 15~25 mg·L−1,TN 50 mg·L−1,NH3-N 30 mg·L−1,TP 0.2~0.4 mg·L−1,Ca2+ 102 mg·L−1,Mg2+ 34 mg·L−1,K+ 11 mg·L−1,Na+ 72 mg·L−1,pH=7.80。Na型粉末树脂和离子交换树脂的动态吸附结果如图8所示。2种树脂对于Ca2+、Mg2+的去除率始终为100%,故未放在本研究中进行对比。为考察粉末树脂工程化运行的可行性,采用目前常用的离子交换柱进行氨氮实验。但由于粉末树脂粒径过小,粉末树脂积压在底部,导致其有效接触面积减少。进水采用升流式并未减轻粉末树脂沉积产生的影响。
利用实际污水对离子交换树脂和Na型粉末树脂进行静态实验,实验结果如表6所示。与离子交换树脂相比,Na型粉末树脂对二价阳离子具有更高的去除率(Mg2+为71.11%、Ca2+为87.03%)。升流式离子交换柱中Na型粉末树脂并未表现出对氨氮的高吸附效率,因此,该运行方式并不适合Na型粉末树脂,故需要进一步研究适用于Na型粉末树脂的其他运行方式。
2.1. 预处理前后对比
2.2. pH的影响
2.3. Na型粉末树脂投加量的影响
2.4. 不同阳离子的影响
2.5. 再生实验
2.6. 吸附等温线
2.7. 吸附动力学
2.8. 粉末树脂与离子交换树脂对比分析
-
1)预处理使得粉末树脂吸附氨氮的能力降低了5%;在中性和酸性条件下,Na型粉末树脂对低浓度氨氮的去除率均可达到99%;每增加2 g·L−1树脂投加量,会提高氨氮去除率20%,但吸附容量下降了2.85 mg·g−1;二价离子对单价离子吸附具有抑制作用,但同价态的离子吸附相互抑制作用较小。Na型粉末树脂再生后依然具有良好的吸附性能。
2) Na型粉末树脂对氨氮的吸附符合Langmuir吸附等温线,理论最大吸附容量为44 mg·g−1;Na型粉末树脂对氨氮的吸附符合准二级动力学方程,H型粉末树脂更快达到吸附平衡状态。
3) Na型粉末树脂虽对氨氮不具有选择性,但其具有较大的吸附容量,且再生几次后依然具有良好的吸附能力。静态实验结果表明,Na型粉末树脂适用于对低浓度氨氮的回收,但动态运行方式下效果并不显著,故需要进一步研究适用于Na型粉末树脂的其他运行方式。
