






























-
硒是人体必不可缺的微量元素之一,具有抗氧化、抗肿瘤、增强免疫力等作用[1 − 4]. 目前,硒元素的补充途径主要包括天然膳食,营养添加剂等,而富硒类植物作为天然膳食最直接便捷的主要来源,研究其中的硒形态及其抗氧化损伤功能对评估营养学价值,保护人体健康,探究科学补硒有着重大意义. 硒存在的形态有很多种,比较常见的有硒酸根(Se(Ⅵ))、亚硒酸根(Se(Ⅳ))、硒代胱氨酸(SeCys2)、甲基-硒代半胱氨酸(MeSeCys)、硒代蛋氨酸(SeMet)等. 葛粉具有降血脂、抗癌、消癌肿、抗氧化等特殊功效,而硒作为人体必需微量元素,可以清除自由基而表现出抗氧化、抗癌、抗炎症[5 − 6]的能力. 因此研究葛粉中硒形态及其抗氧化损伤功能研究具有重要意义.
本工作以葛粉为研究对象,建立高效液相色谱-电感耦合等离子体质谱联用技术测定葛粉中Se(Ⅵ)、Se(Ⅳ)、SeMet、SeCys2、SeEt、MeSeCys共6种硒形态的方法,并对葛粉中主要硒形态进行脂多糖诱导的AML12细胞氧化损伤的影响研究,初步探索了3种硒形态的抗氧化活性,以期为进一步研究富硒产品的功能和硒的生物有效性奠定基础,为科学补硒及评价富硒产品营养价值提供理论依据.
-
Agilent 7900型电感耦合等离子体质谱仪、Agilent 1260型高效液相色谱仪、LabTech EH20A plus 微控数显电热板、MileStone ETHOS A微波消解系统、THERMO GENPURE纯水机、日本Nikon TS100倒置显微镜、德国Innova CO-170 CO2培养箱、高速离心机、水浴温控摇床、漩涡振荡仪等.
Se(Ⅵ)[GBW 10033, (41.5±1.3) μg∙g−1]、Se(Ⅳ)[GBW 10032, (42.9±0.9) μg∙g−1]、SeCys2[GBW 10087, (44.2±1.0) μg∙g−1]、MeSeCys[GBW 10088, (34.8±1.0) μg∙g−1]、 SeMet[GBW 10034, (39.4±1.0) μg∙g−1]、Se[GBW(E)080215, 100 μg∙mL−1]标准溶液(均以Se计),均购自中国计量科学研究院;SeEt标准品的纯度为98%,购自加拿大TRC公司;蛋白酶XIV(P5147-1G)、脂肪酶(L3126-25G)、蛋白酶K(P6556-100MG)购自日本Sigma公司; 木瓜蛋白酶(P164463)、胃蛋白酶(P128678)购自北京百灵威科技有限公司; 柠檬酸、己烷磺酸钠、氨水、乙酸、氢氧化钠均为优级级;实验用水由THERMO GENPURE纯水机制得(>18.2 MΩ·cm). 标准储备液及标准品置于4℃冷藏,混合标准溶液系列均由标准储备液及标准品以超纯水逐级稀释配制,所有工作溶液均当天配制. 色谱进样前所有样品经0.22 μm滤膜过滤.
丙二醛(MDA)试剂盒(A003-1)、超氧化物歧化酶(SOD)试剂盒(A001-3)、谷胱甘肽(GSHx)试剂盒(A005-1)均购自南京建成科技有限公司; MTT细胞增殖及细胞毒性检测试剂盒(C0009s)购自南通碧云天生物技术有限公司; 小鼠肝实质细胞AML12购自中科院上海细胞库(SCSP-550); 脂多糖(L6143-1MG)购自Sigma公司.
-
微波消解条件 消解功率1500 W,升温程序:由室温升至120 ℃,升温时间3 min,恒温3 min;120 ℃升至190 ℃,升温时间5 min,恒温40 min.
色谱条件 色谱柱:Agilent ZORBAX SB-Aq(4.6 mm×250 mm,5 µm),流动相:20 mmol∙L−1柠檬酸,5.0 mmol∙L−1己烷磺酸钠(pH=4.4,甲醇加入量1 %),等度洗脱,流速:1.0 mL∙ min−1,进样量:20 μL.
ICP-MS条件 射频功率1550 W,蠕动泵速率0.4 r∙min−1,雾化室温度2.0 ℃,载气流量1.09 L∙min−1,等离子体气流量15 L∙min−1,采样深度:8.0 mm,积分时间0.5 s,高氦碰撞反应池模式,流量6.0 mL∙ min−1.
-
称取0.5000 g样品于消解罐内,加入6 mL 硝酸和2 mL过氧化氢溶液进行微波消解. 按照1.2条件微波消解. 消解结束后,冷却,打开消解罐,电热板上赶酸至近干,用超纯水洗涤消解罐 3 次,合并洗涤液,再用超纯水定容至25 mL,摇匀. 直接用 ICP-MS 测定硒总量. 同样方法做试剂空白试验.
-
(1)碱提取 称取0.1000 g样品于15 mL离心管内,加入10 mL 0.1 moL∙L−1 氢氧化钠溶液,放入超声波清洗器中90 ℃恒温超声30 min,冷却,在15000 r ∙ min−1的条件下离心15 min后,调至pH 6.5,取两份3 mL样品溶液转移至A、B离心管. A管经15000 r ∙ min−1离心15 min后,上清液经0.22 µm滤膜过滤,测定Se(Ⅵ)、Se(Ⅳ)、SeCys2.
(2)酶提取 将碱提取后的B离心管加入10 mL水、10 mg 蛋白酶XIV、10 mg脂肪酶、10 mg淀粉酶,涡旋3 min后置于37 ℃水浴温控摇床中,在150 r∙min−1条件下振摇18 h,15000 r ∙ min−1离心15 min后,上清液经0.22 µm滤膜过滤,得到待测液B,测定MeSeCys、SeMet、SeEt.
-
(1)分组与处置:以(0(正常组)、0.01、0.1、1、10 μmol∙L−1)的硒代蛋氨酸、硒酸根、亚硒酸根(均以Se计)处理脂多糖诱导的AML12细胞48 h,MTT实验测定细胞活力. 将AML12细胞随机分为5组:正常组(不加任何药物刺激)、模型组(加10 μg∙ml−1 LPS刺激),硒代蛋氨酸低浓度组、硒代蛋氨酸中浓度组、硒代蛋氨酸高浓度组,每组细胞培养48 h. 实验重复3次.
(2)细胞活力测定:按照上述分组与处置将细胞培养48 h后,加入200 μL二甲基亚砜孵育4 h,后加入20 μL的MTT溶液,在酶标仪620 nm处测定细胞吸光值(DO值), DO值即为细胞活力.
(3)细胞上清液中SOD、GSHx活性及MDA含量:按照上述分组与处置将细胞培养48 h后,收集上清液,并分别严格按照试剂盒说明书检测SOD、GSHx活性及MDA含量.
-
结果采用SPSS 20软件进行统计分析,计量资料结果以均数±标准差(x±s)表示,多组间比较采用单因素方差分析、组间两两比较采用t检验,P<0.05说明差异具有统计学意义.
-
以1 µg∙L−1的硒标准溶液进行碰撞池气体流量优化,随着He流量增加,78Se的灵敏度出现先升高后下降的趋势,在氦气流量达到6.0 mL∙ min−1时78Se灵敏度最高,最终确定碰撞池气体氦气的最佳流量为6.0 mL∙ min−1.
-
本实验室前期研究已确定硒形态分离的相关色谱参数,具体研究已发表[7]. 优化后色谱柱选用Agilent ZORBAX SB-Aq(4.6 mm×250 mm, 5 µm),流动相20 mmol∙L−1柠檬酸和5.0 mmol∙L−1己烷磺酸钠(pH=4.4,甲醇加入量1 %),等度洗脱,流速1.0 mL∙min−1,进样量20 μL. 分离色谱图见图1.
-
结合硒形态分析相关文献[8 − 13],以富硒葛粉为样品,考察水提法、醇提法、酸提法和碱提法提取硒形态效果,其中水提法只对Se(Ⅳ)的提取效果较明显,其他形态提取效率低;醇提法和水提法的提取效率相近;酸提法与前二者相比,Se(Ⅵ)的提取效率反而下降且基线波动大,提取效果极差;碱提法对一些水溶性硒蛋白中硒形态具有较高的提取效率,如Se(Ⅳ). 考察木瓜蛋白酶、蛋白酶XIV、胃蛋白酶、蛋白酶K对样品中硒形态提取效果的影响. 蛋白酶XIV提取Se(Ⅳ)、SeMet效率更高,因此蛋白酶XIV是较优选择. 综上,本研究拟定碱提取和酶提取相结合的提取方法. 但由于蛋白酶XIV在中性条件下活性最强. 因此,硒形态提取分两步进行,先碱提取,提取液中和后,采用蛋白酶XIV伴以水浴摇床辅助提取.
-
考察不同温度对提取效果的影响,分别进行室温(25 ℃)、60 ℃、90 ℃超声提取提取30 min,121 ℃提取30 min,结果显示60 ℃超声时Se(IV)的提取量是室温条件下的1.5倍;升高温度到90 ℃,Se(IV)的提取效率显著提高,约为60 ℃提取量的一倍,121 ℃时硒形态之间发生转化. 因此选择90 ℃,超声30 min为碱提取条件.
-
酶在高温条件下结构易被破坏,活性降低或失活,其最佳提取温度为37 ℃,提取率高且不会破坏活性. 且酶解时间的长短会对提取效果造成影响,时间过少会导致硒形态不能完全提取,时间过长又会导致硒形态之间发生一定的转变. 为考察酶解时间对葛粉中硒形态提取效果的影响,分别进行37 ℃水浴酶解4 h、6 h、8 h和18 h,酶解时间为4 h、6 h、8 h时,有机硒形态中仅有SeMet检出,而酶解时间18 h时不仅检出SeMet,还检出大量的SeEt. 许多植物为避免硒中毒,会通过甲基化的方式转变硒的储存形式[14],因此存在SeEt. SeEt 为 SeMet的衍生物. 结果与已有研究结果一致[15]. 前期在探讨无机硒提取效果时,采取醇提法时,也同时出现了SeMet、SeEt. 葛粉中检出SeEt,这与富硒大蒜、富硒西兰花的检出结果一致[14,16]. 有研究表明[17 − 18],植物基质中含有丰富的酚类物质,Se(Ⅳ)与植物中黄酮类、酚酸类或鞣质类等酚类化合物发生相互作用含量逐渐下降,而SeMet, SeCys2, SeMeSeCys 以及 Se(Ⅵ))等则是稳定的. 本研究将采用先提取无机硒,后提取有机硒的两步法,既保证了无机硒形态和有机硒形态的提取率,又可以避免长时间酶解导致Se(Ⅳ)含量逐渐下降. 考虑到葛粉中含有少量淀粉及脂肪,本实验选择蛋白酶XIV、脂肪酶、淀粉酶各10 mg混合加入,37 ℃水浴提取18 h为酶提取条件.
-
使用超纯水将6种硒形态的混合标准液逐级稀释,配制成浓度为0.5、1.0、2.0、5.0、10.0、20.0、50.0、100、200、500 µg∙L−1的混合标准液系列,以各浓度色谱峰面积对应质量浓度绘制标准工作曲线,相关系数均大于0.9998. 以色谱峰信噪比S/N为3∶1时所对应的浓度计算检出限,得出Se(Ⅵ)、Se(Ⅳ)、SeCys2、MeSeCys、SeMet、SeEt的检出限0.017—0.053 mg·kg−1.
-
对富硒葛粉中6种硒形态进行加标回收试验,Se(Ⅵ)、SeCys2、MeSeCys的加标浓度分别为0.1、0.5、2 mg·kg−1,Se(Ⅳ)、SeMet、SeEt的加标浓度分别为0.5、2.5、10 mg·kg−1. 6种硒形态的加标回收率均在71.2%—117.3%之间,相对标准偏差(n=6)均在10%以下.
-
将本研究建立方法用于测定市面上销售的富硒葛粉,并用国家标准GB/T 5009.93-2017 电感耦合等离子体质谱法测定总硒含量. 结果显示总硒含量为2.26 mg·kg−1,对富硒葛粉进行硒形态分析,最终测出富硒葛粉中主要成分为:Se(IV)占30.8%,SeMet(以SeMet和SeEt之和计)占40.1%,SeCys2占3.39%,MeSeCys占2.64%. 6种硒形态的总提取率为76.9 %. 由于分析过程中出现多个未知硒形态峰,以当前已有的硒形态标准物质无法对其进行定性定量分析,计划在今后实验中运用HPLC-MS-MS对未知硒形态深入研究. 富硒葛粉中主要成分为Se(IV)、SeMet,因此在研究硒形态的抗氧化功能时,选取常见无机硒形态Se(IV)、Se(Ⅵ)以及有机硒形态SeMet,考察三种硒形态对脂多糖诱导的AML12细胞氧化损伤的影响.
-
采用0、0.01、0.1、1、10 μmol·L−1 SeMet处理脂多糖(LPS)诱导的AML12细胞48 h后,0.01、0.1、1 μmol∙L−1 SeMet均能提高AML12细胞活力,且0.1、1 μmol∙L−1 SeMet具有显著性(P<0.05),而10 μmol∙L−1 SeMet显著降低AML12细胞活力(P<0.01). 0.01、0.1、1、10 μmol∙L−1 Se(Ⅵ)、Se(Ⅳ)均显著降低AML12细胞活力(P<0.01). 因此,本研究选择0.01、0.1、1 μmol∙L−1 SeMet作为后续研究剂量.
-
如表1所示,与正常组比较,模型组中MDA含量上升(P<0.01),SOD及GSHx活性皆显著降低(P<0.01);与模型组比较,硒代蛋氨酸3个剂量实验组中MDA含量显著降低(P<0.01),SOD及GSHx活性都显著提高(P<0.01).
-
使用Agilent ZORBAX SB-Aq 反相色谱柱(4.6 mm×250 mm, 5 μm),流动相体系为20 mmoL∙L−1的柠檬酸和2.5 mmoL∙L−1的己烷磺酸钠,0.1 moL∙L−1的NaOH溶液在90 ℃下提取样品30 min,加入蛋白酶XIV、脂肪酶、淀粉酶,37 ℃水浴酶解18 h,运用高效液相色谱-电感耦合等离子体质谱联用技术(HPLC-ICP-MS)快速分离六种硒形态. 6种硒形态的检出限为0.017—0.053 mg∙kg−1,标准曲线的相关系数均在0.9998以上. 该前处理方法测定样品的加标回收率均在71.2 %—117.3 %之间,相对标准偏差均在10 %以内.
分析富硒葛粉中主要硒形态对脂多糖(LPS)诱导的AML12细胞氧化损伤的影响,与模型组相比,1 μmol·L−1硒代蛋氨酸处理组脂多糖诱导的AML12细胞活力显著上升,丙二醛(MDA)含量显著下降,超氧化物歧化酶(SOD)和谷胱甘肽过氧化物酶(GSHx)活性显著增加. 结果提示,硒代蛋氨酸对LPS诱导的AML12细胞氧化损伤具有保护作用,可增强植物的抗氧化损伤能力.
富硒葛粉中硒形态分析及其对脂多糖诱导的AML12细胞氧化损伤的影响
Determination of selenium species in Se-enriched puerarin powder and its effect on oxidative damage of AML12 cells induced by Lipopolysaccharide
-
摘要: 建立了富硒葛粉中硒酸根[Se(Ⅵ)]、亚硒酸根[Se(Ⅳ)]、硒代胱氨酸(SeCys2)、甲基硒代半胱氨酸(MeSeCys)、硒代蛋氨酸(SeMet)、硒代乙硫氨酸(SeEt)等6种硒形态的检测方法并分析了主要硒形态的抗氧化损伤功能 0.1 mol·L−1的NaOH溶液在90 ℃下提取样品30 min,加入蛋白酶XIV、脂肪酶、淀粉酶,37 ℃水浴酶解18 h,高效液相色谱-电感耦合等离子体质谱联用(HPLC-ICP-MS)快速分离测定6种硒形态. 分析主要硒形态对脂多糖(LPS)诱导的AML12细胞氧化损伤的影响. 结果表明,6种硒形态的检出限为0.017—0.053 mg·kg−1,标准曲线的相关系数均在0.9998以上. 该前处理方法样品的加标回收率在71.2%—117.3%,相对标准偏差均在10 %以内. 与模型组相比,1 μmol·L−1硒代蛋氨酸处理组脂多糖诱导的AML12细胞活力显著上升(P<0.01),丙二醛(MDA)含量下降30.1%(P<0.01),超氧化物歧化酶(SOD)活性增加43.9%(P<0.01),谷胱甘肽过氧化物酶(GSHx)活性增加44.7%(P<0.01). 硒代蛋氨酸对LPS诱导的AML12细胞氧化损伤具有保护作用,可增强植物的抗氧化损伤能力.
-
关键词:
- 硒 /
- 形态分析 /
- 高效液相色谱-电感耦合等离子体质谱联用 /
- 抗氧化损伤
Abstract: The objective of this study was to establish a method for the detection of six selenium species in Se-enriched puerarin powder, including Se(Ⅵ),Se(Ⅳ),SeCys2,MeSeCys,SeMet and SeEt. Also this study was to analyze the antioxidant damage of the main selenium species. The sample was extracted with 0.1 mol·L−1 NaOH solution at 90 ℃ for 30 min, then protease XIV, lipase and amylase were added. And enzymolysis was performed at 37 ℃ for 18 h in water bath. Six selenium species were rapidly detected by high performance liquid chromatography-inductively coupled plasma-mass spectrometry (HPLC-ICP-MS). The effect of main selenium species on the oxidative damage of AML12 cells induced by Lipopolysaccharide (LPS) was measured. The results showed that the detection limits for the six selenium species were 0.017—0.053 mg·kg−1, and the correlation coefficients of the standard curves were all above 0.9998. The spiked recoveries of the samples determined by this pretreatment method were all between 71.2%—117.3%, and the relative standard deviations were all within 10%. Compared to the model group, the activity of AML12 cells induced by LPS increased significantly (P<0.01) after being applied to SeMet at a concentration of 1 μmol∙L−1, the content of Malondialdehyde (MDA) decreased by 30.1% (P<0.01), the activity of Superoxide Dismutase (SOD) increased by 43.9% (P<0.01), and the activity of Glutathione Peroxidase (GSHx) increased by 44.7% (P<0.01). The results suggest that SeMet has a protective effect on LPS induced oxidative damage in AML12 cells. So it can enhance the ability of plants on antioxidant damage. -
交替式生物反应池(UNITANK)作为一种结构紧凑、运行灵活的活性污泥工艺,在我国城市污水处理中得到了较广泛的应用[1-2]。然而,由于结构上的不足,UNITANK在运行过程中普遍存在污泥分布不均、低负荷释磷不充分、池体容积布局不合理等问题[3-4]。为了解决这些问题,张发根等[4]提出了双流态UNITANK,即将UNITANK工艺中的1个边池改成2个以上,且交替向中间池提供污泥,他们同时利用ASM2D模型和实验验证了双流态UNITANK工艺的可行性,但这种工艺的运行周期调控相对更为复杂。因此,改良型UNITANK工艺应运而生。改良型UNITANK是在UNITANK的基础上增设了单独的厌氧池和缺氧池,同时增加了内回流点,以强化工艺的脱氮除磷效果。目前国内对改良型UNITANK工艺的研究报道较少。朱海敏等[5]、夏海波等[6]比较了改良型UNITANK工艺与UNITANK工艺的实际运行效果,结果均表明,改良型UNITANK工艺可以获得更高、更稳定的脱氮除磷能力。朱海敏等[5]提出,在处理水量持续超设计负荷15%的情况下,改良型UNITANK出水水质仍可以稳定达到一级A排放标准,且产泥量远低于UNITANK工艺。但是,目前有关改良型UNITANK工艺的报道均未对工艺特性进行探讨。
目前,工艺特性研究普遍基于实验方法开展。然而,受进水、环境条件变化、采样代表性等多因素影响,实验研究面临着局限性,无法全面表征污水处理厂复杂工艺面临的实际问题。污水生物处理系统的数学模拟是利用数学模型类比复杂的生化反应,以寻求其中的过程规律。大量研究[7-8]表明,数学模型可以成功应用于污水处理系统的优化,并指导污水处理系统的运行调控。宋纯金等[9]、董姗燕等[10]、张发根等[4]分别采用数学模拟方法对UNITANK工艺特性进行了模拟分析,证明采用模型研究复杂的交替式工艺的可行性和可靠性,为工艺特性研究提供了新工具。此外,我国城市污水处理厂普遍采用季节性调控策略,大部分污水厂的运行实践表明[11-13],夏季出水水质稳定达标;而冬季,由于水温降低影响微生物的活性,导致系统的生物脱氮除磷能力下降,容易出现出水水质浓度波动大、超标等问题。因此,冬季的运行调控一直是城市污水处理厂运行过程中的难点。本研究以苏州某城市污水处理厂改良型UNITANK工艺为研究对象,针对污水厂冬季普遍存在的运行稳定性差等问题,采用模型分析了改良型UNITANK工艺的周期性运行特征,同时利用高通量测序技术分析了微生物群落组成,旨在为实际污水厂改良型UNITANK工艺冬季运行提供参考。
1. 材料与方法
1.1 改良型UNITANK工艺及处理效果
苏州某城市污水处理厂主要接纳生活污水,设计规模1.2×105 t·d−1,采用4组改良型UNITANK工艺。改良型UNITANK出水依次进入高效沉淀池、气水反冲洗滤池、紫外线消毒池,出水执行《城镇污水处理厂污染物排放标准》(GB 18918-2002)一级A标准。改良型UNITANK工艺冬季进、出水水质如表1所示,冬季出水水质波动较大,运行稳定性仍有待进一步提高。
表 1 苏州某城市污水处理厂改良型UNITANK工艺冬季进出水水质Table 1. Influent and effluent quality of modified UNITANK process in a Suzhou sewage treatment plant in winter水样 COD/(mg·L−1) 氨氮/(mg·L−1) TN/(mg·L−1) TP/(mg·L−1) SS/(mg·L−1) 进水 399.2±109.5 29.0±4.4 41.3±6.5 4.3±1.0 163.0±48.0 出水 44.8±14.0 6.0±3.5 10.6±3.1 0.7±0.6 15.1±8.8 | Show Table DownLoad:
CSV
DownLoad:
CSV
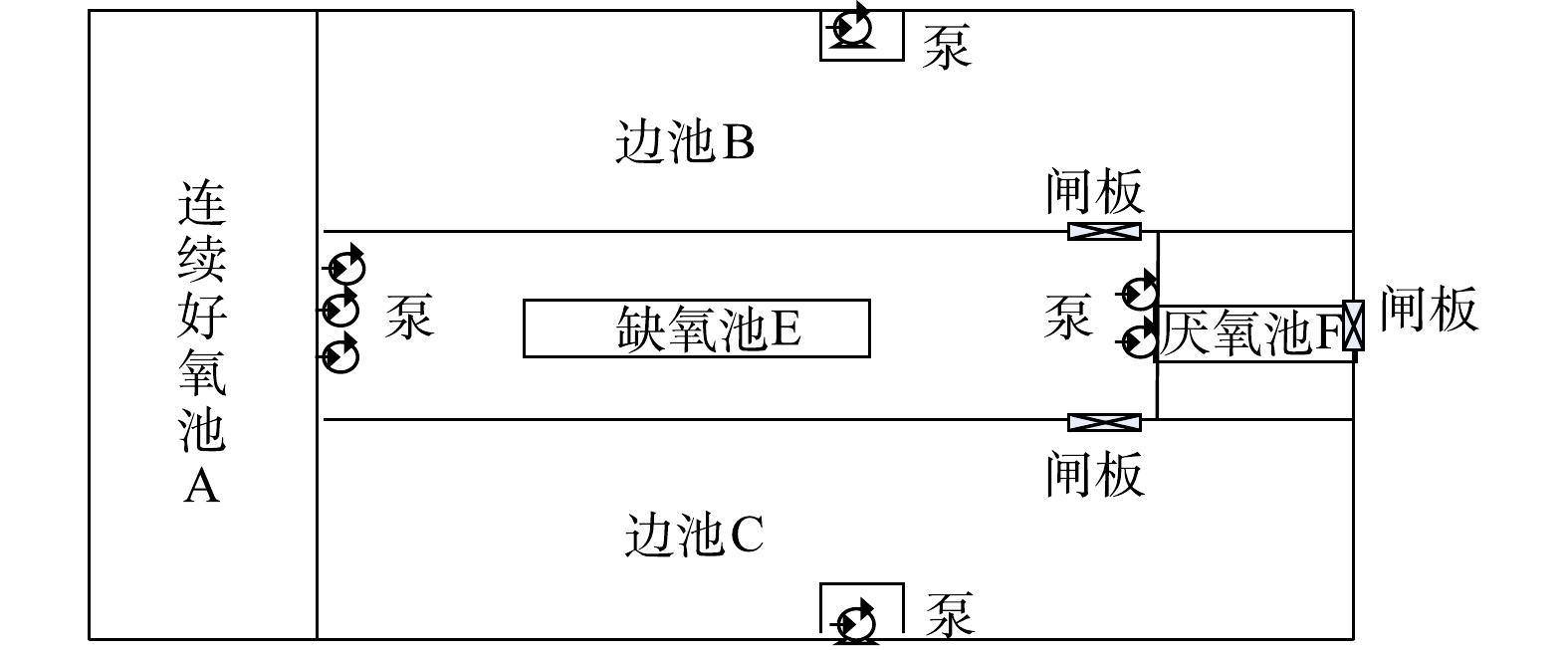
每组改良型UNITANK工艺分隔成5格顺序连通的矩形反应池,分别为1个厌氧池F,1个缺氧池E、1个连续好氧池A、2个边池B和C(图1)。每组平面尺寸为89.85 m×44.85 m,有效水深7.50 m。平均水力停留时间23.72 h,厌氧、缺氧、好氧、边池停留时间分别为1.32、5.63、3.36、13.41 h。厌氧池F与缺氧池E、连续好氧池A与边池之间通过隔墙底部开孔水力连通;缺氧池E至连续好氧池A、缺氧池E到厌氧池F通过泵强制实现混合液流动;边池到缺氧池E通过边池末端底部闸阀控制;剩余污泥由边池两侧的剩余污泥泵定期排放。
改良型UNITANK工艺运行周期为 8 h。上半周期(0~3 h):污水依次进入厌氧池F、缺氧池E、连续好氧池A和边池B(边池B作为曝气池),边池C作为沉淀池(不曝气),出水从边池C通过空气堰排出,剩余污泥从边池C通过泵排放。过渡周期(3~4 h):边池B停止曝气,转换为沉淀模式。污水仍依次进入F、E、A、B,出水从边池C流出。下半周期与上半周期的运行完全一致,通过过渡周期进行衔接。下半周期(4~7 h): B池和C池功能互相转换,边池B作为沉淀池,出水从边池B通过空气堰排出,边池C作为曝气池。过渡周期(7~8 h):边池C停止曝气,转换为沉淀模式。
1.2 样品采集
2020年2月中旬取自改良型UNITANK工艺曝气池末端活性污泥,取3个平行样(标记为M1、M2、M3),以代表系统经过冬季低温后微生物群落的分布特征。样品经过30 min静置并离心(4 ℃、8 000 r·min−1、5 min)后保存于−20 ℃冰箱内,用于后续的分子生物学测定。
1.3 DNA提取,PCR扩增及Illumina MiSeq高通量测序分析
DNA提取采用PowerSoil® DNA Isolation Kit试剂盒,提取后经1%琼脂糖凝胶电泳检测条带完整性。PCR扩增所用引物为338F (5′-ACTCCTACGGGAGGCAGCAG-3′)和806R (5′-GGACTACHVGGGTWTCTAAT-3′)。反应体系为20 μL,PCR扩增管中添加DNA模板10 ng,正反向引物各0.8 μL,灭菌水20 μL,d NTP 2 μL,缓冲液4 μL,FastPfu聚合酶0.4 μL。PCR反应程序:首先95 ℃预变性3 min,然后进行27个循环(95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸45 s),最后72 ℃延伸10 min。扩增结束后,采用2%琼脂糖凝胶电泳检测PCR产物,结果表明,PCR产物条带大小正确,浓度合适,可进行后续实验,委托美吉生物进行Illumina MiSeq高通量测序。
高通量测序获得的原始序列数据经过质控过滤后得到高质量数据,采用USEARCH进行OTU聚类分析,通过贝叶斯算法在97%相似水平对OTU进行物种分类学注释。基于OTU数据,对反映微生物群落丰富度(Sobs, Chao, ACE指数)和多样性(Simpson, Shannon指数)的参数进行统计。
1.4 水质指标与分析方法
氨氮采用纳氏试剂分光光度法测定;
NO−3 PO3−4 1.5 模型构建
以BioWIN软件为平台,选择ASM2D模型为核心机理描述生物碳、氮、磷的去除过程。采用1个非曝气完全混合反应池(CSTR)模拟厌氧池F,4个非曝气CSTR(缺氧池E-1、E-2、E-3、E-4)串联模拟缺氧池E,3个曝气CSTR的串、并联模拟连续好氧池A,5个等体积SBR(边池B-1、B-2、B-3、B-4、B-5/边池C-1、C-2、C-3、C-4、C-5)串联模拟边池B/C的运行,通过分离器的控制实现2组边池的交替运行。
采用该厂改良型UNITANK工艺中2组冬季历史数据进行模型校准和验证,通过调整进水有机物组分(表2)和氨氧化菌最大比生长速率(由默认值0.9 d−1校准为0.7 d−1),改良型UNITANK出水中COD、氨氮、TN、TP、SS模拟值和实测值的绝对误差分别为4.7、1.1、1.6、0.3、2.1 mg·L−1;同时,改良型UNITANK工艺沿程氨氮、
NO−3 SPO3−4 表 2 苏州某城市污水处理厂改良型UNITANK工艺进水有机物组分Table 2. Influent organic matter composition of a modified UNITANK process in a Suzhou sewage treatment plant数值 溶解性不可生物降解COD/(g·g−1) 易生物降解COD/(g·g−1) 发酵产物/(g·g−1) 可发酵的易生物降解COD/(g·g−1) 颗粒性不可生物降解COD/(g·g−1) 颗粒性易生物降解COD/(g·g−1) 校准值 0.05 0.35 0.15 0.85 0.20 0.40 BioWIN推荐值 0.05 0.16 0.15 — 0.13 — | Show TableDownLoad:
CSV
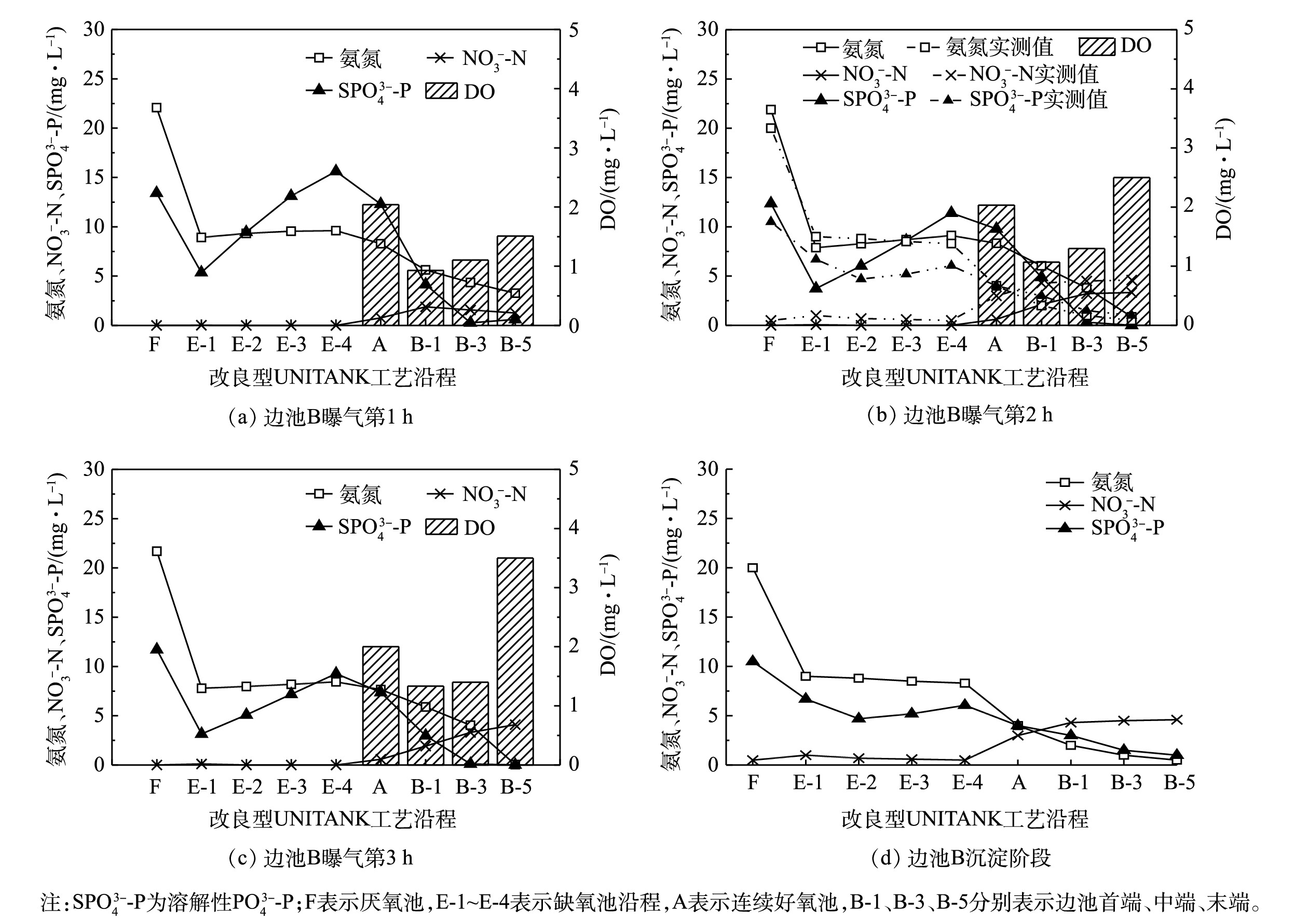 图 2 上半周期改良型UNITANK工艺沿程氨氮、
图 2 上半周期改良型UNITANK工艺沿程氨氮、NO−3 SPO3−4 Figure 2. Changes of ammonium,NO−3 SPO3−4 2. 结果与讨论
2.1 改良型UNITANK工艺运行特性
采用经校准和验证后的模型为工具,模拟冬季低温(12 ℃)条件下,在1个运行周期内,改良型UNITANK沿程氨氮、
NO−3 SPO3−4 NO−3 SPO3−4 由图2(a)可知,在上半周期边池B曝气第1 h,污水首先流入厌氧池F,并与从缺氧池E首端回流的混合液混合,在厌氧池F内利用搅拌形成局部污泥负荷较高的区域,促进磷的释放,因此,厌氧池F内
SPO3−4 SPO3−4 SPO3−4 NO−3 SPO3−4 NO−3 SPO3−4 当边池B处于曝气第2、3 h及沉淀阶段,沿程氨氮、
NO−3 SPO3−4 SPO3−4 NO−3 当工艺进入下半周期,边池B切换为沉淀池,边池B末端通过空气堰出水,改良型UNITANK出水氨氮、
NO−3 SPO3−4 SPO3−4 NO−3 SPO3−4 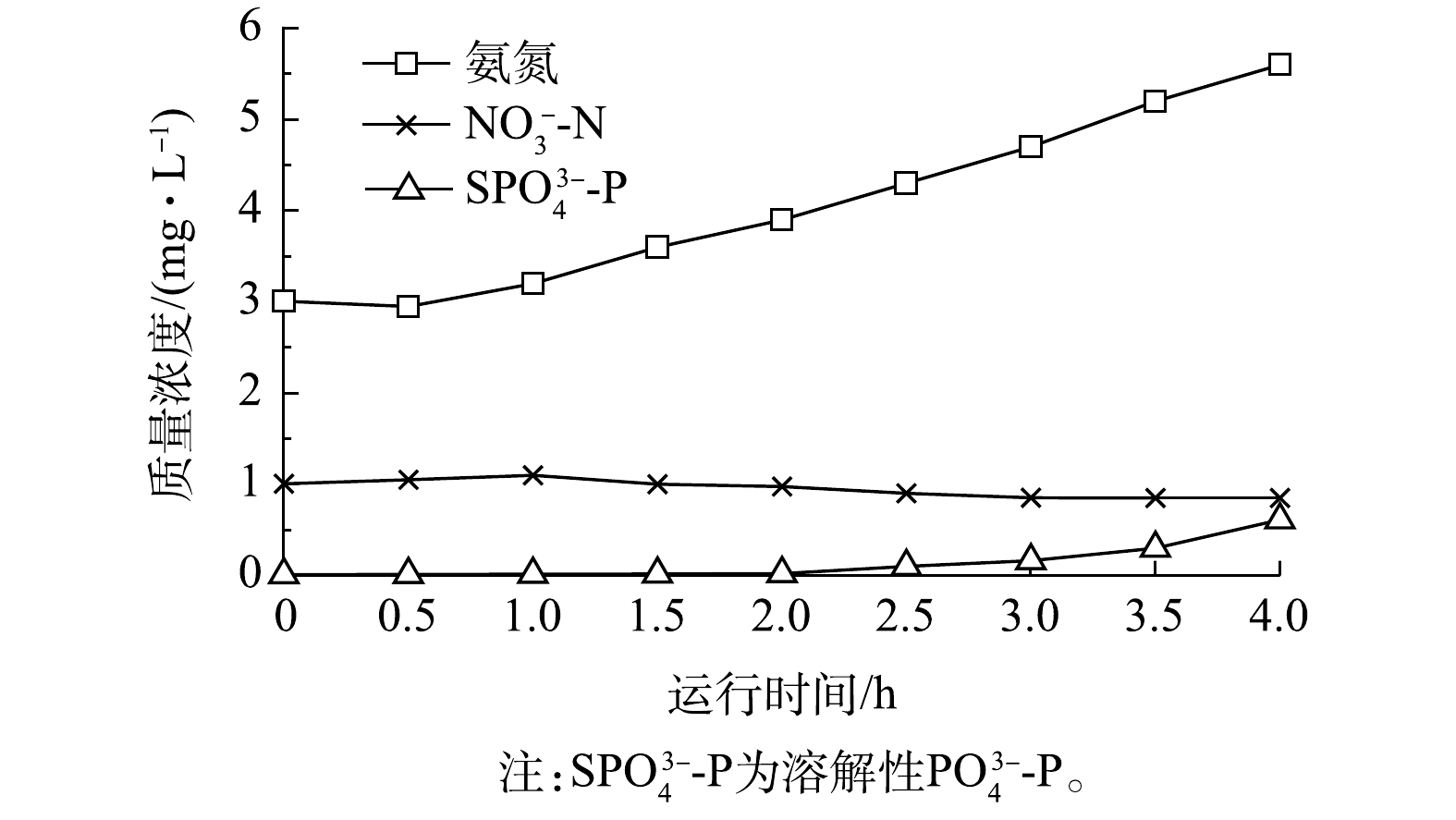 图 3 改良型UNITANK工艺出水水质变化Figure 3. Variation of effluent quality of modified UNITANK process
图 3 改良型UNITANK工艺出水水质变化Figure 3. Variation of effluent quality of modified UNITANK process2.2 活性污泥群落结构分析
1)多样性分析。污水厂改良型UNITANK工艺冬季活性污泥微生物群落的多样性指数见表3所示。3组污泥样品的有效序列为43 774~43 967,经抽平处理后,3组样品的有效序列标准化至43 774。在97%的相似水平上,覆盖率均高于98%,表明本次测序相对于整体样本的覆盖程度极高,测序结果能够较准确地反映改良型UNITANK内冬季的生物特性。3组污泥样品中的Sobs指数为2 017~2 106,略高于张晓红等[14]和韩文杰等[15]的研究结果。张晓红等[14]针对京津冀地区市政污水厂活性污泥种群结构研究发现,5个污水厂AO或AAO工艺中的Sobs指数在1 006~1 965。韩文杰等[15]在长三角地区污水厂低温季节微生物多样性分析中指出,5个污水厂AAO及其变形工艺中Sobs指数为1 014~1 782。因此,该污水厂改良型UNITANK的微生物多样性较高。与张晓红等[14]和韩文杰等[15]对国内其他污水厂的研究结果相比,该厂3组样品的Shannon指数、ACE指数和Chao指数属于较高水平,Simpson指数显著低于文献报道[14-15]的平均水平,证实了生物多样性在改良型UNITANK工艺中较高。ZHANG等[16]提出,进水水质差异是导致系统内微生物多样性的关键因素。而SEIB等[17]发现,除了进水水质,反应器结构也会对污泥群落结构产生影响。本研究中的污水厂进水水质为典型的城市污水,与张晓红等、韩文杰等[14-15]研究的污水厂进水水质处于类似水平。因此,导致该系统生物多样性较高的原因可能与改良型UNITANK独特的工艺运行方式有关,边池交替式的运行控制模式导致微生物多样性较高。郑向阳等[18]发现,微生物多样性与运行条件有关,同一进水条件下,缺氧段多样性明显高于好氧段。彭永臻等[19]在对城市污水处理厂生物脱氮污泥菌群结构分析中发现,更为复杂的运行方式会导致污水处理系统中生物多样性更高。
表 3 改良型UNITANK工艺物种多样性指数及其比较Table 3. Species diversity indices of the modified UNITANK process and their comparison样品 Sobs Shannon Simpson ACE Chao 覆盖率/% M1 2 027 5.936 0.008 2 566 2 606 98.9 M2 2 106 5.964 0.007 2 687 2 681 98.9 M3 2 017 5.909 0.008 2 632 2 716 98.6 | Show TableDownLoad:
CSV
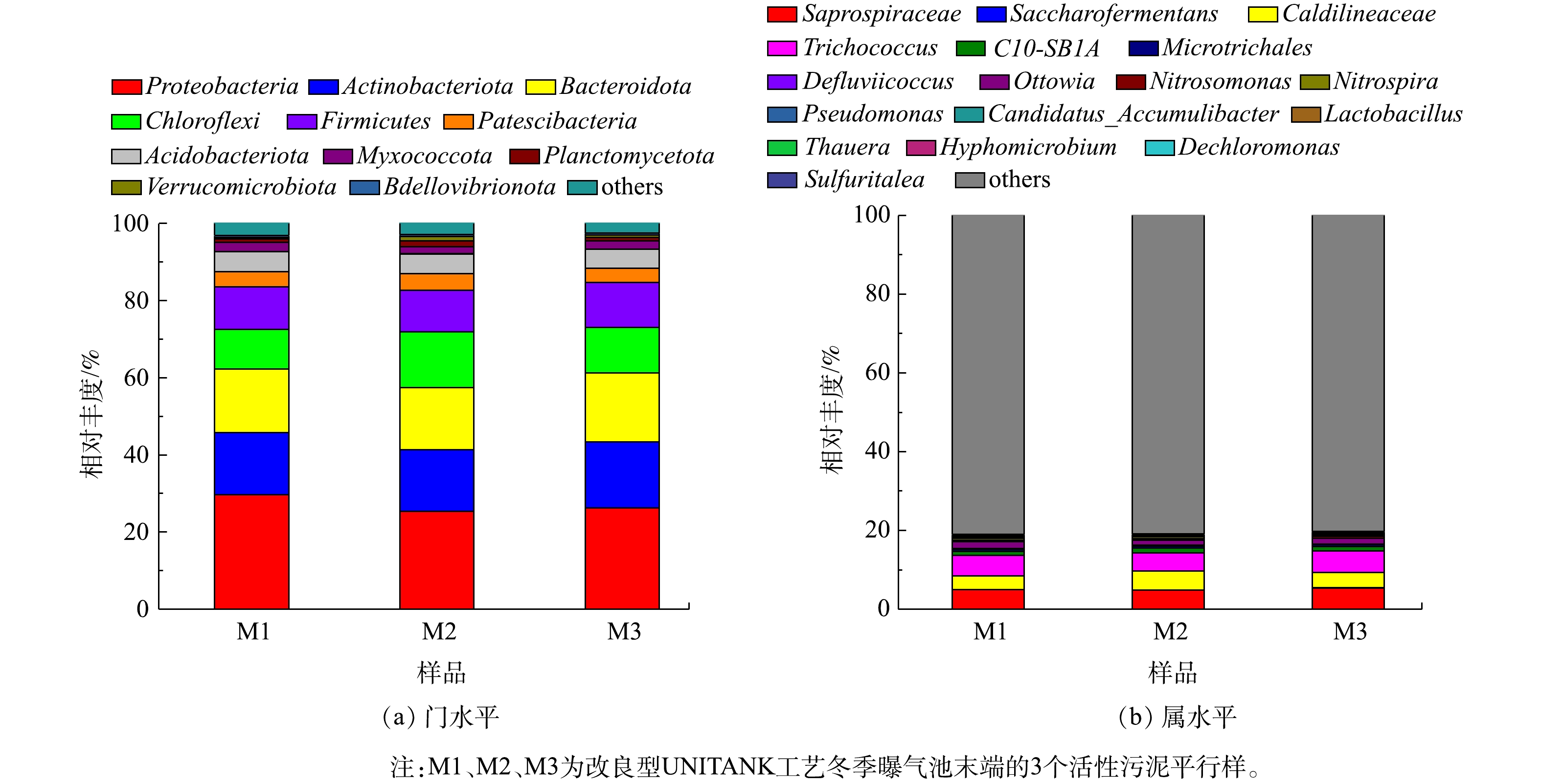
2)物种组成分析。在门水平上,3组污泥样品共检测到48个门,其中,有9种物种的丰度高于1%。由图4可以看出,Proteobacteria、Actinobacteria、Chloroflexi、Bacteroidetes、Firmicutes是3组污泥样品中的主要优势菌群,其总相对丰度达到82.73%~84.76%,与文献报道[14-15]的城市污水厂活性污泥系统中门水平上的微生物组成结论一致。其中,Proteobacteria是样品中丰度最高的菌门,其相对丰度达到25.40%~29.65%;Actinobacteria、Chloroflexi相对丰度分别为15.89%~17.04%、10.31%~14.50%。Proteobacteria和Actinobacteria是生物脱氮除磷和异养生物降解的主要微生物,而Chloroflexi在功能上多与生物除磷有关。王思佳等[20]的研究表明,以乙酸钠为碳源时,会促进Bacteroidetes含量上升。因此,3组污泥样品中门水平上的优势菌群均与生物脱氮除磷功能有关。
在属水平上丰度高于2%的微生物的分析表明,Caldilineaceae、Saprospiraceae、Trichococcus是优势菌属,其相对丰度分别可达3.41%~4.87%、4.83%~5.40%、4.55%~5.52%。 张朝升等[21]的研究表明,Saprospiraceae是以亚硝酸盐为电子受体的反硝化除磷系统中的优势菌种。徐伟峰等[22]的研究表明,延长泥龄,反硝化除磷对系统除磷所起的作用增强。该厂改良型UNITANK工艺冬季的泥龄控制在20 d左右,为Saprospiraceae的生长提供了良好的条件。因此,改良型UNITANK可能存在反硝化除磷现象。有研究[14-15]表明,除了Saprospiraceae,Pseudomonas和Candidatus_Accumulibacter也是污水处理系统中常见的聚磷菌。本研究中,Pseudomonas和Candidatus_Accumulibacter在3组污泥样品中的丰度分别为0.03%~0.05%和0.03%~0.04%,显著低于Saprospiraceae的丰度。Defluviicoccus、Micropruina属于常见的聚糖菌属,在厌氧段会与聚磷菌竞争碳源,从而导致生物除磷效果恶化,这2种菌属在改良型UNITANK工艺冬季污泥样品中的相对丰度仅为0.16%~0.21%、0.51%~0.56%,表明聚糖菌所占比例较低,这与目前的普遍研究结论[13]一致,即在低温条件下,聚磷菌更具竞争优势,而当温度高于20 ℃,聚糖菌的生长占优势。
Nitrosomonas为氨氧化菌(AOB)优势菌属,其功能是将氨氮转化为亚硝酸盐,该菌属在改良型UNITANK工艺内所占比例为 0.31%~0.45%。Nitrospira为亚硝酸盐氧化菌(NOB)优势菌属,其作用是将亚硝酸盐转化为硝酸盐,该菌属所占比例为0.41%~0.60%。在污水处理系统中,常见的NOB菌属包括Nitrospira和Nitrobacter[23]。然而,在改良型UNITANK工艺冬季3组污泥样品中均未检测出Nitrobacter。已有研究[24]表明,Nitrobacter生长速率高,但与底物之间的亲和能力弱,适合生长于基质充足的环境;Nitrospira的生长速率仅为Nitrobacter的1/3,但Nitrospira对基质亲和力更大。因此,在氨氮浓度较低的城市污水处理系统中Nitrospira更具优势。韩文杰等[15]针对长三角地区5座污水厂低温季节微生物检测结果中也指出,在活性污泥系统中Nitrospira为NOB优势菌,相对丰度达0.25%~3.06%,未检测到Nitrobacter。张晓红等[14]在京津冀区域5座城市污水厂的微生物检测中虽发现了Nitrobacter的存在,但其丰度远低于Nitrospira。
3. 结论
1)改良型UNITANK工艺冬季反硝化充分,均匀曝气导致边池前端和中段硝化不充分,在边池的推流作用下,出水氨氮、
SPO3−4 2)改良型UNITANK工艺冬季微生物多样性较高,可能与独特的工艺运行方式有关。Saprospiraceae是优势菌属,其相对丰度可达4.83%~5.40%,工艺内可能存在反硝化除磷现象。Defluviicoccus和Micropruina 2种聚糖菌属的相对丰度仅为0.16%~0.21%和0.51%~0.56%。
3) Nitrosomonas、Nitrospira分别为AOB、NOB的优势菌属,其在改良型UNITANK工艺中的相对丰度分别为0.31%~0.45%、0.41%~0.60%,这2类菌属主要完成了冬季改良型UNITANK工艺脱氮功能。
-
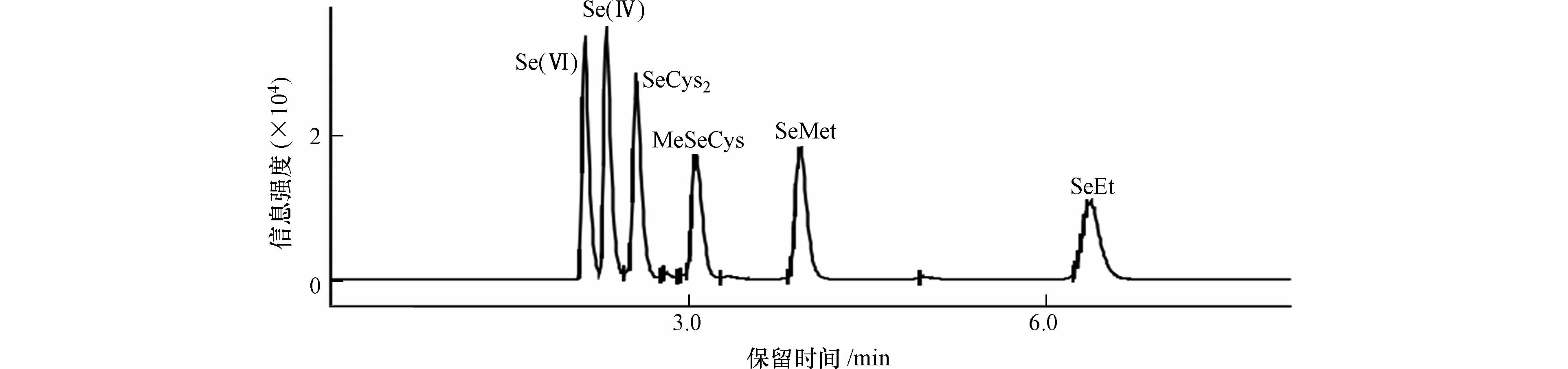
图 1 6种硒形态标准溶液分离色谱图
Figure 1. Separation Chromatograms of standard solutions for six selenium species
表 1 硒代蛋氨酸对LPS诱导的AML12细胞中SOD、GSHx活性及MDA含量的影响(
ˉx±s Table 1. Effects of SeMet on SOD, the activity of GSHx , and the content of MDA in LPS induced AML12 cells(
ˉx±s 组别 MDA/(mmol∙L−1) SOD/(U∙L−1) GSHx/(mg∙L−1) 正常组 5.23±0.54 132.83±11.26 54.28±5.25 模型组 9.41±0.92** 75.32±7.68** 34.47±3.48** 硒代蛋氨酸低浓度组 8.35±0.83## 86.57±8.62## 40.49±3.83## 硒代蛋氨酸中浓度组 7.14±0.72## 98.34±7.83## 45.06±4.12## 硒代蛋氨酸高浓度组 6.58±0.64## 108.36±10.85## 49.87±3.94## (与正常组比较,**P<0.01;与模型组比较,##P<0.01)
下载: 导出CSV
-
[1] KÁPOLNA E, FODOR P. Speciation analysis of selenium enriched green Onions ( Allium fistulosum) by HPLC-ICP-MS[J]. Microchemical Journal, 2006, 84(1/2): 56-62. [2] UDEN P C, BOAKYE H T, KAHAKACHCHI C, et al. Selective detection and identification of Se containing compounds—Review and recent developments[J]. Journal of Chromatography A, 2004, 1050(1): 85-93. doi: 10.1016/j.chroma.2004.05.077 [3] KLEIN E A, THOMPSON I M, LIPPMAN S M, et al. SELECT: The selenium and vitamin E cancer prevention trial[J]. Urologic Oncology: Seminars and Original Investigations, 2003, 21(1): 59-65. doi: 10.1016/S1078-1439(02)00301-0 [4] DUMONT E, VANHAECKE F, CORNELIS R. Selenium speciation from food source to metabolites: A critical review[J]. Analytical and Bioanalytical Chemistry, 2006, 385(7): 1304-1323. doi: 10.1007/s00216-006-0529-8 [5] DERBENEVA S, BOGDANOV A, POGOZHEVA A V, et al. Effect of diet enriched with selenium on the psycho-emotional and adaptive capacity of patients with cardiovascular diseases and obesity[J]. Vopr Pitan, 2012, 81(4): 35-41. [6] 朱磊, 胡婷, 刘德明, 等. 叶面喷施硒对萝卜硒吸收及抗氧化能力的影响[J]. 江苏农业科学, 2019, 47(3): 127-131. doi: 10.15889/j.issn.1002-1302.2019.03.032 [7] 陆秋艳, 张文婷, 林秋莲, 等. 高效液相色谱-电感耦合等离子体质谱联用快速同时分析水中5种砷和7种硒[J]. 环境化学, 2018, 37(7): 1671-1674. [8] 秦冲, 施畅, 万秋月, 等. HPLC-ICP-MS法测定富硒小麦中硒的形态[J]. 食品研究与开发, 2019, 40(2): 140-144. doi: 10.3969/j.issn.1005-6521.2019.02.026 [9] 曾凤泽, 姚宇泽. 微波辅助酶萃取-高效液相色谱-电感耦合等离子体质谱法测定灵芝中6种硒形态[J]. 理化检验-化学分册, 2020, 56(11): 1152-1157. [10] 黄韬睿, 王鑫, 王小平. 反向离子对高效液相色谱-电感耦合等离子质谱联用分析富硒食品中硒的形态[J]. 中国调味品, 2020, 45(11): 140-144. doi: 10.3969/j.issn.1000-9973.2020.11.028 [11] 冯洪燕, 杨从发, 钱亮亮, 等. 市售富硒食品中硒形态及含量的分析[J]. 食品安全质量检测学报, 2021, 12(12): 5059-5065. doi: 10.19812/j.cnki.jfsq11-5956/ts.2021.12.051 [12] 林樾, 陈尚卫, 虞锐鹏, 等. 高效液相色谱-电感耦合等离子体质谱法测定富硒碎米荠中的硒形态[J]. 分析科学学报, 2021, 37(5): 637-642. doi: 10.13526/j.issn.1006-6144.2021.05.011 [13] 孟莉, 许亚丽, 夏曾润, 等. 高效液相色谱-电感耦合等离子体质谱法测定谷类食品中的4种硒形态[J]. 分析科学学报, 2021, 37(6): 843-846. doi: 10.13526/j.issn.1006-6144.2021.06.020 [14] 刘为, 尹金晶, 吴慕慈, 等. 富硒农产品中硒代氨基酸形态及其在不同蛋白组分中的分布[J]. 食品与机械, 2022, 38(6): 45-51, 190. [15] 吴雅颖. 不同外源硒在雷竹笋的富集分布及抗氧化性研究[D]. 杭州: 浙江农林大学, 2014. [16] 张珂, 张钦龙, 张蜀, 等. 高效液相色谱-电感耦合等离子体串联质谱法测定富硒大蒜中硒形态[J]. 中国食品卫生杂志, 2021, 33(5): 577-582. doi: 10.13590/j.cjfh.2021.05.010 [17] CUDERMAN P, STIBILJ V. Stability of Se species in plant extracts rich in phenolic substances[J]. Analytical and Bioanalytical Chemistry, 2010, 396(4): 1433-1439. doi: 10.1007/s00216-009-3324-5 [18] 叶梅. 典型富硒植物中含硒化合物的形态分析与结构鉴定[D]. 无锡: 江南大学, 2022. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 1152
- HTML全文浏览数: 1152
- PDF下载数: 24
- 施引文献: 0
