


















-
新烟碱类杀虫剂化学成分是一种类似于尼古丁结构的化合物,其通过作用于昆虫神经系统中烟碱型乙酰胆碱酯酶受体,阻断昆虫中枢神经系统信号的传导,从而导致害虫麻痹并死亡[1]。自1991年吡虫啉上市以来,新烟碱类杀虫剂迅速发展成为全球第一大类杀虫剂。随着新烟碱类杀虫剂的不断使用,其对环境的危害也进一步凸显出来。一些研究[2-5]表明,新烟碱类杀虫剂不仅对非目标生物(包括蜜蜂、鸟类、水生昆虫等)造成致死作用,而且对人类和哺乳动物的健康也会造成潜在的损害。关于环境中新烟类杀虫剂的迁移转化及去除研究已陆续开展,主要包括化学还原、微生物降解、光催化等,但是存在产物毒性强、降解速率慢、降解产物复杂等方面不足[6-8]。因此,如何高效安全地去除环境中残留的新烟碱类杀虫剂是目前亟待解决的问题之一。
吸附作为去除环境中污染物的有效方法之一,特别是采用环境友好型低成本的吸附剂,由于其使用简单、适用性强、操作灵活、效率高而被广泛应用[9]。生物炭是在缺氧条件下,将生物质经高温裂解产生的富碳产物,具有比表面积大、孔隙发达和含氧官能团丰富等优点,对环境中大部分污染物有良好的吸附和去除效果[10-14]。目前,研究较多的是人工制备的生物炭,对自然环境中生成的生物炭的研究相对较少。自然界中的森林火灾等在一定条件下也能够产生生物炭,且能长时间在自然生态系统中存在,其对环境中污染物迁移转化有着持久的影响[15]。基于此,本研究选用亚热带栽培广、生长快的杉木为原料,通过实验室马弗炉和自然环境下分别制备了2种生物炭,考察了2种生物炭对新烟碱类杀虫剂(啶虫脒(ACE),噻虫胺(CLO))的吸附性能及影响因素,比较了2种生物炭的吸附性能,为新烟碱类杀虫剂在环境中的迁移转化及污染修复提供参考。
-
ACE(C10H11CIN4, >97.0%)和CLO(C6H8CIN5O2S, >97.0%)均购自上海阿拉丁生化科技股份有限公司,2种农药的基本理化性质参数见表1。NaNO3为分析纯,购自无锡市晶科化工有限公司;Ca(NO3)·4H2O为分析纯,购自上海阿拉丁生化科技股份有限公司;HNO3为分析纯,购自无锡市展望化工试剂有限公司。实验用水均为超纯水。
-
高效液相色谱仪(e2695,美国Waters),超纯水装置(Millipore Integral 3,美国Milli-Q),组合式恒温摇床(ZQZY-BF8,上海知楚),马弗炉(SX2-8-10N/NP,上海一恒),型材切割机(355切割机,浙江欧莱德),pH计(FE28,瑞士Mettler Toledo)。
-
本实验的生物炭原料为杉木,生物炭分别采用2种方法制备。方法1:将木材经型材切割机切割成方形小薄块,放入150 mL石英坩埚内,盖上盖子,外层用锡箔纸包裹,置于马弗炉中,于700 ℃条件下高温热解6 h,冷却24 h后取出,在研钵中研磨过100目筛,装于自封袋中备用。将此生物炭标记为BC1。方法2:将木材经型材切割机切割成木块,置于自然环境中燃烧,冷却后刮去表层白色飞灰,研磨过100目筛,装于自封袋中备用。将此生物炭标记为BC2。
采用扫描电子显微镜(SU8100,日本Hitachi)观察BC1和BC2表面形貌;采用傅里叶红外光谱仪(Cary660,美国Agilent)分析生物炭表面官能团;采用全自动比表面和孔隙度测定仪 (Mini Ⅱ,日本Belsorp)测定BC1和BC2的比表面积(SBET)、孔体积和孔径分布。
-
所有实验在250 mL的具塞锥形瓶中进行,反应总体积为100 mL,2种新烟碱类杀虫剂母液浓度为100 mg·L−1,使用0.01 mol·L−1 HNO3和NaOH溶液调节溶液pH,使用1.0 mol·L−1 NaNO3溶液调节离子强度。吸附反应在组合式恒温摇床中进行,摇床转速为200 r·min−1。采用5 mL的针筒注射器取样,经0.45 μm的混合纤维素滤膜过滤后,使用高效液相色谱检测溶液中剩余污染物浓度。
在进行吸附动力学实验时,溶液初始pH为5.0,温度为25 ℃,2种新烟类杀虫剂的浓度初值均为10 mg·L−1,生物炭添加量为0.3 g·L−1,取样时间分别为1.0、2.0、5.0、10、15、20、30、45、60、90、120 min。测定各时间点时溶液中剩余污染物浓度,用准一级动力学模型和准二级动力学模型对实验数据进行拟合,每组实验设置3个平行组和1个空白对照组。
在进行吸附等温实验时,2种新烟类杀虫剂的浓度分别为3.0、5.0、8.0、10、15、18、20 mg·L−1,生物炭添加浓度为0.3 g·L−1,温度分别为15、25和35 ℃。吸附24 h后取样,测定溶液中剩余污染物浓度,用Langmuir和Freundlich模型拟合实验数据。
在考察初始pH和离子强度对吸附的影响时,由于pH≤9,24 h内,ACE基本不会发生水解[19],因此,本实验使用0.01 mol·L−1 HNO3和0.01 mol·L−1 NaOH分别将ACE、CLO溶液初始pH调节为5.0、6.0、7.0、8.0、9.0和3.0、5.0、7.0、9.0、11.0,以考察初始pH对吸附效果的影响。用1.0 mol·L−1的NaNO3和Ca(NO3)2调节溶液中金属离子浓度分别为0、0.01、0.1和0.5 mol·L−1,以考察离子强度对吸附效果的影响,其余条件与吸附动力学实验一致。
-
ACE、CLO采用高效液相色谱检测,吸收波长分别为245 nm和265 nm,色谱柱为XBridge@C18(250 mm×4.6 mm,5 μm),柱温为30 ℃,流动相甲醇与水的体积比分别为70∶30和45∶55。生物炭对ACE和CLO的去除率(R)和吸附量(qe)分别根据式(1)和式(2)进行计算。
式中:R为生物炭对ACE和CLO的去除率;qe为生物炭平衡吸附量,mg·g−1;C0和Ce分别为2种新烟碱类杀虫剂的初始浓度和平衡浓度,mg·L−1;V为反应溶液体积,L;m为生物炭添加量,g。
在吸附动力学实验中,采用准一级动力学方程(式(3))、准二级动力学方程(式(4))和内部扩散方程(式(5))对吸附过程进行拟合[20]。
式中:qt和qe分别为ACE和CLO在t时刻和吸附平衡时新烟碱类杀虫剂被吸附的量,mg·g−1;k1为准一级吸附动力学速率常数,min−1;k2为准二级吸附动力学速率常数,g·(mg·min)−1;k3为颗粒内扩散速率常数,mg·(g·min1/2)−1。
在等温吸附实验中,采用Langmuir方程(式(6))和Freundlich方程(式(7))吸附模型对吸附过程进行拟合[21]。
式中:qe和qm分别为生物炭吸附平衡和最大吸附量,mg·g−1;Ce为吸附平衡时的浓度,mg·L−1;KF和KL分别是Langmuir和Freundlich方程的相关系数;1/n为Freundlich的经验系数。
在吸附热力学实验中,利用式(8)~式(10)计算吸附过程中的自由能ΔG0、焓变ΔH0和熵变ΔS0[22]。
式中:R为理想气体摩尔常数,取值8.31 J·(mol·K)−1;T为热力学温度,K;Kd为吸附平衡常数。
-
图1是BC1和BC2的微观表面形貌图。可以看出,厌氧热解和自然环境下燃烧所形成的炭材料均保持着杉木的自然结构,在形态上无显著差异,杉木细胞空腔呈现出蜂窝状,破碎的导管在BC1和BC2表面形成了大量类似于隧道一样的沟壑,且沟壑中沉积着许多片状颗粒。
采用N2吸附-脱附法分析BC1和BC2的比表面积和孔隙结构,结果如图2(a)和图2(b)所示。根据国际纯粹与应用化学联合会(IUPAC)提出的6种物理吸附等温线[23],BC1和BC2均属于Ⅰ型吸附等温线,表明2种生物炭均具有微孔结构,N2在生物炭表面发生微孔填充现象。BC1吸附-脱附曲线形成了闭合回滞环,属于H4型回滞环。该型回滞环多出现在微孔和中孔混合的吸附剂上和含有狭窄的裂隙孔的固体中。BC2吸附-脱附曲线未形成闭环,这可能是由于BC2中热解未完全的有机物的作用。对比两者的BJH孔径分布曲线图,BC1的孔径主要分布在0~10 nm,没有出现明显的峰,说明孔径分布较均匀,主要以微孔结构为主。BC2的孔径主要分布在0~8 nm,且在2.37 nm时出现尖锐的峰,说明BC2主要含有2 nm左右的孔隙结构,微孔结构更小。根据表2中的数据可知,BC1的BET比表面积明显大于BC2,分别为440.27 m2·g−1和329.15 m2·g−1,主要原因可能是由于马弗炉的热解温度较高且稳定,自然开放环境下烧制的温度差异较大,导致部分生物炭热解不完全。BC1和BC2的平均孔径均在2 nm左右,但是,BC1总孔容大于BC2,这有利于BC1对污染物的吸附。
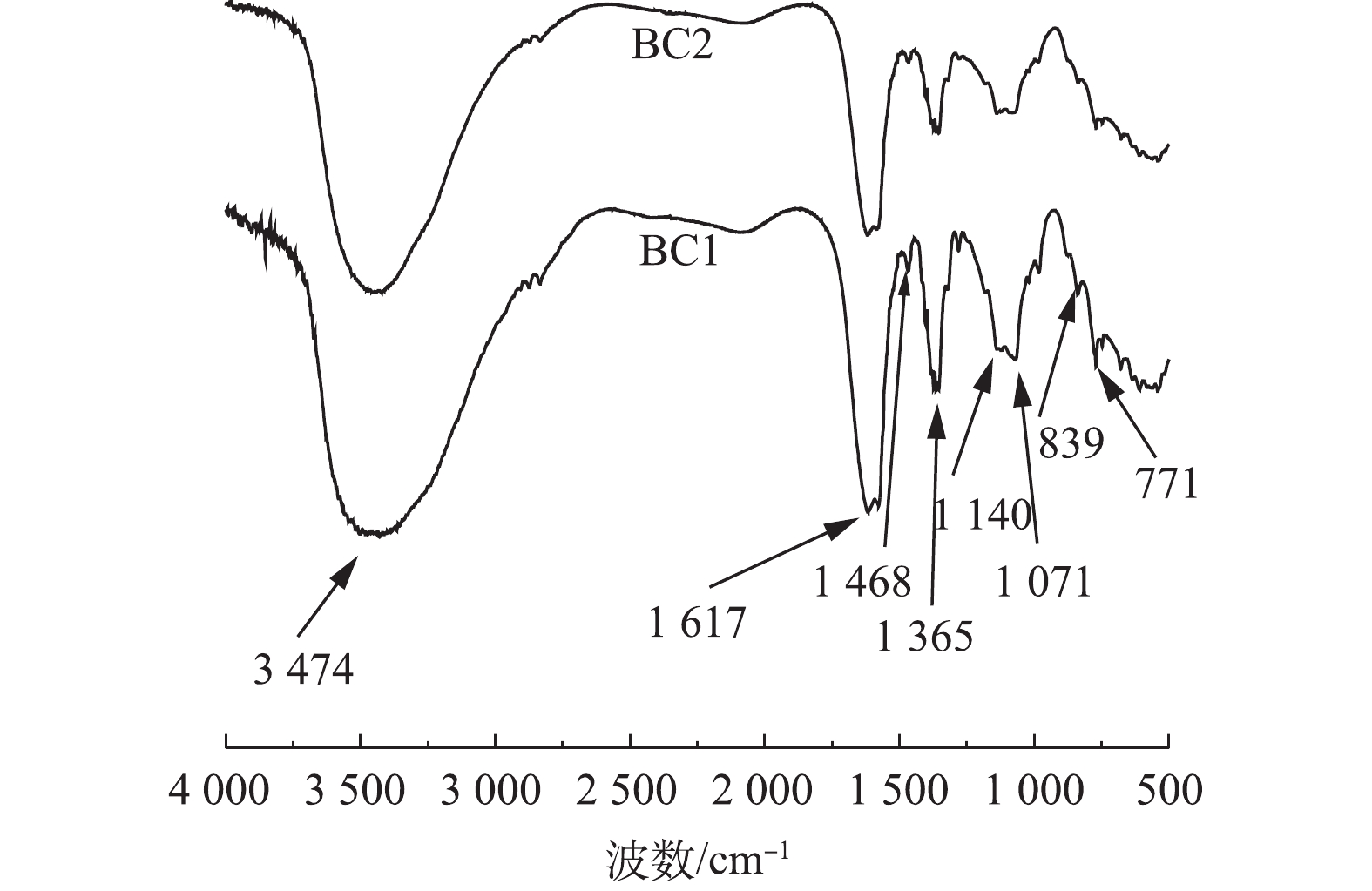
图3是BC1和BC2的傅里叶红外光谱图。可以看出,BC1和BC2具有许多相似的特征峰,这表明BC1和BC2表面含有丰富的官能团。3 474 cm−1处的宽峰是酚羟基(—OH)的伸缩振动峰,1 617 cm−1和1 468 cm−1处的吸收峰是芳香环中的C=O、C=C的伸缩振动峰,1 365、1 140和1 071 cm−1处的吸收峰可以归因于羧基(—OH)的弯曲模式和C—O拉伸振动,839 cm−1和771 cm−1附近的吸收峰主要是由于C—H伸缩振动[12,24-26]引起的。由此得出,BC1和BC2上主要的含氧官能团均为—OH、C=O、—COOH,此外,通过对比BC1和BC2峰强度的大小,可以看出,BC1在1 365、1 140和1 071 cm−1处的吸收峰强度高于BC2,这说明BC1表面的羧基数量可能大于BC2的羧基数量。
-
BC1和BC2对2种新烟碱类杀虫剂的吸附动力学曲线见图4。2种生物炭对溶液中的新烟碱类杀虫剂呈现出相近的吸附动力学过程。生物炭对杀虫剂的吸附量在短时间内(10 min)随着时间的增加迅速增大,之后吸附量增加的速度逐渐变缓,在45 min后,吸附量趋于稳定,此现象与生物炭表面的活性位点的有限性有关。在吸附初始阶段,生物炭表面的活性位点充分,目标污染物能够迅速占据活性位点而呈现出吸附量的急速上升;随着吸附的继续进行,可供吸附的活性位点逐渐减少,进而吸附速率开始下降,最终导致吸附趋于平衡[27]。
为进一步评价2种生物炭的吸附特性及其速率控制步骤和机理,采用准一级动力学方程和准二级动力学方程对2种生物炭吸附新烟碱类杀虫剂的动力学过程进行拟合,结果见表3。由表3可知,准二级动力学的R2均大于准一级动力学,且准二级动力学计算出的理论值qe更加接近于实验测得的数据,这说明准二级动力学方程能够更好地拟合BC1和BC2对2种杀虫剂的吸附过程。BC1的吸附速率普遍小于BC2,但是吸附容量大于BC2。这可能是由于BC1具有更高的孔容,污染物在孔隙中的吸附达到平衡需要更长的时间,总吸附容量也较大,BC2的孔容较小,污染物能够较快地达到吸附平衡,导致吸附容量相对有限。
颗粒内扩散模型反映了吸附过程中的实际控速步骤和吸附机理[28]。由于实验条件及吸附材料和目标污染物的理化性质不同,吸附过程通常可分为吸附2阶段过程和吸附3阶段过程[29-30]。吸附2阶段过程机理为污染物通过液膜扩散到吸附剂表面和污染物在吸附剂表面发生吸附的2个过程。吸附3阶段过程机理为污染物跨水膜扩散到吸附剂表面、污染物在吸附剂孔隙内扩散和吸附趋平衡的3个过程。本研究中生物炭对新烟碱类杀虫剂的吸附过程存在3个阶段,结果如图5所示,线性拟合数据见表4。整体上,BC1吸附过程中的3个阶段较BC2更加明显,BC2的阶段2和阶段3相对模糊,可能是由于BC1具有相对较大的平均孔径和总孔容,致使在阶段2中,新烟碱类杀虫剂于BC1孔隙内扩散积累的量大于BC2所造成。每种生物炭吸附拟合的斜率大小顺序均为k1>k2>k3,这说明污染物向吸附剂的扩散是速率逐渐减小直至达到吸附平衡的过程。BC1和BC2在阶段1中的模型拟合程度均较低,这可能由于该阶段污染物跨水膜扩散到吸附剂表面的作用复杂,影响因素较多所致。2种生物炭在阶段2中的拟合程度最优,R2接近1,这可能是因为孔隙中的环境较稳定,污染物在吸附剂孔隙内能够平稳的扩散所导致。模型对BC1和BC2的阶段3拟合差异最大,BC2的阶段3数据拟合程度明显低于BC1,表明新烟碱类杀虫剂在热解程度较低的生物炭上达到吸附平衡的过程更为复杂。
-
为进一步了解BC1和BC2对ACE、CLO的吸附容量和吸附机理,采用Langmuir和Freundlich 2种吸附模型对数据进行拟合,结果见图6和表5。可以看出,随着ACE和CLO初始浓度的增加,BC1和BC2的吸附量均呈现出先上升后趋于稳定的趋势。此外,随着温度的上升,BC1和BC2的吸附量也同样有所增加,对于BC1,Langmuir模型拟合的相关系数均高于Freundlich,且在3个温度条件下呈现相同的大小关系,说明BC1对2种杀虫剂的吸附属于单分子层吸附,温度的变化对杀虫剂在BC1上的吸附形式没有影响。对于BC2,Freundlich模型拟合的相关系数与Langmuir拟合相关系数并没有一致的大小关系,且随着温度的变化,2种模型相关性系数的大小关系也产生了变化。这说明BC2对目标污染物的吸附存在单分子层和多分子层吸附,温度变化对吸附形式具有一定的影响。BC1对每种杀虫剂的最大理论吸附量均高于BC2,呈现出更好的吸附能力,2种生物炭对杀虫剂的最大吸附量均为qm(CLO)>qm(ACE),此结果与动力学实验一致。KL和KF可指示吸附能力的强弱,KL和KF越大,吸附剂的吸附能力越强。BC1的吸附参数大于BC2,这说明马弗炉高温热解比自然环境烧制的生物炭吸附能力更强。Freundlich拟合模型常数1/n均为0~0.2,这说明BC1和BC2均能较好地吸附2种新烟碱类杀虫剂[31]。
通常使用热力学参数来描述生物炭对新烟碱类杀虫剂的吸附过程中热量的变化情况。在15、25和35 ℃下,2种生物炭吸附ACE和CLO的相关热力学参数见表6。在所选温度下,吉布斯自由能ΔG0均为负值,表明反应是自发进行的。ΔG0在所选温度下基本保持恒定,可见2种生物炭对新烟碱类杀虫剂的吸附受温度影响较小。ΔH0>0,且BC1在吸附过程中的ΔH0明显大于BC2,说明吸附过程需吸收热量,BC1对目标污染物具有更强的吸附能力。ΔS0>0表明在固液界面处生物炭的结合位点上被吸附的2种新烟碱类类杀虫剂的自由度降低,这表明目标污染物分子与生物炭表面活性位点具有较强的结合能力[32]。
-
pH是影响吸附效果的重要因素之一,其可影响目标污染物的解离程度和生物炭的表面电荷[33]。由表1可知,2种新烟碱类杀虫剂的解离常数存在很大差异,ACE无pKa,CLO的pKa为11.1,即在pH为5~9和pH为3~11的范围内,ACE、CLO分别以分子、阳离子的形式存在于水溶液中。初始pH对BC1和BC2吸附ACE、CLO的影响如图7(a)和图7(b)所示。由吸附实验结果可见,随着初始pH的上升,2种生物炭的吸附量均未发生明显变化,为10%~20%,这说明2种生物炭均能在较宽的pH范围内对新烟碱类杀虫剂有良好的吸附效果。溶液初始pH和平衡pH的变化如图8(a)和图8(b)所示,当初始pH为11时,吸附能力的小幅度下降可能是由于吸附平衡后溶液体系为碱性,钝化了生物炭表面的活性位点和减弱了BC1和BC2与CLO分子中O、N原子间的氢键作用力,从而导致生物炭吸附能力的下降[20, 34]。
-
溶液离子强度对BC1和BC2去除2种新烟碱类杀虫剂的影响结果如图9所示。由图9可知,随着Na+和Ca2+浓度的增加,BC1和BC2的吸附容量均有所下降。当Na+离子浓度从0增加到0.5 mol·L−1时,BC1对ACE、CLO的吸附量分别下降了11.29%和5.74%,BC2对ACE、CLO的吸附量分别下降了10.80%和11.44%。Ca2+离子浓度从0增加到0.5 mol·L−1时,BC1对ACE、CLO的吸附量分别下降了20.07%和16.24%,BC2对ACE、CLO的吸附量分别下降了27.66%和18.69%。由图9可知,Na+和Ca2+的添加对2种生物炭吸附新烟碱类杀虫剂均有一定的抑制作用,且Ca2+的抑制作用大于Na+。根据离子强度的定义,相同浓度的Ca2+呈现的离子强度是Na+的4倍,且Ca2+可能会更有效地占据生物炭表面的活性位点[35]。
-
1) 自然条件下烧制的生物炭(BC2)比表面积小于马弗炉烧制的生物炭(BC1),分别为329.15 m2·g−1和440.27 m2·g−1。BC1的总孔容大于BC2,两者官能团种类基本一致,BC1的官能团含量高于BC2,这表明实验室条件下的生物炭较自然条件下的生物炭具有更优的结构和性质,有利于其对污染物的吸附和去除。
2) BC1对2种新烟碱类杀虫剂的吸附能力明显高于BC2。在25 ℃,pH=5时,BC1和BC2对ACE和CLO最大吸附量分别为24.46、31.56、11.13、12.24 mg·g−1。吸附过程更好地符合准二级动力学模型,2种新烟碱类杀虫剂在BC1上发生的是单分子层吸附,在BC2上存在单分子层和多分子层吸附。孔隙结构发达的BC1比BC2呈现出更加明显的跨水膜扩散、孔隙内扩散和吸附平衡3阶段过程。
3) 热力学实验结果表明,2种生物炭对新烟碱类杀虫剂的吸附是自发、吸热的过程。初始pH的升高和离子强度的增加均抑制了BC1和BC2对新烟碱类杀虫剂的吸附作用。
杉木生物炭对水中啶虫脒和噻虫胺的吸附特性
Adsorption properties of acetamiprid and clothianidin in water by Cunninghamia lanceolata biochar
-
摘要: 自然界中生物炭有多种产生途径,影响污染物的迁移转化。为比较实验室和在自然条件下生成的生物炭的吸附性能,以杉木为原料,分别于马弗炉(700 ℃)和自然开放环境中制备了2种生物炭(分别标记为BC1和BC2)。运用氮吸附(BET)、扫描电子显微镜(SEM)和红外光谱(FT-IR)表征分析了生物炭的结构与性质,研究了其对2种新烟碱类杀虫剂(啶虫脒(ACE)和噻虫胺(CLO))的吸附行为,分别考察了初始pH、温度和共存离子对吸附行为的影响。结果表明,700 ℃下裂解制备的BC1吸附能力明显优于自然条件下制备的BC2。BC1对ACE和CLO最大吸附量分别为24.46 mg·g−1和31.56 mg·g−1,BC2对ACE和CLO最大吸附量分别为11.13 mg·g−1和12.24 mg·g−1。BC1和BC2对2种新烟碱类杀虫剂的吸附过程较好地符合准二级吸附动力学模型。颗粒内扩散模型分析结果表明,BC1的吸附较BC2存在更明显的3阶段过程。Langmuir和Freundlich模型拟合结果表明,BC1对2种杀虫剂的吸附属于单分子层吸附,BC2的吸附过程同时存在单分子层和多分子层吸附。热力学研究表明,BC1和BC2对新烟碱类杀虫剂的吸附为自发的吸热过程。随着初始pH的升高和离子强度的增加抑制了生物炭的吸附能力,相同浓度Na+的抑制作用小于Ca2+。以上结果可为水中新烟碱类杀虫剂的去除提供参考。Abstract: Biochar can be formed by many processes in nature and affect the migration and transformation of pollutants in environment. In order to compare the adsorption performance of biochar formed under laboratory and natural conditions, two types of Cunninghamia lanceolata biochar were prepared in a muffle furnace at 700 ℃ and open environment, respectively, which were marked as BC1 and BC2. The structures and properties of these two types of biochar were characterized by nitrogen adsorption (BET), scanning electron microscope (SEM), and Fourier transform infrared spectroscopy (FTIR). The adsorption behaviors of chosen neonicotinoid insecticides (acetamiprid (ACE), clothianidin (CLO)) on biochar were studied, and the effects of initial pH, temperature and coexisting ions on the adsorption behaviors were also investigated. The results showed that the adsorption capacity of BC1 was much greater than BC2, the maximum adsorption capacities of BC1 toward ACE and CLO were 24.46 mg·g−1 and 31.56 mg·g−1, respectively, while the maximum adsorption capacities of BC2 toward ACE and CLO were 11.13 mg·g−1 and 12.24 mg·g−1, respectively. The adsorption kinetics of ACE and CLO on BC1 and BC2 fitted well with the pseudo-second order adsorption model. The intraparticle diffusion model analysis indicated that a more significant three-stage process occurred for BC1 adsorption than BC2 adsorption. The fitting results of Langmuir and Freundlich model showed that the adsorption of two insecticides on BC1 belonged to a single molecular layer adsorption, and on BC2 belonged to both single and multi-molecular layer adsorptions. The thermodynamics experiments indicated that the adsorption processes of neonicotinoid insecticides on BC1 and BC2 were Spontaneous endothermic ones. With the increase of initial pH and ionic strength, the adsorption ability of biochar was inhibited. The inhibitory effect of Na+ was lower than that of Ca2+ at the same concentration. The results can provide a reference for the removal of neonicotinoid insecticides in water.
-
Key words:
- biochar /
- acetamiprid /
- clothianidin /
- adsorption
-
中国酒业协会的数据显示,2019年我国啤酒产量达3 765×104 t,每t啤酒需排放的废水量约为3 t[1],其COD约为780~3 610 mg·L−12]。目前常用的厌氧-好氧法处理啤酒废水效果较好,但会产生大量剩余污泥。光合细菌(PSB)可以高效降解啤酒废水中的污染物,同时利用其中C、N、P合成菌体;PSB菌体中富含蛋白质、多糖、类胡萝卜素、叶绿素、辅酶Q10等高价值物质,可广泛用于畜牧、农业、渔业等领域,也可作为食品与药物的原材料,因此,PSB技术是一项非常有潜力的新型污水资源化技术[3-4]。
PSB几乎不产生胞外聚合物(EPS),但沉降性能差,不易实现菌体回收。膜生物反应器(MBR)结合了传统的生物处理单元与膜分离单元[6],通过膜的高效截留作用使微生物被完全截留在反应器内,因而既有利于废水的一步达标[7],也有利于微生物的高效生长、繁殖和富集[8]。国内外学者将PSB、光生物反应器与膜组件结合起来,开发了光合细菌-膜生物反应器(PSB-MBR),在提高污水处理效果的同时可实现菌体的富集与回收[9]。该方法具有操作简便、处理效果好、生物资源回收率高[10]等优点,其菌体回收率可高达99.5%[11]。
膜污染是MBR应用中的关键问题[12],会对反应器的运行性能与效果产生负面影响。QIN等[13]、彭猛[14]研究了PSB-MBR处理啤酒废水的膜污染,发现膜污染较低,推测其原因是该系统的胞外聚合物(EPS)浓度较低。然而,现有PSB-MBR研究均采用较低的运行通量,如HÜLSEN等[15]设定的PSB-MBR恒定通量为2.1 L·(m2·h)−1,其它研究也在类似水平。然而,实际污水处理厂的MBR通量为20~30 L·(m2·h)−1,比现有PSB-MBR系统通量高10倍。如此大的差异,使得现有研究无法反映未来在工业运行中可能的PSB-MBR膜污染。要实现该技术的工业化应用,将其膜通量调整为工业运行通量,会更具有参考价值。
本研究模拟工业通量,设计了PSB-MBR处理啤酒废水的一系列实验,考察了在不同的运行通量、进水COD、温度和PSB浓度下的膜污染变化情况,以期为PSB-MBR的工业化应用提供参考。
1. 材料与方法
1.1 供试原料与反应器
1) PSB菌种。菌种为通用商业菌种,其中红假单胞菌(Rhodopseudomonas palustris)比例超过80%。
2)供试废水。前期实验结果表明,PSB-MBR工艺处理模拟啤酒废水与实际啤酒废水效果一致,为方便操作,本研究采用人工配制啤酒废水。分别对啤酒稀释10、20、30倍,获得模拟废水COD约为7 400、3 700、2 500 mg·L−1,对应高、中、低浓度废水;加入硫酸铵以补充氮源,控制C/N比约为20。
3) PSB-MBR反应器。反应器结构如图1所示,为长方体玻璃反应器,长
× × × × μ 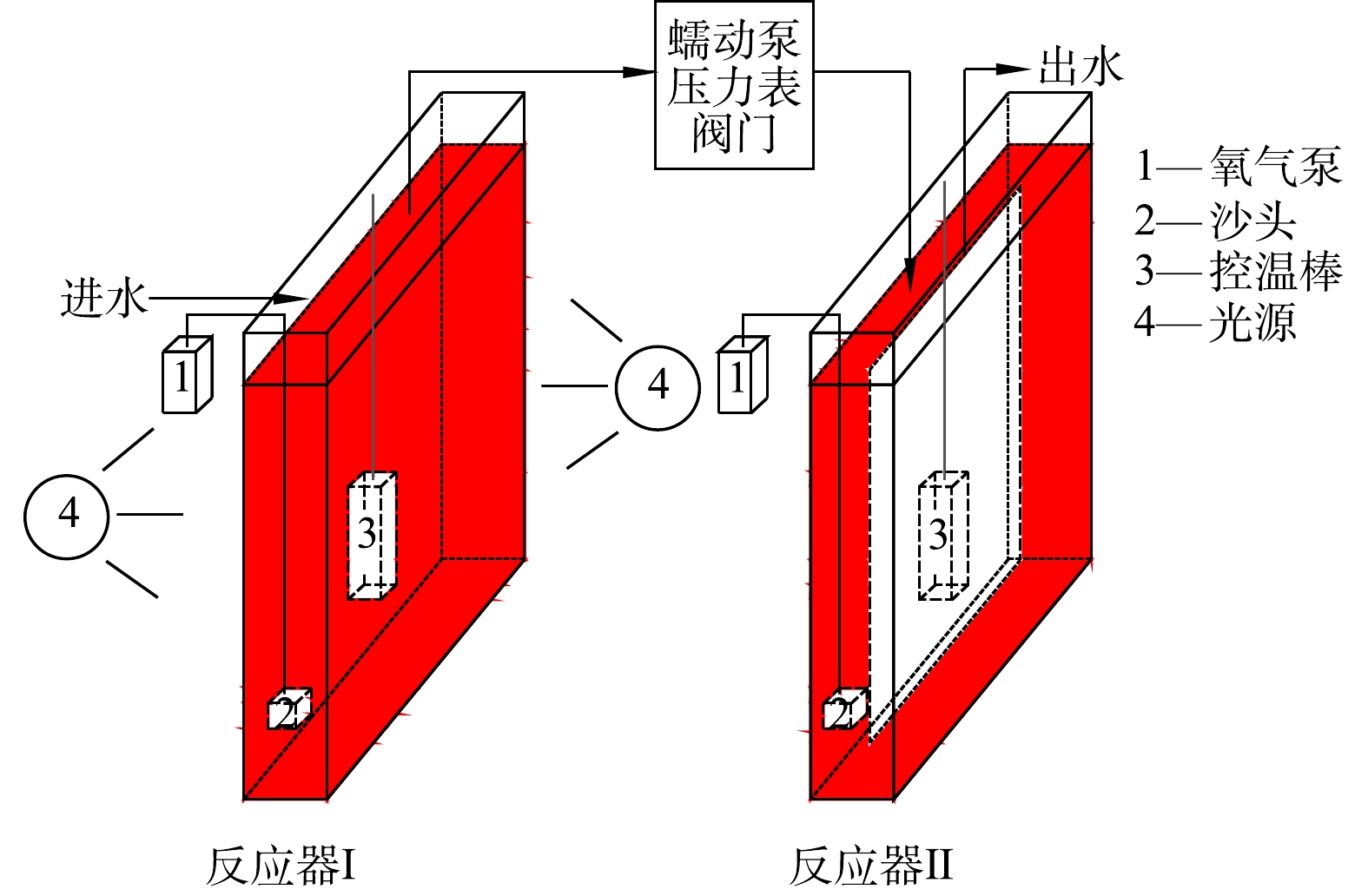 图 1 PSB-MBR反应器示意图Figure 1. Schematic of photosynthetic bacteria-membrane bioreactor (PSB-MBR)
图 1 PSB-MBR反应器示意图Figure 1. Schematic of photosynthetic bacteria-membrane bioreactor (PSB-MBR)1.2 实验方法及运行参数
生物反应器Ⅰ。PSB接种量为200~300 mg·L−1,废水利用氢氧化钠和盐酸溶液调节pH为7.2~7.6。通过氧气泵和DO仪控制溶解氧(DO)为0.2~0.5 mg·L−1,利用控温棒控制温度为约25 ℃,水力停留时间3 d。3种啤酒废水经过PSB处理后,COD分别为171、425、1 005 mg·L−1(均值),COD去除率为80%~93%。
膜分离反应器Ⅱ。该反应器运行参数取值尽量模拟可能的工业运行参数,因此取值范围较窄。生物处理后的废水输送到膜分离反应器Ⅱ中,分别考察不同进水通量、进水COD、运行温度和PSB浓度对运行过程中膜污染的影响。如无特殊说明,反应器Ⅱ的进水COD为170 mg·L−1、PSB浓度为1 000 mg·L−1、温度为25 ℃、运行通量为17.4 L·(m2·h)−1。
运行通量。设置2组反应器的运行通量为17.4、23.4 L·(m2·h)−1,这个范围是常见的污水处理膜通量,比通常的PSB-MBR实验研究高10倍[13-16]。
进水COD。根据生物反应器处理低、中、高废水的出水水质,膜分离反应器进水COD分别设置为170、425、1 000 mg·L−1。
运行温度。利用控温棒控制设定温度分别为常温(25 ℃)、低温(10 ℃),以考察低温下的膜污染。
PSB浓度。前期实验表明,PSB浓度低于1 000 mg·L−1时,污染物去除率低;高于1 500 mg·L−1时,PSB的增值率低;在1 000~1 500 mg·L−1范围内可以同时满足污染物去除与菌体合成以便后续回收利用。因此,控制进入反应器Ⅱ的PSB浓度分别为1 000、1 300、1 500 mg·L−1。
1.3 分析方法
1)水质分析。从反应器中取10 mL的菌-水混合物,在9 000 r·min−1下离心10 min后获得上清液用于水质检测。利用重铬酸钾法快速检测COD[17]。根据国标HJ 535-2009使用TU-1900分光光度计在420 nm的吸光度下分析氨氮[18]。PSB的生物量检测方法参考LU等[19]的方法。
2)膜污染阻力分析。根据达西定律(式1),测量平板膜在过滤过程中随时间变化的膜污染阻力,评估PSB-MBR系统中膜污染程度。
Rt=Rm+Rf=ΔPμ⋅J (1) 式中:Rt为膜的总过滤阻力,m−1;Rm为膜的固有阻力,m−1;Rf为膜丝污染阻力,m−1;ΔP为膜两侧的压力差,Pa;μ为透过液动力学粘度,Pa∙s;J为膜通量,m3·(m2·s)−1。
3)膜污染表征。用扫描电子显微镜(SEM,Hitachi S-4800型)对平板膜表面的微观结构进行观察,对膜表面的污染情况进行表征。将膜片浸泡在超纯水中24 h后,经0.45
μm 4)膜清洗。膜清洗有2种方式,其中物理清洗是用清水洗涤膜表面5 min;化学清洗是将膜片浸入0.75%NaClO溶液中浸泡2 h后,用清水冲洗膜表面5 min [14]。
2. 结果与讨论
2.1 操作参数对膜污染的影响
在传统MBR的运行过程中,高运行通量、高COD均会加剧污染物在膜表面的富集,从而加速膜污染的产生;一定的温度条件可能会引起微生物状态的变化,导致EPS浓度升高,进而加剧膜污染。因此,本研究分别考察了不同的运行通量、膜反应器Ⅱ进水COD与PSB浓度、运行温度等对膜的影响。当膜通量下降至初始值的80%时即判定膜受到污染。图2显示了不同操作参数对膜污染阻力的影响。
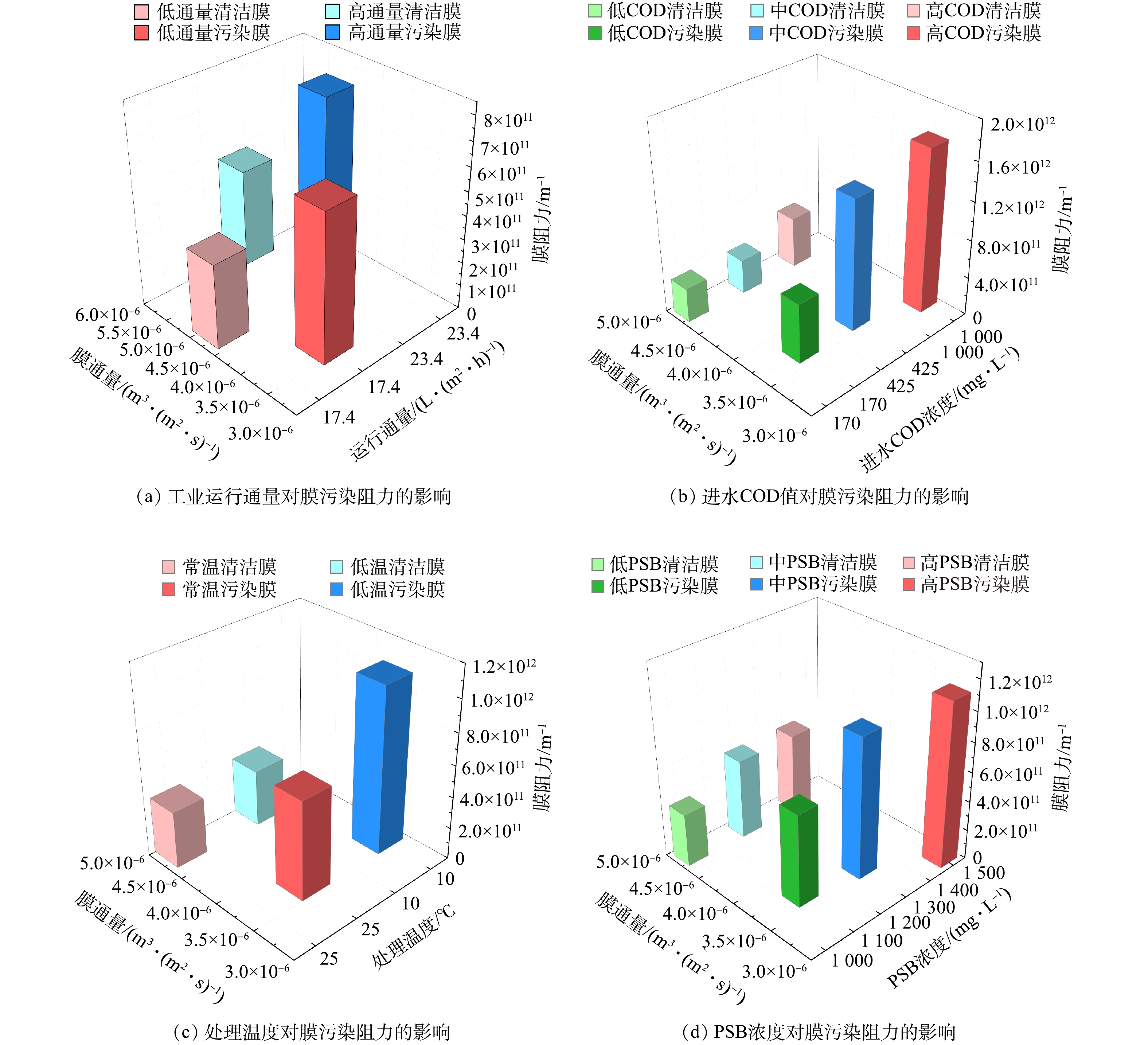 图 2 不同操作参数对膜污染阻力的影响Figure 2. The influence of membrane fouling resistance by different operating parameters
图 2 不同操作参数对膜污染阻力的影响Figure 2. The influence of membrane fouling resistance by different operating parameters在膜设计通量范围内,膜通量随着运行通量的增加而增加(图2(a)),而膜阻力也明显随运行通量增加。以23.4 L·(m2·h)−1通量运行时,污染后膜阻力为7.44
× 膜分离反应器的进水浓度高,也导致了膜阻力增加,通量下降(图2(b))。当膜的进水COD为1 000 mg∙L−1时,膜通量达到3.42
× 运行温度对膜污染阻力影响较大(图2(c))。低温条件下的污染膜污染阻力为1.06
× 膜污染随着PSB浓度的上升逐渐加剧(图2(d)),当PSB浓度为1 500 mg·L−1时,污染后的膜通量为3.17
× 根据膜阻力变化情况,本研究中进水COD是影响膜污染最重要的因素,其次为PSB浓度、温度、运行通量(1.72×1012 m−1 > 1.12×1012 m−1 > 1.06×1012 m−1 > 7.44×1011 m−1)。而在对An MBR运行影响因素的相关研究中,温度是影响其运行特性最主要的因素,其次是污泥(或COD负荷)[21],这主要是由于温度会导致厌氧微生物通过分泌EPS等物质影响污泥的特性从而使得污泥絮体增大、膜污染加剧[25]。PSB几乎不产生EPS,主要可能是由废水中的有机负荷及菌体代谢造成膜污染。因此,进水COD成为了PSB-MBR模拟工业通量运行的主要影响因素,这也与彭猛[14]使用低通量研究时获得的结果一致。
2.2 PSB-MBR中膜污染分析
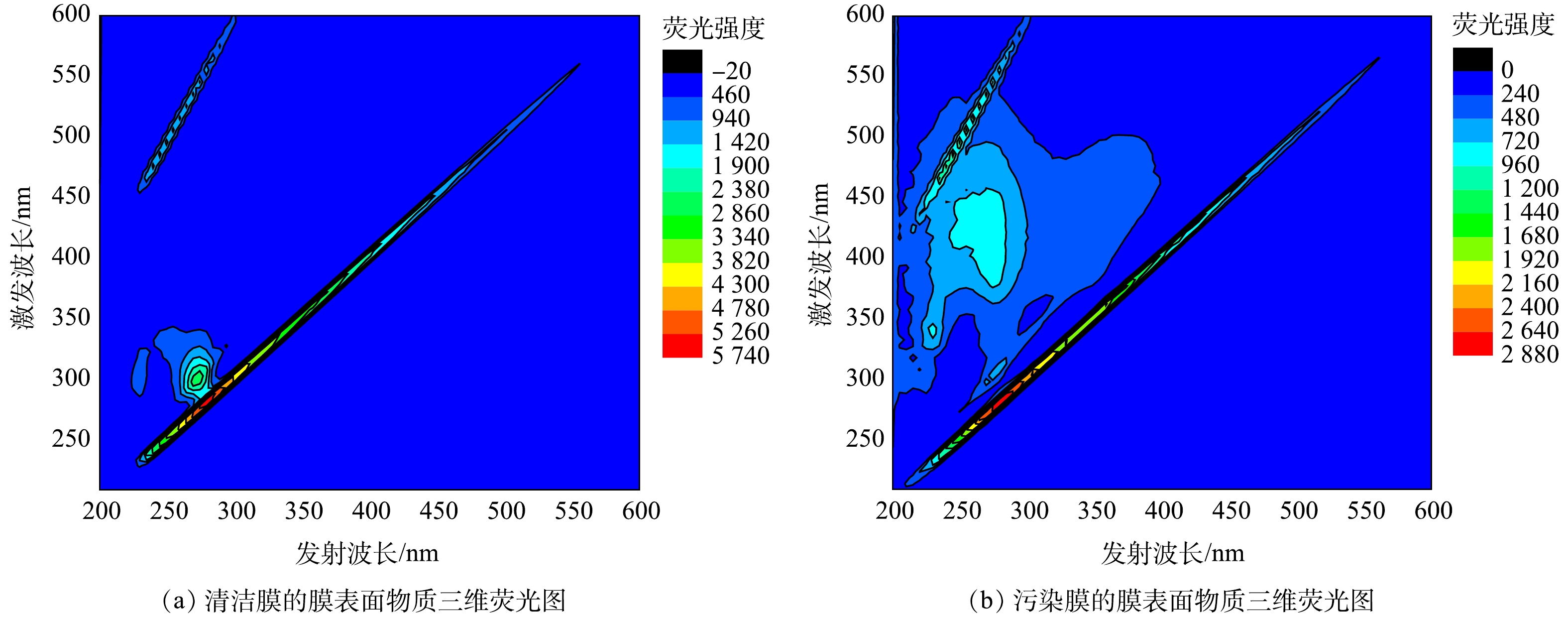
设定多次进行物理清洗后,膜通量无法恢复至初始通量的80%时为严重污染。根据2.1节中的结果,设置参数为,膜分离反应器Ⅱ进水COD 170 mg·L−1、PSB 1 000 mg·L−1、常温(25 ℃)、运行通量17.4 L·(m2·h)−1,在该条件下进行膜污染分析。利用SEM对运行后的PSB-MBR系统中膜的表面特性进行了物理分析。图3(a)显示,清洁膜片在放大倍数为5.00 k时表面平滑,无污染物质存在;在放大30.0 k时呈现明显多孔隙结构,可以判断照片中显示的孔隙即为膜片的过滤孔隙。图3(b)显示,严重污染膜在放大倍数为5.00 k的条件下,表面覆满污染物质;而图3(d)则显示污染物中有一些椭球形物体堆积,无法显示原有孔隙结构,这说明膜孔已被堵塞,大分子污染物堆积在膜表面。因此,在模拟工业通量运行时,PSB-MBR会产生较严重的膜污染,大量污染物会覆盖膜的原有结构,导致反应器的后续运行性能下降。而现有的PSB-MBR研究采用比工业通量低10倍的通量,并未发现这么明显的膜污染[15]。
通过三维荧光光谱法对膜表面物质进行了分析,结果如图4所示。图4(a)显示,清洁膜表面主要为亲水性大分子蛋白类有机物,而图4(b)中则显示膜污染物质中含有亲水性大分子蛋白类有机物与有腐殖质。由于进水为模拟啤酒废水,不含大分子蛋白质与腐殖质[27],结合SEM分析结果(图3),可以推测,造成膜污染的主要是有机污染,来自PSB及其降解或分泌物质。
膜污染通常分为3个阶段,分别为初始污染阶段、缓慢污染阶段及跨膜压差(TMP)跃升阶段[28]。在初始污染阶段,膜表面与混合液发生相互作用,粒径小于膜孔径的污染物颗粒进入膜孔,其中一些被吸附于膜孔内,减小了膜孔的有效直径。因此,膜通量由4.83
× × μ μ × × 2.3 膜清洗
为使已经产生污染的膜恢复膜通量,需要对其进行清洗。膜清洗方法主要分为物理清洗和化学清洗。在反应器运行过程中,对平板膜进行周期性物理清洗(清水冲洗5 min)。由图5所示,初始阶段物理清洗效果很好,膜通量几乎完全恢复。第4次清洗前,膜通量下降到起始膜通量的62%,物理清洗后膜通量100%恢复;第11次物理清洗仍然可以使膜通量恢复至起始膜通量的97%。郭雅妮等的[30]研究表明,传统MBR中物理清洗后膜通量仅恢复至新膜的70%。因此,PSB-MBR系统中物理清洗效果远比传统MBR好,这有利于延长膜片的使用寿命。其原因是,由于PSB几乎不产生EPS,因此,传统MBR中污染最严重也较难清洗的EPS污染在PSB-MBR中几乎没有贡献,从而使得在PSB-MBR系统中物理清洗效果显著。然而,随着物理清洗次数的增加,膜通量恢复效果仍然在缓慢下降,这是因为有机污染逐渐积累,污染层结构逐渐致密化,导致物理清洗效果开始变差。在彭猛[14]的研究中,物理清洗40次,膜通量仍然可以100%恢复。这是因为,其采用的是低通量,其通量仅为本文研究的8%~15%。从图2(a)可知,通量越低,膜阻力越小,越容易恢复。这一结果也表明,在工业通量下,PSB-MBR系统的膜污染虽然比常规MBR低,但是比现有研究低通量PSB-MBR高。
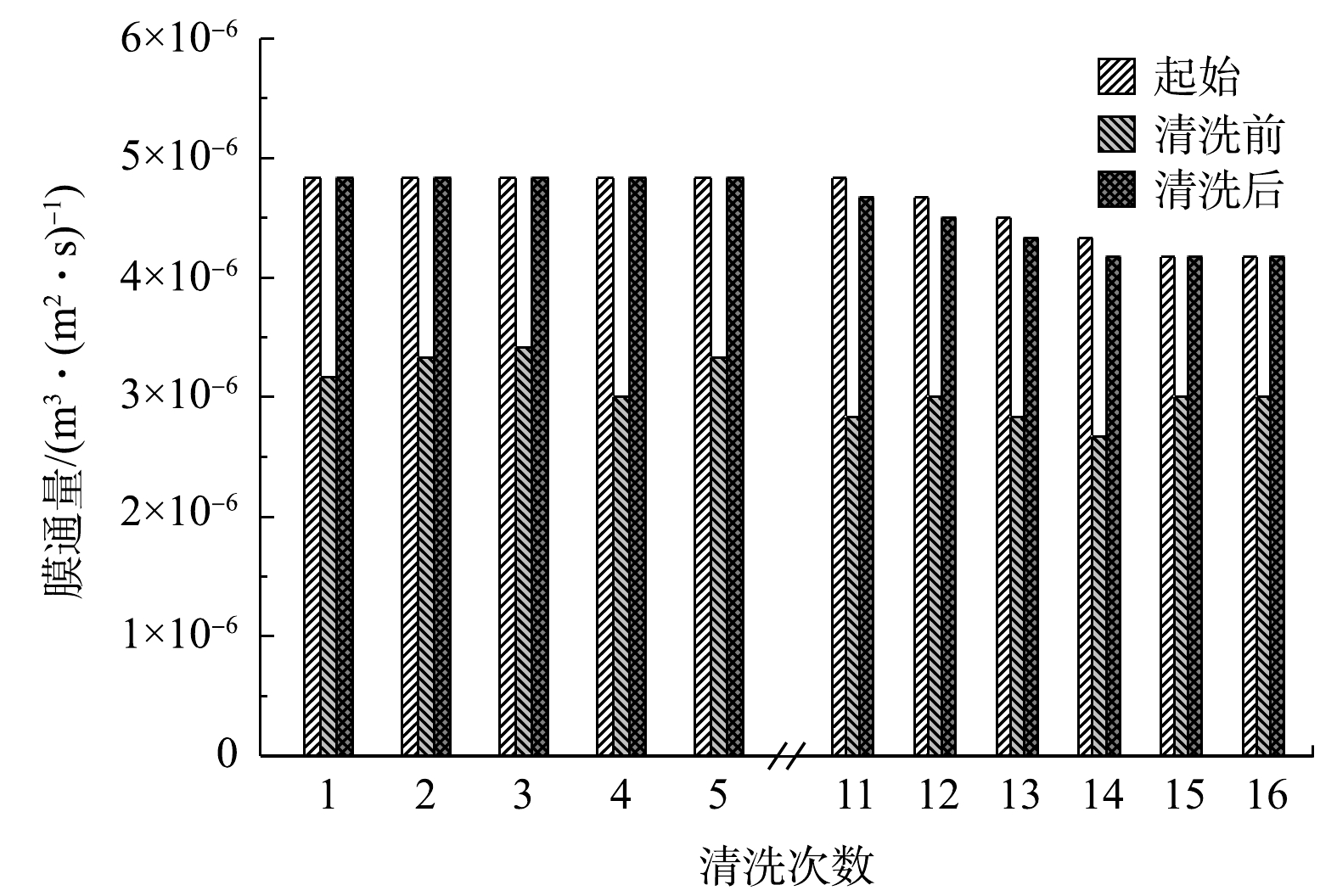 图 5 每次进行物理清洗的膜通量变化Figure 5. Changes in membrane flux after repeated physical cleaning
图 5 每次进行物理清洗的膜通量变化Figure 5. Changes in membrane flux after repeated physical cleaning当物理清洗无法使膜通量恢复至80%时,需要采用化学清洗以恢复膜通量。AHMAD等[31]研究了不同化学清洁剂对化学清洗后膜通量的恢复效果,发现用0.75% NaClO溶液的清洗效果最好,可以恢复约98%膜通量并且使膜孔基本恢复。康永和胡肖勇[32]发现,采用2.0%~5.0%的NaClO溶液清洗膜时,既可以去除污垢,又可以去除膜孔内附着的微生物和蛋白质等有机污染物。谢元华等[33]利用0.1%的NaClO溶液浸泡机械清洗后的膜2 h后,过膜阻力几乎完全恢复。根据以上研究,本研究使用NaClO溶液作为化学清洗药品,具体化学清洗方法为0.75% NaClO溶液浸泡2 h。清洗后膜通量100%恢复,4次清洗后仍然可达到这一效果。
此外,本研究采用三维荧光光谱法分析了化学清洗后的膜,结果见图6。化学清洗后的膜表面,主要污染物质是亲水性小分子蛋白类有机物,而由图4(b)可知,污染膜表面主要污染物是腐殖质和大分子蛋白类有机物,表明化学清洗将膜表面的大分子污染物转化成为小分子物质。其原因是NaClO的氧化性及其对微生物细胞的破坏作用。这一现象与常规MBR膜化学清洗相似[34],NaClO碱洗后膜通量几乎完全恢复,这说明有机污染是造成膜污染的主要原因。
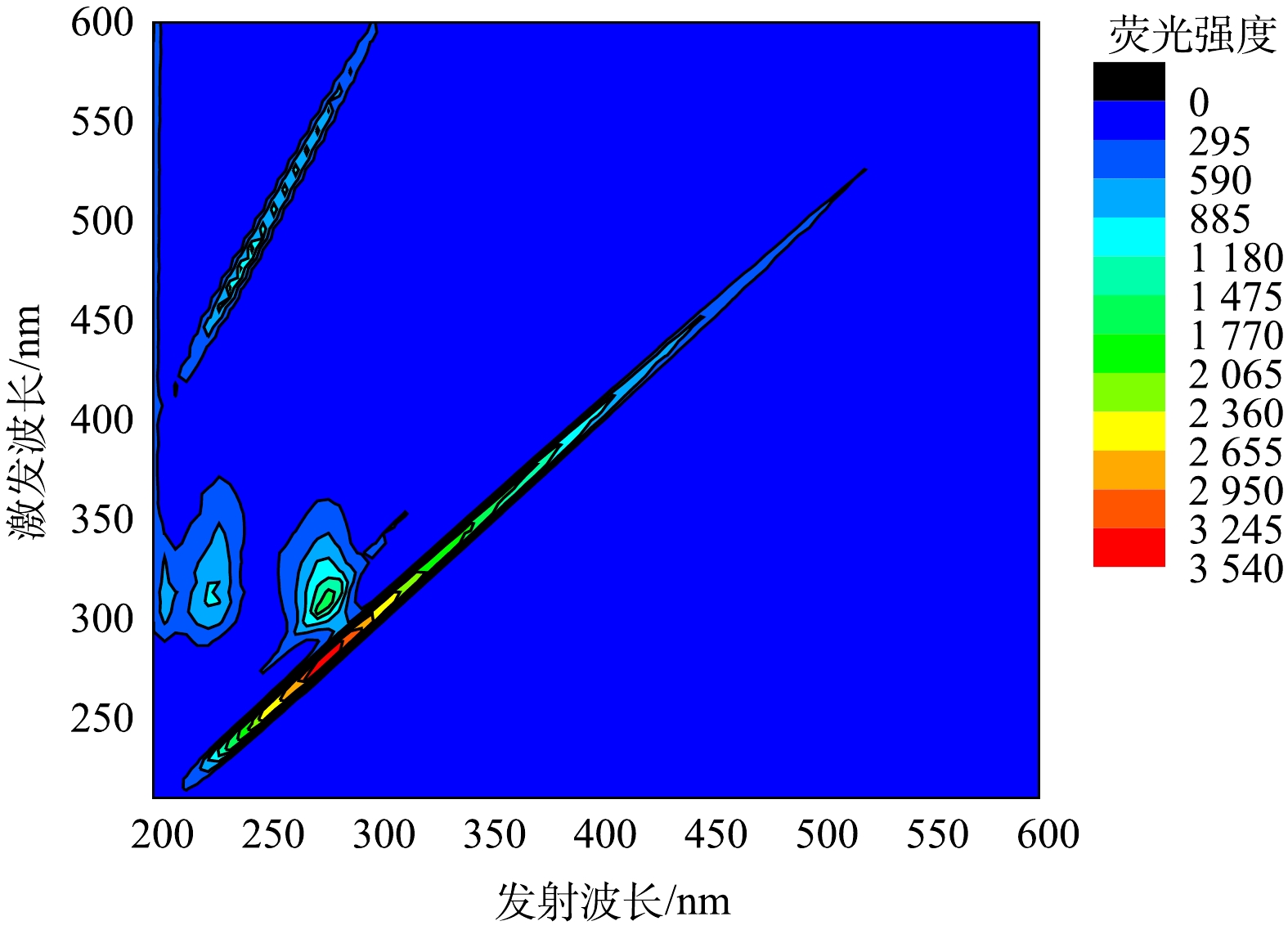 图 6 化学清洗(0.75% NaClO溶液浸泡2 h)后的膜的三维荧光图Figure 6. EEM image of membrane after chemical cleaning (0.75% NaClO, 2 h)
图 6 化学清洗(0.75% NaClO溶液浸泡2 h)后的膜的三维荧光图Figure 6. EEM image of membrane after chemical cleaning (0.75% NaClO, 2 h)3. 结论
1) PSB-MBR模拟工业级别运行通量(17.4 L·(m2·h)−1)时,PSB-MBR膜污染比现有研究中低通量下PSB-MBR膜污染高,但远低于常规MBR污染,其原因是PSB几乎不产生EPS。
2)高运行通量、高PSB浓度、高COD、低温会提高膜阻力,加剧膜污染。
3)造成PSB-MBR系统膜污染的主要来源是PSB菌体本身及其代谢产物。
4)物理清洗无法使膜通量恢复至80%时,化学清洗可以恢复膜通量,通过将原污染物中的腐殖质转变为亲水性小分子蛋白类有机物,解决膜孔堵塞问题。
-

图 2 BC1和BC2的N2吸附-脱附等温线和BJH孔径分布
Figure 2. Nitrogen adsorption and desorption isotherms and BJH pore size distribution of BC1 and BC2
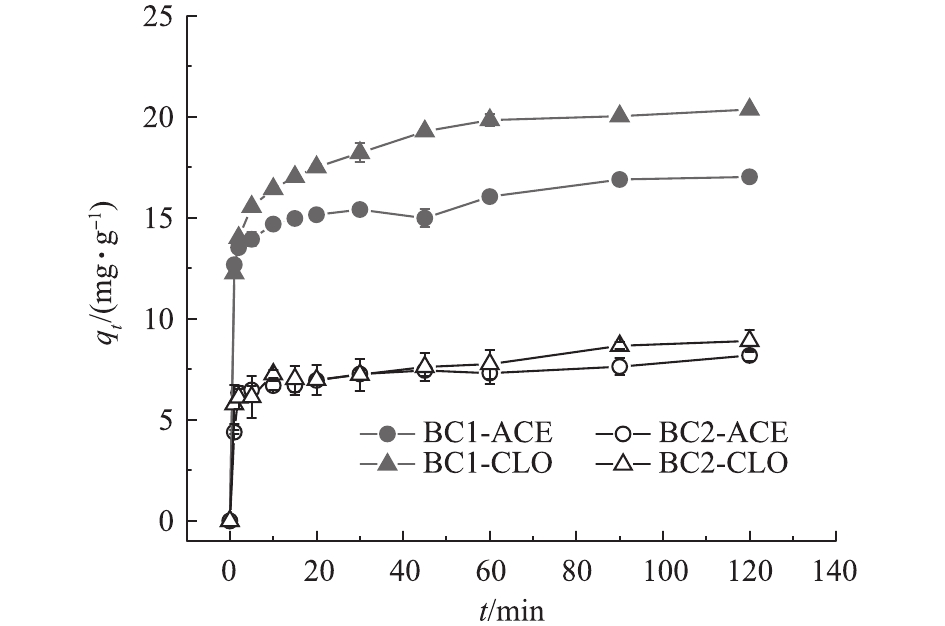
图 4 BC1和BC2对ACE、CLO的吸附动力学曲线
Figure 4. Adsorption kinetics of ACE and CLO on BC1 and BC2

图 5 BC1和BC2吸附ACE、CLO的颗粒内扩散方程拟合
Figure 5. Intraparticle diffusion model fitting of ACE and CLO adsorption on BC1 and BC2

图 6 BC1和BC2对ACE、CLO的吸附等温线
Figure 6. Adsorption isotherms of ACE and CLO on BC1 and BC2
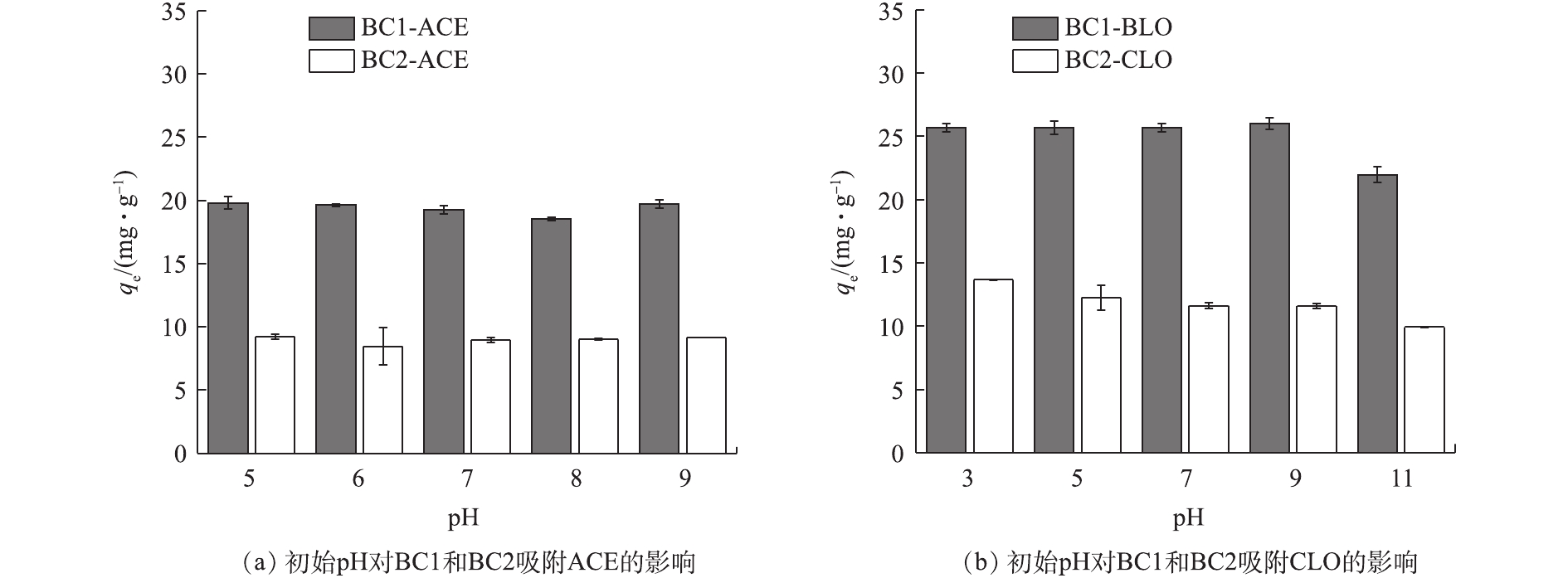
图 7 溶液初始pH对BC1和BC2吸附ACE、CLO的影响
Figure 7. Effect of initial pH on the adsorption of ACE and CLO on BC1 and BC2
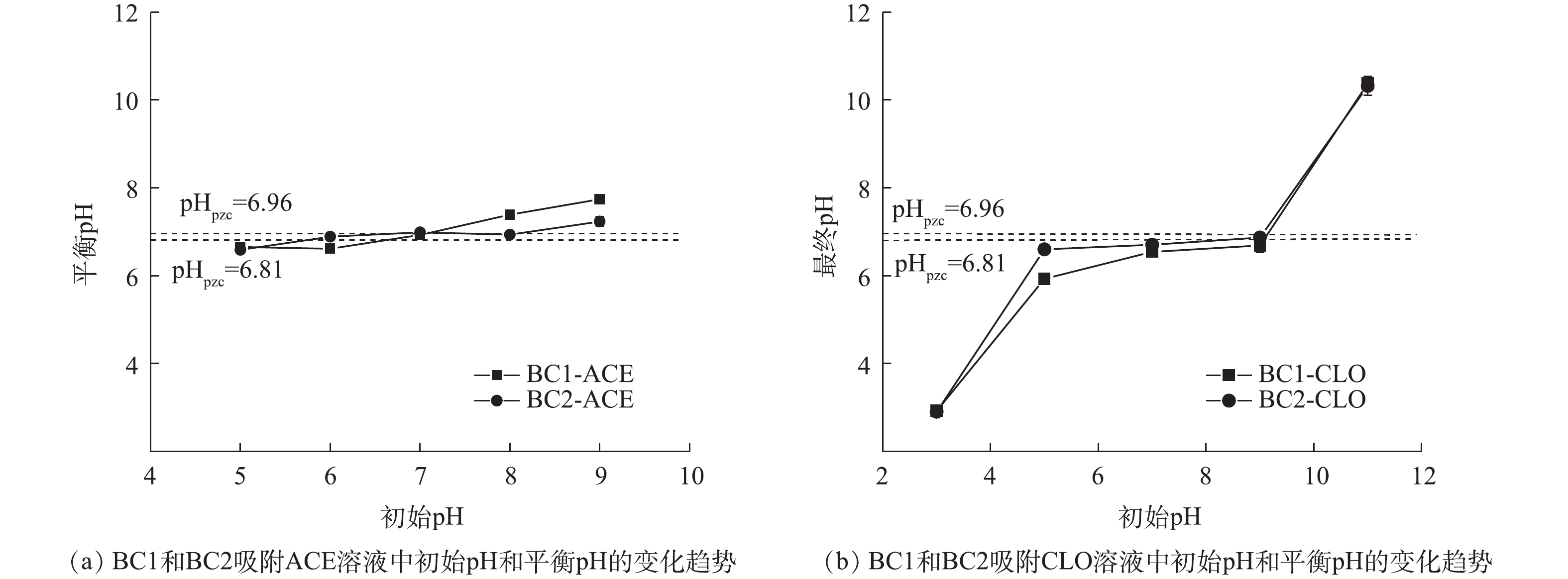
图 8 溶液初始pH与平衡pH变化趋势
Figure 8. Change trend of initial pH and equilibrium pH in solution

图 9 离子强度对BC1和BC2吸附ACE、CLO的影响
Figure 9. Effects of ionic strength on the adsorption of ACE and CLO on BC1 and BC2
表 2 BC1和BC2的BET比表面积和孔隙结构参数
Table 2. BET specific surface areas and pore structure parameters of BC1 and BC2
吸附剂 比表面积/(m2·g−1) 平均孔径/nm 总孔容/(cm3·g−1) BC1 440.27 2.06 0.227 BC2 329.15 1.94 0.160  下载: 导出CSV
下载: 导出CSV
表 3 BC1和BC2吸附ACE、CLO动力学参数
Table 3. Kinetic parameters of ACE and CLO adsorption on BC1 and BC2
吸附剂 杀虫剂 准一级动力学方程 准二级动力学方程 qe/(mg·g−1) k1/h−1 R2 qe/(mg·g −1) k2/(g·(mg·min)−1) R2 BC1 ACE 14.07 61.08 0.988 14.57 5.92 0.997 CLO 16.42 65.85 0.953 17.43 6.29 0.984 BC2 ACE 6.86 59.97 0.992 7.26 14.06 0.996 CLO 6.82 103.22 0.971 7.09 31.30 0.985
下载: 导出CSV
表 4 BC1和BC2吸附ACE、CLO的颗粒内扩散方程拟合参数
Table 4. Fitting parameters of intraparticle diffusion model of ACE and CLO adsorption on BC1 and BC2
吸附剂 杀虫剂 阶段1 阶段2 阶段3 k1 C1 R2 k2 C2 R2 k3 C3 R2 BC1 ACE 10.140 10.644 0.913 3.433 12.521 0.998 0.382 16.205 0.995 CLO 16.858 9.920 0.976 6.423 13.570 0.999 0.227 18.986 0.993 BC2 ACE 6.667 3.605 0.956 0.941 6.618 0.994 0.397 7.745 0.899 CLO 3.242 5.337 0.984 2.483 5.456 0.997 0.149 9.375 0.879
下载: 导出CSV
表 5 BC1和BC2吸附ACE、CLO的吸附等温线拟合参数
Table 5. Fitting parameters of adsorption isotherms of ACE and CLO on BC1 and BC2
吸附剂 杀虫剂 温度/℃ Langmuir模型 Freundlich模型 qm/(mg·g−1) KL/(L·mg−1) R2 1/n KF R2 BC1 ACE 15 19.99 3.61 0.951 0.14 14.31 0.891 25 24.47 3.87 0.979 0.15 17.34 0.866 35 24.95 7.31 0.897 0.14 18.45 0.869 BC2 ACE 15 11.82 2.28 0.953 0.12 8.58 0.890 25 11.13 1.85 0.895 0.14 7.62 0.962 35 13.35 1.62 0.968 0.16 8.62 0.942 BC1 CLO 15 23.96 2.70 0.939 0.14 18.49 0.869 25 31.56 1.20 0.902 0.16 16.51 0.835 35 30.00 4.88 0.949 0.18 20.10 0.900 BC2 CLO 15 13.05 1.31 0.954 0.18 7.95 0.866 25 12.25 3.60 0.930 0.11 9.29 0.881 35 14.11 3.28 0.835 0.12 10.53 0.910
下载: 导出CSV
表 6 BC1和BC2吸附ACE、CLO的热力学参数
Table 6. Thermodynamic parameters for ACE and CLO adsorption onto BC1 and BC2
吸附剂 杀虫剂 温度/℃ lnKd ΔG0/(kJ·mol−1) ΔH0/(kJ·mol−1) ΔS0/(kJ·(mol·K)−1) BC1 ACE 15 15.50 −21.02 38.14 0.262 1 25 16.33 −21.75 35 16.53 −22.48 BC2 ACE 15 14.44 −19.44 12.73 0.164 0 25 14.57 −20.12 35 14.79 −20.79 BC1 CLO 15 17.01 −11.52 16.63 0.180 0 25 16.73 −11.92 35 17.34 −12.32 BC2 CLO 15 14.86 −18.38 13.23 0.168 9 25 14.82 −19.01 35 15.22 −19.65
下载: 导出CSV
-
[1] JESCHKE P, NAUEN R, SCHINDLER M, et al. Overview of the status and global strategy for neonicotinoids[J]. Journal of Agricultural and Food Chemistry, 2011, 59(7): 2897-2908. doi: 10.1021/jf101303g [2] MITCHELL E A D, MULHAUSER B, MULOT M, et al. A worldwide survey of neonicotinoids in honey[J]. Science, 2017, 358(6359): 109-111. doi: 10.1126/science.aan3684 [3] HALLMANN C A, FOPPEN R P B, VAN TURNHOUT C A M, et al. Declines in insectivorous birds are associated with high neonicotinoid concentrations[J]. Nature, 2014, 511(7509): 341-343. doi: 10.1038/nature13531 [4] CAVALLARO M C, MAIN A R, LIBER K, et al. Neonicotinoids and other agricultural stressors collectively modify aquatic insect communities[J]. Chemosphere, 2019, 226: 945-955. doi: 10.1016/j.chemosphere.2019.03.176 [5] HAN W C, TIAN Y, SHEN X M. Human exposure to neonicotinoid insecticides and the evaluation of their potential toxicity: An overview[J]. Chemosphere, 2018, 192: 59-65. doi: 10.1016/j.chemosphere.2017.10.149 [6] 刘晓, 田颖, 周晓慧, 等. 零价铝在碱性条件下还原吡虫啉[J]. 化工学报, 2017, 68(4): 384-389. [7] MULLIGAN R A, TOMCO P L, HOWARD M W, et al. Aerobic versus anaerobic microbial degradation of clothianidin under simulated california rice field conditions[J]. Journal of Agricultural & Food Chemistry, 2016, 64(38): 7059-7067. [8] MIR N A, KHAN A, MUNEER M, et al. Photocatalytic degradation of a widely used insecticide thiamethoxam in aqueous suspension of TiO2: Adsorption, kinetics, product analysis and toxicity assessment[J]. Science of the Total Environment, 2013, 458-460: 388-398. doi: 10.1016/j.scitotenv.2013.04.041 [9] LI H, DONG X, DA SILVA E B, et al. Mechanisms of metal sorption by biochars: Biochar characteristics and modifications[J]. Chemosphere, 2017, 178: 466-478. doi: 10.1016/j.chemosphere.2017.03.072 [10] 程扬, 沈启斌, 刘子丹, 等. 两种生物炭的制备及其对水溶液中四环素去除的影响因素[J]. 环境科学, 2019, 40(3): 1328-1336. [11] NIE T, HAO P, ZHAO Z, et al. Effect of oxidation-induced aging on the adsorption and co-adsorption of tetracycline and Cu2+ onto biochar[J]. Science of the Total Environment, 2019, 673: 522-532. doi: 10.1016/j.scitotenv.2019.04.089 [12] YIN Y, GUO X, PENG D. Iron and manganese oxides modified maize straw to remove tylosin from aqueous solutions[J]. Chemosphere, 2018, 205: 156-165. doi: 10.1016/j.chemosphere.2018.04.108 [13] 李蕊宁, 王兆炜, 郭家磊, 等. 酸碱改性生物炭对水中磺胺噻唑的吸附性能研究[J]. 环境科学学报, 2017, 37(11): 4119-4128. [14] 李政剑, 石宝友, 苏宇, 等. 粉末活性炭粒径对水中菲吸附动力学的影响效应研究[J]. 环境科学学报, 2013, 33(1): 70-75. [15] PRESTON C M, SCHMIDT M W I. Black (pyrogenic) carbon: A synthesis of current knowledge and uncertainties with special consideration of boreal regions[J]. Biogeosciences, 2006, 3: 397-420. doi: 10.5194/bg-3-397-2006 [16] 邵翼飞, 张鹏, 刘爱菊. 生物炭对噻虫胺在土壤中吸附和降解的影响[J]. 农业环境科学学报, 2019, 38(11): 2520-2527. doi: 10.11654/jaes.2019-0460 [17] 庾琴, 周华, 王静, 等. 啶虫脒在环境中的降解代谢及其安全性的研究进展[J]. 农药, 2007, 46(4): 223-226. doi: 10.3969/j.issn.1006-0413.2007.04.002 [18] 管欢, 黄慧俐, 行艳景, 等. 噻虫胺在甘蔗和土壤中的残留分析及消解动态[J]. 现代农药, 2015, 14(2): 48-51. [19] 谢国红, 刘国光, 孙德智, 等. 啶虫脒水解动力学研究[J]. 安徽农业科学, 2007, 35(30): 9629-9630. doi: 10.3969/j.issn.0517-6611.2007.30.078 [20] 杨奇亮, 吴平霄. 改性多孔生物炭的制备及其对水中四环素的吸附性能研究[J]. 环境科学学报, 2019, 39(12): 3973-3984. [21] 张学良, 徐建, 占新华, 等. 微波辅助合成γ-Fe2O3/花生壳磁性生物炭对水体中环丙沙星吸附的研究[J]. 环境科学学报, 2019, 39(11): 3811-3820. [22] 郑晓青, 韦安磊, 张一璇, 等. 铁锰氧化物/生物炭复合材料对水中硝酸根的吸附特性[J]. 环境科学, 2018, 39(3): 262-274. [23] MUTTAKIN M, MITRA S, THU K, et al. Theoretical framework to evaluate minimum desorption temperature for IUPAC classified adsorption isotherms[J]. International Journal of Heat and Mass Transfer, 2018, 122: 795-805. doi: 10.1016/j.ijheatmasstransfer.2018.01.107 [24] OUYANG D, CHEN Y, YAN J, et al. Activation mechanism of peroxymonosulfate by biochar for catalytic degradation of 1,4-dioxane: Important role of biochar defect structures[J]. Chemical Engineering Journal, 2019, 370: 614-624. doi: 10.1016/j.cej.2019.03.235 [25] TAN X L, FANG M, CHEN C L, et al. Counterion effects of nickel and sodium dodecylbenzene sulfonate adsorption to multiwalled carbon nanotubes in aqueous solution[J]. Carbon, 2008, 46(13): 1741-1750. doi: 10.1016/j.carbon.2008.07.023 [26] LIU S, XU W H, LIU Y G, et al. Facile synthesis of Cu(II) impregnated biochar with enhanced adsorption activity for the removal of doxycycline hydrochloride from water[J]. Science of the Total Environment, 2017, 592: 546-553. doi: 10.1016/j.scitotenv.2017.03.087 [27] 孙航, 蒋煜峰, 石磊平, 等. 不同热解及来源生物炭对西北黄土吸附敌草隆的影响[J]. 环境科学, 2016, 37(12): 4857-4866. [28] ZHOU Y Y, LIU X C, XIANG Y J, et al. Modification of biochar derived from sawdust and its application in removal of tetracycline and copper from aqueous solution: Adsorption mechanism and modelling[J]. Bioresource Technology, 2017, 245: 266-273. doi: 10.1016/j.biortech.2017.08.178 [29] 孙绪兵, 吴雪梅, 朱建发, 等. 羧基甲壳素对Pb(Ⅱ)的吸附性能及机理研究[J]. 中国环境科学, 2018, 38(8): 3018-3028. doi: 10.3969/j.issn.1000-6923.2018.08.030 [30] 张连科, 王洋, 王维大, 等. 生物炭负载纳米羟基磷灰石复合材料的制备及对铅离子的吸附特性[J]. 化工进展, 2018, 37(9): 3492-3501. [31] DAI L C, ZHU W K, HE L, et al. Calcium-rich biochar from crab shell: An unexpected super adsorbent for dye removal[J]. Bioresource Technology, 2018, 267: 510-516. doi: 10.1016/j.biortech.2018.07.090 [32] PENG X, HU F, LAM F L, et al. Adsorption behavior and mechanisms of ciprofloxacin from aqueous solution by ordered mesoporous carbon and bamboo-based carbon[J]. Journal of Colloid Interface Science, 2015, 460: 349-360. doi: 10.1016/j.jcis.2015.08.050 [33] LIU P, LIU W J, JIANG H, et al. Modification of biochar derived from fast pyrolysis of biomass and its application in removal of tetracycline from aqueous solution[J]. Bioresource Technology, 2012, 121: 235-240. doi: 10.1016/j.biortech.2012.06.085 [34] LI H Q, HU J T, MENG Y, et al. An investigation into the rapid removal of tetracycline using multilayered graphene-phase biochar derived from waste chicken feather[J]. Science of the Total Environment, 2017, 603-604: 39-48. doi: 10.1016/j.scitotenv.2017.06.006 [35] PEIRIS C, GUNATILAKE S R, MLSNA T E, et al. Biochar based removal of antibiotic sulfonamides and tetracyclines in aquatic environments: A critical review[J]. Bioresource Technology, 2017, 246: 150-159. doi: 10.1016/j.biortech.2017.07.150 -

 点击查看大图
点击查看大图

计量
- 文章访问数: 5471
- HTML全文浏览数: 5471
- PDF下载数: 49
- 施引文献: 0
