

-
漆酶(Laccase, EC 1. 10. 3. 2)是一类含铜多酚氧化酶, 它的起源可追溯到19世纪, 从漆树的汁液中提取分离出来[1]. 漆酶广泛分布于植物、真菌和昆虫中, 如茶树、白腐真菌、灰色链霉菌等[2-4]. 根据它的来源, 植物漆酶和真菌漆酶是主要的两大漆酶类群. 漆酶具有高效的催化氧化功能, 可通过酶促反应氧化耦合环境体系中有机化合物, 它也被视为值得研发的一项生态环保污染修复技术[5]. 然而, 植物漆酶提取工艺的复杂性限制了它的实际应用, 而从真菌中成功提取纯化的漆酶已多达上百种, 并且多数真菌漆酶已经投入到工业化生产和商业化应用中[6-7] .
由于游离漆酶应用的经济成本以及操作不稳定性, 固定化技术为真菌漆酶在环境污染修复的运用开拓了道路. 已有相关研究报道了多种固定化载体的应用潜力, 如纳米氧化物、金属有机骨架材料、多糖物质等[8-10] . 作为优良的吸附剂或催化剂, 活性炭、生物炭、碳纳米管、石墨烯、富勒烯等炭基材料在环境和能源领域的应用吸引了广泛关注[11-12]. 它们均为高度芳香化的富炭材料, 却因其孔隙特征、表面基团、或电子密度差异呈现结构的异质性, 在吸附或催化有机化合物行为上承担着互有不同的作用[13-14]. 以污染物去除角度来看,炭基材料固定化真菌漆酶的复合体以降解环境体系的污染物为主, 且具有明显的去除效果. 此方向研究要求更多的工作进一步挖掘. 本文针对多种炭基材料固定化真菌漆酶的性能以及固定化复合材料催化氧化环境有机物的效果进行了系统归纳和讨论, 旨为拓展真菌漆酶在环境有机污染治理中可循环应用的视野.
-
真菌漆酶为高度糖基化的单亚基蛋白, 其分子量可达60—90 kDa, 大约包含500—550个氨基酸. 氨基酸缩合的肽链缠绕着活性铜(Cu)原子形成螯合物, 如图1所示.
真菌漆酶结构中常含有4个Cu原子与其氨基酸结构的特定位置相结合, 没有其他辅助因子[15]. 在酶促反应过程中, Cu原子构成了真菌漆酶催化氧化的关键位点, 它们起着电子转移的作用. 根据电磁学特征, 可划分为3类: I型Cu2+(T1 Cu)和 II型Cu2+(T2 Cu)均含有一个Cu原子, 为单电子受体, 呈现顺磁性. III型Cu24+(T3 Cu)含有两个铜原子, 且两个铜原子为反磁性的双电子受体, 依靠羟基进行相连. T1 Cu参与分子内的电子传递, 把电子从污染物传递到其他铜原子上. T2 Cu 和T3 Cu共同构成了一个三核铜簇中心, 该三核铜簇中心能够接收T1 Cu传递的电子, 并将其分配给水中溶解的分子氧, 还原转化为水, 其反应机制如下所示:
本质上, 漆酶的催化氧化能力是利用它与化合物的氧化还原电势差将污染物氧化[16-17] . 显然, 真菌漆酶的三核铜簇中心区域也可将电子转移至氧化还原电势低的介质(如没食子酸、2, 2-联氮-二-3-乙基苯并噻唑啉-6-磺酸 (ABTS)), 间接形成特定的自由基或中间体, 进而催化氧化水体污染物[18].
-
真菌漆酶的催化氧化性能被广泛应用于制浆造纸、纺织印染、食品加工、环境修复等多个领域[19-21]. 例如, 真菌漆酶可以协同木聚糖酶可对废弃报纸进行脱墨处理[22]. Couto 等[23]从Trametes hirsute 中提取的粗酶对于4种偶氮染料均有超过50%的脱色效果. Wang 等[24]通过真菌漆酶澄清苹果汁中所含酚类物质, 发现苹果汁的酚类化合物降低了16. 3%, 透光率提高了20. 2%, 有利避免苹果汁长期储存的变质问题. 真菌漆酶在环境污染修复领域的应用也被视为是一种高效、绿色的技术手段. 它可有效降解酚类、羧酸类、芳胺类、以及雌激素等有机污染物. 例如, Chairin 等[25]从Trametes polyzona WR710-1中提取了粗制漆酶, 在1-羟基苯并三唑 (HBT)介体存在的条件下, 发现3 h真菌漆酶可完全去除100 mg·L−1的水体中双酚A. Liu 等[26]利用真菌漆酶对雌二醇(17β-estradiol)进行降解, 发现2 h内10 μmol·L−1的17β-estradiol的去除率可高达99. 24%. Sun 等[27]通过金属离子和天然有机质介导真菌漆酶催化氧化偶联反应过程, 可完全降解水体中的三氯生, 能有效降低三氯生的毒性. 然而, 游离态真菌漆酶的应用也存在局限性. 第一, 稳定性较弱. 漆酶活性易受体系的温度、酸碱度、溶剂成分等因素影响. 通常, 真菌漆酶在适宜的温度区间(30—60 °C)内反应活性相对较高. 若超出最适温度区间, 则漆酶的活性会显著降低[28]. 第二, 重复利用率低. 游离的漆酶是水溶性物质, 通常难以从废水溶液中回收并加以循环利用, 这造成了资源的巨大浪费. 第三, 生产成本高. 现有的真菌漆酶的提纯工艺技术仍难以降低漆酶生产成本, 这是限制真菌漆酶大规模生产应用的主要障碍之一[29]. 因此, 为提高真菌漆酶的循环利用效率, 漆酶固定化技术越来越受到关注, 比如, 利用支撑材料吸附漆酶、或通过共价结合增强负载漆酶效果、或将漆酶包埋在膜或凝胶内等. 固定化漆酶是克服游离漆酶难回收的有效途径, 也是在温度、pH、贮存、以及操作等方面提高漆酶稳定性的有利方法.
-
固定化漆酶在修复技术领域有着巨大潜在应用价值, 特别是针对难处理的环境污染物. 真菌漆酶的固定化方法主要是通过物理或化学方法使水溶性酶与稳定的固态载体相结合, 为了提高酶在连续使用时的稳定性和防止酶活性的丧失[30-31]. 炭基材料如活性炭、生物炭、纳米碳管、石墨等, 因其高度芳香化的炭质结构, 被视为稳定的、机械支撑性能优异的固定化载体[32]. 炭基材料与漆酶分子的相互作用属于弱力作用, 例如范德华力、疏水性作用、离子架桥作用、氢键效应等. 它们不会改变真菌漆酶的原始构型而导致漆酶失活. 炭基材料通常具有多层级孔隙结构或丰富的表面官能团, 为真菌漆酶的固载也提供了重要的位点支撑[33]. 例如,Ramire-Motoya等[34]利用三氯化铁溶液浸渍山核桃壳 于800 °C 在CO2氛围下,裂解制备活性炭. 按1:1000固液比将活性炭分散于漆酶溶液中, 离心分离后获得固定化漆酶, 此漆酶固载率可达63%. 这是由于活性炭丰富介孔结构有利于漆酶的固定. 介孔度越高, 漆酶的负载率越高. Dong等[35]以多层石墨烯为载体制备了石墨烯-漆酶的复合材料, 这种四层石墨烯结构, 因其高比表面积和非共价π-π堆叠作用, 对于漆酶的负载可达211. 1 mg·g−1. 载体表面的酶特异性基团越多, 载体与漆酶的亲和力越高, 从而漆酶在炭基材料表面的负载越多. 松木、猪粪和杏仁壳裂解的生物炭已经被证明是固定粗漆酶的优良吸附材料[36]. 为了能够使生物炭基体高效固载真菌漆酶, 也可通过基团修饰方法引入活性基团如环氧基、羟基、羧基等, 来增加载体表面的结合位点, 促进漆酶的固定化效果(表1). Imam等[37]酸化水稻秸秆裂解的生物炭来丰富炭基表面极性官能团, 发现漆酶的固载效率高达66%. 而且, 经6次循环试验后生物炭对漆酶的固持仍可保留了40%. 而且, 固定化漆酶的储存稳定性和化学耐受能力也可能有所提升. 相较于游离漆酶, 柠檬酸改性猪粪生物炭固定的漆酶, 具有较高的酶活性和贮存能力. 酶性能的提升可能是由于漆酶分子被固相载体多孔结构所保护, 避免了漆酶蛋白的构象变化[38]. 综上, 炭基材料因其物理强度高、表面积大、孔隙率高、表面官能团丰富、热稳定性优良等优点, 为真菌漆酶固定化载体材料提供了一条选择路径. 将炭基材料用于漆酶的固定化载体是一种可探究的途径来实现漆酶的循环利用.
-
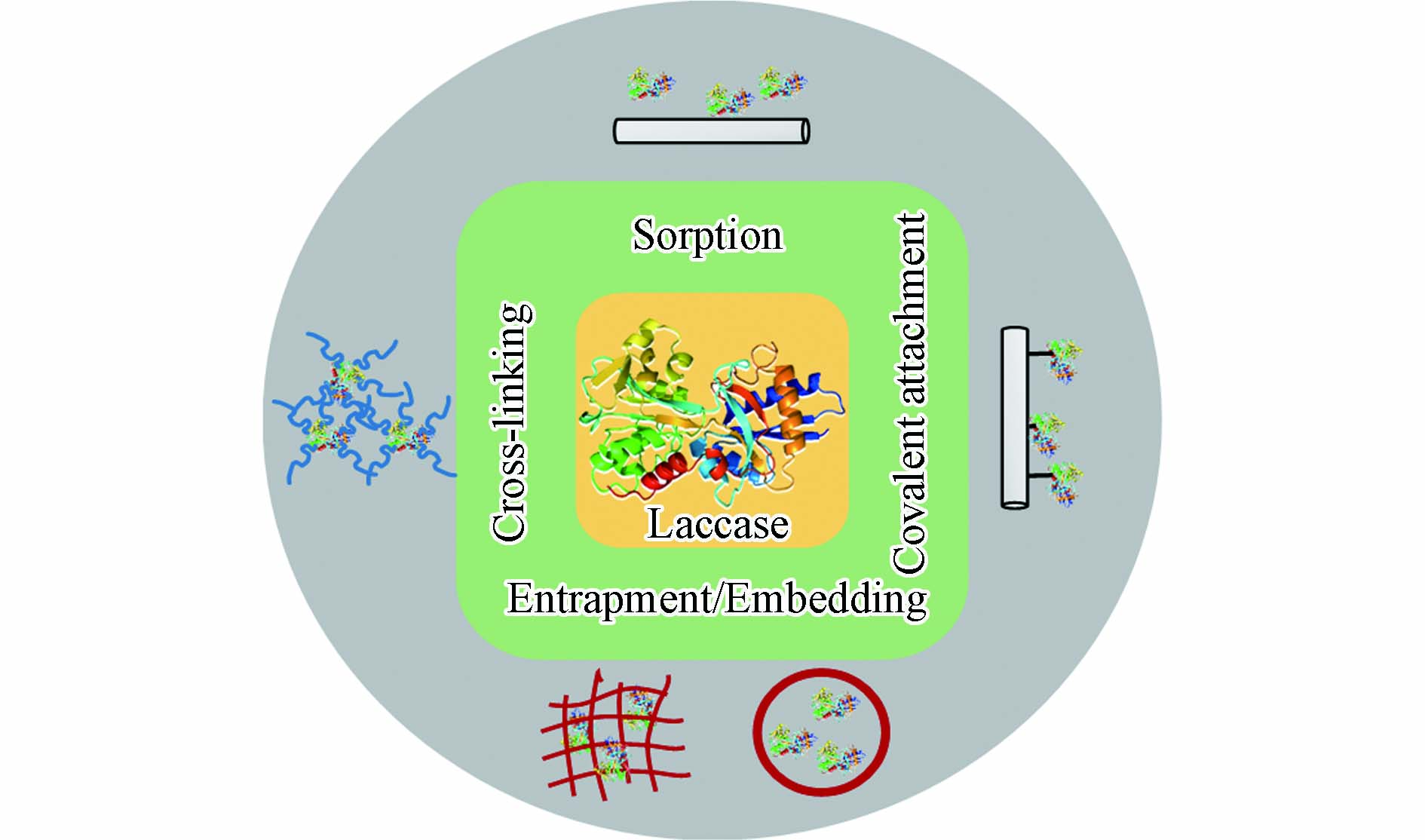
固定化技术是在保持真菌漆酶功能条件下, 通过物理或化学方法使溶液中游离态漆酶固载于不溶性载体上. 一旦真菌漆酶附着或包埋于载体基质后, 游离态漆酶便转化为固定化漆酶. 它有助于漆酶的回收, 并可循环重复使用, 进而降低了操作成本[37-48]. 如图2所示, 炭基材料固定真菌漆酶的方法可以分为物理固定和化学固定.
-
物理方法涉及非特异性相互作用来固定真菌漆酶, 例如, 吸附法、包埋法. 通过范德华力、疏水性作用或氢键等弱力作用将酶吸附于固态载体表面[49-51] . 物理吸附法被广泛应用于酶生物传感器的开发中. 它相对操作简单, 吸附前载体不需要官能化, 载体本身对于酶的固定也没有选择性. 因此, 吸附过程导致真菌漆酶非均一地固定于载体上. 炭基载体吸附真菌漆酶后, 酶结构不会发生明显改变, 活性中心不易被破坏. Lonappan 等[52]利用不同源生物炭吸附漆酶, 于猪粪生物炭、杏仁壳生物炭、松木生物炭上负载量分别为31. 4、24. 3、14. 58 U·g−1. Skoronski等[53], 通过大比表面的石墨烯纳米片层吸附漆酶, 并发现漆酶负载量可达179 mg·g−1. 然而, 由于漆酶吸附于载体的作用力较弱, 循环利用过程中漆酶也易于再次脱落, 漆酶的负载率会不断降低. 包埋或镶嵌法是将漆酶物理包裹或镶嵌于在微囊或高聚物网格内. 由于酶蛋白分子相对较大, 漆酶易于被限制在微囊或网格的空间内, 且漆酶的分子结构也不会发生改变[5,54]. 而分子量较小的污染物则可以畅通进入微囊或网格空间内, 当污染物分子接近该空间时, 被固定化的漆酶将参与污染物的催化氧化过程[55]. Shao等[41]通过空心介孔炭纳米球固定了游离的漆酶, 发现漆酶的固定化效果极为稳定, 且对于抗生素类污染物具有良好的去除效果.
-
化学固定包括共价结合、交联法等, 可以有效提高固定化漆酶的负载率, 增强炭基载体与真菌漆酶之间的结合作用. 相较于物理方法, 化学固定使得固定化漆酶具有更好稳定性, 耐酸碱性, 从而更好地实现固定化漆酶在实际应用中重复利用[56-57]. 共价固定法主要是通过共价键结合的形式, 促使漆酶蛋白的官能团与载体上的活性基团以共价键结合, 避免漆酶蛋白结构的断裂, 促进真菌漆酶更为有效地附着到载体上. 通常, 真菌漆酶结构中半胱氨酸、赖氨酸、天冬氨酸和谷氨酸的侧链可与载体上的醚基、羧基、酰胺键形成共价键[58]. 共价键的连接增强了真菌漆酶在固体载体上负载, 在循环利用过程中真菌漆酶不易发生脱落, 提高了固定化酶的操作稳定性[59]. Xu等[42]利用多壁纳米碳管以共价结合的方式负载漆酶, 获得了具有高稳定性的固定化漆酶. 交联法是使用交联剂促使漆酶分子发生交联反应, 诱导蛋白结构彼此连接, 从而增加漆酶分子负载在固体载体上的效果. 常见的交联试剂有戊二醛、己二胺、顺丁烯二酸酐、双偶氮苯等. 但是, 由于交联剂也会参与漆酶蛋白的结合, 这会导致漆酶本身构象结构或活性中心发生改变, 可能会降低漆酶分子的催化活性[60]. 在许多研究中, 共价结合与交联法被协同运用来增强炭基材料固载漆酶的效果. 例如, Lonappan等[36]以戊二醛作为交联剂连接漆酶蛋白, 并通过酸化过程丰富生物炭界面含氧官能团(如羧基官能团), 以共价结合的方式利用官能化生物炭对交联的漆酶蛋白进行有效固载. 即使在pH =10. 5条件下, 生物炭固定化漆酶的活性也可达到61%.
-
已有大量研究表明, 炭基材料在控制水体有机化合物运移方面具有重要作用. 它们对有机污染物的吸附过程涉及多重吸附机制. 例如, 非特异性的范德华力(色散力, 诱导力, 取向力)被认为普遍存在于炭基材料与有机化合物之间. 通常, 范德华力的能量相对较低, 约在0. 4—4 kJ·mol-1[61]. 由于有机化合物的自身疏水性, 这导致液相溶质化合物易于疏水性分配到固相炭基材料表面. Peng等[62]研究了两种抗生素药物(氧氟沙星和诺氟沙星)在碳纳米管(CNTs)上的吸附行为, 评价它们的结构和疏水性对吸附过程的重要作用. 研究发现, 单点吸附系数(Kd)与高疏水性有显著正相关关系, 而与结构含氧量无显著关系. 这结果表明, 位点特异性吸附没有起到主要作用, 而疏水效应可能是CNTs吸附氧氟沙星和诺氟沙星的重要机制. Dong等[35]以石墨烯为基体, 经过非共价π-π堆叠自组装过程固定化漆酶, 形成石墨烯-漆酶复合材料. 合成的复合材料具有优良的吸附性能和催化活性, 能有效地去除β-受体阻滞剂拉贝洛尔(Labetalol). 漆酶的催化中心的暴露增强了有机化合物的转化过程. 磷酸催化交联机制有助于制备活性炭的活化能降低, 且使活性炭的微孔大量生成. 分子型化合物双酚A 和卡马西平在磷酸改性活性炭的吸附均有数量级增长, 这主要归因于活性炭的微孔填充作用[63]. 固定化漆酶复合结构的炭基材料依然保持着其独特的结构优势, 它有利于富集水相里有机物. Shao等[41]利用空心的碳纳米小球将漆酶封锁于球体内部, 固定的漆酶呈现出优良的循环效率和稳定性能. 为了解碳纳米小球自身在吸附过程中所起的作用, 他们对漆酶固定化炭球进行灭活, 并利用灭活后的复合材料去除四环素和环丙沙星, 去除率分别可达55%和77%. 两种抗生素在炭球上的高效截留可能是由于碳纳米小球的构象结构所致. Ramírez-Montoya等[34]采用介孔炭基支撑漆酶, 该固定化漆酶炭基材料能有效对酸橙7染料溶液进行脱色处理. 研究者发现, 炭基载体除了作为漆酶的固定支撑外, 其本身也通过物理吸附移除有机化合物.
-
固定化漆酶结构中T1 Cu同样承担分解污染物夺取电子的功能, 其氧化还原电势高低直接影响着固定化漆酶的催化氧化能力(图3). Sun等[64]设计了一种海藻酸钙交联捕获漆酶的流化床反应器, 并发现固定化漆酶对雌激素的持续清除能力具有优异潜力. 然而, 他们发现相对较低或较高pH下, 交联捕获的漆酶分子的催化氧化能力减弱, 这主要是由于阻断了雌激素在T1 Cu位点的单电子氧化或阻碍了T1 Cu 和T2/T3 Cu 位点之间的分子内电子转移. Nguyen等[65]采用活性炭固定化漆酶材料催化降解一系列小分子污染物 (如磺胺甲恶唑、卡马西平、双氯酚酸和双酚A). 与等浓度游离漆酶相比, 发现固定化漆酶对污染物的去除率均有所提高, 同时也指出漆酶与污染物之间电子转移受阻可能会限制漆酶氧化效率. 活性炭固定漆酶后, 吸附于炭基的污染物与漆酶分子之间的电子转移增强, 有助于污染物降解转化效率的提升. 曾涵等[66]以聚芳酰胺与碳纳米管为载体, 固定化云芝漆酶后, 将其对玻璃电极进行修饰, 可实现漆酶与电极间的直接电子转移. 这表明炭基材料协助漆酶传递电子的作用. 另一方面, 真菌漆酶本身可能不能有效氧化一些聚合度高的有机化合物, 但可在漆酶作用下形成高氧化还原电势的中间介质或具有强反应活性的自由基参与到污染物的降解过程中. 固定化漆酶以溶解氧作为电子受体间接催化氧化酚类或非酚类化合物, 不仅可以形成小分子产物, 也可以通过聚合形成大分子聚合物, 这极大地拓宽了可反应的目标污染物范围[67]. 例如, 真菌漆酶反应体系中引入介质ABTS, ABTS可被漆酶氧化形成稳定的ABTS+自由基, 可以提高漆酶的底物特异性. Dong等[35]探讨了ABTS 对于石墨烯-漆酶复合材料去除拉贝他洛尔效率的影响. 由于漆酶不能直接作用于拉贝洛尔, 因此将介质ABTS转化为高氧化性能的ABTS+ 自由基有助于拉贝洛尔的降解转化. 研究发现, ABTS+自由基可将拉贝洛尔氧化为过渡态中间体, 随即过渡态中间体在N-Cβ键处断裂, 进而生成两个构象不稳定的中间体. 其中, 一个中间体可与H+反应形成分子量149的最终产物, 而另一个携有一个未配对电子的中间体的酰胺基可能发生水解反应, 形成分子量182的最终产物.
-
近年来, 频繁在湖泊、河流等水体中检测出酚类、雌激素、药物等环境污染物. 它们可在水体生物体内富集, 并影响生物内分泌系统和人类健康. 许多研究已经报道, 固定化漆酶在氧化分解酚类化合物和雌激素污染方面的功效. Wang等[36]利用溴化十六烷基三甲基铵和氢氧化钾溶液改性秸秆生物炭, 并以其固定化漆酶, 利用该复合材料对于2, 4-二氯苯酚的去除效果. 研究者发现, 60 min内生物炭吸附2, 4-二氯苯酚的效率仅为42. 5%, 而固定化漆酶对2, 4-二氯苯酚的去除率接近100%. Lonappan等[47]利用杏仁壳、猪粪和松木制备生物炭进行漆酶固定化, 研究了二氯苯酚在3种固定化漆酶炭材料上的氧化降解过程. 研究发现, 5 h内3种固定化漆酶炭材料对二氯苯酚的去除均可达到100%. Liu等[68]利用磁性介孔炭固定化漆酶对苯酚和对氯苯酚进行去除. 相较于磁性介孔炭, 固定化漆酶对两种酚类污染物的去除率分别提高了58%和64%. Zhang等[40]以丝瓜海绵基磁性碳纳米材料为载体固定漆酶, 去除水中雌激素双酚A. 该团队发现与游离漆酶相比, 100 mg·g−1的双酚A 经固定化漆酶反应4 h后, 目标污染物几乎被完全移除, 脱除效率提高了37. 28%. 工业生产带来的药物、染料类废水的不合理排放也造成了日益突出的环境污染问题, 导致生态环境破坏和生物安全危害. Masjoudi等[44]利用了碳纳米管复合漆酶对水体中卡马西平和双氯芬酸氧化降解. 由于卡马西平含有酰胺(-CONR2)吸电子基团, 它削弱了固定化漆酶结构中T1 Cu捕获电子的能力, 其降解效率约为27%. 然而, 碳纳米管-漆酶复合材料对于双氯酚酸的去除则高达95%. 杨奇等[28]研究了碳纳米管固定化漆酶脱除水中刚果红染料效果. 研究发现, 漆酶固定化前后, 碳纳米管对刚果红染料的最大去除量由363. 4 mg·g−1增加到769. 23 mg·g−1. Habimana等[45]利用交联聚合方法对包覆二氧化硅的磁性碳纳米管进行漆酶的固定化, 并对几种偶氮染料(酸性红88、铬黑T和活性黑5)进行脱色处理. 酸性红88、铬黑T、活性黑5经固定化漆酶氧化分解, 去除率分别可达99%、98%、66%.
-
固定化漆酶的目的在于重复循环利用, 这是工业和环境应用的必要需求. 炭基材料固定真菌漆酶的技术极大程度上避免了游离漆酶在应用上的资源浪费. 例如, 据Habimana等[45]研究报道, 固定化漆酶的稳定性能得到大幅度提高, 通过ABTS的10 次氧化, 其余量酶活仍可保留87% (生物催化剂的运行稳定性得到了提高, 循环10次后仍能保持原有活性的87%. Dong等[35]发现在完全脱除拉贝洛尔物质条件下, 石墨烯-漆酶复合材料可重复使用10次以上, 完美实现了固定化漆酶的循环利用. 通过刚果红染料脱色研究, Samak 等[39]发现螯合了Cu2+的磁性氧化石墨烯所固载的漆酶, 经过5次反复使用, 对于刚果红的脱色率仍高于95%, 10次循环后, 固定化漆酶的降解效率才下降至89. 4%. 据Zhang 等[40]报道, 以丝瓜络制备的磁性活性炭所固载的漆酶, 展现出良好的循环性能. 循环5次测试, 固定化漆酶活性仍可高达92. 51%, 而水中超过80%的双酚A可通过固定化漆酶被净化. Thiyagarajan 等[69]利用Prosopis juliflora 树皮制备的活性碳纳米管用于固定化漆酶, 研究了固定化漆酶的稳定性、循环性以及对于品红染料的吸附性能. 循环性试验中, 6次循环试验后游离漆酶的活性下降到仅20%, 而固定化漆酶的活性仍然保持在80%以上. Li等[70]利用枫树生物炭作为固定化材料, 通过物理吸附固定化漆酶, 并利用ABTS测试了该种固定化漆酶循环性能, 他们发现7次循环试验后, 固定化漆酶的酶活损失了66. 2%. Ramírez-Montoya等[34]利用核桃与坚果壳制备的活性炭对漆酶进行固定, 并利用酸性橙7测试固定化漆酶循环性能. 研究发现, 4次循环试验后, 该种固定化漆酶对酸性橙7的去除率保持在60%. 此外, 相较于其他类型固定化载体, 炭基材料也呈现出优越的机械支撑性能. 同作为固定化载体, 乳酸-甘醇酸共聚体和多壁碳纳米管均可固定化漆酶, 且均能有效降解水体中双氯芬酸. 但是, 乳酸-甘醇酸共聚体固定化漆酶5次循环后双氯芬酸转化效率低于60%, 而多壁碳纳米管固定化漆酶7次循环利用后却仍可保持62.7 %的双氯芬酸去除效率[42,71].
-
综上所述, 炭基材料固定化真菌漆酶是一种具有潜力的绿色环保技术. 该方法可以提高游离漆酶反应活性和贮存稳定性, 降低游离漆酶在水处理应用中产生的成本. 本文结合国内外前沿的科学研究, 系统概述了真菌漆酶结构和性质、炭基材料固定漆酶的方法、以及污染物和固定化漆酶的反应机制和循环效率, 为固定化漆酶技术在绿色化学应用研究提供了新的视角. 目前, 关于炭基材料固定化漆酶技术仍可从以下几方面着手进一步研究:
(1)结合改性修饰等方法, 突破炭基材料固定化漆酶技术, 进一步增强真菌漆酶负载量、贮存稳定性、催化氧化性能、以及循环利用效率等.
(2) 重点关注实际工程应用的局限, 拓宽炭基材料固定化漆酶在环境有机污染治理方面的前景价值.
炭基材料固定化真菌漆酶在污水处理领域的应用
Application of immobilized fungal laccase on carbonaceous materials for wastewater treatment
-
摘要: 漆酶是一种多功能含铜氧化还原酶,被广泛研究或应用于多个学科领域. 然而,实际应用中,漆酶因成本昂贵、易流失,稳定性不足等缺陷,限制了其工业化应用和商业化价值. 炭基材料固定化真菌漆酶技术提供了一种值得探索的途径,来改善真菌漆酶的贮存稳定性、催化氧化活性、以及重复使用的性能. 本文详细介绍了真菌漆酶的来源、结构和性质;系统归纳了生物炭、活性炭、碳纳米管、石墨烯等炭基材料固定化真菌漆酶的物理-化学方法(物理吸附、诱捕包埋、共价结合、交联聚合);概述了酚类、雌激素、染料、药物等环境有机污染物在炭基固定化真菌漆酶复合材料上的去除效果和循环性能,并深入讨论了化合物在炭质界面的吸附机制(如疏水性作用、π-π作用、孔隙填充等)和固定化真菌漆酶利用氧化还原电势差催化氧化有机物的反应机制. 本文旨在为真菌漆酶固定化载体材料的选择提供参考依据,也将为开拓固定化真菌漆酶应用于有机污染治理的视野提供技术支撑.Abstract: Laccase is one of multi-copper containing oxidoreductases, which has been widely studied or applied in multiple subject fields. However, the industrial application and commercial value of laccase is limited in practical production due to expensive cost, difficult recycling, insufficient stability. The immobilization of fungal laccase by carbonaceous materials provides an approach to explore to improve the storage stability, catalytic oxidation activity, and reusability. This work described the origin, structure and properties of fungal laccase, and systematically concluded the physico-chemical methods including physical adsorption, trapping and embedding, covalent binding, crosslinking. Meanwhile, a series of carbonaceous materials such as biochar, activated carbon, carbon nanotubes, graphene was summarized to immobilize the fungal laccase. The removal of organic contaminants (phenols, estrogen, dyes, and drugs) and recycling performance of immobilized laccase were outlined and its reactive mechanisms including sorption and catalytic oxidation were deeply discussed between immobilized laccase and environmental contaminants. This article aims at providing an insight to select the carrier materials to the immobilization of fungal laccase, and also providing a technical support for expanding the application scope of immobilized laccase in organic pollution control.
-
Key words:
- carbonaceous material /
- fungal laccase /
- immobilization methods /
- sorption and degradation /
- reusability
-
塑料因其重量轻、耐腐蚀、化学性质稳定等优点,被广泛应用于人类生产生活[1]. 目前所生产的塑料中,有近80%最终会成为塑料垃圾,被掩埋或堆积在环境中[2]. 环境里的塑料垃圾中有相当大的一部分是直径或长度小于5 mm的颗粒,也被称为微塑料[3]. 微塑料的来源包括较大尺寸塑料垃圾的分解、破碎,以及微米尺寸塑料颗粒的直接生产、使用和处置[4]. 目前针对环境微塑料污染的研究主要集中在陆地环境和水环境. 陆地环境中的微塑料主要来源于垃圾填埋场中塑料制品的破碎、污泥农用、农业地膜破碎和汽车轮胎磨损[5-6]. 水环境中的微塑料主要来源于污水处理厂排放、渔业活动以及船舶运输业释放[7-8]. 微塑料广泛分布在土壤、海洋、河流、湖泊等环境介质中,即便在偏远地区(如极地、高山、深海沉积物中)也均有检出[9-12]. 据估算,陆地环境中每年新增的微塑料超过43万吨[6],而海洋中至少漂浮着5.3万亿个微塑料颗粒[13]. 微塑料易被生物体吞食,诱导氧化应激和炎症反应进而引发颗粒毒性,从而造成肝脏、肾脏、胃肠道等器官损伤[14-15]. 同时,微塑料在生产过程中会混入大量添加剂,例如邻苯二甲酸酯、多溴联苯醚等;生物体摄食微塑料可导致添加剂在体内的释放和积累,进而引发化学毒性,显著抑制生物生长和早期发育[16-17]. 此外,微塑料粒径较小、比表面积较大,进入环境后可成为众多有机污染物、重金属以及微生物的载体,促进了这些污染物的生物富集,造成严重的生态和健康风险[18]. 因此,环境中微塑料的大量存在及其环境风险近年来已在全球范围内引起了广泛关注.
环境中的微塑料可在机械力、紫外线、高温、化学物质和生物等环境因素作用下发生一系列的物理、化学和生物老化[7, 19-22]. 环境老化使得微塑料表面形貌和理化性质发生改变,进而影响微塑料迁移和吸附污染物的能力,改变生物体摄入微塑料的方式和程度. 同时,老化增加了微塑料中添加剂向环境中释放的风险,增强了毒性效应[23-25]. 因此,研究微塑料的环境老化及由此产生的效应具有重要意义. 本文通过综述国内外微塑料环境老化研究的最新进展,总结了物理、化学和生物老化的主要过程与机制,分析了老化对微塑料的环境行为和效应的影响.
1. 微塑料的环境老化机制(Mechanisms controlling environmental aging of microplastics)
微塑料广泛分布在自然环境中,可在机械力、紫外光、高温、化学物质、生物等因素作用下发生老化,其表面形貌和理化性质均可发生变化(图1). 明确微塑料的老化机制对于理解其环境行为和风险至关重要. 本文从物理、化学和生物的3个方面阐述了微塑料在环境中的老化过程及机制(图2).
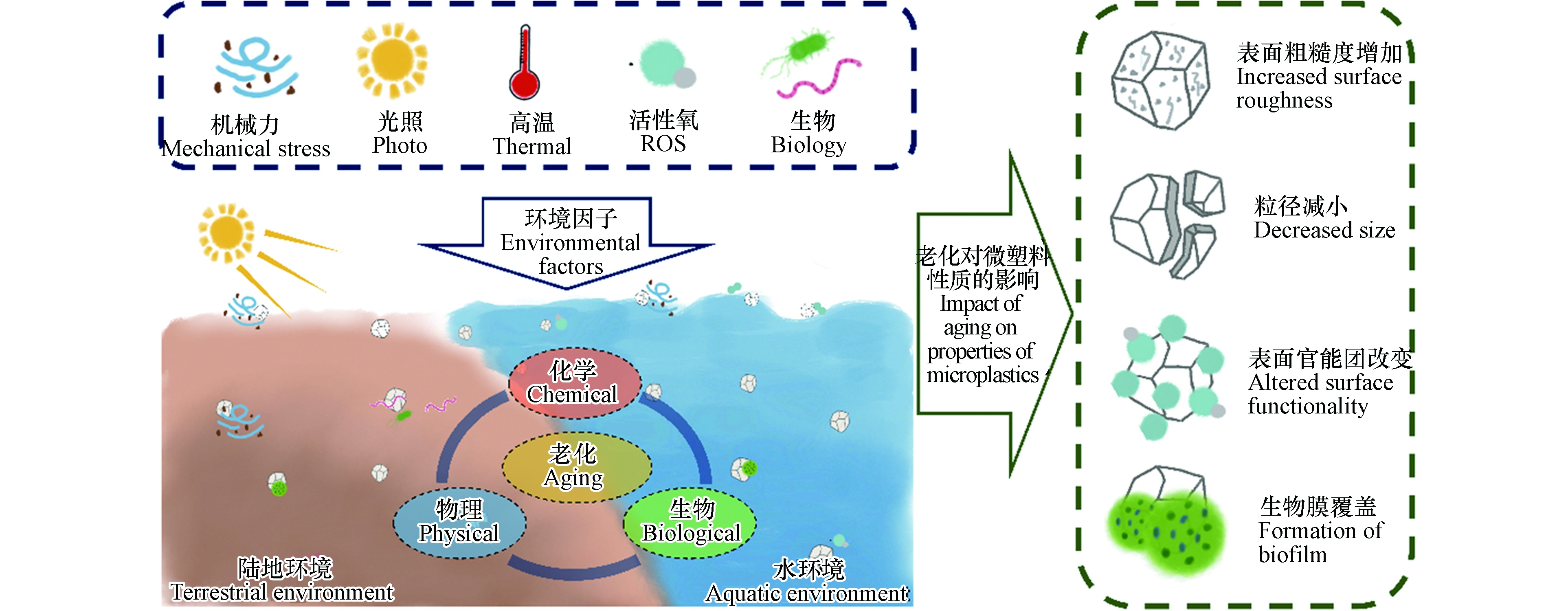 图 1 微塑料进入环境后可在多种环境因子的影响下发生复杂的物理、化学和生物老化,导致其理化性质显著改变Figure 1. Microplastics can undergo a series of physical, chemical, and biological aging processes, resulting in significantly altered physicochemical properties
图 1 微塑料进入环境后可在多种环境因子的影响下发生复杂的物理、化学和生物老化,导致其理化性质显著改变Figure 1. Microplastics can undergo a series of physical, chemical, and biological aging processes, resulting in significantly altered physicochemical properties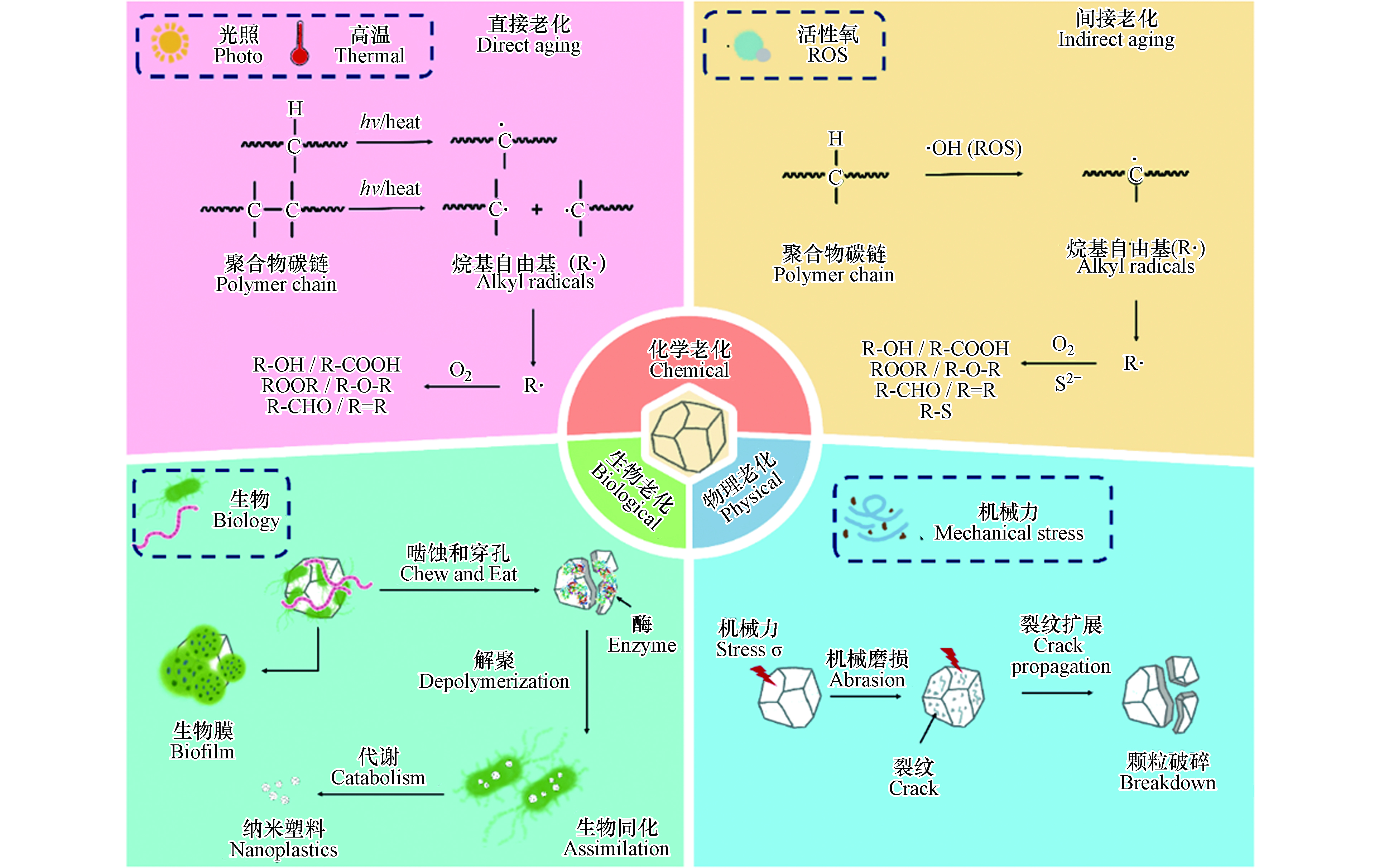 图 2 微塑料进入环境后的物理、化学和生物老化机制Figure 2. Mechanisms controlling physical, chemical and biological aging processes of microplastics
图 2 微塑料进入环境后的物理、化学和生物老化机制Figure 2. Mechanisms controlling physical, chemical and biological aging processes of microplastics1.1 物理老化
进入环境中的微塑料会受到周围环境力的机械作用,如湍流、波浪、沙砾或岩石碰撞及周围塑料碎片的碰撞等,从而发生表面开裂,破碎形成更多小尺寸的微塑料[26],所生成的塑料颗粒数量随尺寸的减小而增加[22]. 有研究显示,从磨砂膏中提取的粒径为398 nm的原生微塑料,可在模拟湍流作用下破碎为50 nm的微塑料颗粒,甚至出现了纳米塑料(<10 nm)[27]. 对海滩中采集到的塑料碎片进行分析发现,物理老化是微塑料表面纹理形成的主要原因,碰撞过程中塑料颗粒棱角会变成近圆形状,同时出现裂缝和凹坑[28]. 同时,已老化的塑料表面更易受沙子磨损等机械力作用而破裂[29]. 微塑料在机械力作用下表面产生裂纹,从而裂纹扩展,进而导致了微塑料颗粒的破碎[27, 30]. 相关研究使用Grady模型来描述微塑料在应力作用和裂纹扩展下的破碎,证明了裂纹在微塑料物理老化中发挥了决定性作用,裂纹越长,扩展时所需最小应力越低,因此,表面有裂纹的微塑料在剪切力的作用下更容易破碎、剥落成更小的颗粒[27]. 自然环境中的微塑料在土壤颗粒、沙砾磨损,海浪、潮汐碰撞等自然机械力作用下,原有的表面形貌被破坏、破碎并形成更多的裂缝,呈现更高的比表面积. 同时,物理老化有利于各种化学物质通过微塑料表面的孔隙和裂纹进入内部,并促进微塑料的进一步老化.
1.2 化学老化
微塑料可在紫外线、高温、活性氧等作用下发生化学老化. 在实验室模拟或自然条件下,老化后微塑料的表面整体呈现粗糙度增加,出现裂纹和缝隙,伴随片状、颗粒状碎片脱落,粒径减小,孔隙度和比表面积增大. 老化后的微塑料往往表面羟基、羰基等含氧官能团含量增高,表面亲水性增强[3, 31-32]. 微塑料化学老化的作用机制可大致分为直接老化和间接老化:直接老化是指紫外光或高温作用使得微塑料聚合物碳键直接断裂及由此引发的一系列变化;间接老化是指活性氧物种等化学物质参与导致化学键断裂而发生的系列化学反应.
1.2.1 直接老化
(1)光老化
光老化是微塑料环境老化最重要的过程[33]. 环境中微塑料的光老化本质上是聚合物碳链中的C—C、C—H等化学键吸收光能,从而直接断裂生成烷基自由基,随后引发的链式反应;光老化通过改变微塑料的化学结构和组成影响其极性、亲疏水性,并通过影响高分子链的排列改变微塑料的机械性能,诱导微塑料进一步破碎形成粒径更小的塑料颗粒,这也是环境中形成微塑料的重要途径. 光老化主要作用于微塑料表面,微塑料表面首先吸收紫外线,紫外线诱导聚合物链化学形态变化,导致链断裂和烷基自由基的形成,随后表面化学官能团解构和重组,从而分子量降低、表面氧化,最终形成惰性产物[3].
光老化过程通常分为3个步骤,分别是链引发、链传递和链终止(图2). 在链引发步骤中,C—C键(375 kJ·mol−1)和C—H键(420 kJ·mol−1)的键解离能相当于320 nm和290 nm处的紫外光能,因此,C—C和C—H键可以直接光解,生成烷基自由基(R· 初始自由基)[34]. 初始自由基(R·)一旦形成,就会迅速的增长、转移,进入链传递过程,而自氧化循环的传递反应对所有碳骨架聚合物都适用. 在链传递步骤中,R·首先与O2反应生成ROO·,ROO·从另一条聚合物链RH中提取氢原子生成氢过氧化物(ROOH),ROOH通过吸收光能裂解为·OH和RO·[35]. 随后RO·可以从RH中提取氢原子生成醇或者经电子离域β-裂解引起聚合物骨架链的断裂而生成酮[7, 34]. 在紫外线照射下,酮经过Norrish Ⅰ型反应生成R·和酰基(R—CO·)(自由基生成),或通过Norrish Ⅱ型反应生成末端羰基(R—CO—CH3)(链断裂)[23, 36]. 当双分子自由基结合或最终裂解形成酮、烯烃和醛等惰性产物时,链反应就会终止[37].
紫外线照射引发聚合物链骨架断裂并释放低分子量链片段,进而加速结构碎片化,导致介孔/微孔的产生和微塑料尺寸的减小[23]. 在半晶态聚合物中,光老化引发的断裂通常发生在无定形区域,导致老化后微塑料结晶度增加[38]. 此外,在模拟太阳光(光照强度分别比北纬0°和50°的自然太阳光高3倍和10倍)照射下,光老化可以使聚苯乙烯微塑料完全矿化成CO2[39].
(2)热老化
微塑料的热老化与光老化相似,都是聚合物链断裂后的氧化反应. 据估计,陆地环境中,深色干燥土壤的理论最高温度可达到90—100 ℃[40],在高温环境下,整个微塑料颗粒都会发生热老化,而光老化只发生在颗粒表面. 微塑料热老化的机制主要是高温有利于达到高分子化合物分子链间的化学键解离能,从而利于分子链的随机断裂和支链的脱落,生成烷基自由基(R·),随后进入自氧化循环(其过程类似于光老化)[7]. 温度是影响动力学反应速率的重要因素,高温可以加速微塑料的老化. 实验室人工老化箱模拟研究显示,随着温度的提高,微塑料薄膜的氧化程度更高,分子量、玻璃态转化温度和机械强度都显著下降,在60 ℃下,聚乳酸(PLA)薄膜的断裂伸长率在1周内便降低至初始值的一半,而在30 ℃下则需要15周[41]. 值得注意的是,自然界中的微塑料很少会经历单纯的高温热老化过程,热老化往往与光老化同时存在,温度的上升提高了紫外光照射下自由基类物质的生成速率,从而加速了微塑料的老化. 此外,自然界中普遍存在的干湿循环和冻融循环也伴随了热老化的过程,显著影响了微塑料的物理强度[42].
(3)其它因素导致的直接老化
环境中很多其它因素也会导致微塑料的直接老化,但相关研究较少. 例如环境中大量存在硫化物,硫离子可以与聚合物链中的C=C键发生亲核取代/加成反应,形成C—S键,进而硫化老化微塑料[43]. 此外,大气中臭氧会破坏不饱和聚合物的不饱和性,与聚合物碳链中C=C键、芳香环等直接加成,从而老化塑料[34]. 因此,即使臭氧浓度很低,也可能造成大气中微塑料的老化. 同时,在地下海底等极端环境中,高温高盐强酸强碱等作用可能导致微塑料的直接老化. 总的来说,目前对于这些环境因素导致微塑料直接老化机制的理解远远不及对光老化和热老化机制的认识,亟待进一步的研究.
1.2.2 间接老化
水环境中,天然有机质(NOM)可在光照下与溶解氧反应生成大量活性氧(ROS,包括羟基自由基(·OH),超氧阴离子(O2·–)和单线态氧(1O2))[44],微塑料中的添加剂(塑料生产过程中会掺入光催化剂,以制备环境友好的可降解塑料)也会在光照下与氧气和水反应生成活性氧[45]. 活性氧具有强氧化能力,可通过与微塑料的作用,诱导微塑料聚合物碳链断裂,从而发生间接老化. 例如,羟基自由基(·OH)可进攻聚合物长链中叔碳原子上的C—H键(叔碳比伯碳和仲碳更活泼),生成烷基自由基(R·). 烷基自由基与氧气反应形成过氧自由基(ROO·)(R·与·OH的反应也可以形成ROO·,但与氧气的反应在热力学上更有利,因为与氧反应的活化能接近于零). 过氧自由基可以从聚合物分子(RH)中提取氢原子,形成氢过氧化物(ROOH)[36]. ROOH裂解脱去—OH生成烷氧基自由基(RO·)[34],RO·可以从聚合物分子(RH)中提取氢从而生成醇类化合物,或在电子离域作用下发生裂解,形成含有C=O和C=C键的化合物[23,36]. 研究显示,模拟太阳光照射下,微塑料的间接老化是导致其表面粗糙、粒径减小、生成大量含氧官能团的重要原因;在加入ROS淬灭剂抗坏血酸后,微塑料老化程度(以羰基指数作为老化程度指标)降低55.5%,老化作用被显著抑制,说明ROS在微塑料老化过程中起重要作用[46].
硫化物是一种亲核试剂和还原剂,生物硫酸盐还原和污水排放使得硫化物普遍存在于环境中[47-48]. 许多富含硫化物的环境(如河口、湖泊和河流沉积物)也是微塑料赋存的重要区域[11, 49-50].本课题组前期研究发现,硫化物诱导的老化是微塑料在自然环境中的重要老化途径[43]. 在中性pH值下,硫化物可在水溶液中自氧化形成超氧阴离子和过氧化氢[51-52]:
HS−+O2→HS⋅+⋅O−2 (1) HS⋅+⋅O−2→S+HO−2 (2) HO−2+H+→H2O2 (3) 硫化物可以作为还原剂与H2O2反应,通过类芬顿反应生成·OH[53],从而促进了微塑料的表面氧化(图2). 硫化物诱导的间接老化可导致微塑料的脆化和开裂,颗粒粒径减小、比表面积的增加,硫化后热塑性聚氨酯(TPU)和聚苯乙烯(PS)的平均粒径分别降低了33.1%和27.3%,同时表面生成含氧官能团(C—O键、C=O键)和C—S键[43]. 有研究使用硫化钠处理聚酰胺(PA)、聚乙烯(PE)和聚苯乙烯(PS)3种微塑料,发现处理后的微塑料表面出现裂缝,比表面积和表面氧含量增加,其中处理后的PA和PS比表面积增加了3倍,PA羰基指数由0.05增长到0.14,3种材料玻璃态转化温度均降低[54],认为是其中硫化物诱导的间接老化起主要作用.
同时,本课题组发现,在硫化物诱导的老化过程中,自由基和小分子之间可能发生交联反应,这会导致聚合物分子量的增加[43]. 硫化体系中可能会出现多硫化物[55],多硫化物作为更强的还原剂,能促进分子氧的激活,加速老化反应. 微塑料硫化反应中C—S键的形成则是通过亲核取代/加成途径[56-57]. 微塑料首先被·OH氧化,在断链过程中生成C=C键,S2-或多硫化物作为亲核试剂进攻共轭双键形成巯基(图2)[43]. 与硫结合的氢原子比与碳结合的氢原子更倾向于与·OH反应[58],因此硫的加成进一步促进了聚合物的氧化.
1.3 生物老化
生物作用是微塑料环境老化中的重要因素. 有关微塑料生物老化的研究目前主要集中在两个方面,一是微塑料被生物摄食后,生物体内特定的酶对微塑料的消化老化过程;二是微塑料进入自然环境后被水体中常见的生物质(如微生物群落、胞外聚合物等)覆盖形成生物膜的过程. 例如,用从蚯蚓肠道中分离出来的革兰氏阳性细菌与微塑料一起孵育21 d后,低密度聚乙烯(LDPE)的降解率可达60%,粒径从53.1—41.3 μm降至35.4—23.6 μm,甚至检测到纳米尺度颗粒[59]. 微生物定殖和生物膜的形成会改变微塑料的比表面积和含氧官能团(C—O和C=O)含量,进而影响其亲疏水性[60]. 微塑料表面生物膜的总量随其在水体中暴露时间显著增加,但随水深而减少[61].
1.3.1 生物摄食消化
微塑料被生物摄食进而被其体内的酶消化后,表面形貌、理化性质等会发生一系列的改变. 动物摄食过程中,微塑料会因动物咀嚼行为产生的物理应力而破碎,随后被吞食进入肠胃;某些动物的肠液、植物的根系中一些特定的酶能将微塑料作为其生长的能量基质进行消耗,使聚合物的分子链断裂、缩短、氧化(图2). 例如,南极磷虾可以通过咀嚼将摄入的微塑料(31.5 µm)破碎成直径小于1 µm的颗粒,并随粪便排出体内[62]. 黄粉虫可以啮食PS,并在24 h内将其降解矿化为CO2或同化为虫体脂肪[63]. 这一过程中,PS首先被黄粉幼虫咀嚼成细小碎片并摄入肠道中,所摄食的碎片在肠道微生物所分泌的胞外酶作用下,解聚成小分子产物,这些小分子产物在多种肠道细菌和黄粉幼虫自身酶的作用下,进一步降解并同化形成幼虫自身组织;残留的碎片与部分降解中间产物,混合肠道微生物,以虫粪的形态排泄出体内[64]. 100只黄粉虫每天可以摄入34—39 mg的PS,其中有47.7%被矿化为CO2,49.2%被老化为微纳米塑料并随粪便排出. 将PE与Zalerion maritimum(一种真菌)培养28 d后,微塑料表面出现不规则断裂,同时表面发生氧化反应,生成羟基羰基等含氧官能团,羟基含量(红外光谱3700—3000 cm−1处峰面积)由原始的1±1增长到了57±2,羰基含量(红外光谱1700—1500 cm−1处峰面积)由原始的0.2±0.3增长到了1±1[65].
1.3.2 生物膜覆盖
为适应不同生存环境,微生物进化出了各种附着机制[66]. 微塑料具有较大的比表面积,微生物群落很容易在其表面附着增殖,黏附并形成生物膜[60, 67]. 微塑料表面的生物膜形成主要分为3个阶段:(1)环境中的生物质和生物群落在范德华力和静电引力的作用下吸附在微塑料的表面,形成微生物有机质层;(2)吸附在微塑料表面的微生物群落释放出胞外聚合物(EPS),形成对微生物群落具有保护效应的EPS层;(3)微塑料表面微生物群落大量增殖,形成复杂的生物膜结构[66]. 在微塑料表面形成生物膜的微生物群落(藻类、真菌、细菌)组成随水体环境条件(pH值、盐度、温度、营养元素)和微塑料表面性质(种类、颜色、粗糙度)的不同而不同,其中细菌是最常被观测到的微生物种类[68]. 例如,在盐度26‰的水体中,定殖在微塑料表面的弧菌丰度是海水和沉积物中的2—10倍,而在盐度小于18‰的水体中,微塑料表面、海水、沉积物中弧菌的丰度没有区别;同时,PS表面生物量显著高于PVC、PP、PE和TPU[69]. 生物膜的形成可显著改变微塑料的理化性质,包括:(1)改变其表面形貌,导致比表面积显著的增大,也可增加其表面的粗糙度;(2)改变其硬度和结晶度;(3)改变其密度,从而影响其在水体中的纵向分布;(4)改变其表面官能团,C—O、C=O含量上升,进而影响其亲疏水性[70]. 研究显示,在海水中原位老化6个月的4种微塑料纤维表面形成了生物膜,表面含氧官能团增多,并出现了多糖类化合物,其中PE的比表面积由(0.38±0.06) m2 g−1增加至(0.65±0.08) m2 g−1,微孔直径由0.76 nm增加至0.95 nm[60].
1.4 其它老化
微塑料在进入环境后可能会经历其它老化过程,其中微塑料对腐殖质等天然有机质(NOM)的吸附受到了广泛关注. 微塑料可以通过疏水作用、静电作用、π-π相互作用等吸附水中的NOM[71],其作用机制和效应与微塑料对EPS的吸附类似. 微塑料吸附NOM后,表面形貌和理化性质显著改变,进而影响其环境效应. 原始PS微塑料颗粒表面光滑,NOM吸附后PS表面粗糙,粒径增大,表面含氧官能团增加[72]. 天然有机质包覆在微塑料表面后增加了微塑料表面电负性,从而加强静电斥力,增强了颗粒稳定性[73]. 一些综述也关注到了腐殖质对微塑料的老化[3,20],未来仍需进一步系统的探索不同类型微塑料与腐殖质不同组分之间的相互作用.
1.5 微塑料环境老化的研究方法
目前研究微塑料环境老化的手段主要有两种,一种是从环境中(如海滩[74]、农田[73]、河流[75]等)直接采集老化的微塑料颗粒开展研究,另一种是人工模拟老化过程,获得老化微塑料进行研究. 人工模拟老化又分为原位自然老化和实验室加速老化[19-20]:原位自然老化是指将购买或制备的微塑料颗粒置于特定自然环境中,老化后回收以进一步研究[60],例如有研究使用长江和太湖的天然水样制备微塑料悬浮液,将其置于屋顶暴露于阳光下5至11个月[45]. 实验室加速老化是指通过机械力磨损、紫外灯照射、化学氧化等方式加速老化微塑料. 例如,有研究将海滩沙砾与微塑料颗粒混合振荡两个月以模拟物理老化过程[22]. 大量研究选择氙灯、汞灯等光源模拟环境中微塑料的光老化[46, 76-77]. 也有很多研究利用Fenton反应和过氧化氢等高级氧化手段,模拟微塑料在活性氧作用下的化学老化[45, 78]. 近期一项研究对比了实验室加速老化与原位自然老化微塑料的性质差异,发现经过高温、紫外、臭氧和超声组合手段老化后的PS微塑料与在室外阳光照射老化11个月的微塑料颗粒表面形貌与理化性质相似,表明多种实验室加速老化手段的综合应用能够较好地评估环境中微塑料的老化过程[79].
2. 老化对微塑料环境行为和生物效应的影响(Effects of aging on environmental processes and implications of microplastics)
环境老化可改变微塑料的表观形貌、分子结构及化学组成等理化性质,从而显著影响其环境行为和生物效应(如图3所示).
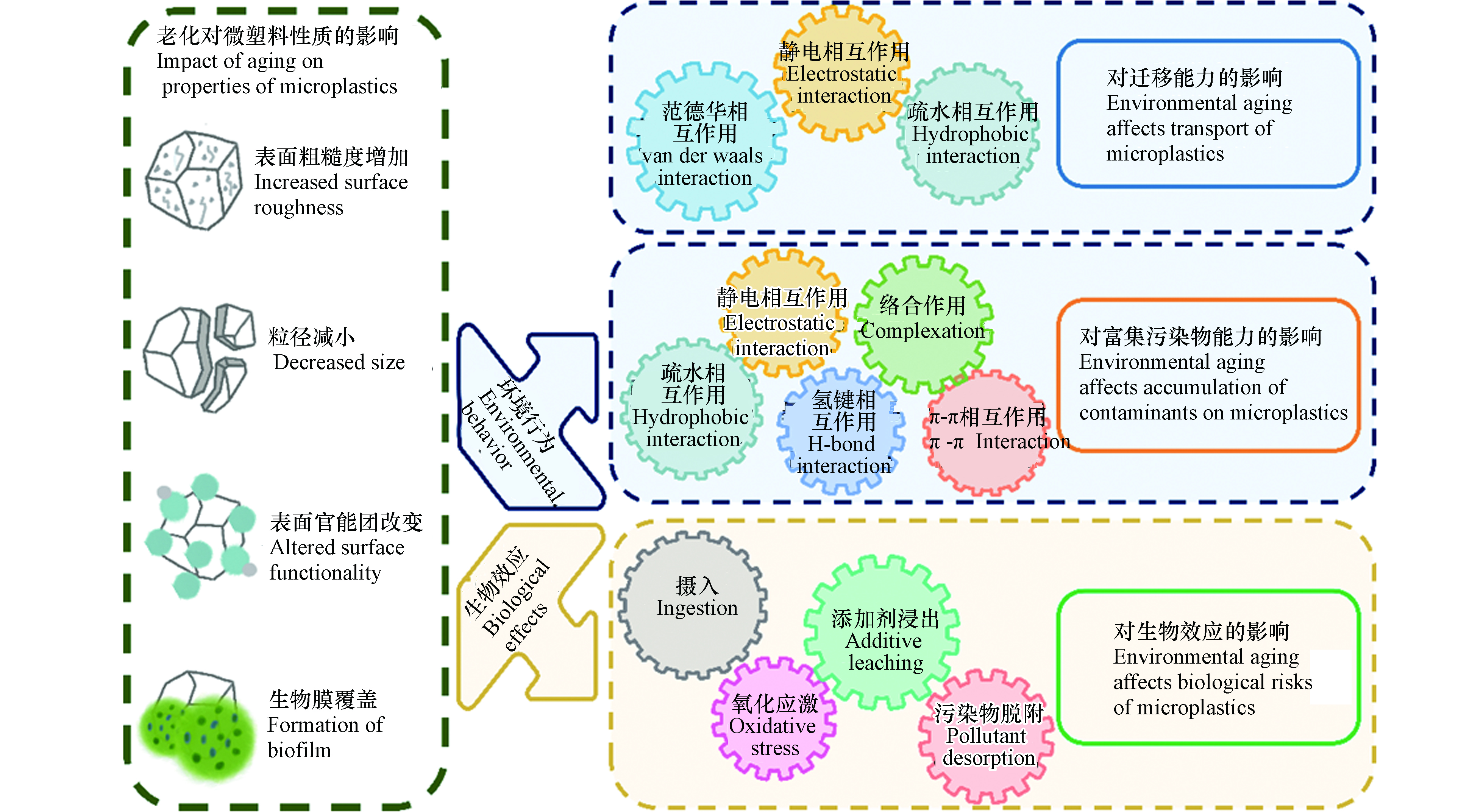 图 3 微塑料的环境老化对其环境行为和生物效应的影响Figure 3. Influences of aging of microplastics on their environmental behavior and biological effects
图 3 微塑料的环境老化对其环境行为和生物效应的影响Figure 3. Influences of aging of microplastics on their environmental behavior and biological effects2.1 环境老化对微塑料迁移能力的影响
微塑料可在环境中长期存在,具有较强的迁移能力. 环境老化过程会显著改变微塑料的物理化学性质,使得微塑料粒径减小、表面含氧官能团增加,影响了其与环境介质间的相互作用,进而显著影响了微塑料的迁移能力(图3). 老化后微塑料粒径的减小会削弱微塑料与多孔介质间的范德华力,微塑料在多孔介质中沉积减弱,迁移能力增强. 在农田自然老化所形成的小尺寸颗粒(15—30 µm)在天然土壤介质中具有更强的向下迁移能力[73]. 老化后微塑料表面含氧官能团含量增加,电负性增强,与带负电的多孔介质间的静电斥力增大,导致微塑料在多孔介质上的沉积减弱,有助于提升迁移能力. 例如,紫外老化的微塑料表面羧基官能团增加,羧基去质子化使其带有更多的负电荷,与带负电的石英砂介质之间静电斥力增大,迁移能力增强[80]. 此外,老化后微塑料表面含氧官能团的增加会显著增加微塑料的亲水性,进而影响其迁移行为. 例如,在背景溶液为0.5 mmol·L−1 NaCl中,原始PS最大穿透率约为43.3%,经紫外(UV)老化后的PS最大穿透率显著提高至90.7%;UV老化后的PS表面含氧官能团增加,这不仅增加了微塑料的表面电负性,还增强了其亲水性,亲水性的增强是老化PS迁移能力增强的关键因素[25]. 值得注意的是,老化后微塑料表面含氧官能团的增加也会导致其在Ca2+等阳离子条件中桥联作用的增加,这可能在一定程度上抑制老化微塑料的迁移能力.
2.2 环境老化对微塑料富集污染物能力的影响
老化过程可导致微塑料吸附污染物的能力发生明显变化. 老化后微塑料的表面更粗糙,出现裂缝和孔隙,粒径减小,比表面积增大;同时,生成的官能团改变了微塑料表面的电负性、极性和亲疏水性,从而影响了静电相互作用、疏水相互作用、π-π相互作用、氢键和络合作用等多种作用机制,进而改变了微塑料富集污染物的能力(图3). 首先,微塑料的吸附容量与比表面积呈正相关,老化后的微塑料产生裂纹或破碎为更小的碎片,粒径减小、比表面积增加,有效吸附位点增多从而具有更强的吸附能力. 例如,紫外照射导致PET微塑料比表面积增大,暴露出更多的有效吸附位点,对Cu2+的吸附能力是原始微塑料的2倍[81]. 其次,老化后微塑料表面生成了含氧官能团,表面氧碳比(O/C)增高,亲水性增加,这将抑制对疏水性污染物的吸附,增强对亲水性污染物的吸附[82-83]. 老化微塑料表面官能团的变化还会影响其与污染物之间的极性作用,增强氢键在吸附过程中的贡献从而改变微塑料的吸附能力. 例如,高温老化的PS表面因交联和断链产生的大量含氧官能团(羧酸、酮和内酯),增强了微塑料与抗生素之间的氢键作用,从而增加吸附容量;而老化后PS与周围的水分子间氢键作用的增强,降低了PS表面吸附位点的可利用性,极大地抑制了多环芳烃的吸附[84]. 在海水中原位老化6个月的微塑料纤维对铅(Pb2+)吸附能力增加4%—25%,这是由于老化后微塑料表面覆盖生物膜,生成的大量含氧官能团可以与Pb2+结合形成金属络合物,提高了吸附能力[60]. 一些带电污染物和微塑料之间的吸附主要由静电相互作用主导[85],老化过程增加了微塑料表面的负电荷,这将增强静电相互作用在吸附过程中的重要性[86]. 例如,自然老化的微塑料表面附着大量有机质,增强了微塑料表面负电荷,有利于对重金属阳离子Pb2+的静电吸引[86]. π-π相互作用是含有苯环的微塑料与芳香族化合物相互作用的主要机制,由于链断裂,老化过程可能会削弱π-π相互作用[3]. 例如,紫外照射后的PS聚合物链断裂、苯环脱落,导致π-π相互作用在PS和双酚A(BPA)的结合中减弱,老化PS对BPA的吸附能力下降[87]. 值得注意的是,微塑料吸附污染物的能力取决于不同吸附机制的整体贡献,而老化对不同机制的影响不尽相同,因此对于吸附的总体影响往往比较复杂[24],应进一步研究微塑料的结构、老化过程、老化程度对不同污染物的吸附贡献率,探明老化影响微塑料吸附行为的主控因子.
2.3 环境老化对微塑料载带污染物能力的影响
微塑料具有较强的胶体稳定性和迁移能力,因此,吸附了污染物之后,可作为载体促进污染物在环境中的运移[88-89]. 微塑料对污染物的载带能力主要取决于3个方面,一是微塑料自身的迁移能力,二是微塑料吸附富集污染物的能力,三是污染物在微塑料上发生不可逆吸附或解吸滞后的程度[90-91]. 前面已提到老化可显著影响微塑料的迁移能力和吸附污染物的能力[73, 92]. 与此同时,环境老化所引发的理化性质变化可显著影响微塑料上有机污染物的解吸行为,进而改变微塑料载带污染物迁移的能力. 首先,老化导致微塑料表面微裂纹、微孔体积增多,这一效应增强了非极性有机污染物在微塑料内部孔隙中的“孔填充”效应,不可逆吸附和解吸迟滞程度明显增强,更易于随微塑料迁移[25]. 其次,老化后微塑料表面羟基、羧基等表面含氧官能团的增多,可提升微塑料与极性有机污染物之间的氢键等极性相互作用,也会导致污染物的不可逆吸附程度的增强[25]. 上述两种效应均使得污染物与微塑料结合得更加紧密,更易随微塑料在环境中迁移.
2.4 环境老化对微塑料生物效应的影响
作为一种新兴污染物,微塑料对环境中生物体影响越来越受到重视[93]. 环境老化后微塑料粒径减小,表面覆盖丰富的生物膜,可导致生物体摄入微塑料方式和数量的改变[3]. 塑料在制造过程中掺入了各种添加剂,相比于微塑料本身,添加剂的释放可能对生态安全构成更大的威胁[94],老化促进了微塑料内有毒添加剂的浸出,增强了其毒性效应. 同时,老化显著改变了微塑料富集污染物的能力,随着生物摄食行为,微塑料可将污染物载带至生物体中,促进了污染物的生物摄入.
许多生物体对食物选择有偏好,例如大小,形状等[95],老化后形成的较小的微塑料颗粒甚至是纳米塑料,更容易被生物体(尤其是贻贝和牡蛎幼虫等无脊椎动物)摄入[96]. 不同大小的PS微塑料在斑马鱼体内的摄取和积累位置不同,7 d暴露实验显示,直径为5 μm的PS主要积累在鱼的鳃、肝和肠中,而直径为20 μm的PS颗粒仅积累在鳃和肠道中;积聚在鱼肝中的较小的微塑料颗粒会诱导氧化应激和鱼肝脏代谢组学的改变[97]. 一般情况下,较小的微塑料颗粒更有可能被生物体摄入,并且在组织中分布更广,毒性更大[98]. 生物膜在微塑料上的形成可影响生物体的感知,在较低或较高的营养级,覆盖生物膜的微塑料的生物可利用性均更强,也更容易被生物体摄食[99-100]. 在海水中自然老化3周的微塑料,表面形成的生物膜掩盖了微塑料的惰性,并散发出化学信号使其更类似于食物颗粒,导致浮游动物对微塑料的摄入增加[99].
环境老化促进了微塑料中添加剂、中间体的浸出,增加了微塑料的生态风险[24, 101]. 例如,老化加速了商业微塑料中含有铬和铅的染料的释放,中性条件下在12 h内原始PE中Cr(Ⅵ)和Pb(Ⅱ)的浸出量分别为0.8 μg·g−1和1.7 μg·g−1,紫外老化后微塑料中两者的浸出量分别提高到了3.0 μg·g−1和11.9 μg·g−1. 相比于原始微塑料,老化微塑料的渗滤液更加抑制了铜绿微囊藻(Microcystis aeruginosa)细胞生长和光合作用[101]. 另一研究表明,紫外线照射诱导了PS中的内源性污染物的浸出,原始PS的渗滤液中没有检测出添加剂,紫外线照射60 d后,老化PS渗滤液中检测到了邻苯二甲酸酯类化合物,其积聚在石斑鱼的肝脏脂肪中,造成了生长抑制和肝脏病变[23]. 老化后微塑料可富集更多污染物,增强了其毒性效应. 例如,在溪流中自然老化4周的PE微塑料,表面出现生物膜,对银离子(Ag)吸附量增加了44%;吸附到老化微塑料上的Ag在Steinberg培养基(通常用于培养淡水浮萍)中释放量为247.5 μg·L−1,显著高于原始微塑料(73.5 μg·L−1)[102]. 紫外老化微塑料吸附了更多的罗红霉素,使其在鲫鱼肠道内积累更多,进而导致了更严重的肠道内炎细胞浸润和纤毛受损,抑制了消化酶活性,并扰乱了肠道菌群,造成了代谢紊乱[103].
3. 结论与展望(Conclusions and prospects)
综上所述,微塑料在自然环境中会经历复杂的物理、化学和生物老化过程. 物理老化是机械力作用下微塑料表面裂纹的产生与扩展,从而导致颗粒破碎,有利于化学和生物因素的进一步作用. 化学老化是在紫外线、高温、活性氧等作用下,微塑料化学键断裂进而导致表面化学组成改变的过程. 生物老化是微塑料在生物作用下的破碎同化,微生物也可通过附着机制在其表面形成生物膜. 老化后微塑料的表面形貌、分子结构及化学组成等物理化学性质发生显著改变,不仅影响其自身环境行为,也改变了微塑料与环境介质中的生命和非生命体的相互作用. 因此,老化是评价微塑料环境行为和生物效应中不可忽视的重要因素.为了系统的认识与评价微塑料的环境老化过程和效应机制,亟需在以下几方面开展深入研究:(1)目前微塑料老化的实验研究中,主要是采用单一人工加速老化手段模拟自然老化过程,如机械研磨、紫外辐射、活性氧进攻等. 微塑料在真实环境中的老化,往往受到多种因素共同作用,单一老化手段可能不足以代表真实的环境老化过程. 因此,在实验室模拟老化研究中,更应关注多个老化过程对其的综合影响,缩小实验室模拟手段与环境自然老化过程之间的差距. (2)微塑料性质迥异,环境老化过程复杂,当前更多是定性研究老化机制,应进一步量化微塑料环境老化程度;明确老化微塑料性质变化对其环境行为的贡献,系统研究与开发微塑料不同类型-老化程度-性质变化-环境效应改变之间的关系曲线与预测模型,实现对微塑料的环境风险预测和评估. (3)微塑料与环境大分子如天然有机质、蛋白质、多糖和其他表面活性物质之间的相互作用值得关注,包括探究微塑料与天然有机质不同组分之间发生选择性吸附的作用机制和影响因素,以及微塑料-NOM复合物的环境行为和毒性机制. (4)目前实验室研究对象主要是微塑料颗粒和碎片,应加强对环境中大量检出的微塑料超细纤维和薄膜的老化过程及效应探索.(5)针对微塑料老化所引发的环境行为和效应变化的研究总体集中在光降解老化微塑料,应关注其他老化过程后所引起的效应变化与机制. (6)目前微塑料老化相关研究主要集中在天然水、土壤环境中,工程系统同样是微塑料的重要赋存环境,对于其中的老化过程,目前还鲜有研究. 因此应加强对污水处理厂和垃圾填埋场中微塑料老化的深入研究,例如水处理中的化学老化和固废处理中的热老化,以及自然条件下微塑料在垃圾渗滤液中的原位老化等.
-
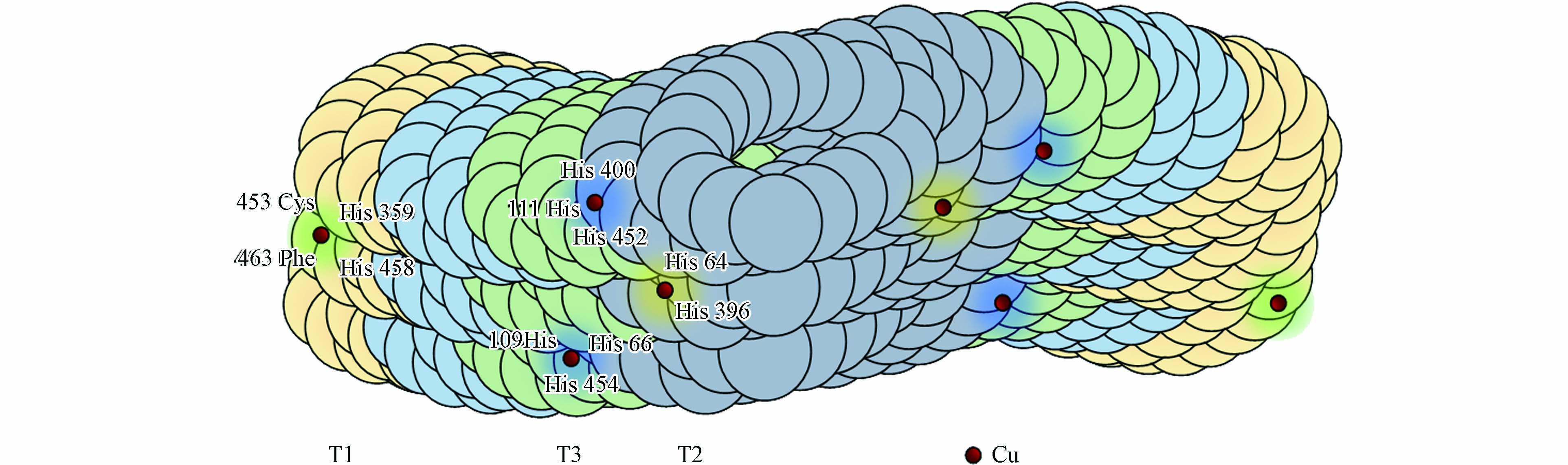
图 1 真菌漆酶的分子结构. T1、T2、T3分别代表漆酶结构中I型、II型、III型Cu2+
Figure 1. The molecular structure of fungal laccase. T1、T2、T3 represented type I, type II, type III Cu2+ of laccase structure, respectively
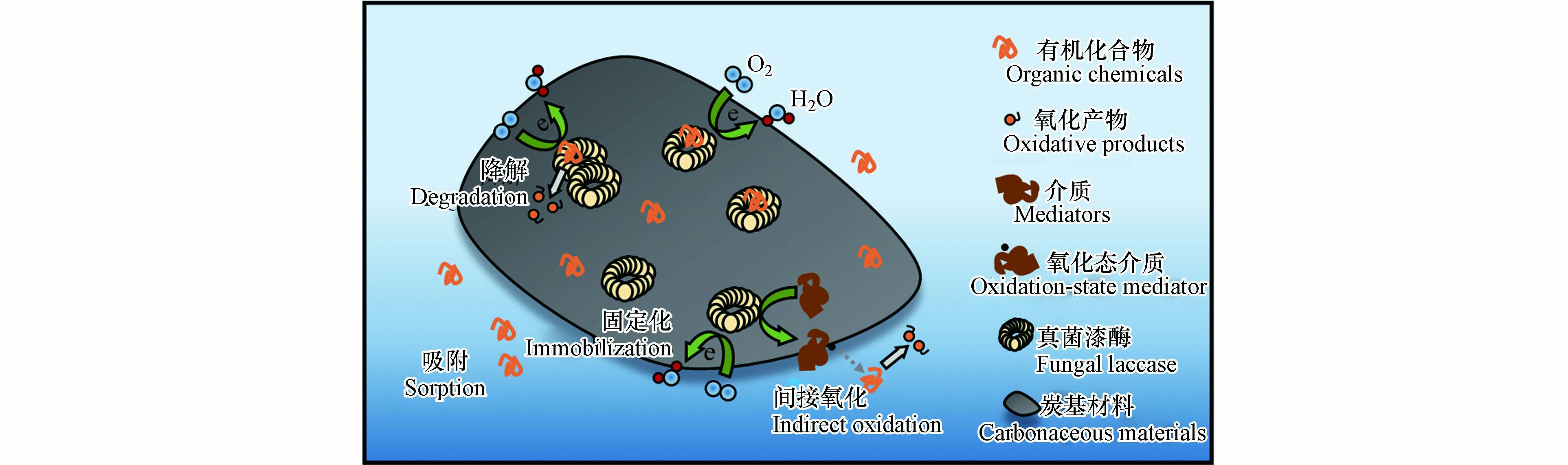
图 3 环境有机污染物在炭基材料固定化漆酶界面上的反应机制
Figure 3. The reaction mechanisms of the environmental organic contaminants on immobilized laccase by carbonaceous materials.
表 1 不同种类的炭基材料固定化漆酶以及其对污染物的去除效率和循环利用率
Table 1. Immobilization of laccase by different types of carbonaceous materials and the efficiency of removal and recycling of organic pollutants
漆酶来源 Source of laccase 炭基材料Carbonaceous materials 炭基原料Feedstock 漆酶固定方法Laccase immobilization methods 固定量 Immobilized amount 目标污染物 Pollutants 去除效率 Removal efficiency 循环活性/去除率Cycle activity 参考文献References Trametes versicolor 石墨烯 石墨烯 物理吸附(非共价自组装) 221. 1 mg·g−1 拉贝洛尔(Labetalol) 100%(初始浓度: 5 µmol·L−1) 10次循环 100%(污染物去除) [35] Bacillus Subtilis 负载Cu2+磁性氧化石墨烯 石墨烯 物理吸附 177 mg·g−1 刚果红(Congo red) 100%(初始浓度: 1 mg·L−1) 10次循环 89. 4%(污染物去除) [39] Trametes versicolor 活性炭 核桃坚果壳 物理吸附 17. 6 mg·g−1 酸性橙7(Acid Orange 7) 100%(初始浓度: 30 mg·L−1) 4次循环 60%(污染物去除) [34] Bacillus Subtilis 磁性活性炭 丝瓜海绵 物理吸附 82. 57 mg·g−1 双酚A(Bisphenol A) 100%(初始浓度: 100 mg·L−1) 5次循环 79. 76%(污染物去除) [40] Trametes versicolor 磁性介孔炭 SBA-15 物理吸附 491. 7 mg·g−1 苯酚(phenol, PHE)对氯苯酚(р-chlorophenol, PNP) PNE: 78%PNP: 84%(初始浓度: 2 mmol·L−1) — [24] Trametes versicolor 空心介孔炭球 四丙氧基硅烷 物理吸附 835 mg·g−1 盐酸四环素(Tetracycline hydrochloride, TCH)盐酸环丙沙星(Ciprofloxacin hydrochloride, CPH) TCH: 高于55%CPH: 接近80%(初始浓度: 20 mg·L−1) 4次循环 70%(酶活) [41] Trametes versicolor 复合纳米纤维膜 多壁碳纳米管 共价结合 907 mg·g−1 双氯芬酸(diclofenac) 100%(初始浓度: 12. 5 mg·L−1) 7次循环 62. 7%(污染物去除) [42] Aspergillus oryzae 功能化多壁碳纳米管 多壁碳纳米管 交联聚合 8620 U·g−1 4-甲氧基苯酚(4-methoxyphenol) 100%(初始浓度: 10 mg·L−1) 3次循环 40%(污染物去除) [43] Trametes hirsuta 纳米复合膜 羟基化多壁碳纳米管 共价结合 30. 40 mg·cm−2 卡马西平(Carbamazepine, CBA)双氯芬酸(diclofenac, DCF) CBZ: 27%DCF: 95%(初始浓度: 5 mg·L−1) 5次循环 22%(酶活) [44] — 介孔二氧化硅负载磁性多壁碳纳米管 多壁碳纳米管 交联聚合 — 铬黑T(Eriochrome Black T, EBT)酸性红88(Acid Red 88, AR 88)活性黑5(Reactive Black 5, RB 5) EBT: 99%AR88: 98%RB5: 66%(初始浓度: 40 mg·L−1) 10次循环 87%(酶活) [45] Trametes versicolor 纳米生物炭 松木 共价结合 5 U·g−1 — — 4次循环 30%(酶活) [46] Trametes versicolor 生物炭(BC) 松木(PW)猪粪(PM)杏仁壳(AS) 交联聚合 BC-PW: 20. 1 U·g−1BC-PM: 40. 2 U·g−1BC-AS: 31. 8 U·g−1 双氯芬酸(Diclofenac) BC-PW: 100%BC-PM: 100%BC-AS: 100%(初始浓度: 0. 5 mg·L−1) BC-PW5次循环46%BC-PM5次循环40%BC-AS5次循环43%(污染物去除) [36] Trametes maxima 功能化生物炭 水稻秸秆 交联聚合 92. 3%(25 mg, 50 U·mL−1) 蒽(Anthracene) 98%(初始浓度: 50 mg·L−1) 3次循环 高于90%(酶活) [37] — 改性生物炭 秸秆 交联聚合 — 2, 4-二氯芬酸(2, 4-diclofenac) 95. 5%(初始浓度: 200 mg·L−1) 6次循环 27. 8%(酶活) [47] 注: “—”的表格代表参考文献中对该内容并未提及.  下载: 导出CSV
下载: 导出CSV
-
[1] YOSHIDA H. LXIII. —chemistry of lacquer (urushi). part I. communication from the chemical society of tokio [J]. J Chem Soc, Trans, 1883, 43: 472-486. doi: 10.1039/CT8834300472 [2] FAURE D, BOUILLANT M, BALLY R. Comparative study of substrates and inhibitors of Azospirillum lipoferum and Pyricularia oryzae laccases [J]. Applied and Environmental Microbiology, 1995, 61(3): 1144-1146. doi: 10.1128/aem.61.3.1144-1146.1995 [3] DWIVEDI U N, SINGH P, PANDEY V P, et al. Structure-function relationship among bacterial, fungal and plant laccases [J]. Journal of Molecular Catalysis B:Enzymatic, 2011, 68(2): 117-128. doi: 10.1016/j.molcatb.2010.11.002 [4] ENDO K, HOSONO K, BEPPU T, et al. A novel extracytoplasmic phenol oxidase of Streptomyces: Its possible involvement in the onset of morphogenesis[J]. Microbiology (Reading, England), 2002, 148(Pt 6): 1767-1776. [5] DARONCH N A, KELBERT M, PEREIRA C S, et al. Elucidating the choice for a precise matrix for laccase immobilization: A review [J]. Chemical Engineering Journal, 2020, 397: 125506. doi: 10.1016/j.cej.2020.125506 [6] LEONOWICZ A, CHO N, LUTEREK J, et al. Fungal laccase: Properties and activity on lignin [J]. Journal of Basic Microbiology, 2001, 41(3/4): 185-227. [7] ZUCCA P, COCCO G, SOLLAI F, et al. Fungal laccases as tools for biodegradation of industrial dyes [J]. Biocatalysis, 2016, 1(1): 82-108. [8] 王苗苗. 介孔二氧化硅磁性复合微球的制备及漆酶固定化[D]. 北京: 北京工业大学, 2012. WANG M M. Mesoporous silica magnetic composite microspheres: Preparation and laccase immobilization[D]. Beijing: Beijing University of Technology, 2012(in Chinese).
[9] 谢柏盛, 贾红华, 谢迎春, 等. 金属框架结构材料MOF-199对漆酶的固定化及其性质 [J]. 生物加工过程, 2011, 9(5): 6-10. doi: 10.3969/j.issn.1672-3678.2011.05.02 XIE B S, JIA H H, XIE Y C, et al. Immobilized laccase and its properties by metal organic framework MOF-199 [J]. Chinese Journal of Bioprocess Engineering, 2011, 9(5): 6-10(in Chinese). doi: 10.3969/j.issn.1672-3678.2011.05.02
[10] 彭凌洁. 壳聚糖基水滑石复合材料合成及漆酶固定化研究[D]. 青岛: 青岛科技大学, 2018. PENG L J. Synthesis of chitosan/layered double hydroxides composite material and immobilization of laccase[D]. Qingdao: Qingdao University of Science & Technology, 2018(in Chinese).
[11] LIU W J, JIANG H, YU H Q. Emerging applications of biochar-based materials for energy storage and conversion [J]. Energy & Environmental Science, 2019, 12(6): 1751-1779. [12] LEE S J, THEERTHAGIRI J, NITHYADHARSENI P, et al. Heteroatom-doped graphene-based materials for sustainable energy applications: A review [J]. Renewable and Sustainable Energy Reviews, 2021, 143: 110849. doi: 10.1016/j.rser.2021.110849 [13] YIN Z F, CUI C J, CHEN H, et al. The application of carbon nanotube/graphene-based nanomaterials in wastewater treatment [J]. Small, 2020, 16(15): 1902301. doi: 10.1002/smll.201902301 [14] JIANG L H, LIU Y G, LIU S B, et al. Adsorption of estrogen contaminants by graphene nanomaterials under natural organic matter preloading: Comparison to carbon nanotube, biochar, and activated carbon [J]. Environmental Science & Technology, 2017, 51(11): 6352-6359. [15] JONES S M, SOLOMON E I. Electron transfer and reaction mechanism of laccases [J]. Cellular and Molecular Life Sciences:CMLS, 2015, 72(5): 869-883. doi: 10.1007/s00018-014-1826-6 [16] CLAUS H. Laccases: structure, reactions, distribution [J]. Micron, 2004, 35(1/2): 93-96. [17] ARESKOGH D, LI J B, NOUSIAINEN P, et al. Oxidative polymerisation of models for phenolic lignin end-groups by laccase [J]. Holzforschung, 2010, 64(1): 21-34. [18] GAVRILAS S, DUMITRU F, STANESCU M D. Commercial laccase oxidation of phenolic compounds [J]. UPB Scientific Bulletin, Series B: Chemistry and Materials Science, 2012, 74: 3-10. [19] FENG Y P, COLOSI L M, GAO S X, et al. Transformation and removal of tetrabromobisphenol A from water in the presence of natural organic matter via laccase-catalyzed reactions: Reaction rates, products, and pathways [J]. Environmental Science & Technology, 2013, 47(2): 1001-1008. [20] 季立才, 胡培植. 漆酶的结构、功能及其应用 [J]. 氨基酸和生物资源, 1996, 18(1): 25-29. doi: 10.14188/j.ajsh.1996.01.010 I L C, HU P Z. Structure, fucntion and application of laccase [J]. Amino Acids & Biotic Resources, 1996, 18(1): 25-29(in Chinese). doi: 10.14188/j.ajsh.1996.01.010
[21] 初华丽, 梁宗琦. 漆酶的潜在应用价值 [J]. 山地农业生物学报, 2004, 23(6): 529-533. doi: 10.15958/j.cnki.sdnyswxb.2004.06.015 CHU H L, LIANG Z Q. New potential application of laccase [J]. Journal of Mountain Agriculture and Biology, 2004, 23(6): 529-533(in Chinese). doi: 10.15958/j.cnki.sdnyswxb.2004.06.015
[22] VIRK A P, PURI M, GUPTA V, et al. Combined enzymatic and physical deinking methodology for efficient eco-friendly recycling of old newsprint [J]. PLoS One, 2013, 8(8): e72346. doi: 10.1371/journal.pone.0072346 [23] COUTO S R. Decolouration of industrial azo dyes by crude laccase from Trametes hirsuta [J]. Journal of Hazardous Materials, 2007, 148(3): 768-770. doi: 10.1016/j.jhazmat.2007.06.123 [24] WANG F, OWUSU-FORDJOUR M, XU L, et al. Immobilization of laccase on magnetic Chelator nanoparticles for apple juice clarification in magnetically stabilized fluidized bed [J]. Frontiers in Bioengineering and Biotechnology, 2020, 8: 589. doi: 10.3389/fbioe.2020.00589 [25] CHAIRIN T, NITHERANONT T, WATANABE A, et al. Biodegradation of bisphenol A and decolorization of synthetic dyes by laccase from white-rot fungus, Trametes polyzona [J]. Applied Biochemistry and Biotechnology, 2013, 169(2): 539-545. doi: 10.1007/s12010-012-9990-3 [26] LIU Q Z, LIU J, HONG D, et al. Fungal laccase-triggered 17β-estradiol humification kinetics and mechanisms in the presence of humic precursors [J]. Journal of Hazardous Materials, 2021, 412: 125197. doi: 10.1016/j.jhazmat.2021.125197 [27] SUN K, KANG F X, WAIGI M G, et al. Laccase-mediated transformation of triclosan in aqueous solution with metal cations and humic acid [J]. Environmental Pollution, 2017, 220: 105-111. doi: 10.1016/j.envpol.2016.09.028 [28] 杨奇. 碳纳米管固定化漆酶吸附降解水中刚果红染料的研究[D]. 广州: 广东工业大学, 2019. YANG Q. Study on the adsorption of Congo red dye in water by carbon nanotubes immobilized laccase[D]. Guangzhou: Guangdong University of Technology, 2019(in Chinese).
[29] KHATAMI S H, VAKILI O, MOVAHEDPOUR A, et al. Laccase: Various types and applications [J]. Biotechnology and Applied Biochemistry, 2022: 1-15. [30] HOMAEI A A, SARIRI R, VIANELLO F, et al. Enzyme immobilization: An update [J]. Journal of Chemical Biology, 2013, 6(4): 185-205. doi: 10.1007/s12154-013-0102-9 [31] GOMES-RUFFI C R, da CUNHA R H, ALMEIDA E L, et al. Effect of the emulsifier sodium stearoyl lactylate and of the enzyme maltogenic amylase on the quality of pan bread during storage [J]. LWT, 2012, 49(1): 96-101. doi: 10.1016/j.lwt.2012.04.014 [32] NJOKU V O, FOO K Y, ASIF M, et al. Preparation of activated carbons from rambutan (Nephelium lappaceum) peel by microwave-induced KOH activation for acid yellow 17 dye adsorption [J]. Chemical Engineering Journal, 2014, 250: 198-204. doi: 10.1016/j.cej.2014.03.115 [33] FANG G D, LIU C, WANG Y J, et al. Photogeneration of reactive oxygen species from biochar suspension for diethyl phthalate degradation [J]. Applied Catalysis B:Environmental, 2017, 214: 34-45. doi: 10.1016/j.apcatb.2017.05.036 [34] RAMÍREZ-MONTOYA L A, HERNÁNDEZ-MONTOYA V, MONTES-MORÁN M A, et al. Correlation between mesopore volume of carbon supports and the immobilization of laccase from Trametes versicolor for the decolorization of Acid Orange 7 [J]. Journal of Environmental Management, 2015, 162: 206-214. [35] DONG S P, JING X P, CAO Y, et al. Non-covalent assembled laccase-graphene composite: Property, stability and performance in beta-blocker removal [J]. Environmental Pollution, 2019, 252: 907-916. doi: 10.1016/j.envpol.2019.05.053 [36] LONAPPAN L, LIU Y X, ROUISSI T, et al. Covalent immobilization of laccase on citric acid functionalized micro-biochars derived from different feedstock and removal of diclofenac [J]. Chemical Engineering Journal, 2018, 351: 985-994. doi: 10.1016/j.cej.2018.06.157 [37] IMAM A, SUMAN S K, SINGH R, et al. Application of laccase immobilized rice straw biochar for anthracene degradation [J]. Environmental Pollution, 2021, 268: 115827. doi: 10.1016/j.envpol.2020.115827 [38] KAUR N, BHARDWAJ P, SINGH G, et al. Applicative insights on nascent role of biochar production, tailoring and immobilization in enzyme industry-A review [J]. Process Biochemistry, 2021, 107: 153-163. doi: 10.1016/j.procbio.2021.05.017 [39] SAMAK N A, TAN Y Q, SUI K Y, et al. CotA laccase immobilized on functionalized magnetic graphene oxide nano-sheets for efficient biocatalysis [J]. Molecular Catalysis, 2018, 445: 269-278. doi: 10.1016/j.mcat.2017.12.004 [40] ZHANG C Y, YOU S P, LIU Y D, et al. Construction of Luffa sponge-based magnetic carbon nanocarriers for laccase immobilization and its application in the removal of bisphenol A [J]. Bioresource Technology, 2020, 305: 123085. doi: 10.1016/j.biortech.2020.123085 [41] SHAO B B, LIU Z F, ZENG G M, et al. Immobilization of laccase on hollow mesoporous carbon nanospheres: Noteworthy immobilization, excellent stability and efficacious for antibiotic contaminants removal [J]. Journal of Hazardous Materials, 2019, 362: 318-326. doi: 10.1016/j.jhazmat.2018.08.069 [42] XU R, TANG R Z, ZHOU Q J, et al. Enhancement of catalytic activity of immobilized laccase for diclofenac biodegradation by carbon nanotubes [J]. Chemical Engineering Journal, 2015, 262: 88-95. doi: 10.1016/j.cej.2014.09.072 [43] COSTA J B, LIMA M J, SAMPAIO M J, et al. Enhanced biocatalytic sustainability of laccase by immobilization on functionalized carbon nanotubes/polysulfone membranes [J]. Chemical Engineering Journal, 2019, 355: 974-985. doi: 10.1016/j.cej.2018.08.178 [44] MASJOUDI M, GOLGOLI M, GHOBADI NEJAD Z, et al. Pharmaceuticals removal by immobilized laccase on polyvinylidene fluoride nanocomposite with multi-walled carbon nanotubes [J]. Chemosphere, 2021, 263: 128043. doi: 10.1016/j.chemosphere.2020.128043 [45] HABIMANA P, GAO J, MWIZERWA J P, et al. Improvement of laccase activity via covalent immobilization over mesoporous silica coated magnetic multiwalled carbon nanotubes for the discoloration of synthetic dyes [J]. ACS Omega, 2021, 6(4): 2777-2789. doi: 10.1021/acsomega.0c05081 [46] NAGHDI M, TAHERAN M, BRAR S K, et al. Pinewood nanobiochar: A unique carrier for the immobilization of crude laccase by covalent bonding [J]. International Journal of Biological Macromolecules, 2018, 115: 563-571. doi: 10.1016/j.ijbiomac.2018.04.105 [47] WANG Z, REN D, WU J, et al. Study on adsorption-degradation of 2, 4-dichlorophenol by modified biochar immobilized laccase [J]. International Journal of Environmental Science and Technology, 2022, 19(3): 1393-1406. doi: 10.1007/s13762-021-03151-2 [48] NAGHDI M, TAHERAN M, BRAR S K, et al. Immobilized laccase on oxygen functionalized nanobiochars through mineral acids treatment for removal of carbamazepine [J]. Science of the Total Environment, 2017, 584/585: 393-401. doi: 10.1016/j.scitotenv.2017.01.021 [49] GENNARI A, FÜHR A J, VOLPATO G, et al. Magnetic cellulose: Versatile support for enzyme immobilization - A review [J]. Carbohydrate Polymers, 2020, 246: 116646. doi: 10.1016/j.carbpol.2020.116646 [50] QIU H J, XU C X, HUANG X R, et al. Immobilization of laccase on nanoporous gold: Comparative studies on the immobilization strategies and the particle size effects [J]. The Journal of Physical Chemistry C, 2009, 113(6): 2521-2525. doi: 10.1021/jp8090304 [51] LIN J H, LIU Y J, CHEN S, et al. Reversible immobilization of laccase onto metal-ion-chelated magnetic microspheres for bisphenol A removal [J]. International Journal of Biological Macromolecules, 2016, 84: 189-199. doi: 10.1016/j.ijbiomac.2015.12.013 [52] LONAPPAN L, LIU Y X, ROUISSI T, et al. Adsorptive immobilization of agro-industrially produced crude laccase on various micro-biochars and degradation of diclofenac [J]. Science of the Total Environment, 2018, 640/641: 1251-1258. doi: 10.1016/j.scitotenv.2018.06.005 [53] SKORONSKI E, SOUZA D H, ELY C, et al. Immobilization of laccase from Aspergillus oryzae on graphene nanosheets [J]. International Journal of Biological Macromolecules, 2017, 99: 121-127. doi: 10.1016/j.ijbiomac.2017.02.076 [54] ASGHER M, NOREEN S, BILAL M. Enhancement of catalytic, reusability, and long-term stability features of Trametes versicolor IBL-04 laccase immobilized on different polymers [J]. International Journal of Biological Macromolecules, 2017, 95: 54-62. doi: 10.1016/j.ijbiomac.2016.11.012 [55] le T T, MURUGESAN K, LEE C S, et al. Degradation of synthetic pollutants in real wastewater using laccase encapsulated in core-shell magnetic copper alginate beads [J]. Bioresource Technology, 2016, 216: 203-210. doi: 10.1016/j.biortech.2016.05.077 [56] QIU X, WANG Y, XUE Y, et al. Laccase immobilized on magnetic nanoparticles modified by amino-functionalized ionic liquid via dialdehyde starch for phenolic compounds biodegradation [J]. Chemical Engineering Journal, 2020, 391: 123564. doi: 10.1016/j.cej.2019.123564 [57] RAHMANI H, LAKZIAN A, KARIMI A, et al. Efficient removal of 2, 4-dinitrophenol from synthetic wastewater and contaminated soil samples using free and immobilized laccases [J]. Journal of Environmental Management, 2020, 256: 109740. doi: 10.1016/j.jenvman.2019.109740 [58] FERNÁNDEZ-FERNÁNDEZ M, SANROMÁN M Á, MOLDES D. Recent developments and applications of immobilized laccase [J]. Biotechnology Advances, 2013, 31(8): 1808-1825. doi: 10.1016/j.biotechadv.2012.02.013 [59] MOHAMAD N R, MARZUKI N H C, BUANG N A, et al. An overview of technologies for immobilization of enzymes and surface analysis techniques for immobilized enzymes [J]. Biotechnology & Biotechnological Equipment, 2015, 29(2): 205-220. [60] YANG J J, MA X X, ZHANG Z S, et al. Lipase immobilized by modification-coupled and adsorption-cross-linking methods: A comparative study [J]. Biotechnology Advances, 2010, 28(5): 644-650. doi: 10.1016/j.biotechadv.2010.05.014 [61] KAH M, SIGMUND G, XIAO F, et al. Sorption of ionizable and ionic organic compounds to biochar, activated carbon and other carbonaceous materials [J]. Water Research, 2017, 124: 673-692. doi: 10.1016/j.watres.2017.07.070 [62] PENG H B, PAN B, WU M, et al. Adsorption of ofloxacin and norfloxacin on carbon nanotubes: Hydrophobicity- and structure-controlled process [J]. Journal of Hazardous Materials, 2012, 233/234: 89-96. doi: 10.1016/j.jhazmat.2012.06.058 [63] CHU G, ZHAO J, HUANG Y, et al. Phosphoric acid pretreatment enhances the specific surface areas of biochars by generation of micropores [J]. Environmental Pollution, 2018, 240: 1-9. doi: 10.1016/j.envpol.2018.04.003 [64] SUN K, HONG D, LIU J, et al. Trametes versicolor laccase-assisted oxidative coupling of estrogens: Conversion kinetics, linking mechanisms, and practical applications in water purification [J]. Science of the Total Environment, 2021, 782: 146917. doi: 10.1016/j.scitotenv.2021.146917 [65] NGUYEN L N, HAI F I, DOSSETO A, et al. Continuous adsorption and biotransformation of micropollutants by granular activated carbon-bound laccase in a packed-bed enzyme reactor [J]. Bioresource Technology, 2016, 210: 108-116. doi: 10.1016/j.biortech.2016.01.014 [66] 曾涵, 龚兰新. 高分子聚合物-多壁碳纳米管复合物固定漆酶及其在玻碳电极上的直接电子转移 [J]. 应用化学, 2012, 29(6): 682-690. ZENG H, GONG L X. Immobilization of laccase on multiwall carbon nanotubes-polymers composites and their direct electron transfer on glassy carbon electrode [J]. Chinese Journal of Applied Chemistry, 2012, 29(6): 682-690(in Chinese).
[67] 陈明雨, 倪烜, 司友斌, 等. 固定化真菌漆酶在环境有机污染修复中的应用研究进展 [J]. 生物技术通报, 2021, 37(6): 244-258. CHEN M Y, NI X, SI Y B, et al. Advances in the application of immobilized fungal laccase for the bioremediation of environmental organic contamination [J]. Biotechnology Bulletin, 2021, 37(6): 244-258(in Chinese).
[68] LIU Y Y, ZENG Z T, ZENG G M, et al. Immobilization of laccase on magnetic bimodal mesoporous carbon and the application in the removal of phenolic compounds [J]. Bioresource Technology, 2012, 115: 21-26. doi: 10.1016/j.biortech.2011.11.015 [69] THIYAGARAJAN P, SELVAM K, SUDHAKAR C, et al. Enhancement of adsorption of magenta dye by immobilized laccase on functionalized biosynthesized activated carbon nanotubes [J]. Water, Air, & Soil Pollution, 2020, 231(7): 1-9. [70] LI N, XIA Q Y, NIU M H, et al. Immobilizing laccase on different species wood biochar to remove the chlorinated biphenyl in wastewater [J]. Scientific Reports, 2018, 8: 13947. doi: 10.1038/s41598-018-32013-0 [71] SATHISHKUMAR P, CHAE J C, UNNITHAN A R, et al. Laccase-poly(lactic-co-glycolic acid) (PLGA) nanofiber: Highly stable, reusable, and efficacious for the transformation of diclofenac [J]. Enzyme and Microbial Technology, 2012, 51(2): 113-118. doi: 10.1016/j.enzmictec.2012.05.001 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 2897
- HTML全文浏览数: 2897
- PDF下载数: 104
- 施引文献: 0
