


 下载:
下载:

-
蓝藻水华暴发是湖泊富营养化的典型特征[1]。蓝藻水华的暴发不仅影响水体自然景观和城市供水,而且有毒蓝藻产生的微囊藻毒素(microcystin,MC)严重危害着水生态系统的安全[2]。能够形成蓝藻水华的藻类包括微囊藻(Microcystis)、鱼腥藻(Anabaena)、颤藻(Oscillatoria)和束丝藻(Aphanizomenon)等,有时直链硅藻(Melosira)也伴随蓝藻大量滋生[3],但微囊藻水华是被公认为发生最多、影响范围最广的蓝藻水华[4]。我国众多湖库均有微囊藻水华暴发的报道[4-7]。
藻类密度是湖库水质和营养型评价中最基本的藻类生物学指标之一[8]。藻类密度是直接反映水体中藻类聚集程度的指标,而浮游藻类的生长受到多种环境因子的制约[9]。特定的水体中,环境因子对藻类的影响作用各不相同。近年来,学者们对不同水体微囊藻密度时空分布及其影响因素开展了大量研究[10-12],但这些研究主要集中在微囊藻水华期间进行,对于微囊藻密度在较长时间尺度下的变化及其与水质因子的关系研究较为少见。
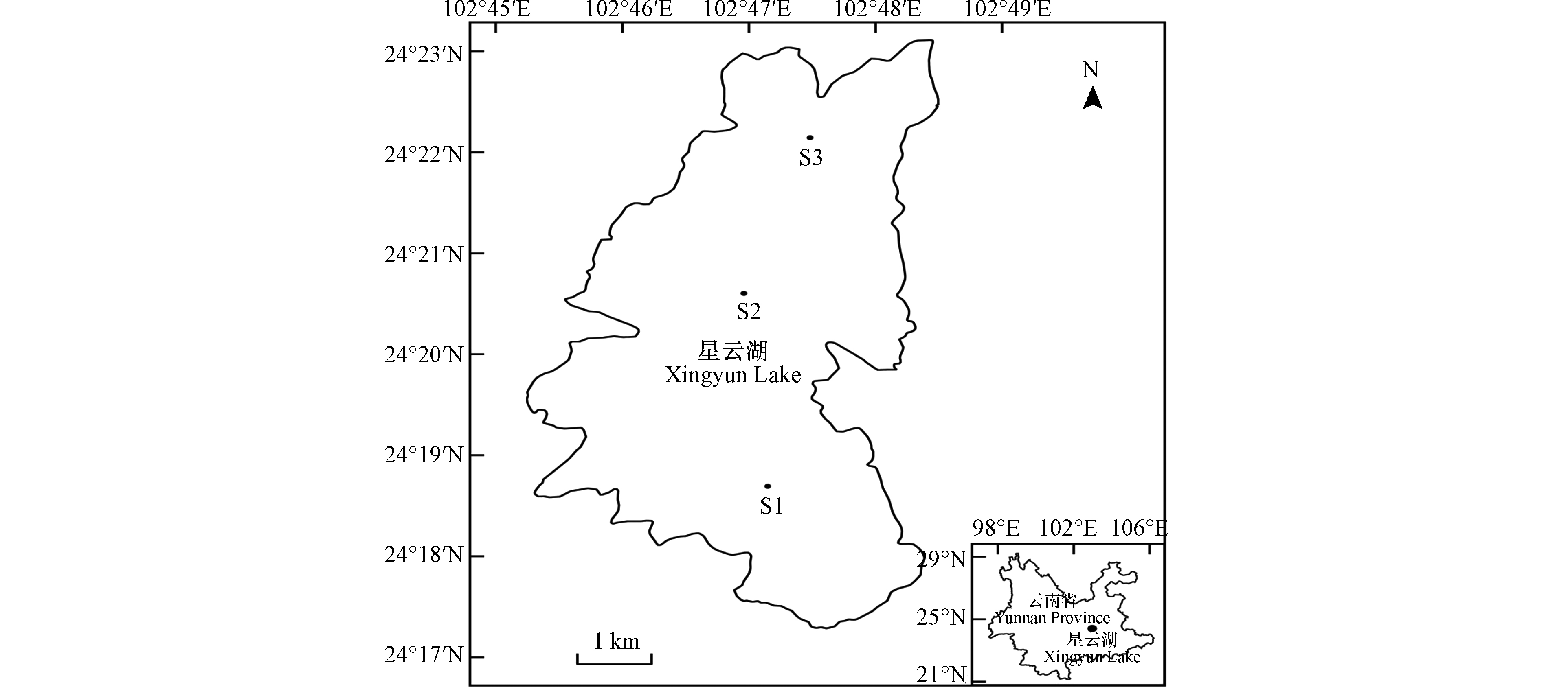
星云湖(24°17′—24°24′N,102°45′—102°49′E)是云南省九大高原湖泊之一,属于珠江流域南盘江水系源头湖泊,水域面积34.7 km2,平均水深7 m[13]。随着流域内社会经济的快速发展,入湖污染负荷总量不断增加,星云湖水体富营养化程度不断加剧。根据玉溪市环境监测站资料,自2002年以来,星云湖以微囊藻为绝对优势类群的蓝藻水华频繁发生,湖泊生态系统日益恶化。因此,有必要对星云湖微囊藻种群数量变化及其机制进行研究。目前,已有学者[14-15]对星云湖浮游藻类群落结构进行了调查,并对微囊藻密度及优势度进行了分析,但以上调查只是针对部分月份进行,并不能全面表征星云湖微囊藻种群数量的动态变化;同时,星云湖微囊藻密度变化与水质指标的相关性研究也尚未见报道。
本研究基于2017年12月至2018年11月星云湖藻类密度及部分水质理化指标监测数据,分析了星云湖微囊藻密度及优势度周年变化特征,同时筛选出了与微囊藻密度变化密切相关的水质指标,旨在为星云湖微囊藻水华的防控及治理提供基础资料。
全文HTML
-
本研究用于分析的数据来源于玉溪市环境监测站2017年12月至2018年11月例行监测数据,具体包括微囊藻密度、藻类总密度、水温(WT)、pH值、电导率(Cond)、氨氮(NH3-N)、总氮(TN)、总磷(TP)、溶解氧(DO)、高锰酸盐指数(CODMn)、五日生化需氧量(BOD5)、化学需氧量(CODCr)和氮磷比(N/P)等13项生物、理化指标。
全湖共设置3个例行监测点位(图1),其中S1位于湖泊南部(省控点),S2位于湖泊中心(国控点),S3位于湖泊北部(省控点)。于每月上旬对各点位浮游藻类及理化指标进行表层(水面下0.5 m)和深层(水面下6 m)采样分析。其中,浮游藻类定量样品为表层和深层混合样(各0.5 L),样品处理参考《水和废水监测分析方法(第四版)》[16]进行,浮游藻类鉴定参考《中国淡水藻类—系统、分类及生态》[17]等资料进行,样品鉴定到属水平。理化指标样品不进行表深层混合,同一点位表深层样品分别测定后取算术平均值代表该点位值。WT采用现场温度计法测定;pH采用玻璃电极法测定;Cond采用实验室电导率仪法测定;NH3-N采用纳氏试剂分光光度法测定;TN采用碱性过硫酸钾消解紫外分光光度法测定;TP采用钼酸铵分光光度法测定;DO采用碘量法测定;CODMn采用酸性高锰酸钾法测定;BOD5采用稀释与接种法测定;CODCr采用重铬酸盐法测定。
-
根据微囊藻密度占比及出现率计算其优势度(Y),公式为:Y=Ni/N × fi,式中,Ni为微囊藻密度,N为藻类总密度,fi为微囊藻的出现率。
微囊藻密度、优势度及水质理化指标取月均值(3个点位算术平均值)进行周年变化分析。将所有数据在Excel2003中建立基础数据库,采用SPSS13.0对微囊藻密度和11项水质指标进行Pearson相关性分析和多元逐步回归分析,筛选出与星云湖微囊藻密度周年变化密切相关的水质指标。相关分析前,经K-S方法检验,所有变量均满足正态分布。
1.1. 数据来源
1.2. 数据处理及分析
-
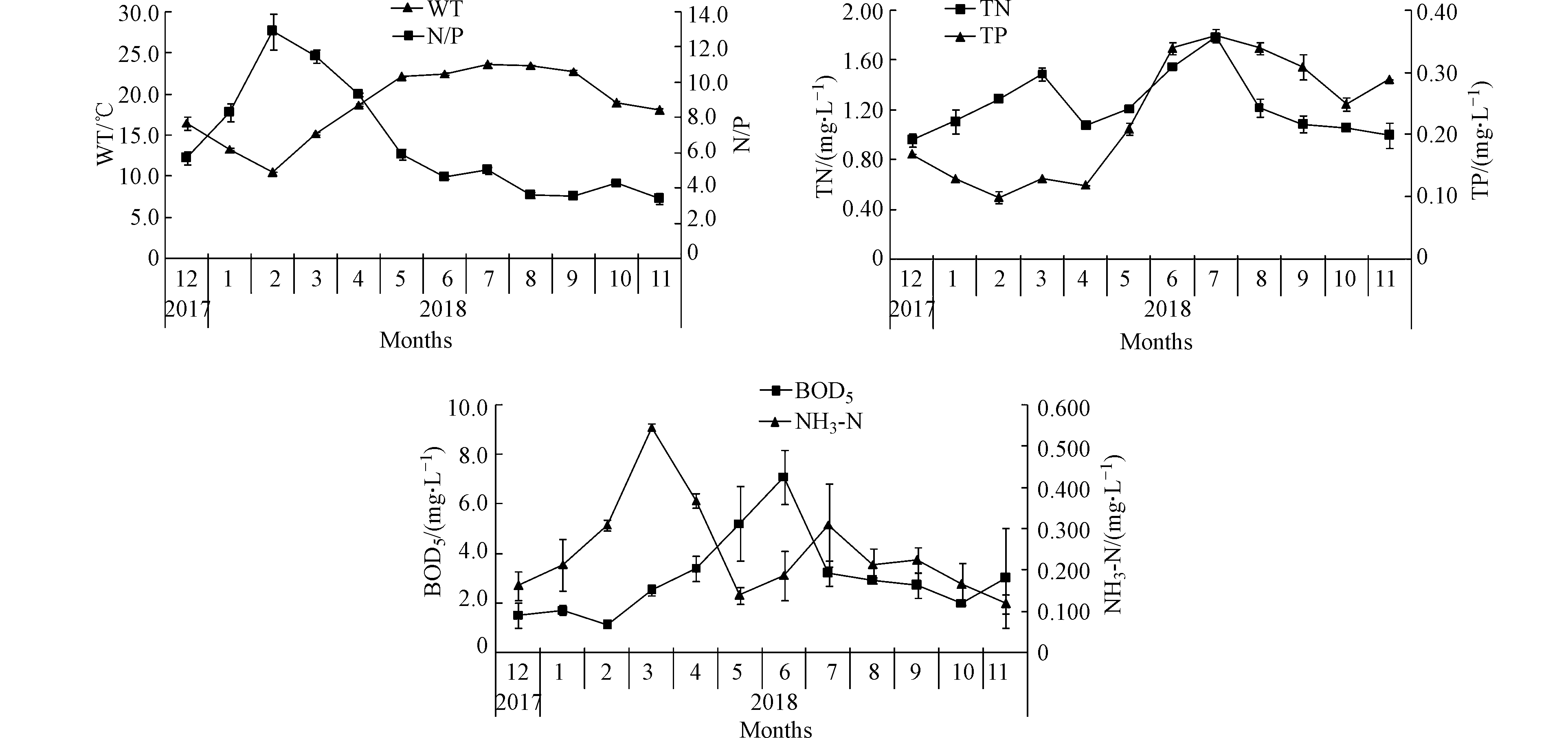
星云湖水质理化指标周年变化情况如表1和图2所示。其中,WT波动范围为10.5—23.6 ℃,年均18.8 ℃,最低出现在2月,最高出现在7月。pH值变化范围8.89—9.40,年均9.16,显示了星云湖水体呈弱碱性,最低出现在3月,最高出现在11月。NH3-N浓度变化范围为0.118—0.546 mg·L−1,年均0.246 mg·L−1,最低出现在11月,最高出现在3月。
TN浓度在0.96—1.79 mg·L−1间变化,年均1.24 mg·L−1,最低为12月,最高为7月。TP浓度在0.10—0.36 mg·L−1间变化,年均0.23 mg·L−1,2月最低,7月最高。N/P变化范围3.4—12.9,年均6.5,11月最低,2月最高。BOD5变化范围1.1—7.1 mg·L−1,年均3.0 mg·L−1,最低出现在2月,最高出现在6月。
-
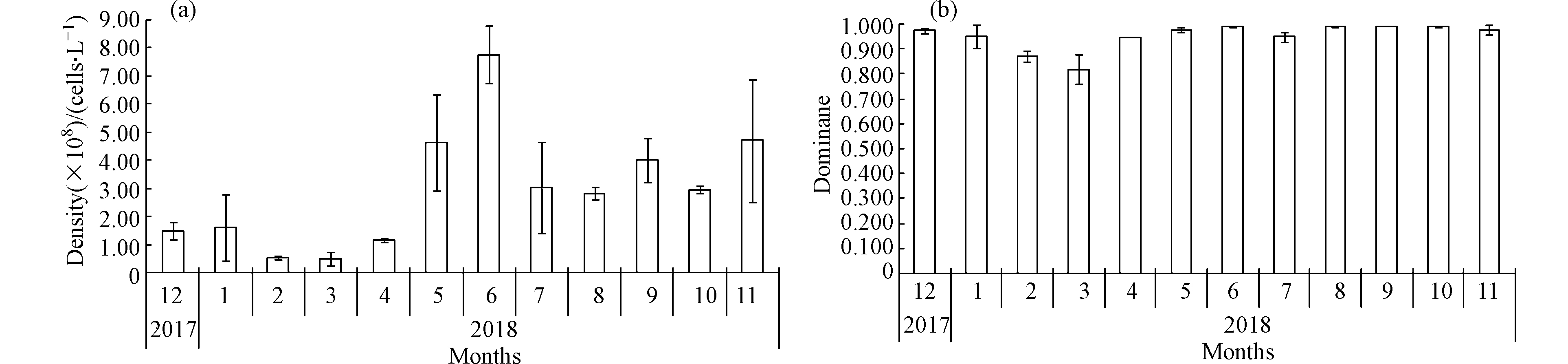
星云湖微囊藻密度周年变化见图3a。结果显示,周年内星云湖微囊藻密度存在明显的月际变化特征,变化范围4.97×107 —7.79×108 cells·L−1,年均2.94×108 cells·L−1。12月、1月微囊藻密度水平基本一致,2月、3月明显降低,3月为全年最低值,4月后密度呈指数增长,6月达最高,7月密度虽有所降低,但仍处于较高值,7—11月间波动不大。孙千千等[18]对太湖梅梁湾和贡湖湾微囊藻周年(2010年7月至2011年6月)变化研究发现,两湾微囊藻平均密度在7月达最高,7—10月间,微囊藻密度虽起伏变化,但总体维持在较高水平,11月后密度大幅降低,次年1—4月水体微囊藻较少,5—6月密度开始逐渐增大。李原等[5]对滇池微囊藻密度年变化(2001年9月至2002年7月)的研究表明,滇池微囊藻密度从9月份开始呈下降趋势,1月份降至最低,而后又逐渐升高,7月份达到最高值。以上表明星云湖与太湖梅梁湾和贡湖湾、滇池水体中微囊藻密度周年变化特征有所相似。从季节上看,夏季(6—8月)和秋季(9—11月)微囊藻密度平均值相对较高,分别达4.55×108 cells·L−1和3.90×108 cells·L−1;春季(3—5月)次之,为2.09×108 cells·L−1;冬季(12—2月)最低,为1.21×108 cells·L−1。表明星云湖微囊藻密度具有明显的季节性变化特征,这种变化趋势与微囊藻水华发生规律较为吻合,夏秋季为星云湖微囊藻水华暴发期,细胞密度较高,而冬春季水华发生频次和规模相对较小,因此微囊藻密度水平也相对较低;星云湖微囊藻水华一年四季均有出现,相同的情况在滇池[5]和太湖[19]也有发生。
周年内星云湖微囊藻密度占藻类总密度比例极高,其优势度高达0.821—0.995(图3b)。微囊藻属是星云湖各月份浮游藻类组成的绝对优势类群,其密度水平决定了星云湖浮游藻类密度的总水平。星云湖浮游藻类优势种群无季节演替,这与Sommer等[20]提出的PEG模型中,浮游藻类冬春季以硅藻和隐藻占优,夏季以绿藻占优,夏末秋初则以蓝藻占优,秋季硅藻再次占优的季节演替规律完全不符,分析认为这可能与湖泊营养状态有关。同时,对比前人研究还发现,星云湖微囊藻密度及优势度水平远高于太湖[21]和滇池[5, 22]等富营养型湖泊。根据况琪军等[8]湖泊营养类型评价中藻类密度指标标准,仅从微囊藻密度加以评价,周年内星云湖除2月、3月水体营养状态为中富营养型外,其余10个月份均为极富营养型,表明星云湖水体受污染程度尤为严重。
本研究中,周年内星云湖微囊藻密度及优势度水平均处于较高值,分析认为引起这种现象的原因可能是多方面的:一是湖泊内外源污染严重,湖滨带生态功能退化。罗玉等[23]研究表明,近20年来,星云湖水体中N、P污染指标呈明显上升趋势,这与流域内面源污染负荷的产生量和入湖量显著增加相关,“十五”、“十一五”和“十二五”期间,星云湖流域TP污染负荷年均产生量分别为382.20 、589.87 、346.80 t,TN污染负荷年均产生量分别为2694.90、3292.44 、1827.11 t。自2000年至今,星云湖湿地建设工作虽持续开展,但湖滨带的建设总体较为分散零星,且缺乏后续的管护资金投入,导致已建成的河口湿地物种组成单一,生物多样性受损,湖滨带生态功能退化[24],失去了应有的缓冲、拦截作用。同时,出流改道后星云湖自西南角出水,微囊藻吸收了水体中营养盐后随常年盛行的西南风逆吞吐水流移动,不利于营养盐随水流输出,导致营养盐在湖体中形成富集机制[15],又一次为微囊藻的生长提供了营养物质。二是沉水植物退化消亡严重。沉水植物是维护水生态系统多样性的基础,能够消耗水中大量的氮、磷等物质,对有毒藻类的生长具有明显的抑制作用[25]。研究表明,星云湖沉水植物分布面积和分布深度分别由1984年的7.85 km2和4.2 m以内缩减至2000年的0.72 km2和2.5 m以内[26];而2008年星云湖沉水植物分布面积约为0.6 km2,盖度约1.8%[14]。三是大型浮游动物种群数量减少。施海彬等[27]研究表明,在太湖新银鱼捕食压力增强的背景下,星云湖桡足类与枝角类等浮游动物的生物量受到了捕食压力的抑制作用,但桡足类下降更为明显;2005年以来,星云湖象鼻溞(Bosmina)个体大小出现明显波动并呈总体下降趋势,同时象鼻溞与Daphnia longispina group生物量总体下降。由此,来自浮游动物对微囊藻的捕食压力也随之减小。四是微囊藻自身生存特点。群体微囊藻对营养盐、温度和DO等具有较广的适应范围[28]。微囊藻细胞聚集成较大群体后,可以有效抵御浮游动物的捕食,在一定程度上促使小型单细胞绿藻、硅藻等生物量的降低,使得大型或群体的藻类占优势[21]。同时,微囊藻在自然界中具有很多竞争优势,如可以通过伪空胞调节自身浮力,以接受更多的光照;通过他感作用影响其他藻类的生长;其数量较大并趋向于水面分布时,还可以通过遮光效应导致其他种类的减少[29]。综合以上分析认为,星云湖多年的污染加剧及治理不善,使水体营养盐等赋存量保持在较高水平,为藻类的生长繁殖提供了丰厚的物质基础,随着沉水植物及大型浮游动物的不断减少,加之微囊藻自身所具备的生存优势等一系列原因,导致了微囊藻成为周年内星云湖浮游藻类的绝对优势类群,并具有较高的密度水平。星云湖由草型湖泊转变为典型的藻型湖泊,标志着星云湖水生态系统功能的严重退化。
-
对星云湖各月份微囊藻密度与11项理化指标进行了Pearson相关分析(表2)。由于微囊藻具伪空胞,易自由漂浮,加之其易受风浪和湖流等影响而发生迁移,因此微囊藻密度的空间分布可能受自身生理特性以及采样期间风向风速、水体流场、水体营养状态等多种因素的共同影响。
本研究旨在时间尺度下分析星云湖微囊藻密度变化特征及其与水质指标的关系,为避免其它因素对分析结果带来干扰,故微囊藻密度及水质指标取月均值进行分析。分析数据显示,微囊藻密度与TP和BOD5呈极显著正相关(P<0.01),与WT和Cond呈显著正相关(P<0.05),与NH3-N和N/P呈显著负相关(P<0.05)。
-
根据相关性分析结果,以TP、BOD5、WT、Cond、NH3-N和N/P等 6项与微囊藻密度具有显著相关的水质指标为自变量,微囊藻密度为因变量,通过多元逐步回归分析进一步筛选与星云湖微囊藻密度周年变化关系最为密切的水质指标,建立最优的多元线性回归方程,并对其进行显著性检验(表3)。由表3可知,入选星云湖微囊藻密度回归方程的水质指标有TP、BOD5、N/P和WT,R值、F值和P值分别为0.981、45.428和0.000,显示了回归方程极显著,表明分析结果可靠。因此,TP、BOD5、N/P和WT为与星云湖微囊藻密度周年变化密切相关的水质指标。
WT是影响藻类生长的关键物理因子[30]。WT能够促进藻类生命活性,使氮、磷等营养盐在水体中的迁移转化过程加快,利于藻细胞对营养物质的吸收,藻类生理活性升高和生物量高峰值增加,从而使藻类细胞完成自身成长增殖[31]。WT的增加能促进微囊藻的生长速率[32]。太湖梅梁湾微囊藻生物量与WT密切相关,随着太湖WT的增加,微囊藻生物量显著增加,但并非简单的线性相关[33]。本研究中,星云湖微囊藻密度周年变化与WT呈显著正相关,冬春转换期(2、3月)星云湖WT较低,微囊藻细胞新陈代谢较慢,增殖较慢,密度为全年最低;而春末(5月)后随着WT上升,微囊藻细胞代谢活动不断增强,生长繁殖较快,密度也处于相对较高水平。表明WT是星云湖微囊藻密度变化的原因之一。
磷是影响微囊藻生长的主要营养元素之一[34]。微囊藻具有较高的磷摄取速率和较强的磷储存能力,对氮磷等营养盐的结合能力也比其它藻类强,这些特点使其可以更有效地利用磷[35]。大量研究表明,TP对微囊藻生长影响较大,微囊藻丰度或生物量与TP浓度呈显著正相关[10, 36-37],但过高的磷浓度也不利于微囊藻生长[38]。同时,微囊藻的大量繁殖可使水体pH增高,降低下层或底层水体DO水平,释放促发颗粒态磷转化释放的磷酸酶等作用机制,从而促进底泥中内源性磷的释放,增加水相颗粒磷库[39]。BOD5与水体有机污染状况有关,常作为衡量湖库水体有机物含量的重要指标,指数越大,表明水体受有机污染越严重。水体中有机物会促进藻类的生长繁殖,同时藻类自身会通过光合作用产生大量的有机物,使水体中有机物升高[40]。沙湖浮游藻类密度与BOD5呈显著正相关[41]。本研究中,星云湖微囊藻密度周年变化与TP和BOD5呈极显著正相关。一方面,星云湖TP和有机物的入湖污染途径主要是入湖河流,而星云湖主要的入湖河流多为季节性河流,冬春季(12—3月)为流域内相对农闲及枯水期,入湖的TP和有机物相对较少,微囊藻密度也相对较低;春末后(5—11月)随着当地大春栽种、农田泄水、雨季来临、小春栽种等一系列原因,入湖污染负荷不断增大,水体中TP和BOD5浓度呈增加趋势,加之WT的升高对营养盐及有机物的转化加快,利于微囊藻细胞对营养物质的吸收利用,微囊藻密度较冬春季节明显增加,水体TP和有机物浓度成为了星云湖微囊藻大量生长的原因。另一方面,周年内星云湖较高的微囊藻密度,使其水体pH也维持在相对较高水平(8.89—9.40),内源性磷的释放导致水体TP浓度也较高;同时,微囊藻在生长代谢和消亡过程中释放大量的有机物,使水体BOD5浓度也随之增加,由此,水体TP及BOD5浓度的增加又成为了微囊藻大量生长繁殖的结果。因此,星云湖微囊藻密度与TP和BOD5浓度变化是互为因果的。
Rhee等[42]发现N/P低于10时有利于微囊藻成为优势种。我国绝大部分湖泊暴发的水华都是蓝藻水华,原因是我国绝大部分湖泊中N/P都比较低,使得没有固氮能力的微囊藻成为优势种[3]。代龚圆等[43]研究认为,较低的N/P有利于滇池微囊藻生物量的增加。Liu等[44]研究发现,太湖微囊藻生物量与N/P呈负相关。在较高的N/P情况下,水体中也会形成蓝藻水华,较低的N/P并不是蓝藻水华形成的条件,而是蓝藻水华产生的结果[45]。夏季太湖梅梁湾蓝藻水华比湖心区更为严重,导致无机氮浓度降低,无机磷浓度升高,进而使N/P降低,低N/P是蓝藻水华发生的结果[46]。本研究中,星云湖水体周年内N/P变化范围3.4—12.9,全年均以微囊藻为绝对优势类群,微囊藻密度变化与N/P呈极显著负相关。这一方面符合Smith[47]提出的低N/P学说;但从另一方面看,星云湖绝对优势类群微囊藻自身并无固氮能力,其大量繁殖必定要消耗水体中大量的氮,同时又促进底泥中内源性磷的释放,最终导致N/P降低。因此,星云湖微囊藻密度与N/P变化的因果关系还有待进一步研究来解释。
2.1. 水质指标周年变化
2.2. 微囊藻密度及优势度周年变化
2.3. 微囊藻密度周年变化与水质指标的关系
2.3.1. 相关性分析
2.3.2. 多元逐步回归分析
-
(1)微囊藻是星云湖浮游藻类组成的绝对优势类群,周年内优势度高达0.821—0.995。星云湖微囊藻密度月变化明显,变化范围4.97×107 —7.79×108 cells·L−1,年均2.94×108 cells·L−1,最大值出现在2018年6月,最小值出现在2018年3月。微囊藻密度夏秋季相对较高,春季次之,冬季最低。
(2)Pearson相关性分析和多元逐步回归分析认为,WT、TP、BOD5和N/P是与星云湖微囊藻密度周年变化密切相关的4项水质指标。
(3)周年内星云湖微囊藻密度及优势度均维持在较高水平,微囊藻水华全年均可暴发。相关管理部门应高度重视,积极采取外源与内源治理并举的措施,最大程度削减湖泊及入湖营养盐输入,尤其是磷输入,逐步减少水华现象的发生,实现湖泊资源的可持续发展和利用。
