



-
城镇污水处理厂生化尾水深度净化脱氮常采用异养反硝化工艺,安全廉价的碳源是该工艺高效运行的关键。与常用液态碳源相比,可生物降解型人工合成聚合物固体碳源因运输安全、释碳稳定而成为反硝化脱氮领域的研究热点。聚乳酸(Polylactic acid PLA)、3-羟基丁酸/戊酸酯共聚物(Polyhydroxy-butyrate-co-valerate PHBV)[1]、聚己内酯(Poly(ε-caprolactone) PCL)[2]和聚丁二酸丁二醇酯(Poly(Butylene Succinate) PBS)[3]等聚合物具有良好释碳性能。但PHBV,PCL和PBS等聚合物在合成过程中存在聚合条件苛刻、催化剂毒性高等问题[4-5],存在安全隐患。PLA能够由乳酸直接缩聚而成,合成方法简单、价格低廉,被认为是最具有发展潜力的可生物降解聚合物[6-7]。
目前,关于PLA作为固体碳源进行反硝化脱氮的研究多以工程应用为主。FAN等[8]研究了PLA反硝化脱氮的污泥培养时间(35 d)与适宜温度(30~40 ℃)。彭书林等[9]优化了PLA反硝化系统的适宜进水pH范围为7.8~10.1。TAKAHASHI等[10]从PLA水解产物角度判断了不同分子量PLA的脱氮效果,指出低分子量PLA脱氮效果更好。但是不同分子量PLA的脱氮路径及相应机制尚不清晰,这是异养微生物脱氮过程高效调控的理论基础。
本研究选取分子量为5 000 g·mol−1,10 000 g·mol−1和50 000 g·mol−1的PLA,在探究其释碳性能和反硝化效果的基础上,通过分析出水溶解性有机质(DOM)组分及微生物群落结构,揭示不同分子量PLA反硝化脱氮路径与机制,以期为PLA应用于低碳脱氮提供参考。
-
本实验选用直接聚合法[11]合成的PLA 。该方法流程简单成本较低,合成的PLA分子量在5 000~50 000 g·mol−1 [12-13]。因此,本实验选取分子量为5 000 g·mol−1,10 000 g·mol−1,50 000 g·mol−1的PLA进行研究。PLA的形态为白色椭球形颗粒,其直径为2~3 mm、长为3~4 mm。其他试剂均采用分析纯。模拟废水的制备方法:在自来水中加入NaNO3和KH2PO4,使NO3−-N质量浓度为30 mg·L−1,PO43−-P质量浓度为6 mg·L−1;微量元素溶液按1 mL·L−1体积分数添加,其组分如表1所示[9]。
-
1)聚乳酸PLA静态释碳研究方法。取3只500 mL锥形瓶,分别加入300 mL去离子水,并加入100 g分子量为5 000 g·mol−1、10 000 g·mol−1、50 000 g·mol−1的PLA,瓶口密封,放置于35 ℃、80 r·min−1恒温培养箱中。每24 h取样并更换去离子水,连续更换17 d[14]。之后,测定水样COD,并计算PLA的累计释碳率(cumulative carbon release rate)。
2) 聚乳酸PLA反硝化研究方法。取3只500 mL锥形瓶,加入300 mL模拟废水和150 mL活性污泥(MLSS为3 g·L−1),分别加入100 g分子量为5 000 g·mol−1、10 000 g·mol−1、50 000 g·mol−1的PLA,瓶口密封,放置于35 ℃、80 r·min−1恒温培养箱中。每24 h取样并更换300 mL模拟废水,连续更换36 d,并于第1天、第17天和第37天取样分析DOM组分,于第37天取样分析微生物群落结构。测定水样NO3−-N、NO2−-N、NH4+-N、COD,并计算反硝化速率(Denitrification rate)。
3) PLA累计释碳率计算。PLA累计释碳率可用CR来表示,计算如式(1)所示。
式中:CRd为第d天的累计释碳率;Ci为第i天的COD,mg·L−1;V为实验溶液体积,L;TC为释碳实验所使用PLA总含碳量,mg。
4) PLA反硝化速率计算。反硝化速率可以用DR表示,如式(2)[2]所示。
式中:DR为反硝化速率,mg·L−1·h−1;C0和Ce分别为NO3−-N初始浓度和出水质量浓度,mg·L−1。本研究中水力停留时间(HRT)统一为24 h。
-
水样经0.45 μm膜滤膜过滤后测定相关参数。其中,NO3−-N采用紫外分光光度法测定;NO2−-N采用N-(1-萘基)-乙二胺光度法测定;NH4+-N采用纳氏试剂法测定;COD采用快速消解分光光度法(DR-1900,哈希,美国)测定。三维荧光光谱采用日立(Hitachi)F-7000荧光分光度计测定,激发波长(λex)扫描范围:200~450 nm,发射波长(λem)扫描范围:250~600 nm;狭缝宽带:λex=5 nm,λem=5 nm;扫描速度2 400 nm·min−1。三维荧光光谱(3D-EEM)根据激发波长和发射波长的不同范围分为5个区域。这5个区域分别为Ⅰ区(λex/λem= (200~250 nm) / (250~330 nm) ,类酪氨酸物质)、Ⅱ区(λex/λem= (200~250 nm) / (330~380 nm) ,类色氨酸物质)、Ⅲ区(λex/λem= (200~250 nm) / (380~550 nm) ,类富里酸物质)、Ⅳ区(λex/λem= (250~400 nm) / (250~380 nm) ,溶解性微生物代谢产物物质)、Ⅴ区(λex/λem= (250~400 nm) / (380~550 nm) ,类腐殖酸物质)[15]。按照荧光区域积分 (FRI) 分析法,通过Origin9.1计算荧光区域的积分体积Фi;对荧光区域的积分体积进行标准化,得到区域i的标准化积分体积Фi,n;再计算总荧光区域标准化积分体积ФT,n;最后计算区域i标准化积分体积占总标准化积分体积之比Pi,n[16]。
微生物群落结构采用16SrRNA高通量测序进行分析。污泥样品采集后置于10 mL离心管内,-20 ℃保存,随后进行样品DNA提取,采用通用引物338F ( ACTCCTACGGGAGGCAGCAG ) 和806R ( GGACTACHVGGGTWTCTAAT ) 对样品 16S rRNA基因V3-V4 可变区进行 PCR 扩增。PCR产物用QuantiFluor™ -ST定量系统(美国Promega公司)检测定量后,最后利用Illumina Miseq PE300平台进行测序。使用Uparse软件 (version 11) ,根据97%的相似度对序列进行OTU聚类。利用RDP classifier (version 2.13) 对每条序列进行物种分类注释,比对Silva 16S rRNA数据库 (version 138) ,设置比对阈值为70%,并在门、科和属水平上统计不同样本的群落组成。
-
分子量为5 000 g·mol−1、10 000 g·mol−1和50 000 g·mol−1的PLA静态释碳性能如图1所示。PLA的分子量增大,其释碳稳定性越好。分子量为5 000 g·mol−1、10 000 g·mol−1的PLA释碳表现出明显的两阶段性:第1天为快速释碳阶段,COD分别降为40 mg·L−1和33 mg·L−1;第2~17天为缓慢释碳阶段,每天稳定释放COD约20 mg·L−1。分子量50 000 g·mol−1的PLA未见明显的阶段性,而周期内COD释放量每天保持约20 mg·L−1。不同分子量PLA在17 d内累计释放率稳定增长,且不超过0.1%。
性能良好的固体碳源在清水中释碳能力较弱,可在微生物作用下释放反硝化脱氮所需碳源[17]。上述数据表明,不同分子量的PLA均可长时间进行低量释放碳源,为一种性能良好的固体碳源。这归因于PLA的水溶性差,在清水中可长时间低量释碳(约为20 mg·L−1),与PCL、PHBV等人工合成碳源释碳量相近(13.35~26.70 mg·L−1)[18-19],远低于玉米芯和花生壳等天然碳源(200~300 mg·L−1)[20];而固体碳源静态释碳过高,应用脱氮系统中会造成出水有机物含量超标。
-
当PLA分子量分别为5 000 g·mol−1、10 000 g·mol−1和50 000 g·mol−1时,出水NO3−-N质量浓度变化、去除率和反硝化速率如图2所示,副产物NH4+-N质量浓度和不同形态N比例变化如图3。
随着PLA分子量的增加,出水NO3−-N质量浓度增大,去除率和反硝化速率降低。第1~16天,不同分子量PLA的出水NO3−-N质量浓度均随时间逐渐下降,去除率和反硝化速率逐渐升高;第17~36天,不同分子量PLA出水NO3−-N质量浓度、去除率和反硝化速率均呈稳定趋势,5 000 g·mol−1分子量PLA的NO3−-N去除率和反硝化速率可分别达到100%和1.29 mg·L−1·h−1。分析其原因,在反硝化过程中,微生物无法直接利用PLA颗粒,而是以PLA的水解产物作为碳源[10]。PLA会在微生物分泌的胞外水解酶作用下,发生内部酯键水解而形成小分子有机酸[21-22] ,胞外水解酶会优先催化水解PLA的非晶区[7, 21-22]。PLA水解出小分子有机物的能力也与其分子量密切相关。低分子量PLA主要以非晶区存在更容易被胞外酶催化水解提供有效碳源[23-24],故其反硝化效果更好。
由图3 (a) 可知,在反应过程中,PLA分子量分别为10 000 g·mol−1、50 000 g·mol−1时,NH4+-N无明显积累(<0.5 mg·L−1)。PLA分子量为5 000 g·mol−1时,反应1 d时出水NH4+-N质量浓度为2.27 mg·L−1。这可能是由于PLA分子量为5 000 g·mol−1时,反应器的接种污泥释放了在污水处理厂吸附的铵[25];而在后续实验阶段,NH4+-N稍有积累,最高质量浓度为0.83 mg·L−1,这可能是由于反应器内发生了硝酸盐异化还原为铵(dissimilatory nitrate reduction to ammonium,DNRA)的反应[26]。PLA分子量为5 000 g·mol−1时,NH4+-N积累量大于PLA分子量为10 000 g·mol−1和50 000 g·mol−1时。这是因为PLA分子量为5 000 g·mol−1时,能释放更多碳源并加强DNRA作用[27]。DNRA作用常见于固体碳源脱氮过程,但NH4+-N积累量会随不同种类碳源变化。一般情况下,天然固体碳源的效果会优于聚合物类固体碳源。而本实验结果表明,PLA为固体碳源时,其出水NH4+-N积累量(<0.83 mg·L−1)会低于PHBV(1.14 mg·L−1)等其他聚合物碳源[28-29]。不同分子量PLA反硝化实验出水NO2−-N无明显积累 (<0.2 mg·L−1) 。异养反硝化反应以有机物为电子供体,分为NO3−-N被还原为NO2−-N与NO2−-N被还原为N2两步,分别由硝酸盐还原酶和亚硝酸盐还原酶催化进行。出水NO2−-N无明显积累表明以PLA为固体碳源的反硝化反应的限速步骤为NO3−-N被还原为NO2−-N过程,而NO2−-N能迅速被亚硝酸盐还原酶还原为N2。
由图3 (b) 可知,稳定期(第17~36天)不同分子量PLA反硝化氮气转化率均高于驯化期(第1~16天),5 000 g·mol−1分子量PLA稳定期氮气转化率达到99%,且不同分子量PLA反硝化出水NO2−-N无明显积累。这表明5 000 g·mol−1分子量的PLA能实现完全反硝化,几乎不生成NO2−-N和NH4+-N等副产物。
-
1) PLA反硝化电子供体有机质来源。分子量分别为5 000 g·mol−1、10 000 g·mol−1和50 000 g·mol−1的PLA对应反硝化出水COD的变化如图4所示,溶解性有机质(DOM)组分变化如图5、6和7所示。
由图4可知,分子量为5 000 g·mol−1、10 000 g·mol−1和50 000 g·mol−1的PLA对应的反硝化实验出水COD较稳定,平均COD分别为16.26 mg·L−1、15.35 mg·L−1和14.17 mg·L−1。固体碳源反硝化系统内COD主要取决于微生物作用下碳源释放过程与异养反硝化等碳源利用过程[30]。图4中出水COD稳定表明脱氮系统内碳源的释放与利用达到平衡,即在脱氮过程中微生物水解PLA释放的碳源会被微生物菌群迅速用于反硝化脱氮及自身生长繁殖。
根据三维荧光光谱(3D-EEM)的特定波长荧光强度数据,能显示水中溶解性有机质(DOM)的组分特征。由三维荧光光谱 (图5) 可知,不同分子量PLA反硝化出水中DOM组分主要为溶解性微生物代谢产物、腐殖酸类、富里酸类及少量蛋白质类物质。为进一步探究DOM的荧光特性,运用荧光光谱FRI对DOM的三维荧光光谱进行定量分析。各样品的总积分标准体积见图6 (a) 。随着实验的进行,反应器中DOM的荧光强度逐渐降低。这说明PLA异养反硝化能去除水中DOM。各荧光组分占比见图6 (b) 。随着反硝化时间的延长,反应器内溶解性微生物代谢产物和类蛋白质物质组分占比降低,而腐殖酸类和富里酸类物质组分占比升高。这是因为溶解性微生物代谢产物等易降解有机物在反硝化过程中会逐渐转化为腐殖酸类和富里酸类等难降解有机物[31],A2O工艺的缺氧池中也存在以微生物代谢产物为电子供体的反硝化过程,同时产生腐殖酸[32]。因此,PLA反硝化脱氮电子供体来源不仅有PLA释放的小分子碳源,还有溶解性微生物代谢产物等有机物。
2) PLA脱氮微生物学机制分析。分子量分别为5 000 g·mol−1、10 000 g·mol−1和50 000 g·mol−1的PLA对应的反硝化脱氮系统微生物群落多样性分析结果:α多样性指数如表2所示,微生物门、科和属水平群落结构如图7和8所示。
表2表明,所有样品覆盖率(coverage)均为0.99,代表测序结果真实可靠。反应器中Shannon和Chao指数基本无变化并保持在较高数值,证明PLA分子量对微生物群落丰富度和多样性影响不大,且PLA反应器内微生物活性较高,同时也表明PLA可作为反硝化固体碳源。
不同污泥样品在门水平上的分布如图7 (a) 所示,主要优势门主要有变形菌门(Proteobacteria)(32.28%~35.12%)、绿弯菌门(Chloroflexi)(25.67%~36.00%)、放线菌门(Actinobacteriota)(5.11%~6.82%)、CPR细菌(Patescibacteria)(3.92%~8.14%)和拟杆菌门(Bacteroidota)(5.47%~6.27%)。常见的还原硝酸盐的反硝化菌属大多在变形菌门(Proteobacteria)和拟杆菌门(Bacteroidota)下[33-34],不同分子量PLA反应器内变形菌门(Proteobacteria)与拟杆菌门(Bacteroidota)相对丰度之和为37.75%~41.38%。这表明各反应器内这2种菌门为优势菌门。绿弯菌门(Chloroflexi)下的微生物能在厌氧条件下吸收来自细胞裂解的肽聚糖等物质,其部分属能利用细胞释放的微生物代谢产物作为电子供体进行反硝化作用[35],S3样品内绿弯菌门(Chloroflexi)相对丰度(36.00%)明显高于S1(27.97%)、S2(25.67%)样品。推测这是由于分子量50 000 g·mol−1的PLA无法释放出足够碳源以供微生物生长所需,使得微生物细胞裂解释放肽聚糖,造成绿弯菌门(Chloroflexi)等菌群相对丰度的增加。放线菌门(Actinobacteriota)与CPR细菌(Patescibacteria)常见于城市污水处理厂活性污泥内[36-37]。
由图8 (a) 可知,S1样品拟杆菌门(Bacteroidota)、放线菌门(Actinobacteriota)和酸杆菌门(Acidobacteriota)相对丰度均高于S2和S3样品。拟杆菌门(Bacteroidota)下的微生物可以降解纸浆等有机物从而释放碳源[34],放线菌门(Actinobacteriota)下的微生物能合成胞外水解酶降解大分子有机物[38],其部分属能够水解PLA[21]。随着PLA分子量的升高,拟杆菌门(Bacteroidota)和放线菌门(Actinobacteriota)丰度降低。这表明5 000 g·mol−1分子量PLA更容易富集PLA水解相关菌门,从而释放更多碳源。酸杆菌门(Acidobacteriota)中的微生物带有narG、nirK和nirS等多种反硝化功能基因能够编码合成硝酸盐还原酶和亚硝酸盐还原酶,并能在厌氧条件下利用乙酸等有机物还原硝酸盐[39-40]。
样品在科水平上的分布如图7 (b) 所示,样品中共检测出32个科。前十个主要优势科是:嗜甲基菌科(Methylophilaceae)(9.82%~14.32%)、厌氧绳菌科(Anaerolineaceae)(3.44%~11.62%)、生丝微菌科(Hyphomicrobiaceae)(4.04%~4.97%)、norank_o_C10-SB1A(2.44%~3.76%)、亚硝化单胞菌科(Nitrosomonadaceae)(2.40%~3.77%)、红环菌科(Rhodocyclaceae)(1.96%~4.53%)、unclassified_p_Patescibacteria(0.18%~5.25%)、腐败螺旋菌科(Saprospiraceae)(2.37%~2.63%)、A4b(1.56%~3.94%)、暖绳菌科(Caldilineaceae)(2.18%~2.80%)。嗜甲基菌科(Methylophilaceae)、厌氧绳菌科(Anaerolineaceae)、生丝微菌科(Hyphomicrobiaceae)、红环菌科(Rhodocyclaceae)和暖绳菌科(Caldilineaceae)均为硝酸盐还原菌[41-43];亚硝化单胞菌科(Nitrosomonadaceae)是一种氨氧化菌(ammonia oxidizing bacterium,AOB),能氧化氨氮为亚硝酸盐[44];腐败螺旋菌科(Saprospiraceae)是一种亚硝酸盐还原菌[45]。上述各菌科均为反硝化脱氮过程的关键功能科,广泛存在于全球各地废水脱氮反应器中。norank_o_C10-SB1A 为硝酸盐异化还原为氨(DNRA)功能菌群[46]。A4b曾在含有PLA的反硝化系统中检测出,是一种能降解大分子有机物的微生物[47]。
如图8 (b) 所示,S1样品生丝微菌科(Hyphomicrobiaceae)、norank_o_C10-SB1A和亚硝化单胞菌科(Nitrosomonadaceae)相对丰度高于S2和S3样品。这表明含有5 000 g·mol−1分子量PLA的反应器中起主要反硝化作用的菌科为生丝微菌科(Hyphomicrobiaceae),且该菌生物活性较高,能在相对丰度小于5%的情况下,完成反硝化脱氮。norank_o_C10-SB1A和亚硝化单胞菌科(Nitrosomonadaceae)丰度较高,可能是因为5 000 g·mol−1分子量PLA能够释放较多碳源以致DNRA菌群(norank_o_C10-SB1A)繁殖[48],使得短时间内NH4+-N得到积累,促进了以NH4+-N为基质的亚硝化单胞菌科(Nitrosomonadaceae)的生长。因此,尽管norank_o_C10-SB1A相对丰度较高,出水NH4+-N却未见明显积累。
样品在属水平上的分布如图7 (c) 所示,共检测出38个属。前十个主要优势属为:甲基娇养杆菌属(Methylotenera) (9.09%~13.26%)、norank_f_Anaerolineaceae (3.03%~10.85%)、生丝微菌属(Hyphomicrobium) (3.50%~4.17%)、norank_f_norank_o_C10-SB1A (2.44%~4.00%)、norank_f_Caldilineaceae (2.10%~2.74%)、norank_f_Saprospiraceae (1.65%~1.91%)、unclassified_p_Patescibacteria (0.18%~5.25%)、norank_f_Steroidobacteraceae (1.34%~2.78%)、norank_f_Blastocatellaceae (0.18%~1.70%)、norank_f_norank_o_norank_c_JG30-KF-CM66 (1.49%~2.62%)。其中,甲基娇养杆菌属(Methylotenera)和生丝微菌属(Hyphomicrobium)是典型反硝化菌属具有还原硝酸盐的作用,常见于反硝化活性污泥[49-50]。norank_f_Anaerolineaceae、norank_f_Blastocatellaceae、norank_f_norank_o_norank_c_JG30-KF-CM66和norank_f_Steroidobacteraceae属均与复杂有机物代谢有关[51-54]。norank_f_Caldilineaceae和norank_f_Saprospiraceae属是通过消耗有机物进行反硝化作用的功能细菌[55-56]。norank_f_norank_o_C10-SB1A为norank_o_C10-SB1A科下的细菌为DNRA作用的关键功能属。
如图8 (c) 所示,S1样品中生丝微菌属(Hyphomicrobium)、norank_f_Steroidobacteraceae、norank_f_Blastocatellaceae和norank_f_JG30-KF-CM45属相对丰度高于S2和S3样品。生丝微菌属(Hyphomicrobium)是隶属于生丝微菌科(Hyphomicrobiaceae)的微生物,是反硝化作用的关键细菌。 norank_f_JG30-KF-CM45属也是反硝化功能属,丰度与有机物浓度成正比[55]。norank_f_Blastocatellaceae属参与代谢有机高分子物质,可能与聚合物水解过程有关[54]。norank_f_Steroidobacteraceae属参与木质素生物降解和解聚过程[52],也可水解有机聚合物。属水平结果表明5 000 g·mol−1分子量PLA能富集更多水解功能菌群。
总体来说,不同分子量PLA脱氮系统内微生物在门、科和属水平上的分布不同。5 000 g·mol−1分子量的PLA更有利于富集水解功能菌群。PLA反硝化生物脱氮路径主要有两条:反硝化功能菌群利用水解大分子有机物的微生物分解PLA释放的小分子碳源作为电子供体进行异养反硝化作用;微生物利用溶解性微生物代谢产物等有机物进行反硝化脱氮。
-
1) PLA作为固体碳源,在清水中能够长期稳定释放碳源,反硝化脱氮过程中释碳主要依靠生物作用。PLA反硝化脱氮效果表明,PLA分子量越低,脱氮效率越高。不同分子量PLA出水NO2−-N和NH4+-N等副产物均未见明显积累。5 000 g·mol−1分子量的PLA作为反硝化固体碳源更具有优势。
2) DOM组分和微生物测序结果表明,5 000 g·mol−1分子量的PLA更有利于富集水解相关功能菌群。PLA脱氮路径有两条:一是反硝化功能菌群利用水解微生物分解PLA释放的小分子碳源作为电子供体进行异养反硝化作用;一是微生物利用溶解性微生物代谢产物等有机物进行反硝化脱氮。
不同分子量聚乳酸的异养反硝化脱氮性能及脱氮反应机制
Heterotrophic denitrification and denitrification mechanism of polylactic acid with different molecular weight
-
摘要: 聚乳酸(PLA)是一种发展潜力巨大的反硝化固体碳源。为探究不同分子量PLA反硝化脱氮路径及机制,以不同分子量PLA为固体碳源研究了PLA的静态释碳性能和反硝化脱氮效果,考察了反硝化出水溶解性有机质(DOM)组分和微生物群落结构。结果表明:PLA释碳稳定,在清水和脱氮反应器中出水COD均能稳定在20 mg·L−1;PLA分子量越低,脱氮效果越好,5 000 g·mol−1分子量PLA的NO3−-N去除率和反硝化速率达到100%和1.29 mg·L−1·h−1。PLA脱氮路径有2条:一是反硝化功能菌群利用水解微生物分解PLA释放的小分子碳源作为电子供体进行异养反硝化作用;一是微生物利用溶解性微生物代谢产物等有机物进行反硝化脱氮。该研究结果可为PLA固体碳源在反硝化脱氮工艺中的高效应用提供参考。Abstract: Polylactic acid (PLA) is a kind of denitrification solid carbon source with great potential. In order to explore the denitrification pathway and mechanism of PLA with different molecular weights, the static carbon release performance and denitrification efficiency of PLA with different molecular weights were studied, and the effluent of dissolved organic matter (DOM) components and the microbial community structure were analyzed. The results showed that PLA carbon release performance was stable, and the effluent COD concentration in both deionized water and nitrogen removal reactors could be stabilized at 20 mg·L−1. The lower the molecular weight of PLA was, the better the nitrogen removal efficiency was. The NO3−-N removal rate and denitrification rate of 5 000 g·mol−1 molecular weight PLA reached 100% and 1.29 mg·L−1·h−1. There were two pathway for PLA denitrification, one was that denitrifying bacteria used the small molecule weight carbon source released by hydrolyzed microorganisms PLA as electron donor for heterotrophic denitrification,the other one was that microorganisms used organic matter such as dissolved microbial metabolites for denitrification. The research results can provide a theoretical basis for the efficient application of PLA as solid carbon source in denitrification process.
-
中国是抗生素生产和使用大国[1]。作为中国最常用的5类抗生素之一,β-内酰胺类抗生素生产量和销售份额在众多抗生素中名列前茅[2],在全球制药行业也具有举足轻重的地位[3-4]。青霉素和头孢菌素是使用最广泛的抗生素。2010年,青霉素和头孢菌素2种抗生素使用量占所有抗生素使用总量的近60%,与2000年相比增长了41%[5]。环境中β-内酰胺类抗生素的残留主要来自制药工业[6-9]、临床[10-12]和畜禽养殖[13-14]。抗生素生产过程产生的制药废水[8-9]及菌渣[15-16]中含有高浓度的抗生素残留[17]。LI等[18]对青霉素G生产废水(废母液)的研究表明,由于溶剂萃取过程的高温酸性环境使大部分青霉素G母体降解(残留浓度为153.0 μg·L−1),降解产物脱羧青霉噻唑酸和青霉醛的浓度可分别达到389.0 mg·L−1和75.3 mg·L−1。与青霉素G生产废水相比,青霉素生产排放菌渣中的青霉素G残留更高,可达5 000 mg·L−1[19],是β-内酰胺类抗生素排放强度最高的排放源。
尽管β-内酰胺类抗生素是生产和应用最广泛的抗生素,但由于其结构中的β-内酰胺环具有易水解和生物降解的特性,通常环境中检出浓度较低,甚至一些类型低于检测限。青霉素生产废水处理后,出水和受纳河流下游青霉素G浓度仅为1.68 μg·L−1和0.35 μg·L−1,但却发现大量对β-内酰胺类和其他各类抗生素具有多抗性的抗药细菌,这表明即使废水中抗生素母体浓度很低,其排放仍然可能导致环境细菌抗药性潜在风险[18]。头孢菌素生产废水中也出现了类似的现象[7]。制药菌渣中的青霉素G[20]、头孢菌素C[16]残留及转化产物导致在好氧堆肥过程中[17, 21]或土壤施用菌渣肥[20]后亦出现抗性基因和抗药菌升高的现象[20, 22]。因此,深入认识环境中β-内酰胺类抗生素的环境行为、抗性的产生和传播、高浓度污染源的抗生素去除等,对于该类物质的环境管理和污染控制至关重要。
本研究在对β-内酰胺类抗生素污染特征、环境行为和控制技术的研究进展进行梳理的基础上,重点关注了β-内酰胺类抗生素中最重要的“母体”抗生素原料药——青霉素G和头孢菌素C的环境行为和控制,并在构筑抗生素和抗性基因控制多级屏障技术体系和危险废弃物制药菌渣无害化处理及资源化利用等方面提出展望,以期为β-内酰胺类抗生素残留效价的削减和抗性的控制提供参考。
1. β-内酰胺类抗生素的性质和主要环境排放源
β-内酰胺类抗生素是指化学结构包含一个β-内酰胺环,具有抗菌活性的天然或经化学手段合成的有机化合物[23-25]。表1和图1列举了主要的β-内酰胺类抗生素及其分子结构。
表 1 主要的β-内酰胺类抗生素Table 1. Major β-lactam antibiotics分类 亚类 化合物名称 青霉素类 天然青霉素 青霉素G 耐酸青霉素 青霉素V、苯氧乙基青霉素 耐酶青霉素 苯唑西林、氯唑西林、双氯西林、氟氯西林 广谱青霉素 氨苄西林、阿莫西林、匹氨西林 抗绿脓杆菌青霉素 羧苄西林、磺苄西林、替卡西林等 头孢菌素类 第1代 头孢噻吩、头孢噻啶、头孢唑啉等 第2代 头孢呋辛、头孢孟多、头孢替安等 第3代 头孢哌酮、头孢噻肟、头孢克肟等 第4代 头孢吡肟、头孢匹罗等 第5代 头孢洛林、头孢托罗、头孢吡普等 非典型类 头霉素类 头孢西丁、头孢美唑、头孢替坦等 拉氧头孢类 拉氧头孢 β-内酰胺酶抑制剂 克拉维酸、舒巴坦等 单环β-内酰胺类 氨曲南、卡芦莫南 碳青霉烯类 亚胺培南,美罗培南,帕尼培南等 | Show Table DownLoad:
CSV
DownLoad:
CSV
青霉素G和头孢菌素C是2种重要的天然抗生素和原料药。青霉素于1928年被发现。1949年,科学家通过X射线晶体学研究揭示了青霉素的完整结构,并于1959年通过全合成证实了该结构[26-27]。1945年,分离得到的头孢子菌可以产生多种成分的抗生素,1955年,分离得到头孢菌素C[28-29]。在不改变核心结构β-内酰胺环基础上,通过人工修饰侧链[30],又发展出了第4代头孢菌素、β-内酰胺抑制剂、碳青霉烯、单环菌素等多种β-内酰胺类抗生素[4, 31]。β-内酰胺类抗生素从20世纪初开始广泛使用,不仅应用于人类疾病的临床治疗,还可用于畜牧水产养殖和植物病虫害的防治[12-14, 32-33]。由于β-内酰胺类抗生素的分子结构与细胞壁黏肽结构中的D-丙氨酰-D-丙氨酸相似,可以竞争性地结合转肽酶,不可逆地抑制肽聚糖的形成,抑制细胞壁的合成,从而起到抗菌作用[34-37]。β-内酰胺环是抗菌活性基团,而侧链基团改变会引起抗菌谱或对β-内酰胺酶的敏感性等药理学特性变化[34, 38]。图2显示了青霉素G和头孢菌素C的分子结构。头孢菌素C-7位基团影响抗菌活性,而C-3和C-4位取代基主要决定药代动力学[30-31]。与青霉素相比,头孢菌素具有较低的致敏性和对β-内酰胺酶的敏感性,因此,被认为是广谱抗生素,可用于治疗革兰氏阴性和革兰氏阳性细菌的感染,并有效地阻止微生物的生长[31]。
环境中β-内酰胺类抗生素主要来自制药工业[6-9]、医用[10-12]和兽用[13-14]。抗生素原料药发酵生产过程产生的制药废水(废母液)[8-9]及菌渣[15-16]含有高浓度的抗生素残留[17],是强度最高的排放源。制药废母液中单一抗生素(如土霉素)的浓度可达1 000 mg·L−1[39]。在印度一个大型药物生产基地的污水处理厂中检出高浓度的氟喹诺酮类抗生素残留[9]。由于β-内酰胺环具有不稳定易水解的特性,与土霉素、氟喹诺酮类等结构相对稳定的抗生素相比,青霉素G残留浓度相对较低,浓度为150 μg·L−1左右。然而,其降解产物脱羧青霉噻唑酸和青霉醛的浓度可以达到389.0 mg·L−1和75.3 mg·L−1[18]。有学者[7]在中国北方一个年产头孢菌素3 000 t的制药园区排放的废水中,发现了700 μg·L−1的头孢呋辛残留。因此,尽管制药行业污水相对生活污水(含医院污水)排放量不大,但其污染物排放强度高,如果不进行有效处理,制药废水中的抗生素残留就可能直接排放到水环境或者通过管网进入城市污水处理厂。现行的污水处理技术对常规有机物处理效果良好,但是对抗生素的去除效率还较低,这也会导致抗生素从污水厂向环境中的排放[40-42]。
临床使用的抗生素可通过粪便、尿液及医疗废弃物排入环境。头孢菌素通过口服和肠胃外途径给药,排泄半衰期为0.25~9 h[43]。大多数头孢菌素以母体的形式排泄,代谢率为5%~65%,使得头孢菌素母体和在体内产生的代谢产物,以及排泄后形成的初级转化产物进入环境中[44]。医院是头孢菌素类抗生素的主要排放源[45],其中第3代和第4代头孢菌素抗生素检出较多。也有研究表明,医院废水不是城市污水中抗生素残留的主要来源。在欧洲,医院抗生素用量仅占总用量的5%~20%,而英国、美国和德国的社区使用量占比为70%~75%[46]。另外,抗生素也广泛用于畜牧水产养殖[47]:美国畜牧商每年将约11 200 t抗生素投加进牛、猪和家禽的饲料中,以促进畜禽的生长[48];兽药的使用也导致残留的抗生素随着动物排泄物进入环境中[13, 49]。排泄物中的抗生素一部分会直接渗入地下水,另一部分则通过粪便进入土壤[50],随着雨水淋洗作用迁移至河流[51]或湖泊[1, 52]。
2. 环境中β-内酰胺类抗生素的污染特征
来自制药、医用和兽用的β-内酰胺类抗生素残留,可通过在污水处理厂和土壤中的迁移进入水体被底泥吸附,或者随水体迁移进入其他水环境中。抗生素残留在医院废水[11, 46]、制药废水[6, 18]、地表水[11]、河流[24, 53]、污水厂废水[11, 54]、污泥[41]和菌渣[15, 17, 22]等不同环境基质中均有检出。β-内酰胺环不稳定,易水解开环[38, 55],同时还可以与β-内酰胺酶发生酶解反应,上述过程均使其丧失抗菌活性。因此,β-内酰胺类抗生素尽管是应用最广泛的抗生素,但通常在环境中浓度较低,甚至低于检测限。
表2列出了不同环境介质中β-内酰胺类抗生素的分布浓度。医院废水中浓度为20.0 ng·L−1~4.1 μg·L−1,与其使用量和预期排放浓度相比,检出浓度较低[56-57];青霉素G、青霉素V和头孢克洛等未检出[11]。在地表水和河水中这类抗生素的残留浓度较低,阿莫西林、头孢克洛、氯唑西林、青霉素G和青霉素V的浓度在0~250 ng·L−1[11];长江、黄河和珠江中β-内酰胺类抗生素残留浓度分别为(123.1±128.2)、(343.5±345.6)及(1 606.3±1 384.3)ng·L−1[24]。头孢菌素在不同基质中浓度在0.30~30 ng·L−1,污水厂废水中浓度最高[31]。城市污水厂中检出氨苄西林、头孢氨苄、头孢噻肟,浓度分别为77.2~383.0、65.7~525.0、38.4~93.0 ng·L−1[54]。制药废水中抗生素浓度比医院废水和污水厂浓度高。青霉素制药废水中青霉素G浓度只有0.153 mg·L−1,而水解产物青霉噻唑酸、脱羧青霉噻唑酸、青霉二酸、青霉异二酸和青霉醛的浓度分别高达8.49、389.00、23.50、1.05和75.30 mg·L−1[18]。相比之下,由于污水处理过程或水循环过程中,抗生素母体被迅速降解,使得活性污泥和河流沉积物中抗生素的浓度较低[58]。在活性污泥、剩余污泥和脱水污泥中,均未检出氨苄西林和阿莫西林[41],但其水解产物可能存在较高的浓度[18]。制药菌渣中,抗生素残留高于制药废水,例如青霉素鲜菌渣中青霉素G残留浓度为5 000 mg·L−1[15-17, 19]。作为β-内酰胺类抗生素强度最高的排放源,菌渣的无害化处理和资源化利用是抗生素残留削减的关键。
表 2 β-内酰胺类抗生素在不同环境介质中的分布浓度Table 2. Distribution concentration of β-lactam antibiotics in different environmental media环境基质 检测方法 抗生素及其转化产物名称 浓度 来源 医院废水 HPLC-MS/MS 阿莫西林、青霉素G、青霉素V、头孢克洛、氯唑西林、头孢氨苄 0~4 100 ng·L−1 [11] 制药废水 HPLC-MS/MS 青霉素G、青霉噻唑酸、脱羧青霉噻唑酸、青霉二酸、青霉异二酸、青霉醛、头孢呋辛、头孢唑啉、头孢噻肟、头孢曲松、阿莫西林、头孢菌素 0.13~703.84 mg·L−1 [7-8, 18, 59] 地表水 HPLC-MS/MS 氨苄西林、哌拉西林、阿莫西林、头孢克洛、氯唑西林、青霉素G、青霉素V 0~250 ng·L−1 [11, 56] 表层海水 HPLC-MS/MS 头孢氨苄 10~180 ng·L−1 [60] 城市污水 HPLC-MS/MS 青霉素G、头孢氨苄、头孢噻肟、阿莫西林、青霉素V、头孢克洛、氨苄西林、氯唑西林、头孢菌素、苯唑西林 0~1 400 ng·L−1 [11, 54, 57, 61-63] 河流底泥 HPLC-MS/MS 青霉素G、青霉噻唑酸、脱羧青霉噻唑酸、青霉二酸、青霉异二酸、青霉醛 0~6.56 mg·kg−1 [18, 53] 活性污泥 HPLC-MS/MS 青霉素G、青霉噻唑酸、脱羧青霉噻唑酸、青霉二酸、青霉异二酸、青霉醛 0.034~470 mg·kg−1 [18, 41] 制药菌渣 HPLC-UVHPLC-MS/MS 青霉素G、头孢菌素C 2 000~5 000 mg·L−1(鲜菌渣)70~420 mg·kg−1(干菌渣) [15-17, 19, 64] | Show TableDownLoad:
CSV
3. β-内酰胺类抗生素的环境行为
β-内酰胺类抗生素的环境行为主要包括吸附、迁移和降解。其中,吸附和迁移是β-内酰胺类抗生素在土壤中的重要环境行为。β-内酰胺类抗生素的生物降解主要由微生物的活动驱动,受到环境条件、微生物种群、化合物结构等因素影响[65]。非生物降解指抗生素不经生物作用而自行降解的过程,通常经过光解、水解等理化作用转化为其他物质。直接光解是去除湖水中头孢曲松钠的主要过程[52],也可能是地表水中头孢菌素类抗生素最重要的消除过程[33],但是在污水处理过程中光解作用贡献较小[40]。β-内酰胺类抗生素中的β-内酰胺环不稳定,在酸、碱、热、氧化剂、紫外线以及极性试剂存在的条件下易水解开环,从而丧失抗菌活性[38, 55, 66]。水解是β-内酰胺类抗生素最主要的降解途径,通过一些强化手段还可能应用于废水中抗生素的去除。因此,下面重点讨论β-内酰胺类抗生素在环境中的吸附迁移和水解过程,并特别关注大量生产的青霉素G和头孢菌素C这2种重要母体抗生素的降解途径和降解产物。
3.1 吸附与迁移
吸附对抗生素在环境中的滞留、迁移及转化有重要影响,也会影响到抗生素在环境基质中的活性和生物转化作用。吸附作用可将抗生素暂时以非生物活性的形态储存在土壤中,降低急性毒性并延长在基质中的停留时间[67]。吸附系数(Kd)用来衡量吸附能力。吸附能力强则容易在环境中累积;而吸附能力较弱则易随雨水发生迁移,并转至其他水体中,严重时还会影响地下水质。土壤的温度、pH、阳离子交换容量(cation-exchange capacity,CEC)、黏土矿物含量和有机碳含量(organic carbon,OC)会影响抗生素的吸附[5]。温度升高会影响活性污泥对不同抗生素的吸附作用。例如,温度升高后,污泥对阿莫西林、头孢噻肟、头孢克洛和头孢唑林的吸附作用增强,对普鲁卡因和头孢曲松的吸附作用降低。与青霉素类抗生素相比,活性污泥对头孢类抗生素的吸附作用受pH的影响更大[68]。
迁移是指抗生素随水体移动到其他环境基质中,或经过土壤渗透进入土壤深层或地下水的过程[69]。青霉素制药废水中,青霉素G及水解产物在处理后会随着废水的排放向收纳河流下游迁移[18]。迁移行为的影响因素包括吸附特性、淋洗速率和降解程度等。由于β-内酰胺类抗生素在土壤中的吸附能力较弱,很可能对地表及地下水造成污染[70]。抗生素在环境中的迁移能力取决于其理化特性、土壤理化特性、有机粪肥的施用和天气条件。酸雨可使抗生素加速从动物粪便向地表土壤迁移,而长时间的降雨还会促进抗生素在土壤中向下迁移[5, 50, 65]。
3.2 水解
水解是有机化合物,特别是酯类和酰胺类化合物在环境中最重要的降解途径。β-内酰胺环不稳定易水解开环。这一特性也是这类抗生素在无大量微生物种群的系统(如河流和地下水)中的主要降解途径[71]。温度和pH是影响水解速率最重要的参数。水解速率通常随着温度的升高而增加[71-72]。氨苄青霉素、头孢洛汀和头孢西丁3种抗生素在通常环境条件(pH=7,25 ℃)下的水解半衰期为5.3~27.0 d。碱性条件下的水解速率明显快于酸性和中性条件下的水解速率。温度升高10 ℃时,水解速率增加2.5~3.9倍[71]。在对阿莫西林的水解研究中也得到了类似结论[55]。
KESSLER等[73]探究了青霉素G在酸性条件下的两种降解途径,确定了青霉胺作为最终降解产物的酸性降解途径。青霉素G在酸性、碱性、中性的水解途径和酶解途径如图3所示[23,74]。在青霉素制药废水中检测到这些产物,并通过定量测定验证了产物之间的转化关系。在制药废水处理过程中,出水的青霉素G浓度为(1.68±0.48)μg·L−1,去除率为96.7%;脱羧青霉噻唑酸是青霉素的主要降解产物,浓度为(44.5±2.5)mg·L−1,摩尔浓度占总摩尔浓度的68.8%。高浓度的青霉素降解产物被排放到河流中,在污水排放的受纳河流下游,脱羧青霉噻唑酸、青霉噻唑酸和青霉二酸都具有相对的环境持久性[18]。
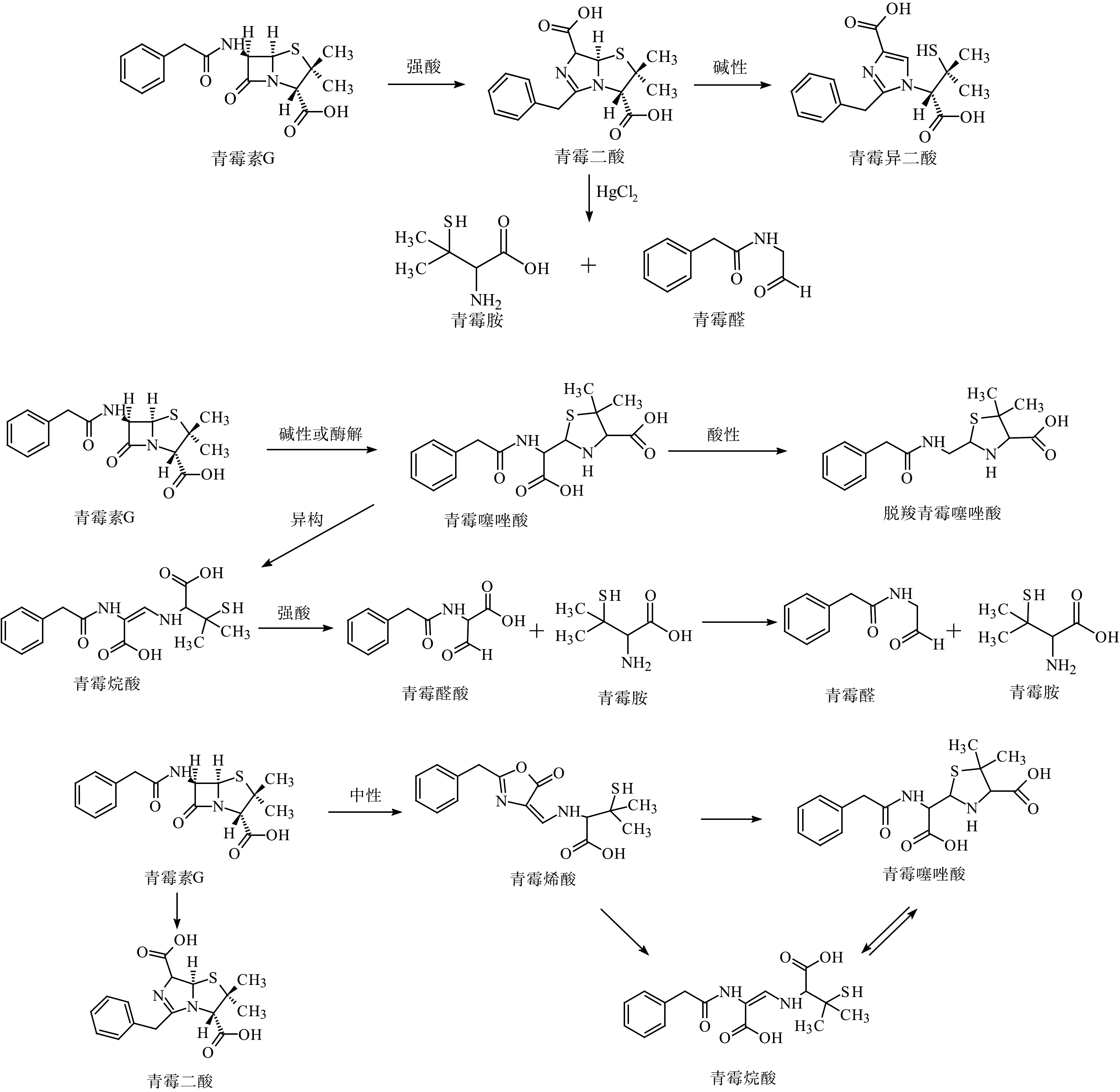 图 3 青霉素G在不同条件下的水解途径Figure 3. Hydrolysis pathway of penicillin G under different conditions
图 3 青霉素G在不同条件下的水解途径Figure 3. Hydrolysis pathway of penicillin G under different conditions与青霉素相比,头孢菌素中四元环和六元环形成的稠合结构对水解反应更稳定,但也会发生各种化学和酶促转化。C-3位取代基、β-内酰胺环、C-4位羧基和C-7位侧链酰胺键都是头孢菌素易发生降解反应的位置(图2(b))[75]。C-3位存在离去基,使得亲核试剂或β-内酰胺酶可以水解β-内酰胺环的C—N键[23]。在酸性、碱性和中性水解中,C-3位侧链分离是头孢菌素的主要降解途径之一。头孢菌素C的水解途径[23]如图4所示。与青霉素类似,β-内酰胺环也是头孢菌素水解反应的关键位点。开环反应根据侧链结构的不同分为:中性条件的一般开环水解、侧链α-氨基参与的分子内氨解及头孢菌素环缩合成噻唑结构。另外,还存在pH依赖型的立体异构,即在中性或酸性环境下C-6位发生立体异构,而碱性环境下C-7位发生立体异构和C-7位侧链的甲氧亚胺键构型发生顺反异构[75-76]。HIRTE等[55]研究阿莫西林在酸性、碱性、中性条件下的降解过程中,识别鉴定出了45个产物,证明了阿莫西林青霉噻唑酸、阿莫西林2',5'-哌嗪二酮和阿莫西林脱羧青霉噻唑酸并非稳定产物,并首次报道了青霉胺氧化成二硫化青霉胺的过程。
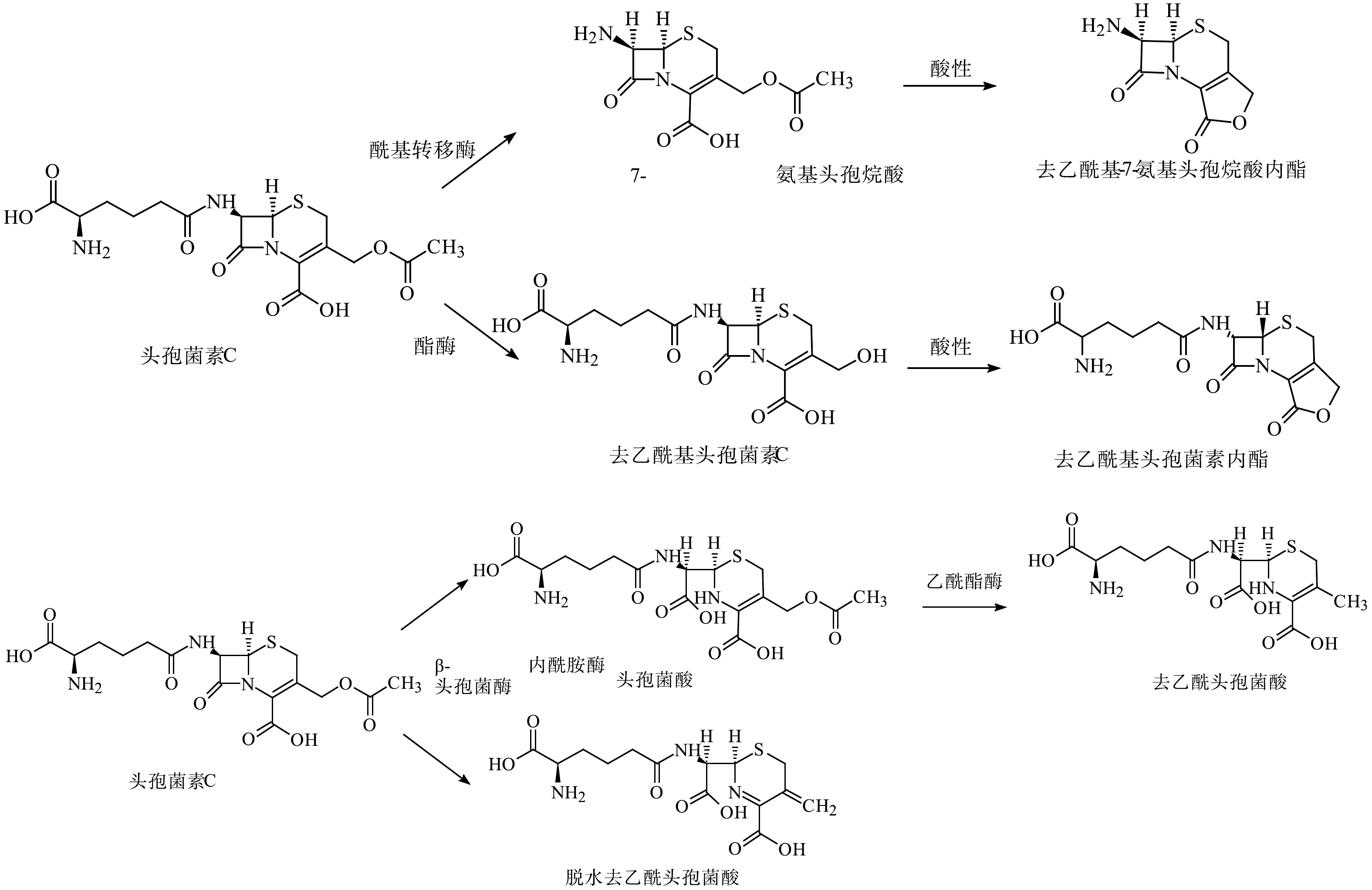 图 4 头孢菌素C在不同条件下的水解途径Figure 4. Hydrolysis pathway of cephalosporin C under different conditions
图 4 头孢菌素C在不同条件下的水解途径Figure 4. Hydrolysis pathway of cephalosporin C under different conditions4. β-内酰胺类抗生素抗性的产生和传播
抗生素的作用目标是细菌。若抗生素在环境中长期稳定存在,则会影响基质中的群落结构,导致环境中微生物菌群失调,诱导微生物抗性的产生,也会对动物、植物产生毒性作用,可能存在潜在的环境和健康风险。环境中抗生素残留对抗生素抗性细菌(ARB)及抗生素抗性基因(ARGs)的形成具有促进作用。
世界卫生组织将抗生素抗性列为一类亟需解决的全球性公共健康问题。各种抗生素第一次观察到抗性的时间序列已被系统总结[38]。19世纪40年代,临床上就出现了青霉素抗性菌。1946—1948年,青霉素抗药菌株比例从14%上升到59%。抗药菌的抗药机制[35, 37-38]主要包括4个方面:抗生素作用靶点突变、酶促反应使抗生素失活、细胞膜特性改变和外排泵机制、改变代谢途径。对于β-内酰胺类抗生素,细菌主要通过分泌β-内酰胺酶,释放到细胞外周胞质中水解药效基团β-内酰胺环,从而使其到达作用靶点前失活。此外,细菌也通过改变抗生素作用对象青霉素结合蛋白(PBPs)的结构产生抗性,但由于PBPs变异比较困难,故分泌β-内酰胺酶是β-内酰胺类抗生素最重要的抗性机制[77]。抗性基因是通常由基因突变产生的并赋予宿主细胞对抗生素有耐受性的一类基因。而抗性基因通过整合到质粒和整合子等水平转移元件上,实现了在不同菌种之间的水平转移[37]。初级抗性在微生物中天然存在,例如铜绿假单胞菌对青霉素G的抗性,相同物种生物通过细胞分裂(垂直转移)“继承”该抗性。在微生物与抗生素接触过程中会产生次级抗性,质粒介导的抗性可在微生物之间转移,染色体外遗传物质通过结合在不同细菌之间转移(水平转移)[78]。
碳青霉烯类抗生素是抗菌谱最广、抗菌活性最强的非典型β-内酰胺类抗菌药物。这类抗生素对超广谱β-内酰胺酶(ESBL)和头孢菌素酶 (AmpC酶)具有高度稳定性,是医学临床治疗中产生超广谱β-内酰胺酶类耐药肠肝菌科细菌等多重耐药革兰氏阴性菌感染的重要药物。随着这些药物广泛应用于临床,碳青霉烯类耐药肠杆菌科细菌等各种广泛耐药细菌出现并流行,碳青霉烯抗菌药物的疗效已经明显降低[79-80]。20世纪80年代,还出现铜绿假单胞菌、沙雷菌等对碳青霉烯类药物耐药的现象。KPC型碳青霉烯酶于1996年在美国发现[81]。目前,常见的碳青霉烯酶类耐药基因主要有KPC、IMP、VIM和NDM等,国内以KPC和IMP为主。碳青霉烯类抗生素抗药性产生的机理主要有3种:1)细菌通过产生碳青霉烯酶产生抗药性;2)产生质粒或染色体介导β-内酰胺酶合并外膜蛋白的缺失或数量的减少;3)药物作用靶位的特异性改变。其中,碳青霉烯酶所造成的危害最为严重[82]。由于碳青霉烯类耐药肠杆菌具有流行范围广、传播速度快、多重耐药以及引起的疾病死亡率高等特点,美国己经将碳青霉烯类耐药肠杆菌列为最为紧急的3大威胁之一[83]。目前,动物养殖、环境和临床细菌的碳青霉烯耐药菌和抗性基因及其关联性研究是一个热点。
不同环境基质中都已检出β-内酰胺类抗生素抗药菌和抗性基因。对自然水体中β-内酰胺酶基因和整合子的发生和分子多样性评估[84]发现,在77.8%的肠杆菌科和10.5%的气单胞菌中检测到β-内酰胺酶基因,最常检出基因是blaTEM。海水中超过90%的细菌菌株对1种以上的抗生素具有抗性,而20%的细菌对至少5种抗生素具有抗性。医院和集约化养殖场的废水可能是环境中抗药菌和抗性基因的主要污染源[85]。污水处理厂则被认为是环境中抗性基因传播的热点地区,因为污水处理过程为抗药菌增殖以及抗性基因在不同微生物之间的水平转移提供了便利条件[38]。目前,已有86种β-内酰胺抗性基因在11个国家的污水处理厂进水、出水和活性污泥中的分布和变化情况被报道[38]。污水处理厂中抗生素残留在抗性菌及抗性基因的维持和传播中起着重要作用,除了选择抗性表型,还可能干扰土著细菌群落。制药废水的抗生素残留进入水体后也能够改变水体的微生物系统[6, 86],通过选择性压力诱导水体中的微生物产生抗性基因,并进行传播扩散[85]。本课题组的前期研究也发现,由于可能受到废水中青霉素G及转化产物的影响,在青霉素生产废水和下游河流中发现了大量抗药菌,对β-内酰胺类抗生素具有极高的抗性,对其他各类抗生素也有广泛的抗性,具有一定的环境风险[86]。此外,携带了各种抗性基因的I型整合子(intI1)在废水和下游河流的菌株中也被检出,进一步说明了抗性基因在细菌间水平转移的可能性。制药菌渣中的青霉素G[20]、头孢菌素C[16]残留及转化产物在堆肥过程[17, 21]或施用菌渣肥[20]后,在土壤中也会诱导产生抗性基因[20, 22]和抗药菌[21]。
当考虑导致抗生素抗药菌和抗性基因出现、扩增和持续存在的因素时,传统的假设是“抗生素的使用是这些过程主要驱动力”[87]。处于亚抑制浓度的抗生素可能会影响细胞功能,并改变毒力因子基因表达或抗生素抗性转移。一些研究表明,医院废水中抗药性的增长与头孢菌素的使用呈正相关[10, 85];但也有证据表明,抗药性并不总与所用药物的量或环境中残留物的浓度有关[48]。肠胃外施用头孢噻呋对胃肠道中头孢噻呋抗药细菌的流行具有有限或短期的影响。虽然这种反应可能足以解释对头孢菌素广泛抗药的模式,但大约2/3的头孢噻呋代谢物会从尿液中排出,这增加了环境选择在扩增和维持抗生素抗药性起重要附加作用的可能性[87]。不同的基因可能具有不同的维持动力学,可能与携带它们的宿主微生物有关[85, 88]。环境中β-内酰胺类抗生素抗药菌和抗性基因发展和传播的机制还需要进一步深入探讨。
5. 废水和菌渣中β-内酰胺类抗生素的控制技术
与临床使用和动物养殖抗生素造成的环境广泛存在的污染相比,制药行业的污染源相对集中,但污染排放强度却是最高,故制药废水和菌渣是β-内酰胺类抗生素排放强度最高的排放源。若不对制药废水和菌渣进行有效控制,抗生素母体及残留效价则可能直接排放到环境中,或通过管网进入城市污水处理厂,从而导致抗生素从污水厂向环境中的排放。因此,必须在源头对抗生素残留效价进行控制,阻断抗生素和抗性基因向环境中排放。
对废水和菌渣中β-内酰胺类抗生素的去除技术,主要有生物处理和物化处理两类。通常采用厌氧、好氧和缺氧等生物处理过程去除制药废水中的COD,并实现氨氮的硝化反应和反硝化脱氮。以山西某制药厂为例,该厂青霉素生产废水生化处理后出水COD为218.0 mg·L−1,无法达到一级排放标准[89]。使用UASB反应器处理制药废水时,COD去除率为39%~85%,效果不稳定;同时6-氨基青霉烷酸和阿莫西林的去除率分别只有19%~33%和13%~47%[90]。当模拟配水中阿莫西林浓度从250 mg·L−1提高至650 mg·L−1时,活性污泥对阿莫西林的去除率从88.79%下降至完全无法去除[91]。因此,尽管β-内酰胺类抗生素容易生物降解,但因制药废水水质成分复杂,残留的抗生素和残留基质、试剂可能会影响生物处理效果[38, 61, 92]。抗生素的浓度、稳定性和对微生物的抑制作用都会造成处理效果的不稳定[31],更为严重的是可能会导致废水生物处理过程中抗性基因的产生和排放[93-95]。解决抗生素废水生物处理技术瓶颈的关键,是在废水进入生物处理之前,利用预处理技术选择性去除废水中的抗生素效价残留,降低对微生物的抑制作用,从而达到稳定的处理效果[96]。
抗生素生产过程除了产生制药废水,还会产生大量生产菌渣。制药菌渣含水率高,主要成分为菌丝体、剩余培养基、代谢中间产物、有机溶媒以及少量残留的抗生素[97]。2008年,抗生素菌渣被列入《国家危险废物名录》,如何实现抗生素菌渣合理有效利用与安全处置已成为制药企业亟待解决的难题[98]。许多研究者通过生物处理技术对菌渣进行无害化处理和资源化利用。例如,厌氧消化可以将头孢菌素C菌渣中低品位的有机质转化为高品位沼气,可实现菌渣资源化利用[99];好氧堆肥可以消除青霉素G菌渣中的抗生素残留,但堆肥过程也会导致抗药菌和抗性基因的产生[100-103],在其他青霉素菌渣好氧堆肥的研究中也得出了相似的结论[17]。最近的研究[104]发现,采用高温(70 ℃)厌氧消化对螺旋霉素菌渣进行预处理具有可行性。高温消化可大幅度降低抗生素的浓度,同时由于相应宿主的减少,抗药基因的丰度也在7 d之内减少。实际上,处理菌渣问题的关键是研发出去除抗生素效价残留的有效技术,以阻断抗性传播为目标,并在此基础上实现菌渣无害化处理及资源化利用。
氧化等一些物理化学技术通常对于水中抗生素的去除具有很好的效果。表3列出了常见的物理化学去除工艺及效果。吸附[105-106]和膜过滤[57]是常规的物理处理技术。这类技术通过分子间作用力或孔径截留作用将抗生素从一个相中分离转移至另一相,但抗生素本身没有发生降解反应。由于头孢菌素具有在相对较宽的波长范围内的光吸收作用,故光催化技术广泛用于头孢菌素类抗生素的去除技术的研究中[33, 52],但目前还难以进行实际工程应用。臭氧、芬顿等高级氧化技术在β-内酰胺类抗生素的去除研究中体现出反应速度快和去除效率高的特点,可以高效去除溶液中抗生素母体[107-108],但在氧化过程产生的降解产物的结构、毒性,特别是在复杂实际废水中的适用性还有待进一步探究[31]。在青霉素制药废水深度处理工艺探究中发现,Fenton氧化法、Cu/Fe催化还原法、臭氧氧化法均可有效降低COD残留。在Fenton氧化最优条件下,出水COD含量可降至22.5 mg·L−1 [89]。采用Fenton氧化和Fe/C微电解2种方法预处理7-氨基头孢烷酸制药废水。结果表明:2种方法均可有效提高COD去除率和废水可生化性;但Fenton氧化法存在药耗量大,处理成本高的问题;而Fe/C微电解发也存在反应时间较长的问题[109]。物理化学处理技术在制药菌渣无害化和资源化过程中也有一些初步研究。例如,青霉素菌渣经过100 ℃热水解作用,98%的抗生素残留被去除,制取的菌渣肥可以作为有机肥料应用于农业生产[15, 20]。也有研究者发现,微波加热温度可显著影响菌渣中残留抗生素的去除率,700 W辐射功率下快速升温至100 ℃条件下持续处理15 min,头孢菌素C的去除率可以达到99.9%[16]。
表 3 常见的物理化学处理工艺及处理效果Table 3. Common physical and chemical treatment process and treatment effect处理工艺 基质 处理条件 抗生素名称 浓度 去除效果 来源 吸附 制药废水 pH 为2~7 温度 30 ℃吸附剂量0.1~3.5 g 阿莫西林 317 mg·L−1 膨润土为吸附剂,去除率88.01%;活性炭为吸附剂,去除率94.67% [59] 膜处理 城市污水 MF微滤2 min,平衡罐5 min,RO反渗透 0.5 min,加氯反冲洗 阿莫西林 90 ng·L−1 头孢氨苄去除率87%~100%,其余抗生素去除率100% [57] 氯化 模拟配水 阿莫西林和头孢拉啶:ClO2和抗生素摩尔比为 0.25~2.0,pH=8.0反应时间1 min;青霉素:ClO2和抗生素摩尔比为 0.25~1.5,pH=3.5反应时间2 h 阿莫西林头孢拉啶青霉素G 1.6 mg·L−11.8 mg·L−125.0 mg·L−1 阿莫西林和头孢拉啶在1 min内全部降解,青霉素可以在2 h内全部降解 [110] 芬顿 模拟配水 pH为3,H2O2浓度25 mmol·L−1Fe3+浓度1.5 mmol·L−1 青霉素G 400 mg·L−1 配水中COD的去除率达56%,TOC的去除率达46% [111] 模拟配水 m(COD): m(H2O2): m(Fe2+)=1:3:0.3pH=3 阿莫西林氨苄青霉素氯唑西林 104 mg·L−1105 mg·L−1103 mg·L−1 2 min抗生素完全降解,10 min生物降解性提高到0.37,60 min COD和DOC降解率为81.4%和54.3% [112] 光催化 模拟配水 pH 为3.0~9.0,太阳光源,催化剂TiO2 0.1~0.7 g·L−1或不同比例铁碳混合物 阿莫西林 100 mg·L−1 最优条件下去除率可以达到85% [113] 模拟配水 pH为 3.0~11.0,紫外光源365 nm,6 W催化剂TiO2 0.5~2.0 g·L−1,H2O2 50~300 mg·L−1 阿莫西林氨苄青霉素氯唑西林 104 mg·L−1105 mg·L−1103 mg·L−1 pH=5,TiO2 1.0 g·L−1,抗生素去除率均达50%,DOC去除率81%;pH=5,TiO2为1.0 g·L−1,H2O2为100 mg·L−1,抗生素30 min内完全降解,24 h矿化率达40% [114] 臭氧 模拟配水 pH为2.5~7.2O3浓度为1.6×10−4 mol·L−1 阿莫西林 210 mg·L−1 4 min阿莫西林去除率90%,20 min阿莫西林矿化率可以达到18.2%,延长时间后,矿化率保持较低水平 [115] 模拟配水 pH为3.0~11.0,O3量为3 g·(h·L)−1H2O2浓度0~200 mmol·L−1 头孢曲松青霉素G − 60 min抗生素去除率95%,TOC去除率45%,提高废水可生化性 [110] 模拟配水 预臭氧pH 为7.0~12.0,O3量1 800 mg·(h·L)−1;H2O2 100 mmol·L−1 青霉素G 600 mg·L−1 pH=7,预臭氧工艺提高废水可生化性,但并不能完全去除生态毒性,还存在严重生物抑制作用;1 h内臭氧COD去除率为37%,而经预臭氧COD去除率达76% [116] 水热 制药菌渣 高压釜内部容积1.0 L,内径70 mm,温度100~220 ℃ 头孢菌素C − 在100~220 ℃和0~60 min内,头孢菌素C的去除率为99.0%~99.9% [66] 热水解 制药菌渣 盐酸2 mol·L−1,投加量10~60 mL,温度50~100 ℃ 青霉素G 9 g·mL−1(效价) 最优条件盐酸投加40 mL浸泡1 h,90 ℃水浴中加热机械搅拌3 h,过滤,效价去除率80.93% [64] 制药菌渣 加水倍数为3,水解温度60 ℃,水解时间30 min 青霉素G 5 000 mg·kg−1 水解后菌渣青霉素残留小于0.5 mg·kg−1,凯式氮削减率大于45%,有效降低青霉素残留及凯氏氮对厌氧消化的影响,为厌氧消化高效、稳定进行创造有利条件 [19] 热水解 制药菌渣 温度60~100 ℃ 青霉素G 2 000 mg·L−1 60 ℃时,20 min去除率为20%;100 ℃时,20 min去除率为98% [15] 微波 制药菌渣 温度100 ℃,微波功率为300、500、700 W 头孢菌素C 420 mg·kg−1 前150 s降解效率较低,此后降解速率明显提升,不同功率条件微波辐射15 min,降解率均超过99.9% [16] | Show TableDownLoad:
CSV
综上所述,尽管生物处理过程能削减部分抗生素,但抗生素残留会导致处理效果不稳定和抗性基因产生、排放问题。高级氧化技术可以在生物处理之后进一步去除残留抗生素,阻断抗生素向环境排放。但该技术主要用于模拟废水的研究,在实际废水处理中还存在操作困难、选择性差、成本高等问题,且可能产生具有较强毒性的未知中间产物[117]。因此,如何从制药废水中高效选择性去除残留抗生素及其相关物质仍然是一个技术挑战。阻断抗性基因向环境中排放的关键,是在生物处理工序之前有效削减和控制抗生素效价残留,并根据抗生素特性,开发针对抗生素抗菌活性基团的源头预处理技术。笔者所在研究团队根据四环素类抗生素药效官能团易水解的特性,探究了四环素类抗生素的水解动力学及其效价变化,通过优化水解条件可以显著加快四环素和土霉素的水解,并有效降低抗生素对生物处理微生物的抑制[118]。在此基础上进行了现场中试实验[119],验证了强化水解预处理工艺,并与生物处理结合进行现场工程应用,实现了常规指标、抗生素及抗性基因的源头控制。该技术思路有望实现β-内酰胺类抗生素废水的无害化处理。同时,抗生素及其代谢产物残留,还有抗性基因均是菌渣资源化利用造成环境风险的重要安全隐患,在以后的研究中需要从这两个方面对菌渣资源化利用进行安全性评估,为菌渣利用安全性评估方法和控制标准的制定提供科学基础。
6. 研究展望
1)由于β-内酰胺类抗生素具有易水解和生物降解的特性,在环境中通常浓度很低或低于检测限。然而,抗生素母体的削减并不意味着环境效应的消除,各种水解、降解产物等相关物质可能会贡献残留效价,导致环境中β-内酰胺抗药菌和抗性基因的产生和传播。因此,需要对β-内酰胺类抗生素的降解产物和降解途径进行深入探究,进一步明确降解产物与细菌抗性产生和传播之间的关系。
2)对β-内酰胺类抗生素残留的去除,除了母体之外,还应关注抗生素残留效价,将具有抑菌活性的中间产物、前驱体或者生产副产物,以及经过物化或者生物降解等过程后保留抗菌活性的中间转化产物纳入β-内酰胺类抗生素污染控制评价体系。为了明确残留抗生素和效价的控制目标和标准,需要对抗生素在促进抗性基因产生和发生水平转移方面的浓度阈值进行研究。
3)针对β-内酰胺类抗生素制药废水等高浓度抗生素残留的废水,需要开发选择性地去除抗生素效价的预处理技术,从源头解决由于抗生素效价残留造成生物处理效果不稳定的问题,阻止抗性的产生;同时采用末端保障深度处理技术,阻断抗生素残留、抗性菌和抗性基因向环境的传播,保障废水的安全排放。为了构筑抗生素和抗性基因控制多级屏障技术体系,应当将源头控制预处理技术和末端保障深度处理技术联合起来。
4)包括β-内酰胺类抗生素在内的发酵类抗生素的制药菌渣已被列入《国家危险废物名录》。针对其产生量大、处理难度大等现实问题,并根据《制药工业污染防治技术政策》(征求意见稿) 中提出的“鼓励开发发酵菌渣在生产工艺中的再利用技术、无害化处理技术、综合利用技术”政策建议,如何实现抗生素菌渣合理有效利用与安全处置已成为制药企业亟待解决的难题[98]。未来的研究将以去除抗生素效价残留、阻断抗性传播为目标,开发和优化菌渣处理技术,同时围绕菌渣资源化利用过程中抗生素残留效价和抗性发展进行系统评价,为菌渣无害化处理及资源化利用提供参考。
-
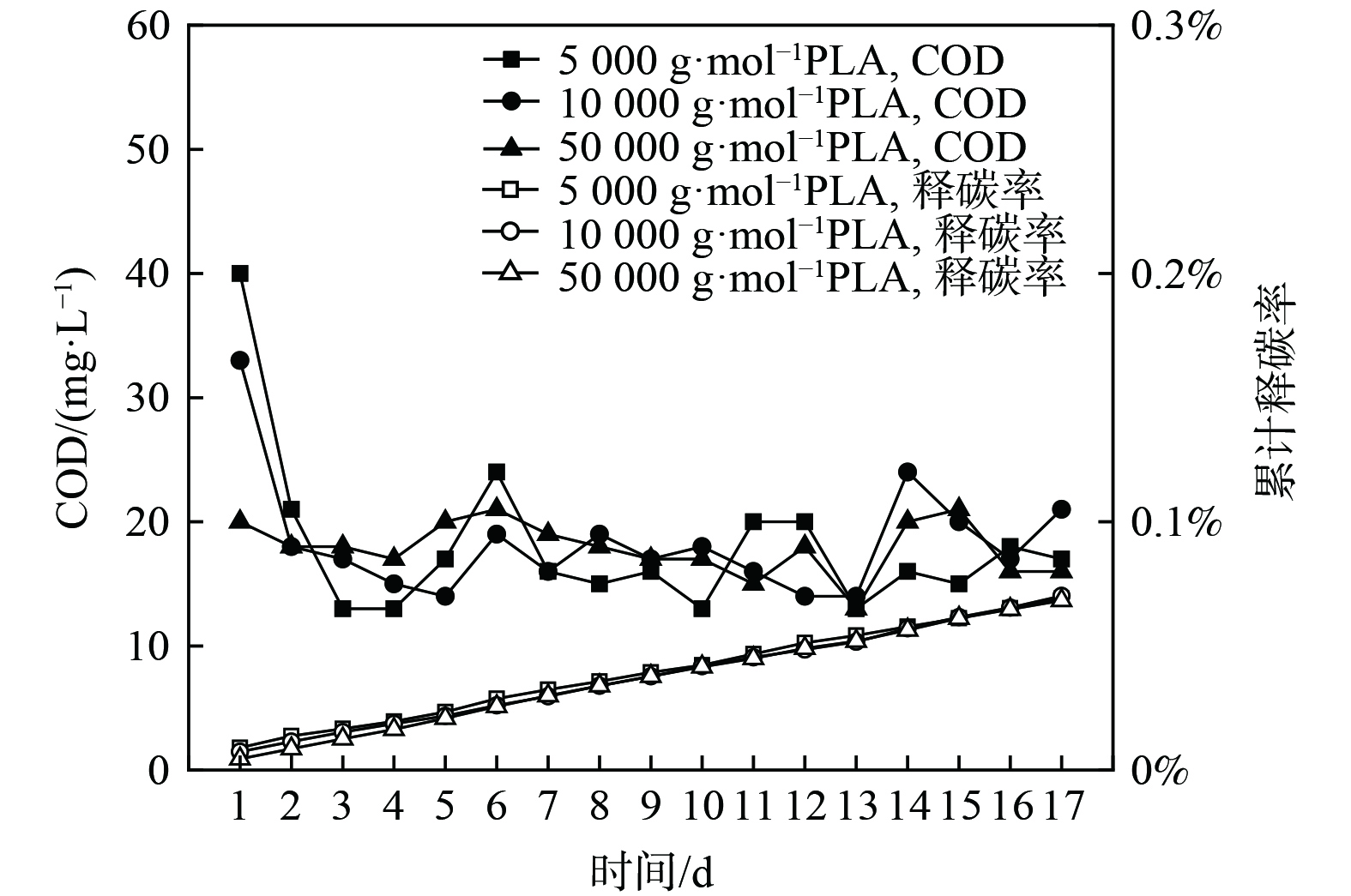
图 1 不同分子量PLA释碳性能
Figure 1. Carbon release properties of PLA with different molecular weights
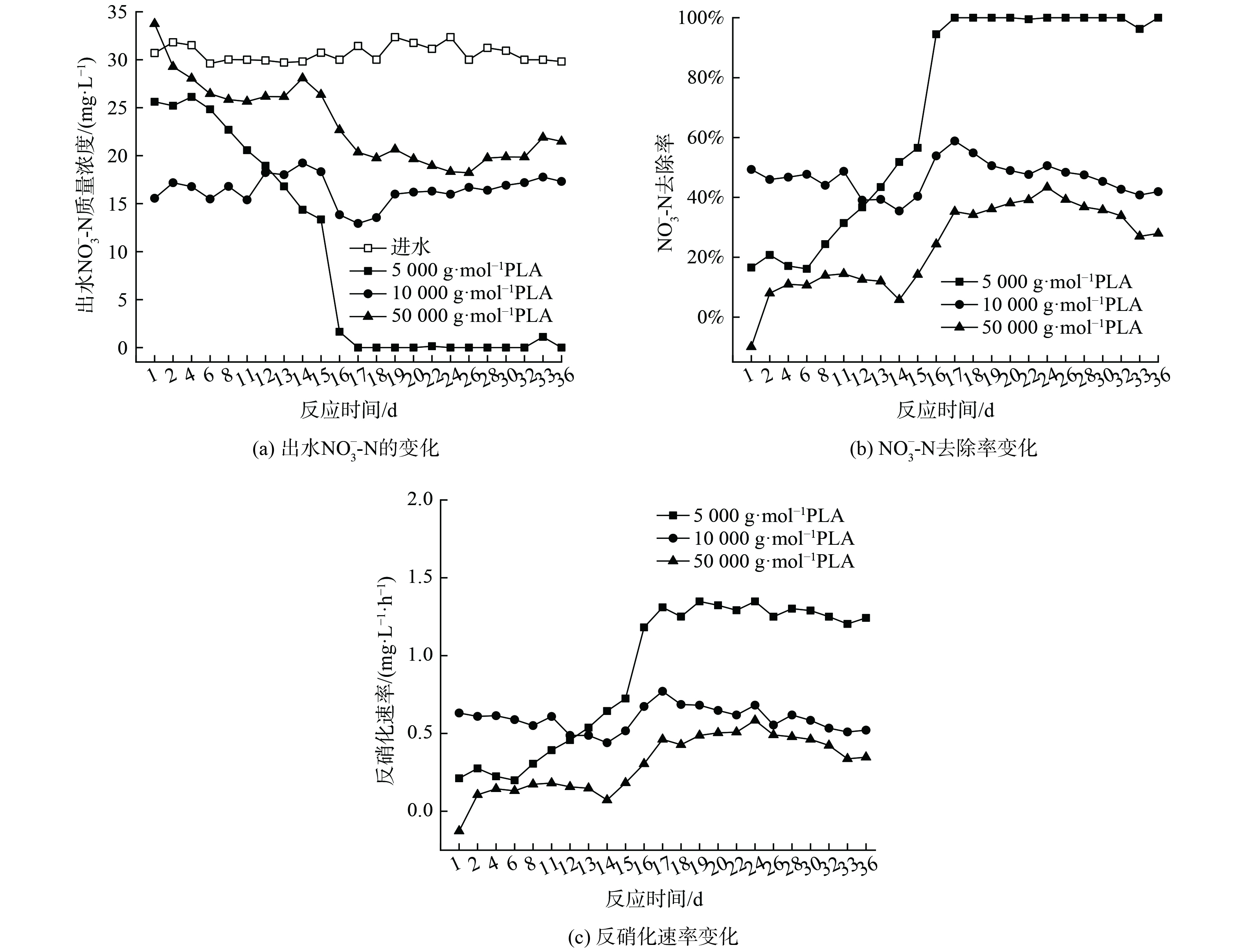
图 2 不同分子量PLA的脱氮性能对比
Figure 2. Comparison of PLA denitrification properties of different molecular weights
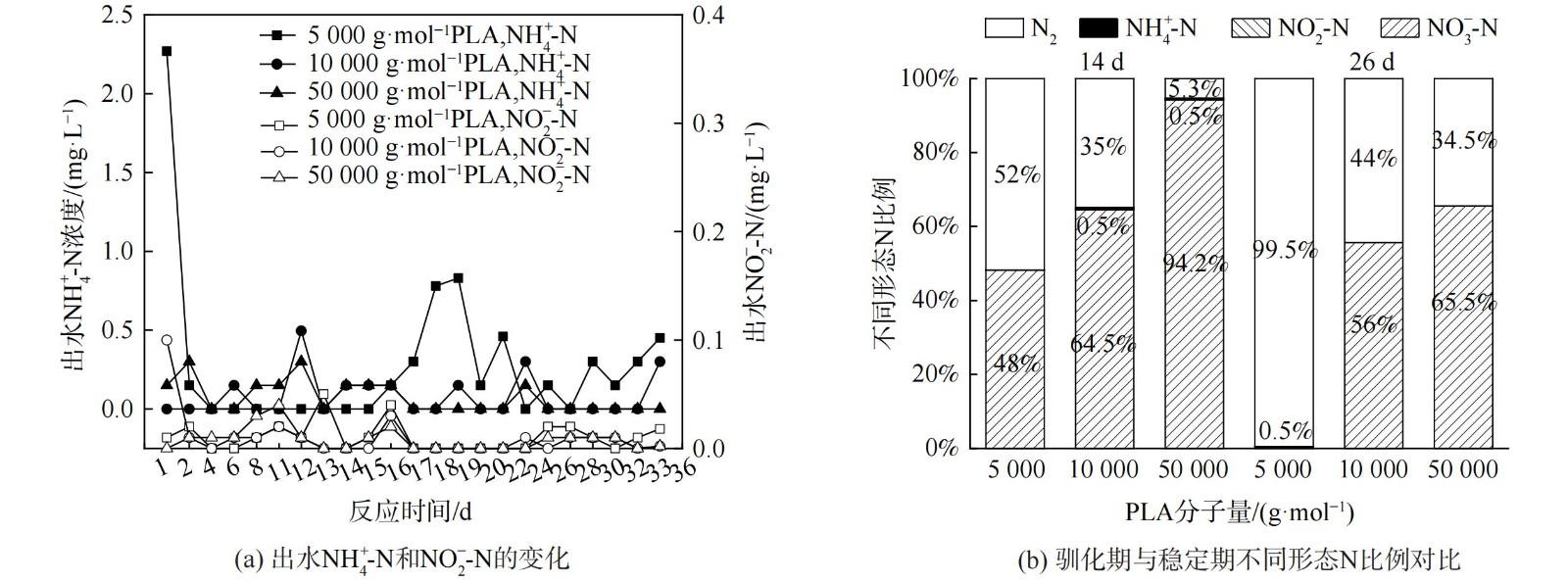
图 3 不同分子量PLA反硝化实验出水NH4+-N质量浓度、NO2--N质量浓度和不同形态N比例
Figure 3. Concentrations of NH4+-N, NO2--N and different morphological N ratios in denitrification effluent from PLA with different molecular weights
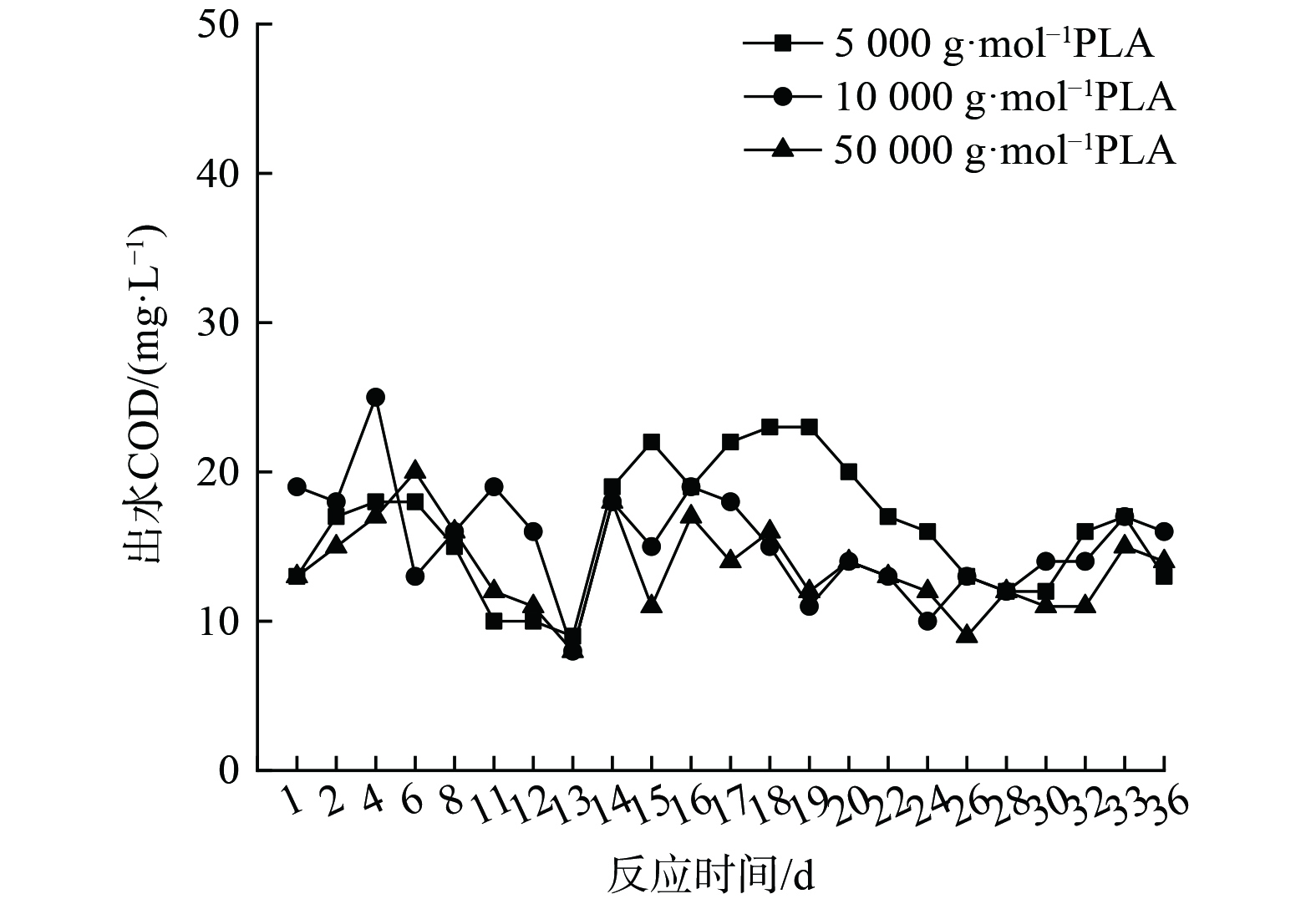
图 4 不同分子量PLA反硝化实验出水COD
Figure 4. Concentrations of COD in denitrification effluent of PLA with different molecular weights

图 5 不同分子量PLA反硝化实验1 d、17 d、37 d出水三维荧光光谱(3D-EEM)图
Figure 5. Three-dimensional fluorescence spectra (3D-EEM) of effluent from denitrification experiments with different molecular weights at 1 d, 17 d and 37 d
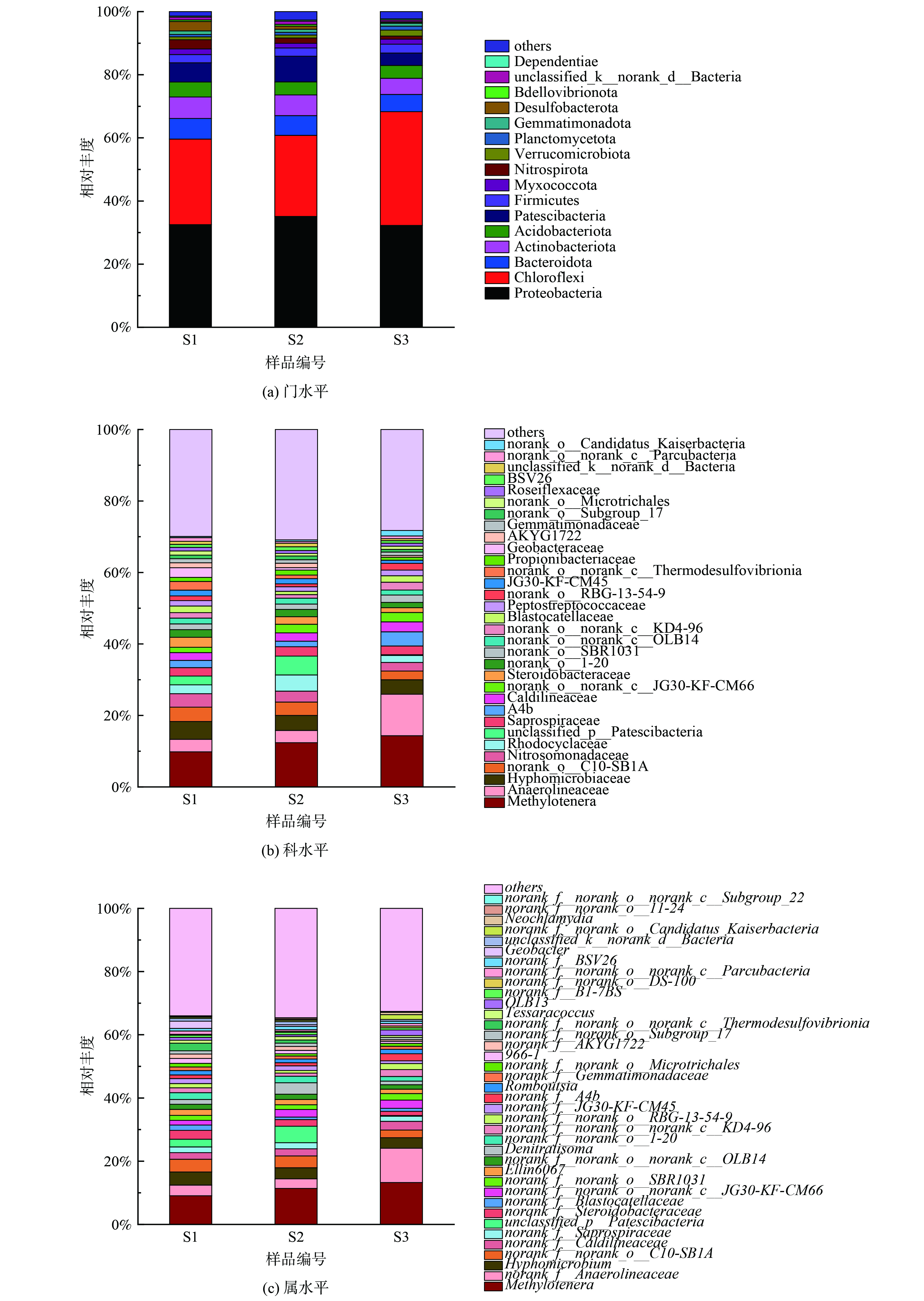
图 7 不同样品微生物门水平(a)、科水平(b)和属水平(c)群落结构
Figure 7. Microbial phylum level (a), family level (b) and genus level (c) in different samples

图 8 不同分子量PLA影响的主要菌群门水平(a)、科水平(b)和属水平(c)
Figure 8. Major microbial phylum (a), family level (b) and genus level (c) affected by PLA of different molecular weights
表 1 微量元素溶液成分组成
Table 1. Composition of trace element solution
成分 质量浓度/(g·L−1) 成分 质量浓度/(g·L−1) ZnSO4·7H2O 2.2 CuSO4·5H2O 1.6 CoCl2·6H2O 1.6 MgSO4·7H2O 5.0 FeSO4·7H2O 5.0 (NH4)6Mo7O24·4H2O 1.1 CaCl2·2H2O 5.5 NiSO4·6H2O 0.42 MnCl2·4H2O 5.0 EDTA 5.0
下载: 导出CSV
表 2 微生物群落α多样性指数
Table 2. α diversity index of microbial community
样品编号 序列数目 OTUs Shannon指数 Chao指数 coverage S1 43 896 1 205 5.50 1 375.29 0.99 S2 36 628 1 273 5.47 1 402.56 0.99 S3 40 559 1 172 5.41 1 298.73 0.99 注:5 000 g·mol−1分子量PLA反应器37 d污泥样品S1;10 000 g·mol−1分子量PLA反应器37 d污泥样品S2;50 000 g·mol−1分子量PLA反应器37 d污泥样品S3。
下载: 导出CSV
-
[1] XU Z S, CHAI X L. Effect of weight ratios of PHBV/PLA polymer blends on nitrate removal efficiency and microbial community during solid-phase denitrification[J]. International Biodeterioration & Biodegradation, 2017, 116: 175-183. [2] JIANG L, WU A Q, FANG D X, et al. Denitrification performance and microbial diversity using starch-polycaprolactone blends as external solid carbon source and biofilm carriers for advanced treatment[J]. Cheosphere, 2020, 255: 126901. doi: 10.1016/j.chemosphere.2020.126901 [3] 芦婷, 杨璐华, 杨飞飞, 等. 高效反硝化菌强化固相碳源生物脱氮特性研究[J]. 北京大学学报(自然科学版), 2017, 53(5): 957-963. doi: 10.13209/j.0479-8023.2017.114 [4] 陕洁, 牟芬, 王新芳, 等. 可生物降解塑料的合成及其改性[J]. 聚酯工业, 2021, 34(3): 23-26. doi: 10.3969/j.issn.1008-8261.2021.03.006 [5] 李静, 刘景江. 聚(β-羟基丁酸酯)和β-羟基丁酸酯-β-羟基戊酸酯共聚物与可生物降解高分子共混改性研究进展[J]. 高分子通报, 2003(6): 33-43. doi: 10.3969/j.issn.1003-3726.2003.06.006 [6] 林世东, 杜国强, 顾君, 等. 我国生物基及可降解塑料发展研究[J]. 塑料工业, 2021, 49(3): 10-12+37. doi: 10.3969/j.issn.1005-5770.2021.03.002 [7] PANG X, ZHUANG X L, TANG Z H, et al. Polylactic acid (PLA): research, development and industrialization[J]. Biotechnology Journal, 2010, 5(11): 1125-1136. doi: 10.1002/biot.201000135 [8] FAN Z X, HU J, WANG J L. Biological nitrate removal using wheat straw and PLA as substrate[J]. Environmental Technology, 2012, 33(21): 2369-2374. doi: 10.1080/09593330.2012.669411 [9] 彭书林, 赵丹. 进水pH值对PLA反硝化系统的影响[J]. 塑料工业, 2021, 49(10): 158-162. doi: 10.3969/j.issn.1005-5770.2021.10.033 [10] TAKAHASHI M, YAMADA T, TANNO M, et al. Nitrate removal efficiency and bacterial community dynamics in denitrification processes using poly (l-lactic acid) as the solid substrate[J]. Microbes and Environments, 2011, 26(3): 212-219. doi: 10.1264/jsme2.ME11107 [11] 许明奕, 逄宇帆, 刑涛等. 聚乳酸合成方法的研究进展及市场分析[J]. 应用化工, 2022, 51(12): 3614-3618+3624. doi: 10.3969/j.issn.1671-3206.2022.12.038 [12] 夏璐, 黄鹏, 刘丹丹等. 磷钨杂多酸直接法催化合成聚乳酸[J]. 当代化工, 2010, 39(06): 628-631. doi: 10.3969/j.issn.1671-0460.2010.06.005 [13] 刘斌基. 聚乳酸的合成过程的影响因素[J]. 新疆有色金属, 2015, 38(06): 49-51. doi: 10.16206/j.cnki.65-1136/tg.2015.06.018 [14] 杨惠兰, 张丹, 兰书焕, 等. 聚己内酯复合固体碳源的制备及其深度脱氮性能研究[J]. 环境科学学报, 2022, 42(05): 263-273. doi: 10.13671/j.hjkxxb.2021.0415 [15] GAO L J, HAN F, ZHANG X W, et al. Simultaneous nitrate and dissolved organic matter removal from wastewater treatment plant effluent in a solid-phase denitrification biofilm reactor[J]. Bioresource Technology, 2020, 314: 123714. doi: 10.1016/j.biortech.2020.123714 [16] 姚璐璐, 涂响, 于会彬等. 三维荧光区域积分评估城市污水中溶解性有机物去除[J]. 环境工程学报, 2013, 7(02): 411-416. [17] 陈思. 生物脱氮固体碳源筛选及初步脱氮效果[D]. 重庆: 重庆大学, 2013. [18] 汪鲁, 刘军, 李永富, 等. 基于固态碳源的同步硝化反硝化反应器对海水养殖废水中氮的去除性能[J]. 环境工程学报, 2022, 16(6): 2001-2009. doi: 10.12030/j.cjee.202112196 [19] YANG M, WANG X N, LIU S, et al. Carbon release behaviour of polylactic acid/starch-based solid carbon and its influence on biodenitrification[J]. Biochemical Engineering Journal, 2020, 155. [20] 王玥, 秦帆, 唐燕华, 等. 农业废弃物作为反硝化脱氮外加碳源的研究[J]. 林业工程学报, 2019, 4(5): 146-151. [21] 孙策, 吕闪闪, 张化腾等. 聚乳酸及其复合材料降解的研究进展[J]. 塑料, 2018, 47(6): 114-117. [22] 任永琳, 王达, 刘合等. 聚乳酸水解机理及水解性能改进方法研究进展[J]. 石油化工, 2022, 51(09): 1129-1136. doi: 10.3969/j.issn.1000-8144.2022.09.018 [23] 焦旗, 田广华, 杨坚, 等. 聚丙烯结晶性能研究[J]. 工程塑料应用, 2015, 43(7): 109-113. doi: 10.3969/j.issn.1001-3539.2015.07.024 [24] 樊新, 陈剑, 阮建明, 等. 聚乳酸类生物可降解材料研究进展[J]. 粉末冶金材料科学与工程, 2008(4): 187-194. doi: 10.3969/j.issn.1673-0224.2008.04.001 [25] 梁捷, 缪恒锋, 任洪艳, 等. 以聚己内酯作为生物反硝化固体碳源的研究[J]. 环境工程学报, 2015, 9(2): 633-638. doi: 10.12030/j.cjee.20150221 [26] SHEN Z Q, ZHOU Y X, WANG J L. Comparison of denitrification performance and microbial diversity using starch/polylactic acid blends and ethanol as electron donor for nitrate removal[J]. Bioresource Technology, 2013, 131: 33-39. doi: 10.1016/j.biortech.2012.12.169 [27] VAN DEN BERG E M, BOLEIJ M, KUENEN J G, et al. DNRA and denitrification coexist over a broad range of acetate/N-NO(3)(-) ratios, in a chemostat enrichment culture[J]. Frontiers in Microbiology, 2016, 7: 1842. [28] HU R T, ZHENG X L, ZHENG T Y, et al. Effects of carbon availability in a woody carbon source on its nitrate removal behavior in solid-phase denitrification[J]. Journal of Environmental Management, 2019, 246: 832-839. [29] YANG Z C, SUN H M, ZHOU Q, et al. Nitrogen removal performance in pilot-scale solid-phase denitrification systems using novel biodegradable blends for treatment of waste water treatment plants effluent[J]. Bioresource Technology, 2020, 305: 122994. doi: 10.1016/j.biortech.2020.122994 [30] LI J, TABASSUM S. Remediation of nitrate-contaminated groundwater by a combined treatment method of novel Mass Bio System and solid organic carbon sources: In-depth study[J]. Cleaner Engineering and Technology, 2021, 4. [31] WU L N, ZHANG L Y, XU Y Y, et al. Advanced nitrogen removal using bio-refractory organics as carbon source for biological treatment of landfill leachate[J]. Separation and Purification Technology, 2016, 170: 306-313. doi: 10.1016/j.seppur.2016.06.033 [32] 万鹏亮, 刘玉玲, 朱妮平, 等. A2O工艺处理城市污水过程中DOM组分变化分析[J]. 西安建筑科技大学学报(自然科学版), 2021, 53(5): 765-772. doi: 10.15986/j.1006-7930.2021.05.020 [33] CHEN D, WANG H Y, YANG K, et al. Performance and microbial communities in a combined bioelectrochemical and sulfur autotrophic denitrification system at low temperature[J]. Chemosphere, 2018, 193: 337-342. doi: 10.1016/j.chemosphere.2017.11.017 [34] ZHAO J M, FENG C P, TONG S, et al. Denitrification behavior and microbial community spatial distribution inside woodchip-based solid-phase denitrification (W-SPD) bioreactor for nitrate-contaminated water treatment[J]. Bioresource Technology, 2018, 249: 869-879. doi: 10.1016/j.biortech.2017.11.011 [35] MIURA Y, WATANABE Y, OKABE S, et al. Significance of Chloroflexi in performance of submerged membrane bioreactors (MBR) treating municipal wastewater[J]. Environmental Science & Technology, 2007, 41: 7787-7794. [36] FUJII N, KURODA K, NARIHIRO T, et al. Metabolic potential of the superphylum Patescibacteria reconstructed from activated sludge samples from a municipal wastewater treatment plant[J]. Microbes and Environments, 2022, 37 (3). [37] YE L, SHAO M F, ZHANG T, et al. Analysis of the bacterial community in a laboratory-scale nitrification reactor and a wastewater treatment plant by 454-pyrosequencing[J]. Water Research, 2011, 45(15): 4390-4398. doi: 10.1016/j.watres.2011.05.028 [38] VETROVSKY T, STEFFEN K T, BALDRIAN P. Potential of cometabolic transformation of polysaccharides and lignin in lignocellulose by soil Actinobacteria[J]. PLoS One, 2014, 9(2): 89108. doi: 10.1371/journal.pone.0089108 [39] 周星煜, 张金松. 活性污泥反硝化速率与功能基因活性关联分析[J/OL]. 给水排水: 1-7 2-11-24].
[40] KRISTENSEN J M, SINGLETON C, CLEGG L A, et al. High diversity and functional potential of undescribed "Acidobacteriota" in Danish wastewater treatment plants[J]. Frontiers in Microbiology, 2021, 12: 643950. doi: 10.3389/fmicb.2021.643950 [41] 肖晶晶, 郭萍, 霍炜洁, 等. 反硝化微生物在污水脱氮中的研究及应用进展[J]. 环境科学与技术, 2009, 32(12): 97-102. doi: 10.3969/j.issn.1003-6504.2009.12.022 [42] CAO J S, ZHANG T, WU Y, et al. Correlations of nitrogen removal and core functional genera in full-scale wastewater treatment plants: influences of different treatment processes and influent characteristics[J]. Bioresource Technology, 2020, 297: 122455. doi: 10.1016/j.biortech.2019.122455 [43] RISSANEN A J, OJALA A, FRED T, et al. Methylophilaceae and Hyphomicrobium as target taxonomic groups in monitoring the function of methanol-fed denitrification biofilters in municipal wastewater treatment plants[J]. J Ind Microbiol Biotechnol, 2017, 44(1): 35-47. doi: 10.1007/s10295-016-1860-5 [44] WANG Z Y, ZHENG M, MENG J, et al. Robust nitritation sustained by acid-tolerant ammonia-oxidizing bacteria[J]. Environmental Science & Technology, 2021, 55(3): 2048-2056. [45] KONDROTAITE Z, VALK L C, PETRIGLIERI F, et al. Diversity and ecophysiology of the genus OLB8 and other abundant uncultured Saprospiraceae genera in global wastewater treatment systems[J]. Frontiers in Microbiology, 2022, 13: 917553. doi: 10.3389/fmicb.2022.917553 [46] ZHANG S Q, KONG Z, WANG H, et al. Enhanced nitrate removal by biochar supported nano zero-valent iron (nZVI) at biocathode in bioelectrochemical system (BES)[J]. Chemical Engineering Journal, 2022, 433. [47] SUN H M, ZHOU Q, ZHAO L, et al. Enhanced simultaneous removal of nitrate and phosphate using novel solid carbon source/zero-valent iron composite[J]. Journal of Cleaner Production, 2021, 289. [48] 张鹏程, 李晓玲, 王晓婷等. 活性污泥体系中C/N/S对硝酸盐还原过程的影响[J]. 中国环境科学, 2021, 41(5): 2117-2122. doi: 10.3969/j.issn.1000-6923.2021.05.015 [49] MARTINEAU C, MAUFFREY F, VILLEMUR R. Comparative analysis of denitrifying activities of Hyphomicrobium nitrativorans, Hyphomicrobium denitrificans, and Hyphomicrobium zavarzinii[J]. Applied and Environmental Microbiology, 2015, 81(15): 5003-5014. doi: 10.1128/AEM.00848-15 [50] ZHANG Y P, DOUGLAS G B, KAKSONEN A H, et al. Microbial reduction of nitrate in the presence of zero-valent iron[J]. Science of the Total Environment, 2019, 646: 1195-1203. doi: 10.1016/j.scitotenv.2018.07.112 [51] JIA L X, ZHOU Q, LI Y, et al. Integrated treatment of suburb diffuse pollution using large-scale multistage constructed wetlands based on novel solid carbon: nutrients removal and microbial interactions[J]. Journal of Environmental Management, 2023, 326: 116709. doi: 10.1016/j.jenvman.2022.116709 [52] LEVY-BOOTH D J, NAVAS L E, FETHEROLF M M, et al. Discovery of lignin-transforming bacteria and enzymes in thermophilic environments using stable isotope probing[J]. The ISME Journal, 2022, 16(8): 1944-1956. doi: 10.1038/s41396-022-01241-8 [53] SUOMINEN S, VAN VLIET D M, SANCHEZ-ANDREA I, et al. Organic matter type defines the composition of active microbial communities originating from anoxic baltic sea sediments[J]. Frontiers in Microbiology, 2021, 12: 628301. doi: 10.3389/fmicb.2021.628301 [54] ZHANG L, HAO S W, WANG Y P, et al. Rapid start-up strategy of partial denitrification and microbially driven mechanism of nitrite accumulation mediated by dissolved organic matter[J]. Bioresource Technology, 2021, 340: 125663. doi: 10.1016/j.biortech.2021.125663 [55] HUANG X, YAO K, YU J H, et al. Nitrogen removal performance and microbial characteristics during simultaneous chemical phosphorus removal process using Fe3+[J]. Bioresource Technology, 2022, 363: 127972. doi: 10.1016/j.biortech.2022.127972 [56] YIN Y N, HU Y M, WANG J L. Co-fermentation of sewage sludge and lignocellulosic biomass for production of medium-chain fatty acids[J]. Bioresource Technology, 2022, 361: 127665. doi: 10.1016/j.biortech.2022.127665 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4264
- HTML全文浏览数: 4264
- PDF下载数: 120
- 施引文献: 0
