

-
河流生态系统是连接陆地与海洋的纽带,也是重要的氮汇和碳汇,对陆地生态系统输出营养盐和有机质的迁移转化过程起主要作用。而沉积物作为河流生态系统的重要组成部分,不仅是流域C、N、P等营养元素的重要蓄积库,也是许多环境污染物的接收器[1]。输入河流中的 NH4+ 和NO3−发生着以各种微生物为媒介的硝化 (nitration) 、反硝化 (denitrification) 、异化还原成铵 (dissimilatory nitrate reduction to ammonium,DNRA) 和厌氧氨氧化 (anaerobic ammonium oxidation,ANAMMOX) 等活跃的生物地球化学反应,使河流中的氮元素以氮气形式返回大气圈,以达到河流中除氮和减少温室气体氧化亚氮 (N2O) 排放的目的[2-4]。甲烷 (CH4) 对全球变暖的贡献占22%,是除了N2O和CO2气体外最重要的温室气体[5-7]。河流也是大气中CH4主要排放源之一。目前,大气中有50%CH4来自于河流、河口等水生生态系统[8-10]。在水环境中广泛存在着厌氧甲烷氧化 (anaerobic oxidation to methane,AOM) ,而反硝化型厌氧甲烷氧化 (denitrifying anaerobic methane oxidation,DAMO) 能在厌氧条件下使甲烷直接作为碳源,以硝酸盐作为电子供体发生脱氮反应。该反应能减少甲烷排放的同时,还能达到除氮目的,因此,被认为是淡水环境中连接碳和氮循环的新型微生物过程,对于厌氧环境中甲烷消除具有重要意义[7,11-14]。国内外研究学者针对湖泊[12,15]、湿地[16-17]及河流[13,18]等多地淡水环境开展了相关研究,但针对易受农业径流污染的河流及河口环境研究较少[11,13,15-18]。城市内陆河流营养盐及电子供体较为丰富、有机质和硝酸盐含量较高、氧化还原电位梯度频繁交迭,为各种以硝酸盐为电子受体的生物化学反应提供了最大反应驱动力,促进了碳氮循环。
西溪为厦门市最大的内陆入海河流。该河流及河口环境的厌氧和好氧交界面发生着多种生物地球化学反应[12,19-20]。西溪所在流域存在较多人类活动,不同地区受人类影响的程度不同。因此,针对西溪中脱氮反应速率及连接碳氮循环的反硝化型厌氧甲烷氧化反应速率研究是很有必要。
本研究选取西溪中下游为研究对象,在2021—2022年期间沿溪流进行季节性采样。基于15N以及13C同位素配对技术,开展沉积物培养实验,评估西溪流域中下游沉积物季节性的反硝化和厌氧氨氧化潜在速率及脱氮贡献,同时也验证反硝化型厌氧甲烷氧化反应的发生并测定其反应速率,并结合上覆水等理化特征分析,探讨了沉积物中生物化学反应潜在速率时空及季节性差异的原因,以掌握河流内部反硝化、厌氧氨氧化作用及反硝化型厌氧甲烷氧化 (DAMO) 的变化规律,从而揭示生态环境中河流生态系统在甲烷参与下碳、氮的动态变化过程,以期为探究该河流中碳氮循环并开展下游沉积物脱氮治理提供参考。
-
西溪起源于同安区西北部的寨尖尾山,全长大约34 km,流域面积约为494 km2,多年平均年径流量约为4.66×109 m3,其东南方向流经东咀港入海。西溪流域地处南亚热带海洋性季风气候区。该区域内气候温和、雨量充沛、热量充足,且河流沿途有莲花溪、澳溪和汀溪等水流汇入,故流域境内主要水系多呈树枝状,有丰富的农林牧渔水利气候资源,是同安区主要工农业、生活用水的来源。西溪中下游流经农田村庄,沿途穿过城市居民区域,且末端接近入海口,存在工农业含氮污水、沿岸生活污水的流入及较为复杂的河口环境,可能会对河流中的碳氮循环过程起到负面作用,其周围环境复杂,对河流生态系统中的碳氮循环过程亦有重要影响。
-
考虑到河流流域周围环境的复杂性及不同时间尺度的可对比性,分别在2021—2022年对西溪中下游河流进行了季节性样品采集。从上游到下游共布置了5个采样点 (图1) ,分别从农业区、城市居民生活区,再到入海口地区。每一个采样点采集0~10 cm 的沉积物混合样 500 g 装入自封袋,并采集相应上覆水样,现场测定溶解氧 (DO) 、电导率、pH及温度等指标。
-
潜在的反硝化和厌氧氨氧化速率利用15N 同位素配对法测定。分别取2组沉积物加入厌氧瓶中并加入200 mL上覆水,将泥样与水样中气体用高纯氦气吹扫干净后密封。通过注射器向2组培养瓶内分别添加 100 μm 和 200 μm 的 K15NO3 溶液。反应稳定一段时间后,保持采样当天的流速与温度,在黑暗条件下培养72 h 。当反应过程结束后,在厌氧手套箱中用引流法将培养好的上覆水样注入经氦气吹扫过的顶空瓶中,并添加饱和 ZnCl2溶液终止反应,使用GasBench-IRMS在线连续分析氮气 (28N、29N、30N) 同位素后进行速率计算[21-25]。计算公式见式 (1)~(5) 。
式中:
V 是不同浓度体系添加15NO3− 浓度比值;r14 为还原体系中14NO3−/15NO3−比值;式(1)和式(2)分别为加入100 μmol·L−1 和 200 μmol·L−1的 K15NO3培养体系;P29N2 和P30N2 分别为同位素的 29N2和30N2的生成量;P14 为总N2实际的生成量;P14,A 为厌氧氨氧化所产生28N2的量;P14,D 为反硝化所产生28N2的量。利用13C 同位素配对法测定潜在的反硝化型厌氧甲烷氧化速率。将沉积物泥浆在厌氧条件下至少培养2~3 d,尽可能去除残留的氧气。随后将泥浆分为2组:1) 13CH4;2) 13CH4+NO3− (处理组) 。在培养瓶中加50 g沉积物并加入50 mL高纯水以确保土样处于饱和状态,再将此混合物转移到 500 mL培养瓶中,采用高纯氦气进行吹扫,拧紧盖子密封;再用注射器通过厌氧培养瓶顶端加入KNO3 溶液 (处理组) 和高纯水 (对照组) ,稳定一段时间后,通过向培养瓶中注入13CH4气体。保持采样当天的流速及温度,避光在恒温培养箱中进行培养。分别在培养12 h与48 h进行破坏性采样,用注射器抽取培养瓶顶部空间气体,注入铝箔气体袋中,随后使用DeltaV Advantage同位素质谱仪测定培养瓶中产生的13CO2丰度值。培养瓶顶部空间气体中产生的13CO2浓度计算参考GUPTA等[26-30]的算法。首先根据测定结果按照式 (6) 计算出13C的原子分数(AT%),再根据等式 (7) 计算13CO2浓度值。
式中:
δ13CPDB 指将样品的13C稳定同位素值(δ 13C)进行国际标准PDB处理的结果;Rstandard 为所采用的标准物质的同位素比值,取值为0.011 237 2;AT% 指13C原子分数;[13CO2]指13CO2浓度值,mmol·L−1;[CO2] 指CO2浓度值,mmol·L−1。 -
采样点上覆水及沉积物孔隙水的理化特征见表1所示。西溪中下游上覆水中氮的总质量浓度 (TN) 为3.02~5.68 mg·L−1,这说明西溪中下游地区水体处于富营养水平。研究区域内无机氮的赋存形态以硝氮和氨氮为主,但不同采样点之间无机氮的主要赋存形态不相同,这与水体中发生的生物化学反应强弱及速率快慢有关。其中,上覆水中硝氮质量浓度最大值为X2点的1.585 mg·L−1,平均值为1.200 mg·L−1;氨氮质量浓度最大值为X4点的2.391 mg·L−1,平均值为0.630 mg·L−1;总有机碳的质量浓度为7.56~16.29 mg·L−1,平均值为9.84 mg·L−1。沉积物孔隙水中主要氮素仍以硝氮和氨氮为主,其中硝氮质量浓度最大值为X2点1.14 mg·L−1 ,平均值为0.88 mg·L−1;氨氮质量浓度最大值为X4点3.80 mg·L−1,平均值为0.96 mg·L−1,总有机碳的质量浓度为3.00~5.68 mg·L−1,平均值为4.22 mg·L−1。研究区域内不同采样点沉积物孔隙水氨氮质量浓度明显高于上覆水氨氮,总有机碳的质量浓度孔隙水低于上覆水。表2表明,沉积物中氮素主要以硝氮和氨氮形式存在,除X4点外,硝氮质量浓度均大于氨氮。其中,硝氮质量浓度最大值在X2点,为10.58 mg·kg−1,平均值为8.62 mg·kg−1;氨氮质量浓度最大值在X4点,为15.81 mg·kg−1,平均值为7.09 mg·kg−1;西溪中下游全氮的质量浓度为2 717~5 376 mg·kg−1,平均值为3 962 mg·kg−1。
-
从空间分布趋势来看,上覆水中总氮与氨氮空间变化趋势相同,沿程均呈现逐渐升高的变化趋势;硝氮与亚硝氮空间变化趋势相同,沿程均呈现先升高后降低的变化趋势 (图2) 。上覆水及沉积物氮素指标中X4点相比其他点位氨氮质量浓度偏高而硝酸盐质量浓度较低,氨氮作为水体硝化过程的反应底物会收到硝化作用及沉积物吸附的影响。结合沉积物氮素含量变化趋势 (图3) 可知,X4点位沉积物中氨氮质量浓度为15.81 mg·kg−1。上覆水中硝氮质量浓度在X2点最高后又沿程降低。西溪中下游污染源输入及水体的自净过程对硝氮沿程变化起主要作用。
从时间变化上看,上覆水及沉积物中总氮、硝氮及亚硝氮等指标的变化呈冬季>秋季>春季>夏季的变化趋势。由于春季属于丰水期,河流氮循环加快,而秋冬季节属于枯水期,河流水量减少,氮循环过程减缓导致氮素浓度升高。上覆水及沉积物中氨氮质量浓度变化呈夏季>冬季>秋季>春季的变化趋势,在夏季呈明显升高的现象。在夏季采样过程中发现,夏季温度较高导致河水蒸发量较,水体呈现大面积干涸,河岸以及水体中漂浮了大量水生动物尸体。有研究表明,水生动植物腐烂会首先进行氨化反应,即水体中的微生物分解有机氮转化为氨氮的过程。这一环境因素对水体中氨氮的升高起到主要作用,也致使许多水生动植物死亡腐烂,水中的氨氮质量浓度升高[31]。水体经过高温日晒水体温度会升高,并导致相关菌活性受抑制,从而使氨氮的迁移转化效率下降。同时,有研究表明氨氮质量浓度过高时,水体中游离氨 (free ammonia,FA) 会对水体中的硝化菌产生抑制作用。这就使得水体中的氨氮不能在硝化菌的作用下转化为硝氮,从而致使X4点硝酸盐或亚硝酸盐的质量浓度降低[32-33]。
西溪中下游水体中TOC为3.563~14.38 mg·L−1,平均质量浓度为8.52 mg·L−1;TIC为10.06~58.396 mg·L−1,平均质量浓度为28.11 mg·L−1 (图4) 。水体中的TOC是外源输入、内源产生及河流内迁移转化等多个过程动态平衡的结果。从空间分布来看,各碳素分布呈现明显沿程增加的变化趋势,最大浓度在X4点。从时间分布来看,TOC与TIC呈秋冬季节较高,相对较低的是春季。
以上水质检测结果表明,上覆水及沉积物中硝氮质量浓度较高。因此,针对自然水体硝氮的15N、18O进行相关丰度测定,再结合不同来源硝酸盐δ15N-NO3−及δ18OV-SMOW典型值域分布图[34],这表明水体中硝酸盐氮的主要污染来源。另外,西溪中下游水样δ15N-NO3−为10.84‰~67.23‰,δ18OV-SMOW为1.69‰~24.90‰,这表明西溪中下游污染物主要来源于有机肥和污水。
-
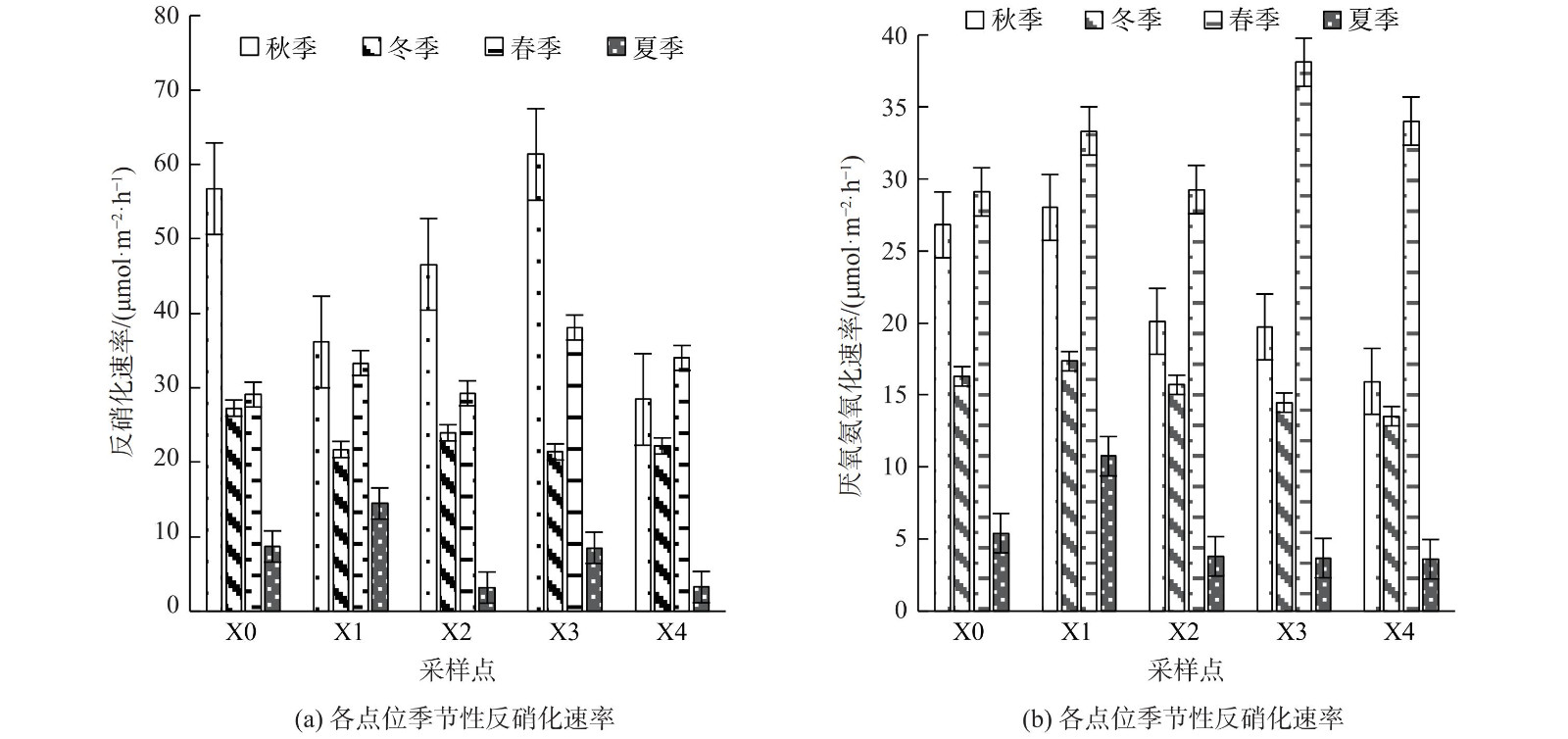
西溪中下游沉积物的反硝化速率为3.233~61.341 μmol·m−2·h−1,最大反应速率为秋季X3点,平均速率为27.386 μmol·m−2·h−1,脱氮贡献率为56.30%~66.76%,在空间上呈现先升高后降低的变化趋势,在时间上呈现秋季>春季>冬季>夏季的变化趋势。厌氧氨氧化速率为3.604~38.101 μmol·m−2·h−1,最大反应速率为春季X3点,平均反应速率为18.953 μmol·m−2·h−1,脱氮贡献率的变化范围为33.24%~43.70%,从沿程呈现逐渐降低的空间变化趋势,在时间变化上厌氧氨氧化潜在反应速率呈现春季>秋季>冬季>夏季的变化趋势 (图5) 。
-
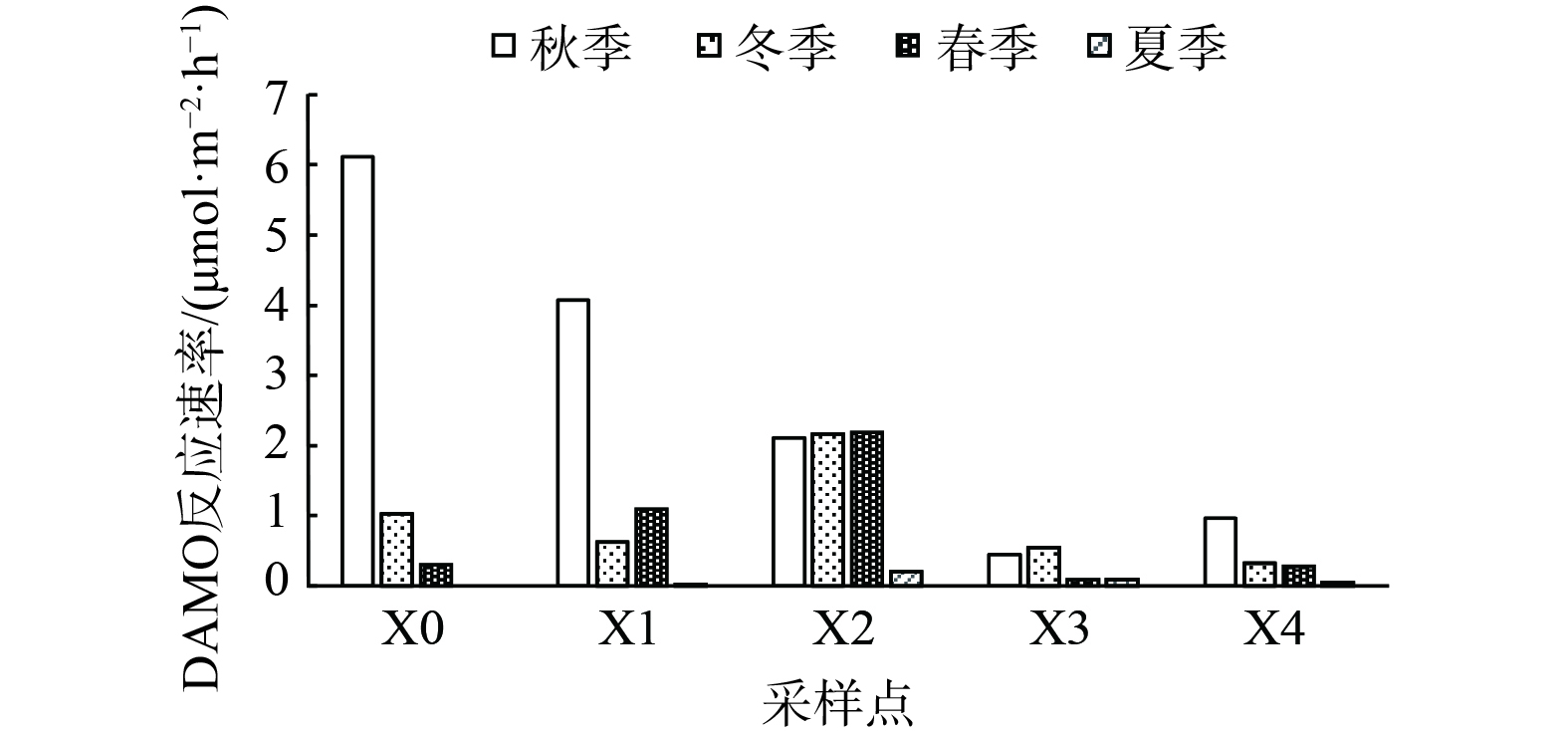
沉积物样品对照组与处理组的顶部空间气体中均检测产生13CO2,且二氧化碳质量浓度随培养时间呈上升的变化趋势,因此,各采样点的沉积物样品均可在厌氧条件下将甲烷氧化转化成为二氧化碳。这也说明西溪中下游沉积物中均有DAMO反应发生。西溪中下游沉积物中DAMO反应速率为0~6.122 µmol·m−2·h−1,平均速率为1.139 µmol·m−2·h−1,最大反应速率点在秋季的X0点,最低反应速率在夏季的X0点。从空间上看,西溪中下游各采样点在空间特征上变化具有较大差异性。从时间上看,西溪中下游各采样点的DAMO速率秋季最强,春季和冬季次之,夏季反应速率最低 (图6) 。
-
由于营养物可用性和环境条件的差异,各种沉积物中生物化学反应的潜在反应速率会受到多种因素及其之间相互作用的影响。溶解氧对厌氧氨氧化产生抑制作用,且厌氧氨氧化相比于反硝化对低氧环境的依赖性更强[35]。西溪中下游河流中有机质的输入也为反硝化细菌提供了充足碳源,厌氧氨氧化细菌在针对氨氮及亚硝氮的竞争中处于劣势,导致西溪中下游河流沉积物中脱氮以反硝化反应为主[36]。值得注意的是,X4点氨氮含量较高,但同时厌氧氨氧化的反应速率明显较低。这是由于该点氨氮含量过高会对硝化细菌及亚硝化菌产生毒害作用,致使没有足够的硝酸盐或亚硝酸盐作为电子受体进行厌氧氨氧化反应[21]。适宜的硝酸盐底物质量浓度对反硝化速率的起到促进作用,结合上覆水中硝酸盐质量浓度可知,在流经城区的X2与X3点,硝酸盐质量浓度高于其他采样点,故反硝化速率也较高[37]。在季节上呈现厌氧氨氧化与反硝化速率春秋两季大于冬季的变化趋势[38]。受营养盐及温度的双重控制,虽然冬季处于枯水期,导致河流断流,致使河流沉积物中没有充足硝酸盐作为反硝化反应的底物,水体氮循环减弱,导致反硝化速率较低[39-40]。针对淡水河流中的脱氮研究中反硝化脱氮贡献率均大于厌氧氨氧化脱氮贡献率,西溪中下游沉积物中的反硝化速率与潮白河 (25.82±0.98)~(142.32±17.06) μmol·m−2·h−1,以及太湖地区西部6条河流(18.5±2.8)~(133.2±27.1) μmol·m−2·h−1相当[41-43]。
DAMO反应过程会受到多种因素及其相互作用的干扰。河流周围不同的环境条件与河流系统中氮素与碳素含量有着密切关系,从而也会主要影响DAMO速率的快慢 [44-46]。其中,X0和X1点氮肥的冲刷及生活污水的排放为河道带来大量氮,王超等[45]发现硝酸盐可用性可提高反硝化程度,能进一步增加DAMO速率。同时,本研究经相关性分析发现冬季与春季硝酸盐质量浓度与DAMO反应速率相关性为0.977 5与0.959,在0.01级别 (双尾) 呈现显著相关性。值得注意的是,除秋季以外,靠近入海口的X4点DAMO反应速率较低而盐度与沉积物中DAMO细菌的丰度及活性呈负相关,会抑制DAMO的速率[47]。秋季X4点接近入海口但反应速率较高,这可能是由于碳素的累积效应所造成的,也有研究表明TOC对DAMO菌群呈显著正相关[9][17]。从时间上看,秋季DAMO速率较大,由于适宜的环境温度能对DAMO的相关微生物生长代谢起到促进作用,长时间更适宜厌氧甲烷氧化菌参与反应,进而也会对DAMO的反应速率起到提升作用,从而促进DAMO的反应过程[48-49]。
厌氧氨氧化反应能以硝酸盐或者亚硝酸盐为电子受体将氨氮转化为氮气,DAMO反应能利用NO3−作为电子供体将甲烷氧化,最终产生二氧化碳与氮气。两种反应均可实现减少水体氮素污染及甲烷排放的目的。厌氧氨氧化与反硝化过程存在竞争,且DAMO与反硝化过程存在一定协同作用。当反硝化反应速率较高时,则厌氧氨氧化速率相应较低。这是由于硝酸盐转化为亚硝酸盐含量的多少及反应竞争强度所决定的。当反硝化过程较强,能将硝酸盐转化为亚硝酸盐进而转化为N2O,因此,反应体系中所含的亚硝酸盐就会被消耗进而会使厌氧氨氧化速率较低。DAMO反应能利用硝酸盐作为电子受体将甲烷氧化成二氧化碳,并在此过程中完成脱氮。因此,反硝化过程所产生的电子受体能促进甲烷氧化,2种生物化学反应之间存在一定的协同作用。由图4与图5可知,反硝化与厌氧氨氧化反应速率在空间上变化趋势相反,而反硝化与DAMO反应速率在空间变化上趋势相同。3种反应相互协同为西溪中下游沉积物脱氮及碳减排做出重要贡献,因此,3种生物化学反应之间存在碳氮交互过程,并且硝酸盐对城市内陆河流沉积物中发生的生物化学反应起到重要作用,因此,进一步研究硝酸盐质量浓度变化对三者反应速率的影响,以及脱氮减排的作用途径至关重要。
城市内陆河流的环境复杂容易受到污染源干扰,为保证城市水生态环境的健康可持续发展,应加大对城市内陆河流水质管控。根据本研究结果,河流沉积物中的反硝化、厌氧氨氧化、DAMO对河流中的脱氮,以及甲烷氧化方面都起着一定的作用,且厌氧氨氧化过程最终产物N2相较于反硝化的产物N2O,降低了温室气体排放的可能性,甲烷转化成二氧化碳排放也降低了温室气体的暖化能力,间接地有助于碳减排。因此,一方面可以对河流中的底泥进行修复,去除河流沉积物中的污染物质减少河流中污染源的释放,从而改善河流水质;另一方面可向受污染水体中投加厌氧氨氧化菌等能提高污染物代谢能力的高效菌种微生物,利用微生物的分解脱氮能力降低水质的污染程度,还可投加促进功能菌微生物生长和提高微生物活性的物质,从而加快污染物的降解和转化,提高沉积物中的脱氮速率。
-
1) 各形态氮及碳素在空间上呈逐渐累积的变化趋势,总氮变化范围为3.02~5.68 mg·L−1,上覆水体中硝氮的主要污染来源为有机肥和污水。西溪中下游沉积物中厌氧氨氧化反应平均脱氮贡献率为40.13%,反硝化平均脱氮贡献率为59.87%,溪流沉积物中脱氮主要是以反硝化作用进行的,但同时厌氧氨氧化也发挥着不小的作用;西溪中下游沉积物中发生着DAMO反应,但不同的点位存在差异性且平均反应速率为1.139 µmol·m−2·h−1,硝酸盐质量浓度与DAMO反应速率相关性较大。
2) 盐度、硝酸盐负荷及碳源的输入对DAMO过程至关重要,也可能对于甲烷汇及氧化过程起到主导作用,因此,进一步探究内陆淡水河流沉积物中反硝化作用对厌氧甲烷氧化过程影响的方式也具有重要生态意义。
3) 针对城市内陆河流水质管控,可对河流底泥进行修复,减少污染物的释放;沿程向受污染水体中投加反硝化、厌氧氨氧化菌等高效菌种微生物及促进功能菌微生物生长和提高微生物活性的物质,提高水体环境中的脱氮速率,加快污染物的降解和转化。
沉积物碳氮交互效应及其对河流水体脱氮治理的影响——以厦门市西溪中下游为例
Interaction effect of sediment carbon and nitrogen and its influence on nitrogen removal in river water —A case study of the middle and lower reaches of Xixi River in Xiamen
-
摘要: 通过福建厦门市内陆河流西溪中下游上覆水及沉积物中碳氮理化指标的时空分布特征,利用碳 (13C) 、氮 (15N) 稳定同位素技术研究了沉积物中厌氧氨氧化 (ANAMMOX) 、反硝化 (denitrification) 及反硝化型厌氧甲烷氧化 (DAMO) 的反应速率及其分布规律。结果表明:西溪中下游上覆水水质总氮为3.02~5.68mg·L−1;利用15N同位素技术测得西溪中下游沉积物中厌氧氨氧化的平均速率为18.953 μmol·m−2·h−1,平均脱氮贡献率为40.13%,反硝化平均速率为27.386 μmol·m−2·h−1,平均贡献率为59.87%;起主要脱氮贡献的为反硝化反应,但厌氧氨氧化的脱氮作用仍不可忽视。利用13C同位素技术测得西溪中下游沉积物中DAMO反应速率为0~6.122 µmol·m−2·h−1,平均速率为1.139 µmol·m−2·h−1。这表明西溪中下游沉积物中存在DAMO反应,可在厌氧条件下以硝酸盐作为电子受体将CH4转化为CO2。在空间上,西溪中下游DAMO速率存在较大差异性;而在时间上,DAMO速率秋季高于冬春季。反硝化与厌氧氨氧化作用对西溪生态系统中脱氮具有重要意义,DAMO在西溪中下游沉积物虽然反应速率较低,但仍然证明了西溪沉积物生态系统存在连接碳氮循环的微生物反应。反硝化、厌氧氨氧化和DAMO的相互协同,在西溪中下游沉积物脱氮治理及相应过程中的温室气体减排中起到重要作用。
-
关键词:
- 河流 /
- 沉积物 /
- 同位素 /
- 脱氮速率 /
- 反硝化型厌氧甲烷氧化
Abstract: Based on the spatial and temporal distribution characteristics of carbon and nitrogen physical and chemical indexes in overlying water and sediments of the middle and lower reaches of Xixi River in Xiamen City, Fujian Province, the reaction rates and distribution of Anaerobic ammonia oxidation ( Anammox ), Denitrification and denitrifying anaerobic methane oxidation ( DAMO ) in sediments were studied by using carbon ( 13C ) and nitrogen ( 15N ) stable isotope techniques. The results showed that the total nitrogen concentration of overlying water in the middle and lower reaches of Xixi River ranged from 3.02 to 5.68 mg·L−1. The average rate of anaerobic ammonia oxidation in the sediments of the middle and lower reaches of Xixi River was 18.953 μmol·m−2·h−1, and the average contribution rate of denitrification was 40.13 %. The average rate of denitrification was 27.386 μmol·m−2·h−1 and the average contribution rate was 59.87 %. The main contribution of denitrification was denitrification, but the denitrification effect of anaerobic ammonia oxidation could not be ignored. The DAMO reaction rate was 0~6.122 μmol·m−2·h−1 and the average rate was 1.139 μmol·m−2·h−1 measured by 13C isotope technique in the sediments of the middle and lower reaches of Xixi River, indicating that there was DAMO reaction in the sediments of the middle and lower reaches of Xixi River, which could convert CH4 into CO2 with nitrate as electron acceptor under anaerobic conditions. In space, the DAMO rate in the middle and lower reaches of Xixi River was quite different; in terms of time, the DAMO rate in autumn was higher than that in winter and spring. Denitrification and anammox were important for nitrogen removal in Xixi ecosystem. Although the reaction rate of DAMO in the sediments of the middle and lower reaches of Xixi was low, it still proved that there were microbial reactions connecting carbon and nitrogen cycles in Xixi sediment ecosystem. Denitrification, anammox and DAMO synergistically played an important role in sediment nitrogen removal and greenhouse gas emission reduction in the middle and lower reaches of Xixi River.-
Key words:
- river /
- sediment /
- isotope /
- nitrogen removal rate /
- denitrifying anaerobic methane oxidation(DAMO)
-
近年来,随着我国畜禽养殖业的快速发展,养殖废水的不合理排放带来了严重的面源污染[1]。与此同时,养殖废水中磷污染问题也受到了极大关注。一方面,磷是养殖废水中的主要污染物,当排放浓度达到0.02 mg·L−1时,便可引起水体富营养化[2];另一方面,磷矿是一种不可再生资源,对农业生产和磷化学工业具有重要意义[3]。因此,采用高效经济的畜禽养殖废水处理技术使其中的磷元素得以去除的同时实现资源化利用具有重要的现实意义。
目前,常用的除磷技术包括化学沉淀法、生物除磷法、离子交换法以及吸附法[4]。吸附法由于操作简单、成本低廉,尤其是在低磷酸盐浓度条件下吸附效率高,已越来越受到重视[5]。传统的磷吸附剂,如活性炭[6]、海泡石[7]和沸石[8]等,由于处理成本较高,容易产生二次污染或除磷能力有限等问题,在工程化应用中受到限制[9]。在过去的10年中,以果皮、木屑和作物秸秆等天然材料及废弃物材料为基础的生物质吸附剂受到了极大的关注[10]。
生物质材料由于易获取且无二次污染而被认为是一种经济有效且环境友好的吸附剂[11]。其中,海藻能够提供一系列的官能团,包括氨基、羟基、羧基、硫酸盐和咪唑等[12],且作为富营养化的产物,来源广泛,成本低廉。因此,它们可能是一类潜在的生物质吸附材料。但纯海藻类生物质存在导电性差、表面积小、孔隙率低、pH敏感性低等缺点[13],降低了其应用价值。因此,改性是提高其吸附性的必要措施。
镧是一种相对便宜和环境负效应较低的稀土元素[14],能与磷形成稳定沉淀(溶解性产物磷酸镧,pK=26.16)[15]。因此,将镧加载在载体上吸附废水中的磷是一种经济有效的方法。目前,已有研究者采用镧改性膨润土[16]、沸石[17]以及粉煤灰[18]等以增加材料的吸附性能,但采用镧改性海藻等生物质材料用于废水中磷吸附剂的研究还鲜见报道。改性海藻吸附磷酸盐后可直接用作肥料,实现磷元素的资源化再利用,可有效降低吸附工艺产生废物的后续处理费用和对环境的影响。
本研究拟采用镧改性4种海藻作为养殖废水除磷吸附剂,通过SEM和FT-IR等表征手段观察镧改性前、后海藻的表面特征和官能团变化,并探讨在模拟废水中不同吸附剂用量、pH对吸附性能的影响,观察其等温吸附及动力学吸附过程的特点;通过在养猪废水中进行验证,为养殖废水中磷的科学去除和资源化利用提供参考。
1. 材料和方法
1.1 实验材料
本研究采用海带(LJ)、石莼(UL)、红藻(RP)和浒苔(EP)4种海藻作吸附废水中磷的基础材料。实验用海藻均购自青岛海兴源生物科技有限公司。海藻经自来水清洗2次后,再用去离子水润洗5次,然后将其置于60 ℃烘箱中干燥24 h,粉碎研磨,过60目筛后,贮存于聚乙烯瓶中备用。
实验用试剂包括氯化镧(LaCl3·7H2O)、氢氧化钠、磷酸二氢钾、钼酸铵、酒石酸锑氧钾、抗坏血酸等,均为分析纯,购自上海国药集团化学药品有限公司。
在制备模拟废水时,称取一定量的KH2PO4溶解于去离子水中,浓度为25 mg·L−1。实验用养猪废水取自四川省某养殖基地,该废水磷酸根浓度为16.83 mg·L−1,pH为8.32,氨氮浓度为498.65 mg·L−1。
1.2 镧改性海藻的制备
准确称取粉末状的LJ、UL、RP和EP各10 g,放于150 mL烧杯中,与质量分数为1%的氯化镧溶液以1∶10的固液质量比混合;于磁力加热搅拌器上,30 ℃恒温搅拌9 h,调节pH至9,再继续搅拌3 h,离心分离;用去离子水反复冲洗至上清液为中性,于60 ℃烘箱干燥24 h,粉碎后,过60目筛保存备用。改性后的4种海藻经强酸(HNO3、HClO4、HF)消解至澄清,上清液于0.45 μm微孔滤膜过滤,稀释一定倍数后,用ICP发射光谱仪(ICP,Optima 8000,PerkinElmer,美国)测定其镧元素负载量。经测定,La-LJ、La-RP、La-UL和La-EP中镧的负载量分别为2.02%、1.83%、1.79%和1.46%。
1.3 材料表征
4种海藻经镧改性前后的表面形貌用扫描电子显微镜(SEM,Hitachi S4800,日本日立)观察,样品先进行真空喷金,再进行SEM测试。特征官能团采用傅里叶红外光谱仪(FT-IR,Nicolet S10,Thermo Scientific,美国)测定。结构分析采用X射线衍射仪(XRD,D8 ADVANCE,布鲁克公司,德国)测定,用MDI Jade 6.0进行数据分析。
1.4 模拟废水吸附实验
称取一定质量的吸附材料于100 mL锥形瓶中,加入50 mL一定浓度的模拟废液,在25 ℃的振荡器中以120 r·min−1的频率振荡。反应一段时间后,以3 500 r·min−1的频率离心5 min,离心后的上清液经0.45 μm微孔滤膜过滤,稀释一定倍数后,用钼锑抗分光光度法测定上清液中磷酸根的浓度。实验中磷酸根的吸附量按照式(1)进行计算。
Qe=(C0−Ce)Vm (1) 式中:Qe为平衡吸附量,mg·g−1;C0和Ce为溶液在吸附前后的磷酸根浓度,mg·L−1;V为加入废液的体积,L;m为加入吸附剂的质量,g。
1)吸附剂用量实验。各吸附剂用量分别为0.02、0.05、0.1、0.15、0.2和0.25 g,模拟废液pH为6,浓度为25 mg·L−1,吸附时间为2 h。
2)初始pH实验。各吸附剂用量均为0.1 g,用0.2 mol·L−1的NaOH或HCl溶液将模拟废液pH调为3、4、5、6、7和8,浓度为25 mg·L−1,吸附时间为2 h。
3)动力学实验。各吸附剂用量均为0.1 g,模拟废液pH为6,浓度为25 mg·L−1,吸附时间分别为30、60、90、120、150、180、360和540 min。采用准二级动力学模型(式(2))、粒子内扩散模型(式(3))和Elovich模型(式(4))拟合吸附数据,探究吸附动力学过程。
qt=k2q2et1+k2qet (2) qt=kip√t+c (3) qt=(1βs)ln(1+αsβst) (4) 式中:qt为t时刻镧改性的4种海藻对磷的吸附量,mg·g−1;qe为平衡时的吸附量,mg·g−1;k2为准二级吸附速率常数,g·(mg·min)−1;kip为粒子间扩散速率常数,g·(mg·min1/2)−1;c为边界层厚度,mg·g−1;αs为初始吸附速率,mg·(g·min)−1;βs为任意一次实验的解吸常数,g·mg−1。
4)等温吸附实验。各吸附剂用量均为0.1 g,模拟废液pH为6,浓度分别为10、20、40、60、80和100 mg·L−1,吸附时间为2 h。用Langmuir(式(5))和Freundlich(式(6))模型拟合实验数据。
qe=qmkace1+kace (5) qe=kfc1/ne (6) 式中:qe为平衡时的吸附量,mg·g−1;qm为理论最大吸附量,mg·g−1;ka为Langmuir模型常数,L·mg−1;ce为吸附平衡浓度,mg·L−1;kf为Freundlich方程常数;n为常数,1/n表示吸附强度大小。
1.5 养猪废水磷吸附实验
称取4种镧改性的海藻各0.1 g于100 mL锥形瓶,加入经滤膜过滤且pH调为6的养猪废水50 mL(磷酸根浓度16.83 mg·L−1),在25 ℃的振荡器中,以120 r·min−1的转速振荡,反应9 h后,再以3 500 r·min−1的转速离心5 min,离心后的上清液经0.45 μm微孔滤膜过滤,稀释一定倍数后,用钼锑抗分光光度法测定上清液中磷酸根的浓度。
1.6 数据处理
用SPSS Version 19.0(SPSS Inc.,芝加哥,伊利诺斯州)统计软件进行数据统计分析。采用单因素方差分析(One-Way ANOVA),每种镧改性海藻在不同吸附剂用量和初始pH条件下,对磷吸附量的差异性以及在同一吸附剂用量或初始pH条件下4种镧改性海藻对磷吸附量的差异性进行检验,采用L-S-D法分析比较平均值,当P<0.05时被认为具有显著差异。
2. 结果与讨论
2.1 样品表征
1) SEM微观形貌。经镧改性后,4种海藻干化粉末的微观表面均变得粗糙且不规则,表明镧已经成功附着在海藻表面(图1)。EP在未改性前呈表面光滑的不规则体,改性后,其表面粗糙,有大量小颗粒附着,结构出现坍塌,并产生大量孔道,为磷的附着提供了大量的有效吸附位点。UL和RP在未改性前,表面结构均较为致密,整体呈块状结构。经改性后,UL表面变为粗糙的片状结构且形成了很多较小的空隙结构,La-RP呈凹凸状,褶皱区域增加。LJ在改性前,相较于其他3种材料,粗糙多孔的特点最突出,经改性后,La-LJ呈层状表面结构,有较大孔道。
 图 1 镧改性前后4种海藻SEM图谱Figure 1. SEM images of four types of seaweeds before and after lanthanum modification
图 1 镧改性前后4种海藻SEM图谱Figure 1. SEM images of four types of seaweeds before and after lanthanum modification2) FT-IR。4种海藻未改性前,特征吸收峰大致相似(见图2)。在3 422 cm−1处,4种海藻均有较宽谱峰,由—OH伸缩振动产生[19];在2 974 cm−1处的吸收峰是由烷基对碳氢的拉伸振动所产生的[19],1 655 cm−1处的吸收峰为海带中游离水H—O—H键的弯曲振动峰[20],1 059 cm−1处的吸收峰归属于C—OH伸缩振动吸收峰[21]。
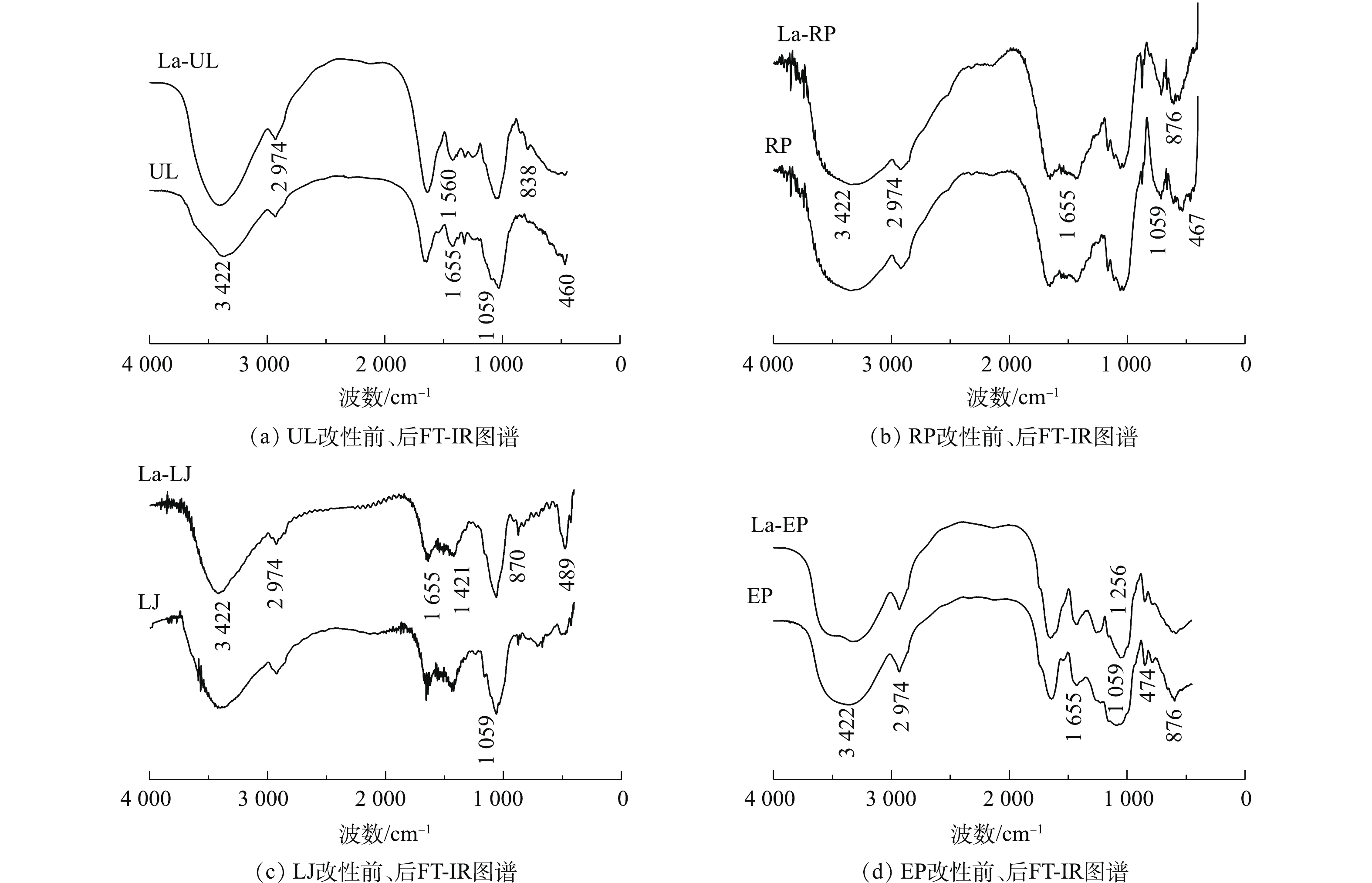 图 2 镧改性4种海藻前后FT-IR图谱Figure 2. FT-IR spectra of four types of seaweeds before and after lanthanum modification
图 2 镧改性4种海藻前后FT-IR图谱Figure 2. FT-IR spectra of four types of seaweeds before and after lanthanum modification经镧改性后,4种海藻在位于3 442 cm−1和1 059 cm−1处的特征吸收峰均一定程度地变弱或变窄。3 422 cm−1处羟基的红外吸收峰减弱的原因可能是由于镧覆盖了部分表面羟基基团,导致海藻表面羟基总量下降,且由于—OH部分被OH−取代,使振动峰变窄[3];1 059 cm−1处C—OH吸收峰的减弱是因为镧的负载形成了C—O—La配位键,从而削弱了C—OH振动峰[21]。此外,La-EP在1 256 cm−1处有新的特征吸收峰,这可能是C—H弯曲振动峰所形成的,而镧改性使其生成C—La键[15]。La-UL、La-RP、La-LJ和La-EP分别在460、467、489和474 cm−1处产生新的特征吸收峰或特征吸收峰有变宽趋势,这是由于La—O键的形成所导致的[22],表明经过改性处理,镧已成功附着在几种海藻的表面。
3) XRD。镧改性前后4种海藻的XRD图谱均出现较明显的变化(图3)。未改性前LJ和UL的主要晶相为方解石和石英,EP的主要晶相为方解石、白云母和石英,RP的主要晶相为NaCl和CaSO4。经改性后,镧主要以氧化物的形式存在于海藻表面。LJ、UL和EP主要在13.82°、25.44°、29.07°和47.15°出现La2O3的衍射峰,表明镧已经成功负载于海藻表面。但RP经改性后特征衍射峰消失,呈现非晶态的衍射特征。这可能是由于镧改性使其晶型发生了改变,RP由晶态转变成了非晶态而未显示出特征衍射峰。
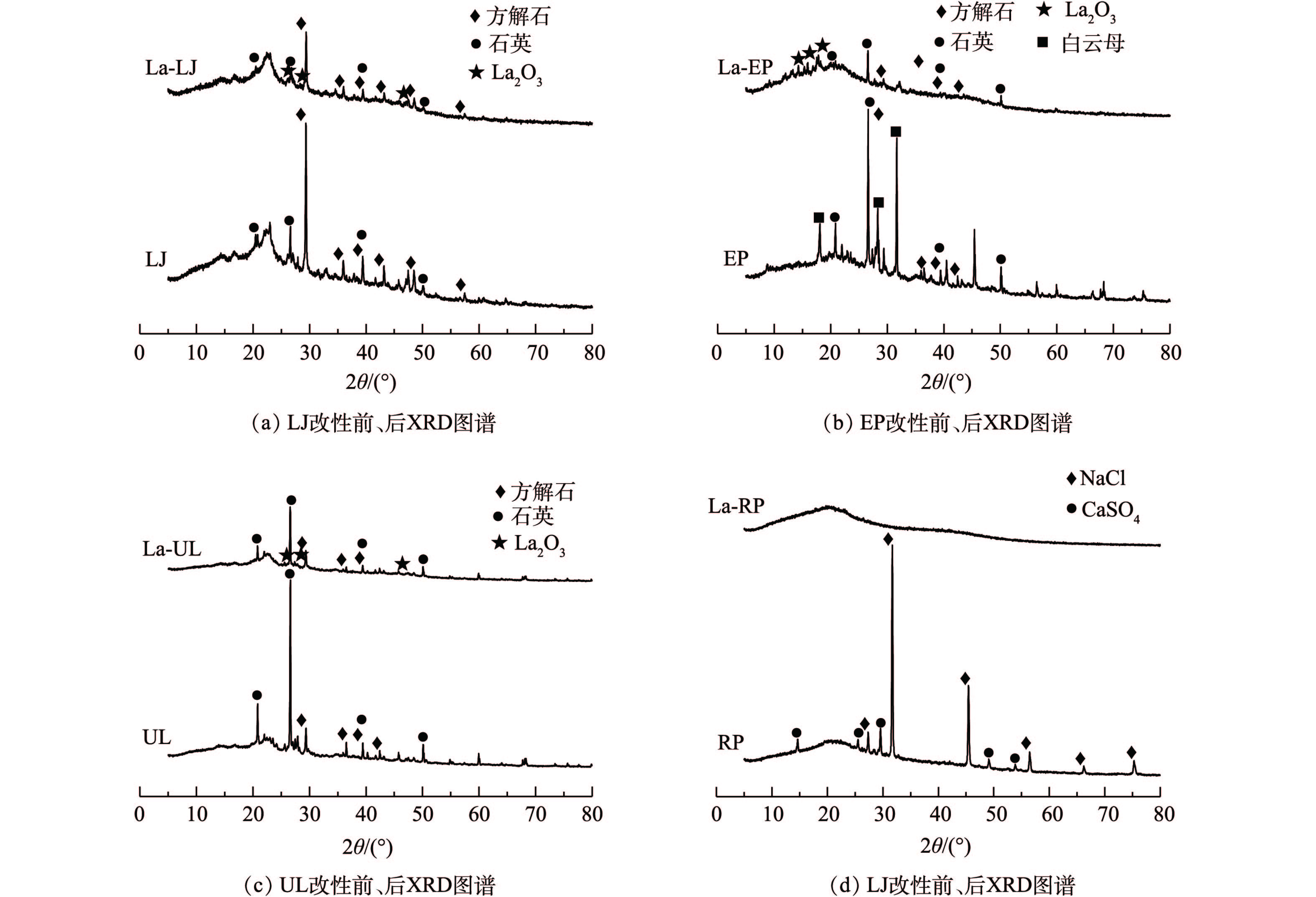 图 3 镧改性4种海藻前后XRD图谱Figure 3. XRD patterns of four types of seaweeds before and after lanthanum modification
图 3 镧改性4种海藻前后XRD图谱Figure 3. XRD patterns of four types of seaweeds before and after lanthanum modification2.2 改性海藻在模拟废水中对磷的去除性能
1)吸附剂用量对吸附效果的影响。吸附剂的投加量是一个既影响处理效果又影响处理成本的重要技术参数[20]。在本研究中,当吸附剂用量从0.02 g增加到0.25 g时,4种镧改性海藻对磷的吸附率随吸附剂用量的增加而呈指数趋势(La-LJ、La-UL、La-RP和La-EP的拟合度分别为0.953、0.974、0.977和0.972)升高(图4(a),P<0.05)。这是由于在吸附初期,吸附面积增大,吸附位点增加使吸附率不断增加,但随着吸附的进行,吸附位点逐渐达到饱和,吸附率趋于稳定。而4种镧改性海藻对磷的吸附量则呈指数趋势减少(图4(b))。这可能是由于4种材料的吸附点位未达到吸附饱和状态。当吸附剂用量为0.02 g时,4种镧改性海藻单位质量吸附剂的吸附量均为最高,La-LJ、La-UL、La-RP和La-EP分别为10.85、9.44、9.99和8.81 mg·g−1。在4种材料中,La-LJ的吸附量显著高于其余3种材料(P<0.05),La-EP显著低于其余3种材料(P<0.05)。当吸附剂用量增加至0.25 g时,4种镧改性的海藻对磷的吸附率均达到了90%以上,La-EP对磷的吸附率甚至达到了99%。
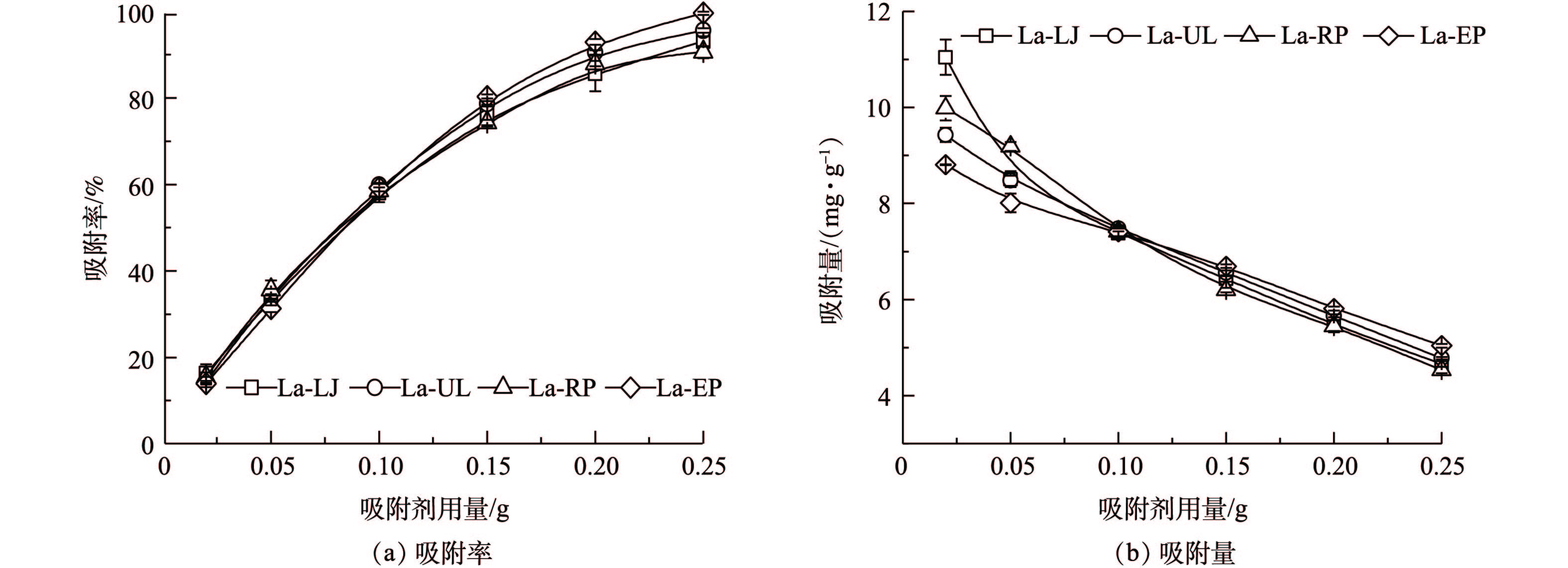 图 4 吸附剂投加量对4种镧改性海藻去除模拟废水中磷的吸附量和吸附率的影响Figure 4. Effects of dosage on adsorption capacities and rates of phosphorousin simulation wastewater with the four types of La-seaweeds
图 4 吸附剂投加量对4种镧改性海藻去除模拟废水中磷的吸附量和吸附率的影响Figure 4. Effects of dosage on adsorption capacities and rates of phosphorousin simulation wastewater with the four types of La-seaweeds在本研究中,磷的吸附量和吸附率随4种镧改性海藻用量的增加所呈现的变化趋势,与镧负载的聚乙烯醇/海藻酸钠水凝胶珠从废水中吸附磷的变化趋势相似[23]。为了兼顾吸附性能与经济效益,本研究在模拟废液初始pH对磷吸附效果、动力学过程和等温吸附过程实验中,均选取0.1 g作为吸附剂投加量。
2)模拟废液初始pH对磷吸附效果的影响。溶液pH通过影响溶液中阳离子存在形态及吸附剂表面电荷从而影响吸附量的变化[19]。在本研究中,当初始pH从3上升到8时,4种镧改性海藻对磷的吸附量随pH的升高呈现不同的变化趋势(图5)。其中,La-LJ和La-RP对磷的吸附量随着初始pH的升高而呈现下降的趋势(P<0.05)。这2种材料对磷的吸附量随pH增加的变化趋势与MgO负载的生物炭对废水中磷吸附的变化趋势相似[4]。这可能是由于pH的升高使吸附剂表面的OH−增多,形成带负电荷的反离子层,导致吸附剂与磷酸盐之间的静电排斥力增大,因而吸附阻力也增大造成的[24]。La-UL和La-EP对磷的吸附量则随pH的升高呈现上升的趋势,且2种吸附剂在不同的pH条件下差异显著(P<0.05)。这可能是由于废液pH的升高使负载在海藻表面的镧与水中的羟基结合形成镧羟基化合物,而镧氢氧化物能络合磷酸根离子,从而吸附去除水中的磷。当pH为3时,4种镧改性海藻间吸附量差异显著(P<0.05),La-LJ和La-RP对磷的吸附量达到了最大,分别为7.18 mg·g−1和7.33 mg·g−1。当pH为8时,4种镧改性海藻对磷的吸附量差异显著(P<0.05),La-UL和La-EP对磷的吸附量达到最大,分别为8.06 mg·g−1和8.86 mg·g−1。镧改性的4种海藻在pH升高时,呈现2种不同变化趋势的原因可能是镧的负载形式不同。LJ和RP中镧可能负载在内部,pH的影响主要由表面羟基的静电作用所决定[25];UL和EP中镧可能负载在表面,pH的影响主要由镧氢氧化物的络合作用所决定。因此,La-LJ和La-RP与磷酸盐的反应机制可能由表面的配体交换或离子交换引起,而La-UL和La-EP与磷酸盐的反应机制可能由内部的复合反应引起。
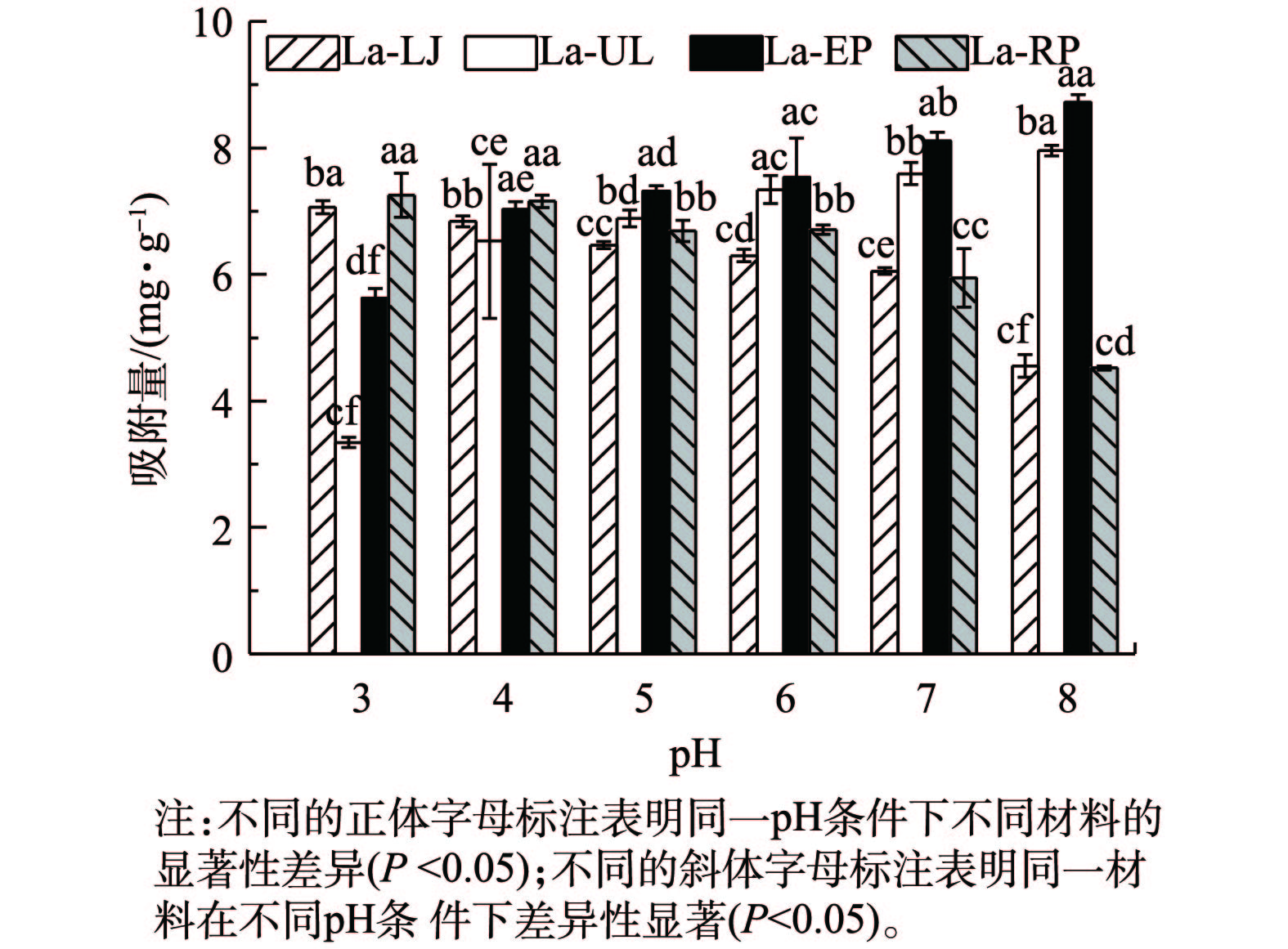 图 5 4种镧改性海藻在不同pH条件下对模拟废水中磷吸附量的影响Figure 5. Effects of pH on adsorption capacities and rates of phosphorous in simulation wastewater withthe four types of La-seaweeds
图 5 4种镧改性海藻在不同pH条件下对模拟废水中磷吸附量的影响Figure 5. Effects of pH on adsorption capacities and rates of phosphorous in simulation wastewater withthe four types of La-seaweeds3)动力学吸附过程。吸附时间也是影响废水中磷吸附量的因素之一[26]。当时间从30 min增加至560 min时,4种镧改性海藻对磷的吸附量不断增加并呈现趋于稳定的趋势(图6(a))。这是因为在反应初期,镧改性的海藻提供了大量吸附位点,磷酸根离子快速迁移至吸附剂表面,但随着吸附位点的减少,反应速率逐渐减慢直至达到平衡[4]。其中La-UL相较于其他3种镧改性海藻,随着吸附时间的增加,其吸附量的增加趋势更加明显。这一现象表明,La-UL相较于La-EP、La-RP和La-LJ 3种材料,其对废水中的磷酸根具有更高的亲和能力。
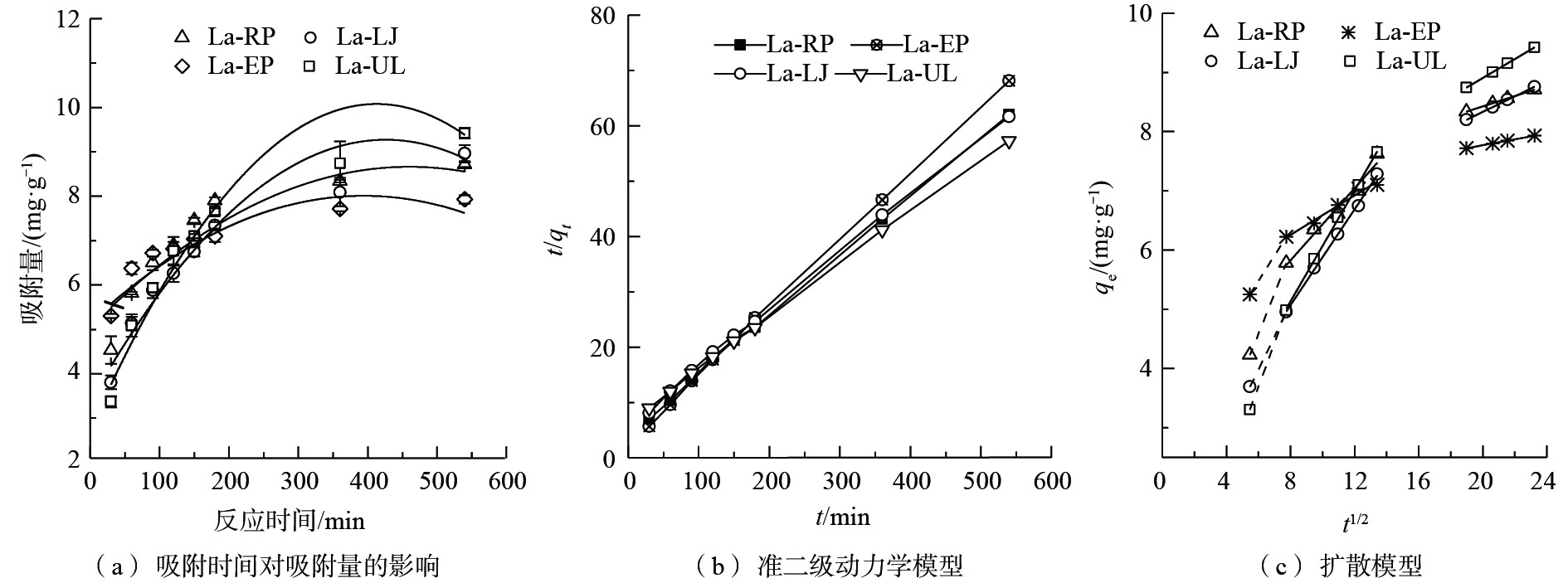 图 6 4种镧改性海藻对模拟废水中磷的吸附动力学Figure 6. Adsorption kinetic models of phosphorous in simulation wastewater with the four types of La-seaweeds
图 6 4种镧改性海藻对模拟废水中磷的吸附动力学Figure 6. Adsorption kinetic models of phosphorous in simulation wastewater with the four types of La-seaweeds本研究采用准二级动力学模型(图6(b))、粒子内扩散模型(图6(c))和叶诺维奇模型进行拟合。表1和表2给出了准二级动力学模型、叶诺维奇模型和粒子内扩散模型的部分参数。准二级动力学模型假设吸附体系为单层吸附体系,能更好地描述吸附反应的全过程,如液膜扩散、表面吸附和内扩散等[27]。由表1可知,准二级动力学模型的可决系数较高(R2均为0.99以上),表明准二级动力学模型能很好地描述4种镧改性的海藻对废水中磷的吸附过程。这也表明,4种镧改性海藻对磷的吸附速率受化学吸附机理控制,而不是受溶液内的扩散过程所控制[28]。准二级动力学模型表明该吸附过程涉及到吸附剂与吸附物质之间的电子共用和电子转移[29]。通过准二级动力学模型的吸附速率常数,4种镧改性的海藻k2为0.10~0.12 g·(mg·min)−1,远高于镧和铈改性的沸石(k2分别为0.036 9 g·(mg·min)−1和0.004 0 g·(mg·min)−1)[30]。
表 1 4种镧改性海藻对模拟废水中磷吸附的准二级动力学模型和叶诺维奇模型相关参数Table 1. Parameters of pseudo-second order and Elovich models for phosphorus adsorption in simulationwastewater with the four types of La-seaweeds吸附剂 准二级动力学模型 叶诺维奇模型 k2 qe R2 αs βs R2 La-LJ 0.10 9.63 0.999 0.56 0.56 0.991 La-UL 0.10 10.48 0.999 0.47 0.47 0.990 La-EP 0.12 8.22 0.999 1.10 1.10 0.977 La-RP 0.11 9.34 0.999 0.65 0.65 0.974 | Show Table DownLoad:
CSV
表 2 4种镧改性海藻对模拟废水中磷吸附的粒子内扩散模型相关参数Table 2. Parameters of intra-particle diffusion model for phosphorous in simulation wastewaterwith the four types of La-seaweeds
DownLoad:
CSV
表 2 4种镧改性海藻对模拟废水中磷吸附的粒子内扩散模型相关参数Table 2. Parameters of intra-particle diffusion model for phosphorous in simulation wastewaterwith the four types of La-seaweeds吸附剂 第1阶段 第2阶段 第3阶段 kip1 C1 R2 kip2 C2 R2 kip3 C3 R2 La-LJ 0.56 0.65 1 0.40 1.83 0.999 0.13 5.71 0.999 La-UL 0.74 −0.74 1 0.47 1.39 0.999 0.16 5.71 0.999 La-EP 0.43 2.90 1 0.17 4.93 0.973 0.05 6.78 0.999 La-RP 0.68 0.48 1 0.30 3.39 0.959 0.09 6.68 0.999 | Show TableDownLoad:
CSV
叶诺维奇模型也能较好地拟合本研究的吸附过程。该模型用来描述化学吸附过程,且假设实际的固体表面能量分布不均匀[11]。比较由叶诺维奇模型获得的吸附速率常数(表1),La-EP的吸附速率常数(1.10)大于其他3种镧改性海藻(La-RP 0.65、La-LJ 0.56、La-UL 0.47)。因此,La-EP比其他3种镧改性海藻能更有效地吸附废水中的磷。
由图6(c)可以看出,4种镧改性的海藻对磷的吸附过程可拟合为3个阶段。每一阶段都可通过拟合直线斜率的变化来确定。曲线的斜率表征了粒子内扩散的速率参数,而截距与边界的厚度成正比。第1阶段在开始前30 min内,直线斜率最大(表2),吸附量随时间的增长快速增加,吸附开始时,反应主要由膜扩散控制,且由于外表面存在大量的有效吸附位点,膜扩散速率很快。在此过程中,磷酸盐从溶液中扩散到镧改性的海藻周围的边界层,吸附剂与磷酸盐之间主要以静电吸引的方式进行吸附[27]。第2阶段为60~180 min,直线斜率减小,此时磷酸盐穿过边界层到达吸附剂的内表面,颗粒内扩散为速率限制步骤。第3阶段为360~540 min,直线斜率最小,在此阶段中,吸附物质与吸附位点相互作用,逐渐达到吸附平衡。4种材料的拟合直线均没有经过原点,表明该吸附过程不仅仅由粒子间的扩散过程控制,还由膜扩散过程控制[26]。此现象与La(OH)3改性蛭石吸附磷的动力学研究结果[28]相似。
4)等温吸附过程。为探究4种镧改性干化海藻粉末对磷吸附去除的可能性及最大吸附量,采用了Freundlich模型和Langmuir模型模拟实验数据(图7)。由表3可知,通过比较2种模型的R2,4种镧改性的海藻对磷的吸附过程用Freundlich模型拟合比用Langmuir模型拟合效果更好。这表明在实验磷酸根初始浓度为20~100 mg·L−1时,4种镧改性的海藻对磷的吸附过程是一种非均相吸附,并且可以存在多层吸附[28]。由Langmuir模型可知,La-LJ、La-UL、La-RP和La-EP的最大吸附量分别为10.80、8.94、11.25和9.90 mg·g−1,是未改性前(LJ 0.449 mg·g−1、UL 0.237 mg·g−1、RP 0.169 mg·g−1和EP 0.387 mg·g−1)的24~66倍。相比于镧负载的多孔陶粒[29]、镧和铈改性的沸石[30]等,镧改性的海藻具有良好的吸附效果。在Freundlich拟合方程中,参数1/n表示吸附剂吸附性能的强弱,1/n越大,表明吸附性能越强,一般认为1/n为0.1~0.5时吸附容易进行[31]。在本研究中,4种吸附剂的1/n值均小于0.5,表明镧改性的4种海藻对磷的吸附过程均很容易进行(表3)。而且La-RP和La-EP的1/n值均远小于La-LJ和La-UL,表明前两者对磷的吸附性能更好。
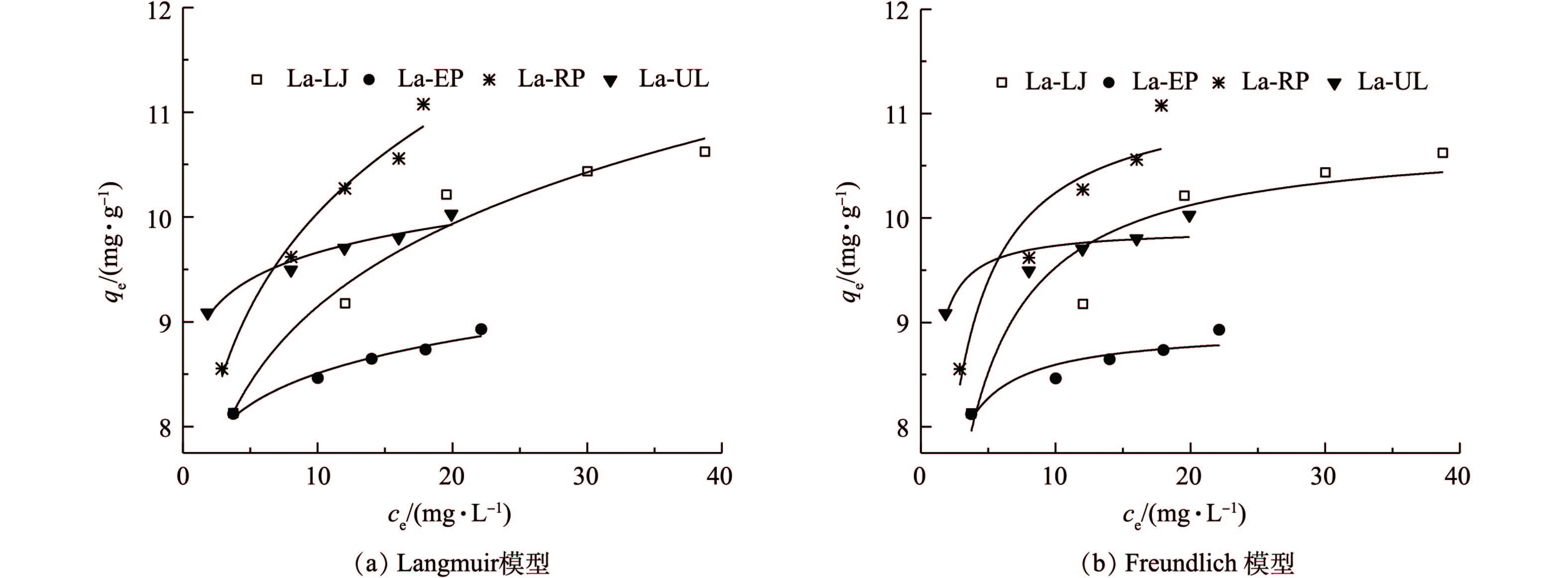 图 7 4种镧改性海藻对模拟废水中磷的等温吸附模型拟合曲线Figure 7. Isothermal parameters adsorption models of phosphorous in simulation wastewaterwith the four types of La-seaweeds表 3 4种镧改性海藻对模拟废水中磷的等温吸附模型Table 3. Isothermal adsorption model parameters for phosphorous in simulation wastewater with the four types of La-seaweeds
图 7 4种镧改性海藻对模拟废水中磷的等温吸附模型拟合曲线Figure 7. Isothermal parameters adsorption models of phosphorous in simulation wastewaterwith the four types of La-seaweeds表 3 4种镧改性海藻对模拟废水中磷的等温吸附模型Table 3. Isothermal adsorption model parameters for phosphorous in simulation wastewater with the four types of La-seaweeds材料 Langmuir Freundlich qm/(mg·g-1) ka R2 kf 1/n R2 La-LJ 10.80 0.75 0.89 6.95 0.12 0.96 La-EP 8.94 2.48 0.85 7.56 0.05 0.97 La-RP 11.25 1.02 0.87 7.33 0.14 0.97 La-UL 9.90 5.78 0.77 8.83 0.04 0.94 | Show TableDownLoad:
CSV
2.3 改性海藻在养猪废水中对磷的去除性能
本研究采用养猪废水对比研究4种海藻用镧改性前后对磷吸附的实际效果(图8)。未改性前,LJ、UL、RP和EP对养猪废水中磷的去除率分别为5.34%、2.81%、2.01%和4.60%,而经镧改性后,La-LJ、La-UL、La-EP和La-RP对磷的去除率分别达到85.54%、94.11%、92.88%和85.81%,这表明4种镧改性后的海藻在组成复杂的养猪废水中具有较强的抗干扰能力。其中UL未改性前对磷的去除率较小,但La-UL对养猪废水中磷的吸附率在4种改性海藻中达到了最高。但是,LJ作为未改性前4种海藻中对磷去除率最高的材料,经镧改性后,在养猪废水中对磷的去除率却最低。吸附后废水中剩余磷含量均小于2.5 mg·L−1,可达到《畜禽养殖业污染物排放标准》(GB 18596-2001)。因此,镧改性的4种海藻在废水除磷的应用中具有潜力。此外,本研究采用的镧改性海藻成本相对较低,经吸附后的材料可直接用作肥料,更具有经济价值。
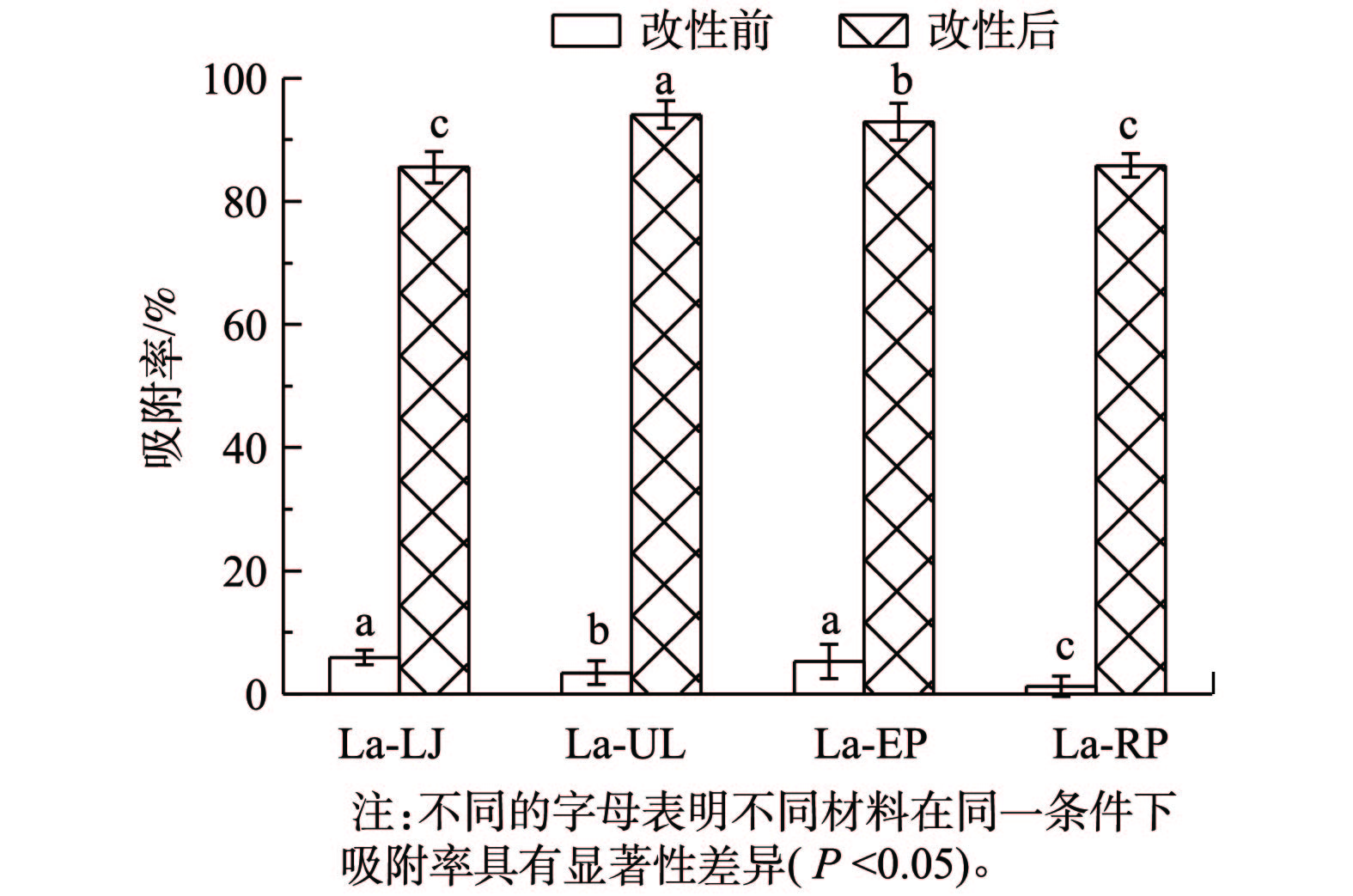 图 8 4种未改性海藻和镧改性海藻对实际废水中磷的吸附率Figure 8. Adsorption rates of phosphorous in swine wastewater with the four types of La-seaweeds
图 8 4种未改性海藻和镧改性海藻对实际废水中磷的吸附率Figure 8. Adsorption rates of phosphorous in swine wastewater with the four types of La-seaweeds3. 结论
1)镧负载改性的海藻干化粉末对磷的吸附去除能力显著提高,La-LJ的吸附效果最好,最大吸附量可达10.85 mg·g−1。
2)当吸附剂用量为0.25 g时,4种镧改性海藻对模拟废水中磷的吸附率均达到90%以上。当pH为3时,La-LJ和La-RP对磷的吸附量最大,分别为7.18 mg·g−1和7.33 mg·g−1;当pH为8时,La-UL和La-EP对磷的吸附量最大,分别为8.06 mg·g−1和8.86 mg·g−1。
3) 用准二级动力学模型和Freundlich吸附等温线模型能更好地拟合4种镧改性海藻粉末对磷的动力学吸附过程和等温吸附过程。
4) 4种海藻经镧改性后对养猪废水中磷的去除率明显提高,能达到85.54%以上,可实现磷达标排放。
-
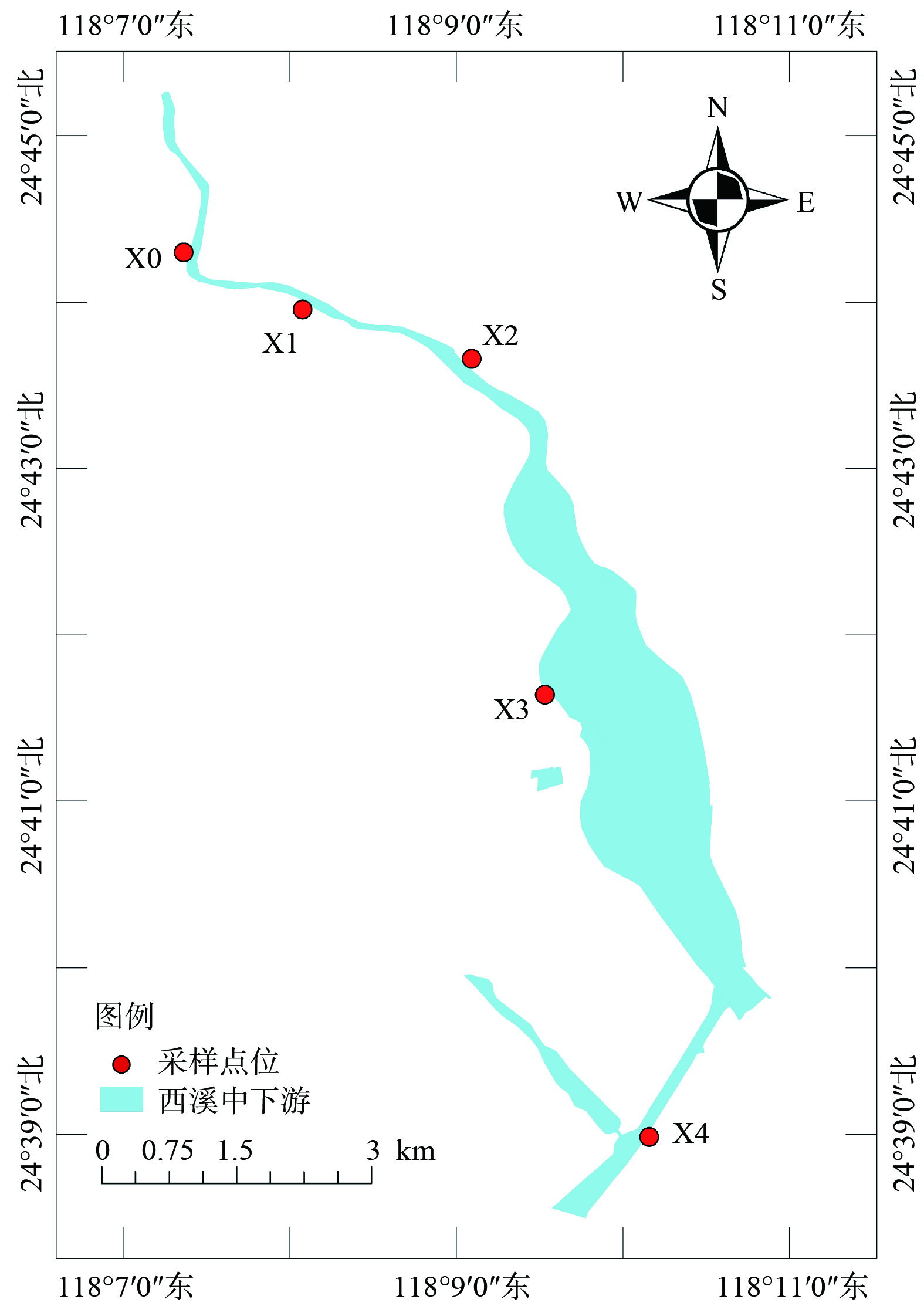
图 1 研究区所在位置及各采样点地理位置图
Figure 1. Location of the study area and geographical map of each sampling point
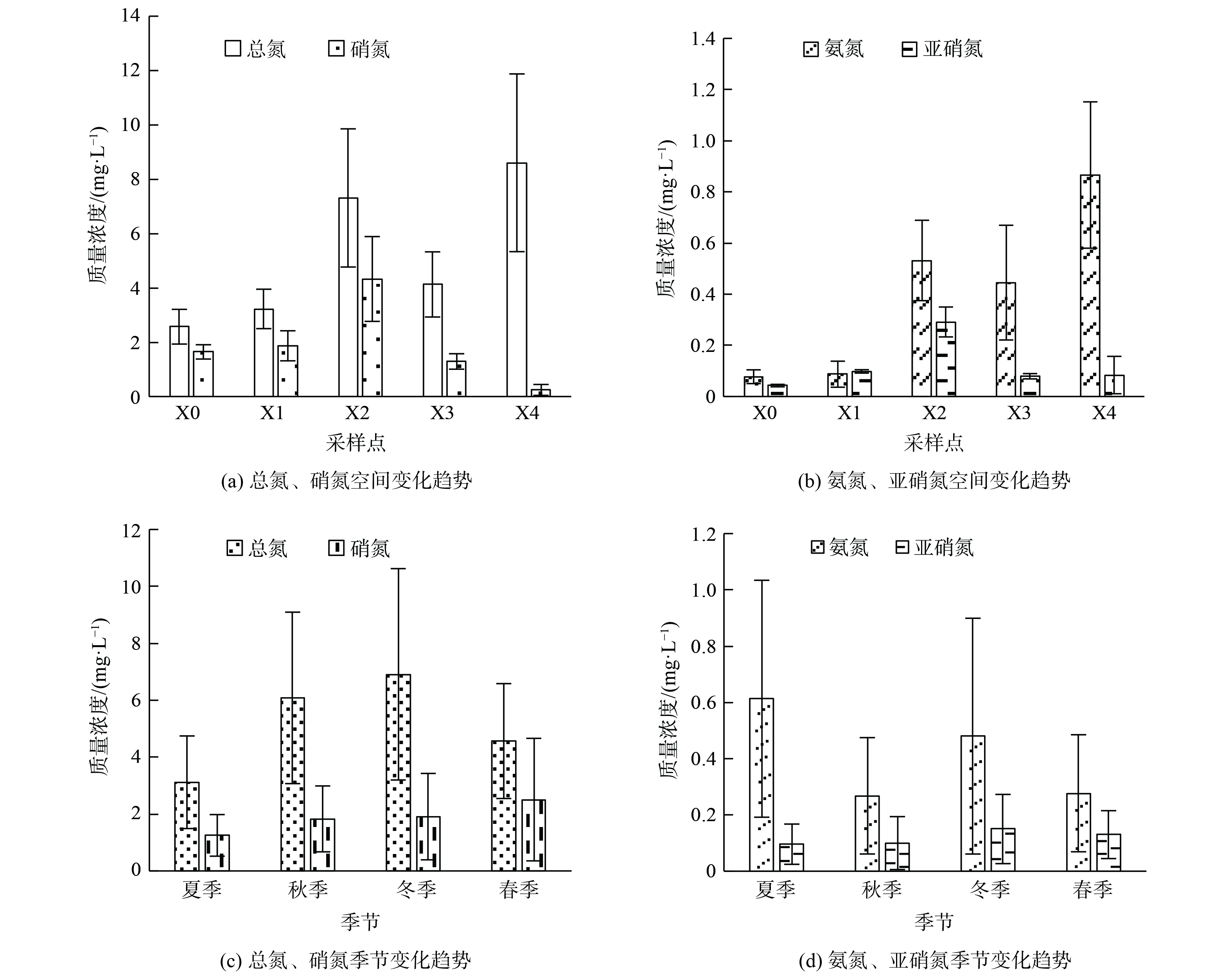
图 2 上覆水中各氮素时空变化趋势
Figure 2. Temporal and spatial variation trend of nitrogen in overlying water

图 3 沉积物中各氮素时空变化趋势
Figure 3. Temporal and spatial variation trend of nitrogen in sediment
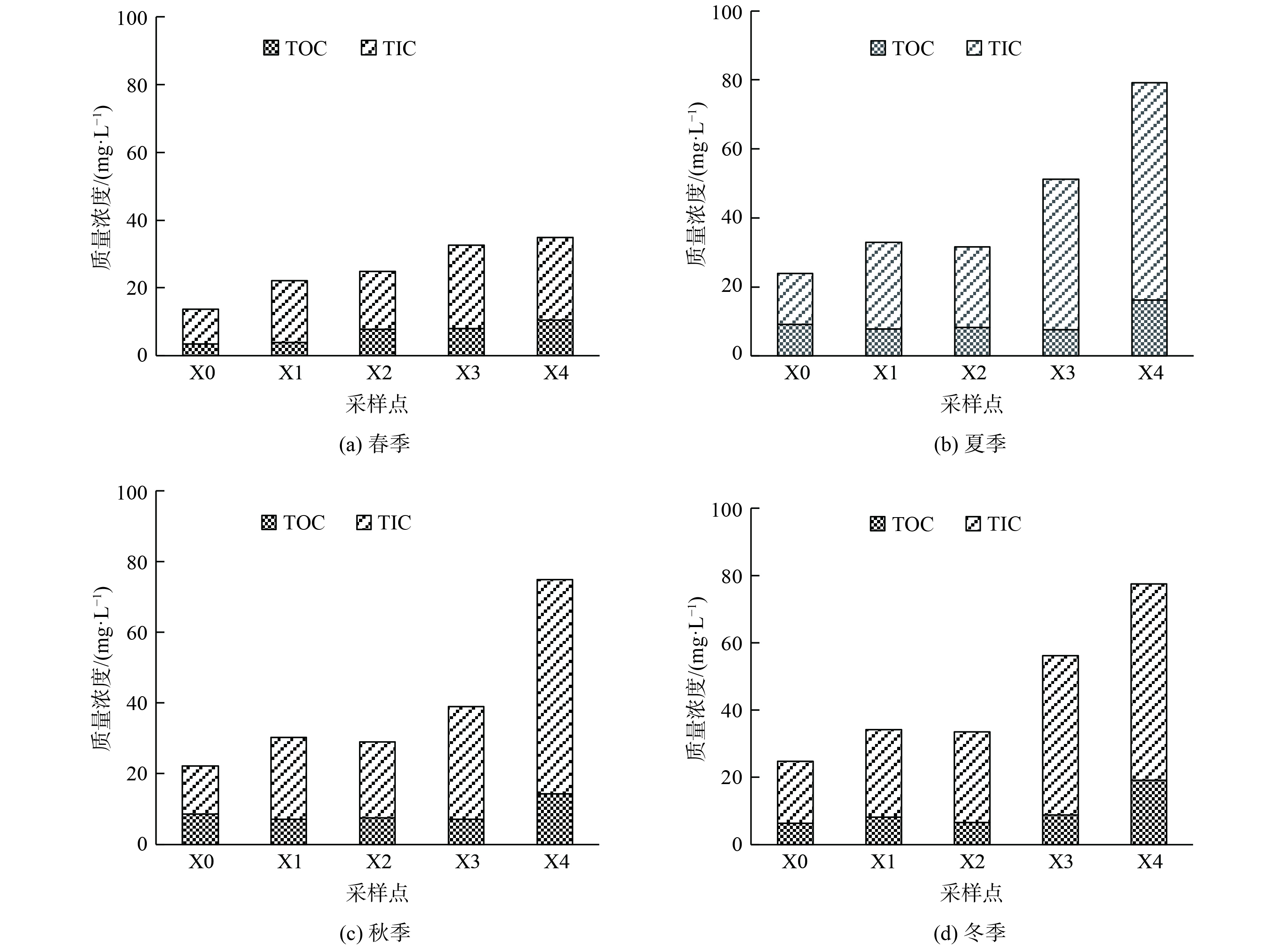
图 4 不同季节上覆水中各碳素空间变化趋势
Figure 4. Temporal and spatial dynamic variation trend of carbon in overlying water in different seasons
表 1 研究区域上覆水及沉积物孔隙水中理化特征
Table 1. Physicochemical characteristics of overlying water and sediment pore water in the study area
mg·L−1 采样点 上覆水 孔隙水 硝氮 亚硝氮 氨氮 总氮 总有机碳 硝氮 亚硝氮 氨氮 总有机碳 X0 0.850 0.035 0.100 3.02 9.23 0.63 0.03 0.17 3.00 X1 0.744 0.035 0.292 3.22 7.89 0.66 0.03 0.36 3.01 X2 1.585 0.094 0.175 4.89 8.21 1.14 0.09 0.20 4.89 X3 1.475 0.047 0.214 4.52 7.56 1.01 0.04 0.29 4.52 X4 1.324 0.328 2.391 5.68 16.29 0.96 0.30 3.80 5.68 平均值 1.200 0.110 0.630 4.26 9.84 0.88 0.10 0.96 4.22
下载: 导出CSV
表 2 沉积物中各氮素理化特征
Table 2. Physicochemical characteristics of nitrogen in sediment
mg·kg−1 采样点 硝氮 亚硝氮 氨氮 有机氮 全氮 X0 7.34 0.24 4.42 2 705 2 717 X1 6.6 0.24 5.38 2 903 2 915 X2 10.58 0.54 4.77 4 558 4 574 X3 10.4 0.35 5.05 4 212 4 228 X4 8.18 1.71 15.81 5 350 5 376 平均值 8.62 0.61 7.09 3 946 3 962
下载: 导出CSV
-
[1] XU G P, WANG X F, CHEN L J. Application of principal component analysis for the estimation of source of heavy metal contamination in sugarcane soil[J]. Applied Mechanics and Materials, 2014, 651-653: 1402-1409. doi: 10.4028/www.scientific.net/AMM.651-653.1402 [2] 朱广伟, 陈英旭. 沉积物中有机质的环境行为研究进展[J]. 湖泊科学, 2001, 13(3): 272-279. doi: 10.3321/j.issn:1003-5427.2001.03.012 [3] WANG J F, CHEN J G, DING S M, et al. Effects of seasonal hypoxia on the release of phosphorus from sediments in deep-water ecosystem: A case study in Hongfeng Reservoir, Southwest China.[J]. Environmental Pollution, 2016, 219: 858-865. doi: 10.1016/j.envpol.2016.08.013 [4] HARIKA, NARUMANCHI, DISHANT, et al. In situ, high-resolution evidence of phosphorus release from sediments controlled by the reductive dissolution of iron-bound phosphorus in a deep reservoir, southwestern China[C]// 2018 IEEE International Conference on Smart Cloud. 0. [5] WANG J, CAI C, LI Y, et al. Denitrifying anaerobic methane oxidation: A previously overlooked methane sink in intertidal zone[J]. Environmental Science & Technology, 2019, 53(1): 203-212. [6] SIVAN O, ADLER M, PEARSON A, et al. Geochemical evidence for iron-mediated anaerobic oxidation of methane[J]. Limnology and Oceanography, 2011, 56(4): 1536-1544. doi: 10.4319/lo.2011.56.4.1536 [7] RAGHOEBARSING A A, POL A, VAND P K, et al. A microbial consortium couples anaerobic methane oxidation to denitrification[J]. Nature, 2006, 440: 918-921. doi: 10.1038/nature04617 [8] 保琼莉, 黄益宗. 湿地反硝化型甲烷厌氧氧化研究进展[J]. 生态学杂志, 2019, 38(10): 3202-3210. doi: 10.13292/j.1000-4890.201910.004 [9] 吴红宝. 典型河流生态系统碳氮温室气体扩散传输研究[D]. 芜湖: 安徽师范大学, 2017. [10] BEAULIEU J J, SHUSTER W D, REBHOLZ J A. Nitrous oxide emissions from a large, impounded river: The Ohio River[J]. Environmental Science & Technology, 2010, 44(19): 7527. [11] 王瑞飞, 王亚利, 杨清香. 淡水生态系统中反硝化型厌氧甲烷氧化微生物的研究进展[J]. 环境污染与防治, 2018, 40(12): 1443-1448. doi: 10.15985/j.cnki.1001-3865.2018.12.023 [12] DEUTZMANN J S, STIEF P, BRANDES J, et al. Anaerobic methane oxidation coupled to denitrification is the dominant methane sink in a deep lake[J]. Proceedings of the National Academy of Sciences, 2014, 111(51): 18273-18278. doi: 10.1073/pnas.1411617111 [13] SHEN L D, ZHU Q, LIU S, et al. Molecular evidence for nitrite-dependent anaerobic methane-oxidising bacteria in the Jiaojiang Estuary of the East Sea (China)[J]. Applied Microbiology & Biotechnology, 2014, 98(11): 5029-5038. [14] ETTWIG K F, BUTLER M K, PASLIER D L, et al. Nitrite-driven anaerobic methane oxidation by oxygenic bacteria[J]. Nature, 2010, 464(7288): 543-548. doi: 10.1038/nature08883 [15] KOJIMA H, TOKIZAWA R, KOGURE K, et al. Community structure of planktonic methane-oxidizing bacteria in a subtropical reservoir characterized by dominance of phylotype closely related to nitrite reducer[J]. Scientific Reports, 2014, 4: 5728. doi: 10.1038/srep05728 [16] SHEN L D, LIU S, HUANG Q, et al. Evidence for the cooccurrence of nitrite-dependent anaerobic ammonium and methane oxidation processes in a flooded paddy field[J]. Applied & Environmental Microbiology, 2014, 80(24): 7611-7619. [17] HU B L, SHEN L D, LIAN X, et al. Evidence for nitrite-dependent anaerobic methane oxidation as a previously overlooked microbial methane sink in wetlands[J]. Proceedings of the National Academy of Sciences, 2014, 111(12): 4495-4500. doi: 10.1073/pnas.1318393111 [18] YAN P, LI M, WEI G, et al. Molecular fingerprint and dominant environmental factors of nitrite-dependent anaerobic methane-oxidizing bacteria in sediments from the Yellow River Estuary, China[J]. PLOS ONE, 2015, 10(9): e0137996. doi: 10.1371/journal.pone.0137996 [19] SHEN L D, LIY S, ZHU Q, et al. Distribution and diversity of nitrite-dependent anaerobic methane-oxidising bacteria in the sediments of the Qiantang River[J]. Microbial Ecology, 2014, 67(2): 341-349. doi: 10.1007/s00248-013-0330-0 [20] GUILLAUME B, DIDIER J, CORINNE B P, et al. Production and consumption of methane in freshwater lake ecosystems.[J]. Research in Microbiology, 2011, 162(9): 832-847. doi: 10.1016/j.resmic.2011.06.004 [21] SONG G D, LIU S M, KUYPERS M M M, et al. Application of the isotope pairing technique in sediments where anammox, denitrification, and dissimilatory nitrate reduction to ammonium coexist[J]. Limnology and Oceanography:Methods, 2016, 1(9): 63-73. [22] TRIMMER M,NICHOLLS J C,DEFLANDRE B. Anaerobic ammonium oxidation measured in sediments along the Thames estuary, United Kingdom[J]. Applied and Environmental Microbiology, 2003, 69(11): 6447-6454. [23] KUYPERS M M M, SLIEKERS A O, LAVIK G, et al. Anaerobic ammonium oxidation by anammox bacteria in the Black Sea[J]. Nature, 2003, 422(10): 608-611. [24] RISGAARD-PETERSEN N, NIELSEN L P, RYSGAARD S, et al. Application of the isotope pairing technique in sediments where anammox and denitrification coexist[J]. Limnology and Oceanography:Methods, 2003, 1(1): 63-73. doi: 10.4319/lom.2003.1.63 [25] CAO W Z, YANG J X, LI Y. Dissimilatory nitrate reduction to ammonium conserves nitrogen in anthropogenically affected subtropical mangrove sediments in Southeast China[J]. Marine Pollution Bulletin, 2016, MPB-07832: 1-7. [26] GUPTA V, SMEMO K A, YAVITT J B, et al. Stable isotopes reveal widespread anaerobic methane oxidation across latitude and peatland type[J]. Environmental Science & Technology, 2013, 47(15): 8273-8279. [27] WEISS R F. The solubility of nitrogen, oxygen and argon in water and seawater[J]. Deep-Sea Research and Oceanographic Abstracts, 1970, 17(4): 721-735. doi: 10.1016/0011-7471(70)90037-9 [28] WANNINKHOF R. Relationship between gas exchange and wind speed over the ocean[J]. Journal of Geophysical Research:Oceans, 1992, 97(C5): 7373-7382. doi: 10.1029/92JC00188 [29] 陈能汪, 吴杰忠, 洪华生. 九龙江河口区夏季反硝化作用初探[J]. 环境科学, 2011, 32(11): 3229-3234. doi: 10.13227/j.hjkx.2011.11.027 [30] 施瑶. 氮输入对泥炭沼泽碳氮转化的影响研究[D]. 长春: 东北师范大学, 2019. [31] 蒋艾青. 残饵、死鱼及排泄物腐解对山区精养池养殖水质的影响[J]. 现代农业科技, 2007(10): 143-144. doi: 10.3969/j.issn.1007-5739.2007.10.101 [32] 孙洪伟, 尤永军, 赵华南, 等. 游离氨对硝化菌活性的抑制及可逆性影响[J]. 中国环境科学, 2015(1): 95-100. [33] 刘富龙. 氨氮浓度对硝化反应影响的研究[D]. 太原: 太原理工大学, 2009. [34] 徐志伟, 张心昱, 于贵瑞, 等. 中国水体硝酸盐氮氧双稳定同位素溯源研究进展[J]. 环境科学, 2014, 35(8): 3231-3238. doi: 10.13227/j.hjkx.2014.08.056 [35] 王超, 单保庆, 赵钰. 滏阳河水系沉积物硝化速率分布及溶解氧的限制效应[J]. 环境科学学报, 2015, 35(6): 1735-1740. doi: 10.13671/j.hjkxxb.2014.0897 [36] 赵锋, 许海, 詹旭, 等. 太湖春夏两季反硝化与厌氧氨氧化速率的空间差异及其影响因素[J]. 环境科学, 2021, 42(5): 2296-2302. doi: 10.13227/j.hjkx.202008303 [37] 俞琳, 王东启, 邓焕广, 等. 上海市苏州河与崇明岛河流沉积物反硝化速率及其影响因素[J]. 生态与农村环境学报, 2015, 31(1): 100-105. doi: 10.11934/j.issn.1673-4831.2015.01.015 [38] 王子聪, 许海, 詹旭, 等. 天目湖流域沟塘湿地脱氮速率的时空差异[J]. 环境科学研究, 2022, 35(4): 979-988. doi: 10.13198/j.issn.1001-6929.2022.02.01 [39] 王正文, 姚晓龙, 姜星宇, 等. 季节与水文影响下鄱阳湖碟形湖湿地沉积物氮去除功能变化[J]. 长江流域资源与环境, 2022, 31(3): 673-684. [40] 王静, 刘洪杰, 雷禹, 等. 三峡库区小江支流沉积物硝化反硝化速率在蓄水期和泄水期的特征[J]. 环境科学, 2017, 38(3): 946-953. doi: 10.13227/j.hjkx.201606202 [41] 秦红益. 太湖沉积物厌氧氨氧化细菌分布多样性及其活性研究[D]. 南京: 南京师范大学, 2017. [42] 赵永强, 夏永秋, 李博伦, 等. 利用膜进样质谱同时测定河流沉积物反硝化和厌氧氨氧化[J]. 农业环境科学学报, 2014, 33(4): 794-802. doi: 10.11654/jaes.2014.04.026 [43] 彭强, 关傲梅, 齐维晓, 等. 基于同位素示踪和分子生物学技术对潮白河沉积物中氮还原功能特征的研究[J]. 环境科学学报, 2021, 41(11): 4615-4624. doi: 10.13671/j.hjkxxb.2021.0193 [44] 沈李东. 湿地亚硝酸盐型厌氧氨氧化和厌氧甲烷氧化微生物生态学研究[D]. 杭州: 浙江大学, 2014. [45] 张亚迪, 宋永会, 彭剑峰, 等. 浑河底泥反硝化厌氧甲烷氧化菌群落多样性的时空分布特征及其与环境因子关系分析[J]. 环境科学, 2018, 39(8): 3670-3676. doi: 10.13227/j.hjkx.201710162 [46] 袁梦冬, 朱静, 吴伟祥. 微生物厌氧甲烷氧化反硝化研究进展[J]. 微生物学报, 2014, 54(2): 129-138. doi: 10.13343/j.cnki.wsxb.2014.02.002 [47] SHEN L D, HU B L, LIU S, et al. Anaerobic methane oxidation coupled to nitrite reduction can be a potential methane sink in coastal environments[J]. Applied Microbiology & Biotechnology, 2016, 100(16): 7171-7180. [48] XU Y, TONG T, CHEN J, et al. Nitrite-dependent methane-oxidizing bacteria seasonally and spatially shift in a constructed wetland used for treating river water[J]. Ecological Engineering, 2018, 110: 48-53. doi: 10.1016/j.ecoleng.2017.10.005 [49] LOU J, LV J, YANG D Y. Effects of environmental factors on Nitrate-DAMO activity[J]. Water, Air, & Soil Pollution, 2020, 231: 263. doi: 10.1007/s11270-020-04640-9 -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4814
- HTML全文浏览数: 4814
- PDF下载数: 168
- 施引文献: 0











