

-
我国2018年环境状况公报显示,鄱阳湖等5大淡水湖水体均处于富营养状态[1]。水体中氮磷营养物质过多,超过湖泊的自我净化能力,未经降解的污染物通过沉积或颗粒物吸附存蓄在底泥中,造成底泥不同程度的污染,被污染的底泥将成为水体污染物的“汇”[2-4]。随着水体环境条件的变化,底泥中的氮磷营养元素被水生植物生长吸收利用,有一部分被释放到上覆水中而成为水体内源性污染的“源”[5-7],其中磷在限制水体生态系统的初级生产力和调控富营养化状态起到关键作用[8-10]。因此,研究氮磷在湖泊水体中的变化和释放规律对于控制湖泊富营养化具有重要的作用。
乌梁素海流域内有河套灌区和一些工业企业,每年有大量未经处理的工业废水和农田灌溉水排入湖泊,导致水体中氮磷营养物质过多,湖泊的内源污染严重。有研究表明:2004—2006年,该湖泊中100多个监测点数据显示水体总磷和总氮质量浓度平均值为0.16 mg·L−1和6.91 mg·L−1[11-12];2015年,水体中总磷和总氮质量浓度平均值为0.085 mg·L−1和2.52 mg·L−1[13];2001年,河套灌区全年农田排水总氮和总磷质量分别为644.47 t和10.07 t[14];2013年,乌梁素海工业污水和城镇生活污水的排放量分别为1 915.81 t和3 033.96 t[15];2019年,入湖TN、TP的污染负荷为2 037.23 t和55.82 t,湖泊整体水质呈Ⅴ类,仍处于富营养化状态[16]。控制水体富营养化一方面可从源头上减少营养物质排入水体,另一方面可降低水体和底泥中的营养物质。
本研究在农田排水的九排干入湖口附近的七作业区域设置示范区,采用原位生态修复技术,增设曝气增氧设施以增加水体中溶解氧浓度,优化水生植物群落,放养鱼类,收获水生植物并开展了在溶解氧(DO)和pH值变化下原柱状底泥氮磷质量浓度变化的室内模拟实验,分析了水环境因素DO和pH值的变化对底泥中营养物质释放规律的影响。本研究成果可为制定乌梁素海切实有效的富营养化治理措施提供参考。
-
乌梁素海(40°36'N~41°03'N,108°43'E~108°57'E) 位于内蒙古高原巴彦淖尔市乌拉特前旗境内,南北长35~40 km,东西宽5~10 km。目前湖泊面积为293.08 km2,其中177 km2为开阔水域,其余为天然及人工芦苇区。平均水深为1.78 m,在1.1 m与2.77 m之间波动。每年大约在11月开始结冰,至翌年3月为冰封期。该湖泊是历史上由黄河改道形成的河迹湖泊,是我国8大淡水湖之一,也是内蒙古高原具有重要生态功能的天然湖泊湿地,于2002年被国际湿地公约组织列为国际重要湿地[17]。其所在地区属中温带大陆性季风气候,日照充足,历年平均日照时间3 202 h,流域多年平均气温5.2 ℃。流域内多年平均降雨量为200~250 mm,降雨多集中在6—9月,雨季的降雨量占全年降雨量的80%左右。
-
在乌梁素海九排干入湖口附近七作业区设置原位修复实验示范区(图1),示范区面积为3.3 km2。这里是农田排水入湖口附近,营养盐浓度较高,紧挨芦苇荡,水流速率较低。2019年5月,采用松木桩子,防腐钢绳挂网,下挂石笼网固定,用高密度聚乙烯双层拦网建立双层防水围隔,水面上露0.5 m。示范区水深为1.4~1.6 m。2019年5—8月,对示范区的水环境、底泥和生物进行本底调查。其中水体总氮和总磷平均含量分别为2.23 mg·L−1和0.24 mg·L−1,底泥(0~10 cm)中总氮和总磷质量分数分别为1.86 g·kg−1和0.61 g·kg−1。挺水植物以芦苇(Phragmites australis)占优势,沉水植物有龙须眼子菜(Potamogeton pectinatus)、菹草(Potamogeton crispus)等,但数量较少;主要鱼类有鲫鱼(Carassius aumtus)、鲤鱼(Cyprinus carpio)、草鱼(Ctenopharyngodon idellus)、花鲢(Hypophthalmichthys nobilis )等。
乌梁素海富营养化导致芦苇生长旺盛、水下光照和氧气不足,沉水植物群落结构简单,鱼类种类和数量较少。本示范工程的基本思路是:建立围网,将示范区与对照区分开;采用太阳能曝气设施,增加水体溶解氧浓度;优化沉水植物结构群落,放养本地鱼类,收获芦苇等措施,促进湖中营养盐的吸收和转出(图2)。
实验处理于2020年5月初开始,示范区内每隔330 m布设1台推流式太阳能曝气机(LBPQ,苏州鲁班环保科技有限公司,昆山市),以太阳能作为设备运转的直接动力,设置独特的转盘式复氧叶轮,通过转盘叶轮旋转拨动水循环,将水横向推流输送至远端,底部缺氧水体向上补充,实现循环推流、混合和复氧等多重功效。 共布设30台推流式太阳能曝气机,以增加水体中氧气浓度,利于水生生物的生存。5月底投放重量为250~350 g的草鱼幼苗1.5×104 kg。同时种植轮藻(Chara fragilis)、狐尾藻(Myriophyllum verticillatum)、轮叶黑藻(Hydrilla verticillata)、苦草(Vallisneria natans )、金鱼藻(Ceratophyllum demersum)、龙须眼子菜等,1 m2种植25丛。示范区内累计种植水生植物0.2 km2。2020年8—9月,收割芦苇,收割深度约为1 m,累计收割芦苇达5.76×104 t。以示范区以外的湖区作为对照,从北向南以不同间隔分别设置12个采样点,用GPS定位(图1(a));在实验示范区设置8个采样点(图1(b))。在2020年5—11月和2021年1月,在各采样点分别采集水样和底泥样品,进行氮磷含量测定。
-
2020年5—11月,每月在湖区进行水样和底泥样品的采集,冬季冰封期的2021年1月开冰取样。用水样采集器(CG-00,格雷斯普科技开发公司,北京) 采集水样后,将水样装入取样瓶中,放入便携式冰箱,带回实验室后立即进行总氮和总磷的分析测定。总氮(TN)的测定采用碱性过硫酸钾消解-紫外分光光度法,总磷(TP)采用碱熔-钼锑抗分光光度法。现场用便携式水质分析仪(SL1000,HACH,Loveland,CO) 测定水体pH值、电导率、水温、溶解氧等指标。
采集水样后,同时用柱状取泥器(Kajak,KC-Denmark,Copenhagen) 采集底泥样品,采集深度为0~10 cm,装入封口袋中,排去袋中空气,密封放入便携式冰箱,带回实验室后,在阴凉通风处将样品风干,剔除砾石、贝壳及动植物残体等杂质,研磨后过100目筛,充分混匀后装入样品瓶,进行总氮和总磷的分析测定。底泥中总氮(TN)的测定采用半微量凯氏法,总磷(TP)的测定采用硝酸-硫酸消化法。
-
2020年8月,在七作业示范区中部,用柱状取泥器采集底泥样品9份,每3份为一组,采集深度为0~10 cm,将所取柱状泥样直接推入预先制好的直径10 cm,高度66 cm的有机玻璃圆柱中,避光低温保存运回实验室并放入低温培养箱,在25 ℃下进行营养盐释放实验。同时采集水样,放入便携式0~4 ℃的冰箱带回实验室,经0.45 μm 孔径的滤膜过滤,滤去藻类等浮游生物,以减小其对实验的影响。滤膜在使用前用稀盐酸浸泡并消煮,消除有机质对水体的干扰。
圆柱顶盖上开有取样孔,一根气管伸入筒内,气管的另一端与氧气和氮气袋相连;另一根通气管与大气相连,调节柱内溶解氧,用于排除柱内其余气体,并保证其气密性,平时用橡胶塞封闭。将经滤膜过滤后的水样沿圆柱筒壁缓缓注入实验柱中,尽量不要搅动底部的泥样,泥样和水样的高度比值接近1∶1。加入水样后,静沉平衡24 h,调节控制实验条件。向实验圆柱中分别通入氧气,以调节上覆水中溶解氧(DO)质量浓度。有研究表明:当水体中溶解氧质量浓度低于2 mg·L−1时,会导致鱼虾窒息死亡,水体处于缺氧状态;而溶解氧质量浓度在4 mg·L−1以上时,可满足大多数鱼类生存[18]。 SCHUBLE等[19]把溶解氧质量浓度低于3 mg·L−1定义为缺氧。我国地表水溶解氧质量浓度以6、5、3、2 mg·L−1为分级标准[20]。结合文献和实际情况,本实验将溶解氧分别控制为好氧(DO>4 mg·L−1)、缺氧(2 mg·L−1<DO≤4 mg·L−1)、厌氧(DO≤2 mg·L−1) 3种状态。在实验期间,用NaOH和HCl调节上覆水中的pH 值,pH值分别控制为 6.5、8、9。实验进行14 d,每2 d 取1次水样,每次取样15 mL,之后补充水样至原刻度。采用文献中报道的方法[21]测定水样中溶解态无机磷(DIP)、溶解态磷(DTP)、总磷(TP)、总氮(TN)。
-
为了稳定方差异质性,水温、电导率和pH值的数据取自然对数后进行统计分析。实验所测数据均采用SPSS 17.0软件(SPSS,Chicago, IL, USA)进行方差分析,其中实验区和示范区的水体环境参数、水体和底泥氮磷质量浓度数据采用单因素方差分析(ANVOV)(显著性水平α= 0.05),各变量随时间变化的差异采用T-test检验(显著性水平α= 0.05),底泥中营养盐在不同时间下释放规律采用重复测量方差分析(显著性水平α= 0.05)。所有数据均用平均值±标准误差表示。采用 Origin 9.0 软件进行制图。
-
实验示范区和对照区的水温变化差异不显著(P=0.052),但水温随季节变化的差异显著(P<0.001)。实验示范区7月水温最高,平均水温为25.1 ℃,变化区间为24.7~25.9 ℃;1月水温最低,平均水温为0.24 ℃,变化区间为0.1~1.0 ℃(图3(a))。对照湖区7月水温最高,平均水温为27.1 ℃,变化区间为24.1~31.4 ℃;1月水温最低,平均水温为0.19 ℃,变化区间为0.9~1.0 ℃(图3(b))。无论实验示范区还是对照区,5—9月的水温均在15 ℃以上,10月水温降至9.5~14.6 ℃。实验示范区表层水体溶解氧质量浓度为5.9~11.6 mg·L−1,底部水体溶解氧质量浓度为1.2~2.5 mg·L−1,示范区表层和底部水体溶解氧浓度均显著高于对照区(P<0.001)(图3(c)、(d))。
实验示范区和对照区的电导率变化差异显著(P<0.001),对照区平均电导率为3.46 mS·cm−1,示范区为1.54 mS·cm−1。当年5月至翌年1月,电导率基本呈相对缓慢的逐渐降低趋势,11月的电导率最低,到翌年1月,有所回升(图4(a)、(b)),对照区电导率随季节变化的差异显著(P<0.001)。对照区水体平均pH值为8.65,变化区间为8.22~8.75;实验示范区平均pH值为8.43,低于对照区(P=0.045),变化区间为7.80~8.80(图4(c)、(d)),单点最高pH值为9.3,最低值为7.5。示范区水体pH值随季节变化差异显著(P<0.001)。
-
与对照相比,实验示范区水体中总氮(TN)质量浓度显著减低(P<0.001),平均TN质量浓度为0.99 mg·L−1,对照湖区水体平均TN质量浓度为2.89 mg·L−1 (图5(a)、(b)),原位生态修复技术对水体中总氮的削减率达到 66%。TN质量浓度在各月之间的变化差异显著(P<0.001),5月水体中TN质量浓度相对较高,之后随着水生植物的生长,其值有所降低,呈现不规律波动式的变化;11月进入冬季以后,沉水植物的新陈代谢活动基本降为零,此时水体中TN达到最低值;到翌年1月以后,随着气温回升,冰冻层下方微生物对水体中有机物质进行分解,水体中TN质量浓度有所回升。
实验处理显著降低了水体中总磷(TP)质量浓度 (P<0.001),实验示范区平均TP质量浓度为0.11 mg·L−1,对照湖区的平均TP含量为0.34 mg·L−1 (图5(c)、(d)),总磷的削减率达到68%。各月水体中TP质量浓度的变化差异显著(P< 0.001),5月水体中TP含量相对最高,之后进入夏季随着水生植物的生长,其值呈现快速降低的趋势,尤其是8月和9月降低幅度最大,11月达到最低值,到翌年1月,随着有机物的分解,水体中TP质量浓度有所回升。
-
实验处理显著减低了底泥(0~10 cm)中总氮(TN)质量分数(P<0.01)。实验示范区底泥平均TN质量分数为1.43 g·kg−1,对照湖区的底泥平均TN质量分数为1.71 g·kg−1 (图6(a)、(b)),实验处理对底泥总氮的平均削减率为16%。对照湖区底泥TN质量分数在各月之间的变化幅度较小,差异不显著(P=0.194)。实验区底泥中TN质量分数变化较大,5月底泥中TN质量分数相对较高,为2.04 g·kg−1,1月降到0.71 g·kg−1。
与对照区相比,实验示范区底泥中总磷(TP)质量分数显著减低(P<0.001),平均TP质量分数为0.43 g·kg−1,对照湖区底泥平均TP质量分数为0.60 g·kg−1 (图6(c)、(d)),实验处理对底泥中总磷的平均削减率为 28%。对照区各月底泥中TP质量分数变化差异不显著 (P=0.073)。实验区底泥中TP质量分数5月最高,为 0.84 g·kg−1,随着夏季水生植物的生长,其值呈现快速降低的趋势,8月底泥中TP质量分数为 0.33 g·kg−1,1月降到 0.25 g·kg−1。
-
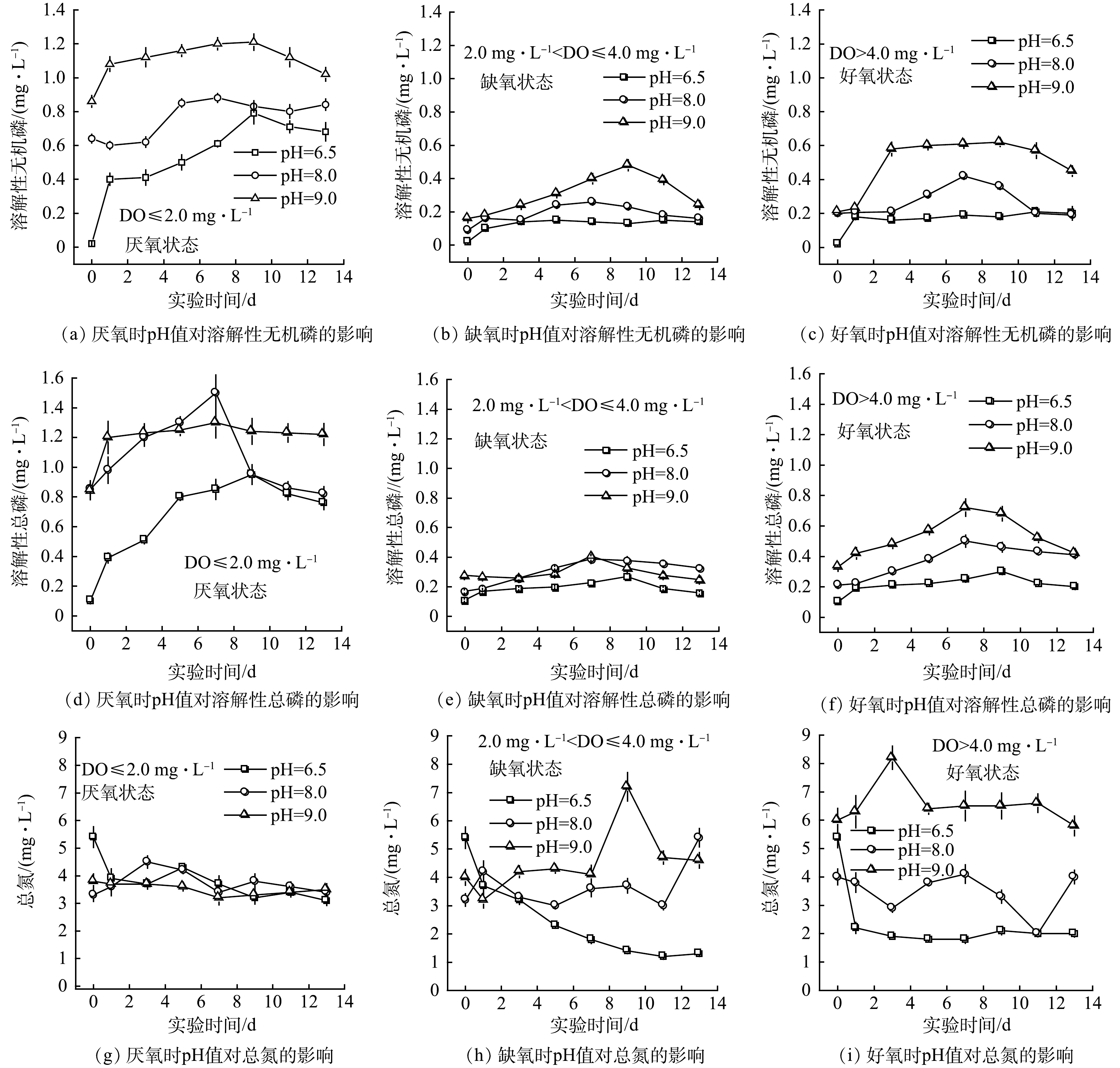
溶解氧浓度显著影响底泥磷的释放(P<0.001),在pH值不变的情况下,溶解氧质量浓度低于2.0 mg·L−1时,上覆水体中溶解态总磷(DTP)的质量浓度显著提高。这说明底泥中磷的释放速度较快。当溶解氧质量浓度为 2.0~4.0 mg·L−1时和高于4.0 mg·L−1时,底泥中磷的释放不显著,水体中DTP的质量浓度较低,两者相差不大(图7)。溶解态无机磷(DIP)质量浓度的变化趋势相似。随pH值的增加,底泥中磷的释放速率显著增加(P=0.003)。在厌氧状态下,pH值从6.5变化到9.0时,底泥中磷的释放速率逐渐增加。在缺氧和好氧状态下,pH值为9.0时,底泥中磷的释放速率有所提高,但仍低于厌氧状态。
DO和pH值的变化显著影响底泥中氮的分解和释放(P<0.001)。在好氧和缺氧状态下,当pH值为6.5时,上覆水体中的总氮质量浓度呈降低趋势;当pH值增加到8.0时,水体中总氮质量浓度比初始值显著增加,猛增后又呈降低趋势;当pH值从8.0增加到9.0时,水体中总氮质量浓度增加后又减少。总之,底泥中氮的分解和释放对pH值比较敏感,变化幅度较大。
-
本研究中原位修复技术显著降低实验示范区湖泊水体的电导率,实验区电导率只有对照区的45%,5—11月电导率呈逐渐降低的趋势,翌年1月有所回升。这表明:实验处理技术可显著降低水体中离子浓度(图4(a)、(b)),在冬季,尽管水面处于冰封状态,但水体中微生物对植物残体和底泥中营养物质的分解仍在进行;实验处理对水温没有显著影响,水温随季节变化而显著变化。本研究发现,乌梁素海的水温在7月最高,平均水温为27.1 ℃,单点最高温度达到31.0 ℃。已有研究[22]表明,随着水体温度的升高,微生物和水生生物活性增强,使底泥中有机磷不断转化为无机磷。但本研究实验区从5月开始,水体中氮磷质量浓度呈逐渐减低的趋势,其原因可能是水草生长过程中能有效吸收释放到水体中的氮磷,使示范区的水体和底泥中氮磷的含量低于对照区(图6(c)、(d)),同时在一定程度上抑制了藻类的生长。其原理包括2个方面。第一,水生植物可以通过自身新陈代谢作用吸收底泥中的营养物质。芦苇和菖蒲(Acorus calamus)对氮的去除效果比较稳定,美人蕉(Canna indica)和芦苇对磷的去除效果较好[23]。挺水植物芦苇生长可固定大量氮和磷,收获后可转移氮磷营养负荷[24]。通过模型模拟发现,挺水植物的存在对氮磷转移具有关键作用[25]。第二,沉水植物对氮磷有很好的吸收作用。使用沉水植物改善水质,不会引入污染物,同时可增强微生物活性而降解污染物,养育水中的食草性鱼类。已有研究[26-27]表明,苦草、黑藻、马来眼子菜、苦草4种沉水植物对水体中TN的去除率均在80%左右,苦草对总磷的去除效果最好,去除率与水体污染物浓度有关。金鱼藻对氮和磷的去除效果也较好[28]。值得一提的是,水生植物修复底泥需要在秋季及时收割打捞,否则到冬春季节植物残体会在水体中被微生物分解,其中所含的营养元素会重新进入底泥中,从而影响修复效果。
磷是控制水体富营养化的关键性与限制性营养盐之一[29]。而且我国近60年来的生产经营活动已经加速了磷的循环[30],西部地区湖泊水体中磷主要来自流域内畜禽养殖业、城乡居民生活污水和农田排水[31]。本研究采用曝气和种植水生植物的方式,显著增加了水体中溶解氧的质量浓度(P<0.001),再加上水生植物生长从水中吸收氮磷,从而降低了乌梁素海水体中营养盐浓度。本实验示范区对水体中TN和TP的削减率达到66%和68%,尽管低于已有研究,溶解氧的增加使上覆水体中氨氮及总磷去除率达到71.6%和 92.6%[32],但已达到地表水III类水的总氮总磷标准;而对照区水体中平均TN、TP质量浓度为2.89 mg·L−1和0.34 mg·L−1,仍为IV类和V类水,甚至超过地表水环境质量标准(GB 3838-2002)中V类标准值。本实验处理对底泥中氮磷的削减率相对水体中比较低,但依然达到显著水平(P<0.01)。其可能原因是:底泥中营养盐的释放一方面受水环境的溶解氧、pH值、微生物活性等因素的影响,另一方面也受底泥中总磷含量较高、水体中营养盐浓度和水生植物吸收转化的影响。乌梁素海底泥平均总磷质量分数为1.07 g·kg−1[33],高于太湖、鄱阳湖等。底泥中磷的削减主要通过植物生长和根系吸收作用,已有研究[34]表明黑藻和苦草可有效降低间隙水中溶解性总磷质量浓度。尽管本实验中种植了苦草、黑藻等沉水植物,可能因种植时间较短,其吸收能力还不足。后期会持续监测沉水植物对底泥中磷质量分数的影响。对武汉南湖的原位生态修复技术表明,围隔构建、鱼类清除、水生植被重建等生态修复措施可改善水质状况,尽管内源污染控制仍需进一步研究[35]。本示范研究经过原位生态修复技术处理后,底泥颜色大部分从黑色转化为黄色,无明显臭味(作者观测)。这表明原位生态修复技术显著降低了水体中氮磷营养物质的质量浓度并改善了水环境条件。构建水生植被、完善食物链、曝气增氧、收获芦苇等措施能有效控制水体富营养化程度。
在实验中,当上覆水体为厌氧条件时,水体中可溶性磷(DIP)质量浓度最高,可达好氧条件下的2倍多(图7),表明好氧环境会抑制底泥中磷的释放。其可能原因:一方面在好氧条件下,底泥中氧化还原电位较高,容易产生胶体沉淀,增强了对磷的固定作用 [36-37];另一方面,在好氧条件下,底泥微生物活性增强,从而降低了底泥中磷的释放速率[38]。底泥-水界面的pH值影响底泥对磷的吸附作用和离子交换作用。已有研究[39]表明,在强碱条件下,总磷的最大释放量是中性条件的1.9倍。从本实验结果看,pH值越高,DIP和DTP的释放越强烈,且高pH值和低溶解氧对DIP和DTP释放有很强的协同作用。本研究实测pH值介于 7.5与9.3之间,底部水体溶解氧质量浓度基本低于2.5 mg·L−1,容易引起底泥磷的释放,如能充分利用水生植物生长吸收释放出的磷,在秋季及时收获并带出水体可有效降低湖泊的富营养化程度。
-
1) 原位生态修复技术显著降低了示范区水体的总氮(TN)和总磷(TP)质量浓度,与对照湖区相比,总氮和总磷的削减率分别达到66%和68%。5月水体中TN和TP质量浓度相对较高,随着水生植物生长,其质量浓度显著降低,并在11月达到最低值;翌年1月,其质量浓度有所回升,这可能是由于植物体的腐烂分解将部分N、P释放到水体中。
2) 原位生态修复技术显著降低了示范区底泥(0~10 cm)中总氮(TN)和总磷(TP)质量分数,对0~10 cm 底泥中总氮和总磷的削减率分别为16%和28%。5月底泥中TN和TP质量分数较高,翌年1月质量分数最低。
3) 较低的溶解氧和较高的pH值(碱性环境)会加速底泥中溶解性无机磷和溶解性总磷的分解和释放,有利于水生植物吸收利用,但会抑制氮的释放。
致谢 感谢北京爱尔斯生态环境工程有限公司武学军、宋国庆、孟路在野外实验实施和采集样品工作中给予的帮助!
乌梁素海水体富营养化原位生态修复实验及效果分析
Experiment study and effect analysis of in situ ecological restoration for eutrophication in Wuliangsu Lake, China
-
摘要: 为探讨原位生态修复技术对乌梁素海水体富营养化的治理效果,在乌梁素海农田排水入湖口作业区用双层防水围隔建立了实验示范区,测定了水体中氮磷质量浓度和底泥中氮磷质量分数;在室内进行模拟实验,研究了溶解氧(DO)和pH值变化下柱状底泥营养盐的静态释放规律。室外实验结果表明:示范区水体中总氮(TN) 和总磷(TP)质量浓度显著低于对照区(P<0.001),削减率分别为66%和68%,水体平均TN质量浓度为0.99 mg·L−1,TP质量浓度为0.11 mg·L−1;就水质年度变化而言,5月份水体TN和TP质量浓度相对较高,随着水生植物生长,其质量浓度显著降低,11月达到最低值,翌年1月,其值有所回升;实验示范区底泥中的平均TN和TP质量分数分别为1.43 g·kg−1和0.43 g·kg−1,显著低于对照区的数值(P<0.01),削减率分别为16% 和28%。室内模拟实验结果表明:当DO质量浓度低于2.0 mg·L−1时,底泥中磷的释放速度较快;当DO质量浓度高于2.0 mg·L−1时,随着pH值的增加,底泥中磷的释放速率显著增加(P<0.01);在厌氧状态下,pH值对底泥氮释放影响较小;在好氧和缺氧状态下,底泥中氮的释放速率显著增加后又下降;DO和pH值对底泥氮磷释放有很强的协同作用。综上所述,原位生态修复技术可显著降低水体和底泥中氮磷营养盐质量浓度,达到净化水质,减缓内源污染的目的,有望成为治理乌梁素海富营养化的一种有效措施。Abstract: In order to explore the effect of in situ ecological restoration technology on the eutrophication control of Wuliangsu Lake, an experimental demonstration area and a double-layer waterproof enclosure were established in the operation area of farmland drainage into the entrance of Wuliangsu Lake, and the mass concentrations of nitrogen and phosphorus in the water body and sediment were measured. A simulation experiment was set up in the laboratory to study the static release law of nutrients in columnar sediments with the changes of dissolved oxygen (DO) and pH. The outdoor experiment results showed that the mass concentrations of total nitrogen (TN) and total phosphorus (TP) in the water body of the demonstration area were significantly lower than those of the control area (P<0.001), their reduction rates were 66% and 68%, respectively, and their average mass concentrations were 0.99 mg·L−1 and 0.11 mg·L−1, respectively; In terms of annual change in water quality, the mass concentrations of TN and TP in the water body were relatively high in May, then decreased significantly with the growth of aquatic plants, reached the lowest values in November, and rose again the following January. the average mass fractions of TN and TP in the sediments of the experimental demonstration area were 1.43 g·kg−1 and 0.43 g·kg−1, respectively, which were significantly lower than the values in the control area (P<0.01), their reduction rates were 16% and 28%, respectively. The results of laboratory simulation experiments showed that: when the DO concentration was lower than 2.0 mg·L−1, the release rate of phosphorus in the sediment was faster; when DO concentration was higher than 2.0 mg·L−1, the release rate of phosphorus in the sediment increased significantly with the increase of pH value (P<0.01). Under anaerobic conditions, pH value had slight effect on nitrogen release from sediment. Under aerobic and anoxic conditions, the release rate of nitrogen in the sediment increased significantly at first, and then decreased. DO and pH had a strong synergistic effect on the release of nitrogen and phosphorus in the sediment. To sum up, in situ ecological restoration technology can significantly reduce the concentrations of nitrogen and phosphorus nutrients in water and sediment, achieve the purpose of purifying water quality and slowing down endogenous pollution, and is expected to become an effective measure to control the eutrophication of Wuliangsu Lake, China.
-
近年来,我国城镇居民用水量不断增大,给水厂自来水产量也随之提高[1]。在自来水生产的同时,会产生大量的污泥,这部分污泥源于生产工艺中的沉淀池排泥水和滤池反冲洗水,主要为原水中的无机颗粒、有机物以及净水过程中投加的混凝剂[3]。其产量约占自来水总生产水量的4%~7%,为避免资源浪费,应设法将其回收利用[2-4]。
化学调理脱水是污泥脱水常用的方法[5]。常用的化学调理剂有PAM等有机高分子调理剂和铁系、铝系等无机调理剂[6]。PAM在较少的投加量下便可达到较理想的效果,如给水厂中常用阳离子PAM来对污泥进行调理,但其具有轻微毒性,泥水分离后的水不能回用,会对给水厂造成较大的损失。铁系混凝剂絮体形成快,但本身不稳定,且投加到水体中存在色度问题[7-8]。对于铝系混凝剂而言,由于铝形态的不同,其絮凝效果和机理也相应不同[9]。羟基铝盐絮凝剂的形态可分为Ala、Alb、Alc。Ala为低聚态铝,如单体铝AlCl3;Alb为中聚态铝,如Al13(分子式为[AlO4Al12(OH)24(H2O)12]7+);Alc为高聚态铝,如Al30。相对于单体铝来讲,聚合铝具有投加量少、污泥产生量少、电中和能力高、对pH和温度有更高的适应能力等优点[10]。Al13是铝离子水解过程的中间产物,是一种高电荷的纳米粒子。与其他混凝剂相比,Al13电中和能力更强,其在混凝、除氟和污泥调理等方面均有应用[11]。刘沛[8]和CAO等[9]分别将AlCl3和Al13应用于市政污泥脱水,发现经2者调理后的污泥脱水性能均有提高,而且Al13的效果更为显著。
本研究采用无机盐调理剂AlCl3与自制Al13溶液对给水厂污泥进行调理,探究不同的药剂投加量对毛细脱水时间和污泥比阻的影响,并分析污泥调理过程中絮体粒径及形态的变化,从而得出2种调理剂形成的絮体特征;最后,结,2种药剂对有机物的去除率、调理后污泥上清液的余铝含量,比较其实际应用潜力。
1. 材料与方法
1.1 供试污泥和试剂
本研究所用污泥取自北京市某给水厂污泥浓缩池,该水厂以南水北调南干渠原水为水源,日产水量50×104 t。污泥基本性质见表1。
表 1 污泥基本性质Table 1. Basic properties of sludge含水率/% 有机质/% pH CST/s d(0.5)/μm SRF×1013/(m·kg−1) 98.5 30.9 7 77.5 51.13 1.63 注:d(0.5)表示污泥中体积累积百分比为50%时颗粒的最大直径。 | Show Table DownLoad:
CSV
DownLoad:
CSV
六水合氯化铝(AlCl3·6H2O)、氢氧化钠(NaOH)、硫酸钠(Na2SO4)、氯化钡(BaCl2)均为分析纯;铝十三溶液(Al13,Al含量9.6 g·L−1)于实验室采用慢速滴碱法自制[12]。经Ferron比色法[9]检测,AlCl3和Al13溶液的Ala、Alb、Alc含量分别为95.32%、3.91%、0.77%和3.44%、93.96%、2.60%。
1.2 实验及检测方法
调理方法。烧杯搅拌实验在六联搅拌器上进行,6个烧杯中分别倒入500 mL污泥,AlCl3的投加量(以Al含量计)为0.300、0.600、0.800、0.900、1.000、1.200 g·L−1;Al13投加量(以Al含量计)分别为0.024、0.096、0.192、0.288、0.336、0.360 g·L−1。搅拌器程序设定为:250 r·min−1快搅30 s,加药后200 r·min−1搅拌1 min,再以40 r·min−1慢搅10 min。
毛细吸水时间(CST)由CST测定仪(RTC-304B,英国Triton公司)直接测定;污泥比阻(SRF)在压力为0.6 MPa下将一定量污泥倒入布氏漏斗中抽滤测定[14];含水率和有机质使用电热恒温干燥箱(DHG-9146A,北京天林恒泰科技有限公司)和箱式电阻炉(SX2-2 5-10TP,上海一恒科学仪器有限公司)测定[13]。
污泥上清液有机物的测定。参考NIU等[14]分离EPS的方法分步分离有机物,用荧光分光光度计(F-7000,日立公司)测定三维荧光谱图,采用150 W氙灯为激发光源,激发波长(Ex)200~400 nm,发射波长(Em)220~550 nm,狭缝宽度5 nm。DOC由总有机碳测定仪(岛津,岛津公司)测定。
污泥上清液余铝的测定。调理后的污泥上清液过0.45 μm膜,稀释一定倍数用电感耦合等离子光谱仪(9800,岛津公司)测定。
絮体粒径和分形维数的动态变化用马尔文激光粒度分析仪(Mastersizer 2000,马尔文公司)测定。混凝程序:A段快搅阶段,250 r·min−1快搅30 s,加AlCl3或Al13后200 r·min−1搅拌1 min;B段慢搅阶段,40 r·min−1慢搅15 min;C段破碎阶段,250 r·min−1快搅1 min破碎絮体;D段恢复阶段,40 r·min−1慢搅15 min使絮体再生成,参考SUN等[15]的方法计算分形维数。
污泥絮体微观结构用场发射扫描电子显微镜(SU-8020,日立公司)观察。
2. 结果与讨论
2.1 AlCl3和Al13调理对污泥脱水性能的影响
图1为经不同浓度Al13和AlCl3调理后污泥的CST和SRF。由图1(a)可知,随着Al13投加量的逐渐增加,污泥CST以较快的速度下降,在投加量为0.336 g·L−1时,CST从79.1 s减小到33.1 s,再继续增加投加量,CST反而上升;相比之下,随着AlCl3投加量的逐渐增加,CST下降速度较为缓慢,当其降至最小值34.8 s时,所对应的最佳投加量为0.800 g·L−1,继续增加投加量,CST缓慢上升。由图1(b)可知,Al13投加量为0.024和0.096 g·L−1时,SRF改善效果不明显,继续增加到0.336 g·L−1时,SRF改善效果最佳,为4.98
× 1012 m·kg−1,继续增加投加量,SRF增大,污泥脱水效果变差;AlCl3投加量为0.300 g·L−1时,SRF达到6.53× 1012 m·kg−1,继续增加投加量至0.600、0.800、0.900 g·L−1,SRF维持在5.50× 1012 m·kg−1左右,最小值为5.45× 1012 m·kg−1,再继续增加投加量,污泥脱水效果变差。在最佳投加量的基础上继续增加投加量,CST和SRF效果都变差,这是因为调理剂投加过多,会引起颗粒复稳,脱水性能将变差[16]。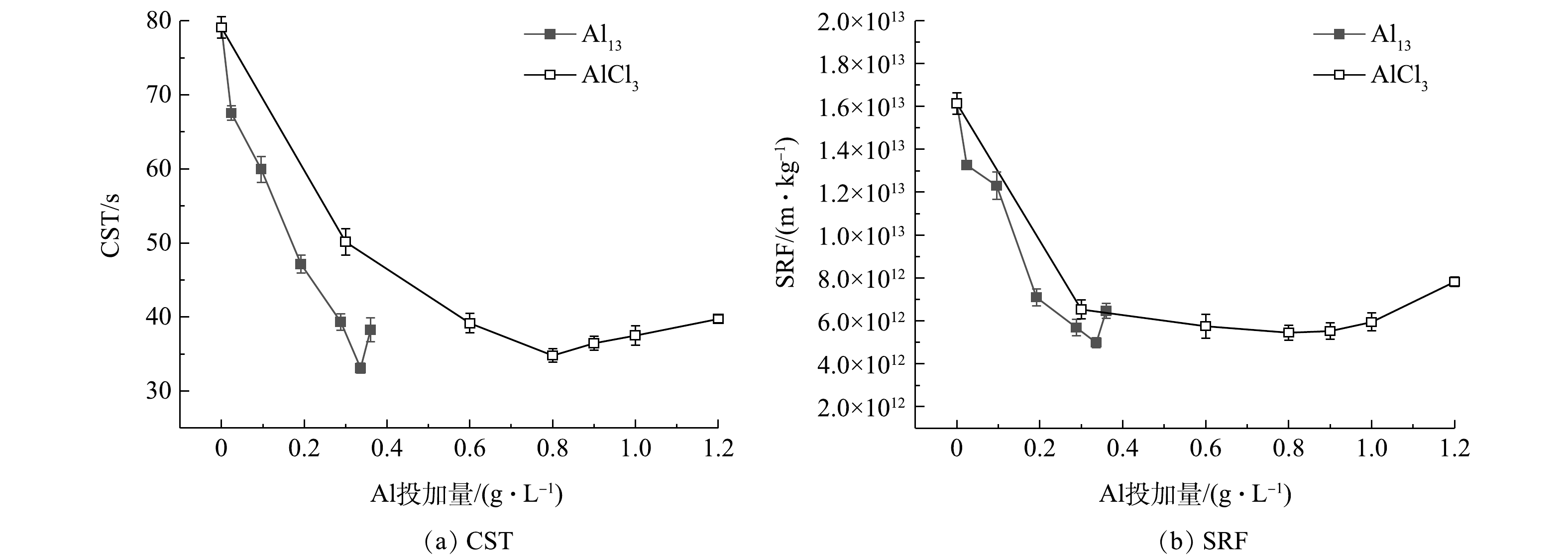 图 1 不同浓度Al13和AlCl3调理后的污泥的CST和SRFFigure 1. CST and SRF of sludge treated with different concentrations of Al13 and AlCl3
图 1 不同浓度Al13和AlCl3调理后的污泥的CST和SRFFigure 1. CST and SRF of sludge treated with different concentrations of Al13 and AlCl32.2 AlCl3和Al13调理对污泥中有机物的影响
给水厂污泥主要成分为原水中的无机颗粒、有机物以及净水工艺中投加的混凝剂[17]。污泥中水分分为自由水、毛细结合水、表面吸附水和内部结合水[18]。与污水厂污泥不同,给水厂污泥微生物含量较少,内部结合水的含量不足以影响污泥脱水效能,因此在本研究中可忽略。有机物相对于无机颗粒更易于与水分结合,其存在状态在给水厂污泥中有3种:有机物单独存在、有机物吸附在无机颗粒上、有机物被净水过程中投加的混凝剂包裹。经过Al13和AlCl3调理后,有机物与调理剂结合,结合在有机物上的水分子脱出。
图2是原泥及AlCl3、Al13最佳投加量调理后的污泥上清液有机物三维荧光图,原泥游离的有机物中主要呈现2个峰:Ex/Em 265-290 nm/320-375 nm、252-272 nm/430-440 nm,分别为微生物代谢产物峰和腐殖酸峰[9]。原泥和Al13、AlCl3调理过的污泥上清液各区域的荧光强度按照游离的有机物、吸附在无机颗粒的有机物、被混凝剂包裹的有机物的顺序依次减小,这说明有机物含量逐渐降低。Al13调理过的污泥游离有机物的荧光强度低于AlCl3,而混凝剂包裹的有机物荧光强度略高于AlCl3,因此Al13对游离有机物去除率相对较高,对混凝剂包裹的有机物去除率相对较低。有研究表明,污泥脱水性能与蛋白质含量相关[19]。蛋白质是污泥絮体的主要组成部分[20],其水化层会包围水相中的水分,使这部分水不易排出[21]。三维荧光图谱中Ex/Em 200-250 nm/280-380 nm为蛋白质区域[9],由图2(a)、图2(d)、图2(g)可知,Al13调理后的污泥上清液在蛋白质区域的荧光强度相对原泥明显减少,而AlCl3变化较小,因此Al13对蛋白质的去除效果优于AlCl3。该结果验证了污泥比阻实验中,Al13调理后的污泥易脱水的结论。
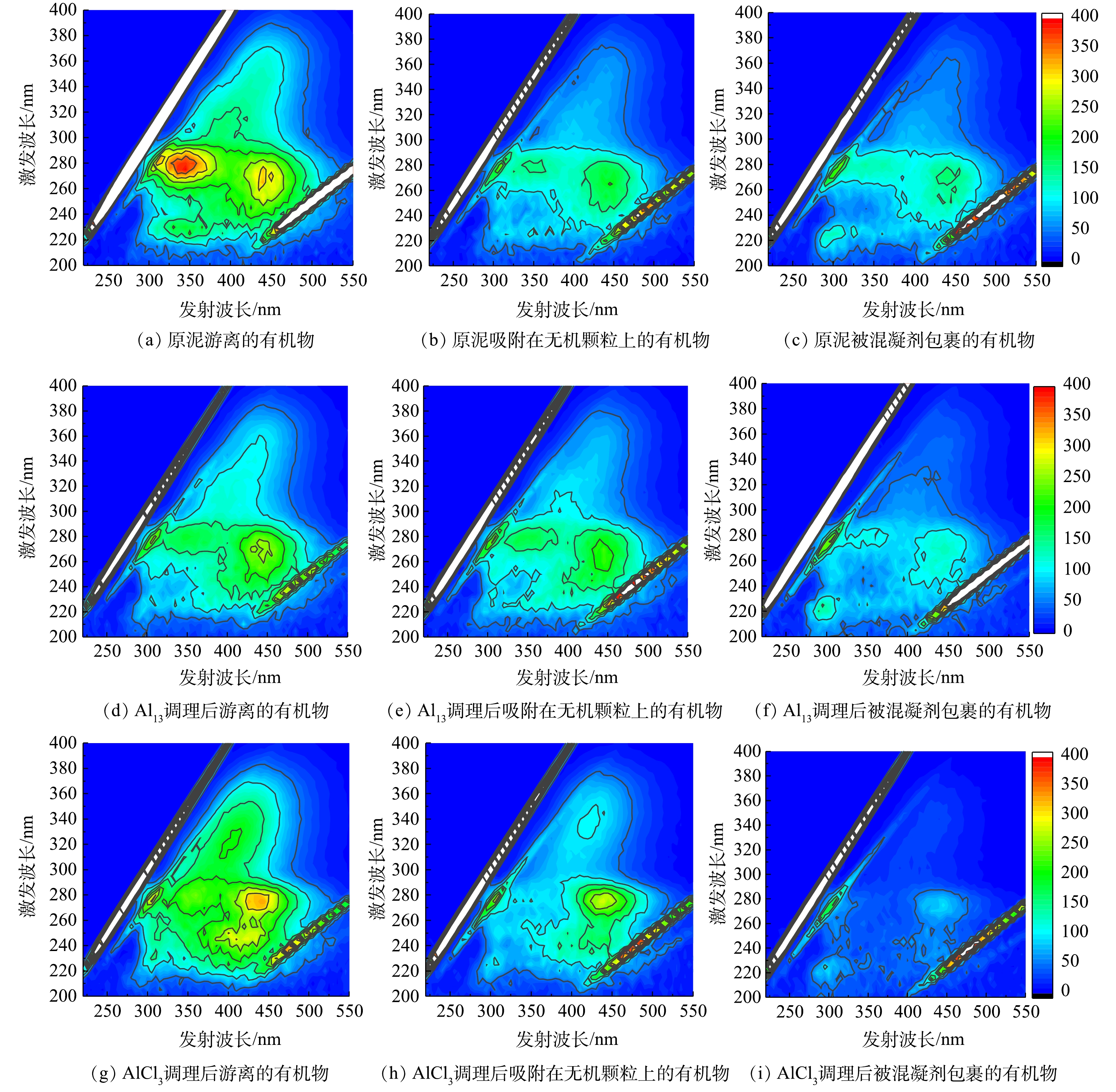 图 2 原泥以及Al13和AlCl3最佳投加量调理后污泥上清液的三维荧光图谱Figure 2. 3D-EEM spectra of sludge supernatant about raw sludge and conditioned sludge using Al13、AlCl3
图 2 原泥以及Al13和AlCl3最佳投加量调理后污泥上清液的三维荧光图谱Figure 2. 3D-EEM spectra of sludge supernatant about raw sludge and conditioned sludge using Al13、AlCl3图3为原泥和Al13、AlCl3调理过的污泥上清液不同层次有机物的DOC浓度(以总悬浮物(TSS)计)。图中3种污泥上清液的浓度按照游离的有机物、吸附在无机颗粒上的有机物、被混凝剂包裹的有机物的顺序减少,与三维荧光谱图中的趋势相同。经Al13和AlCl3调理过后的污泥各组分DOC浓度均有不同程度降低。原泥和Al13、AlCl3调理后的污泥游离有机物DOC浓度分别为2.16和1.03、2.15 mg·g−1,Al13调理过后的污泥游离有机物DOC浓度比原泥降低了1.13 mg·g−1,而AlCl3调理过后的污泥游离有机物DOC浓度仅比原泥降低了0.01 mg·g−1,因此Al13调理过后的污泥更易脱水。
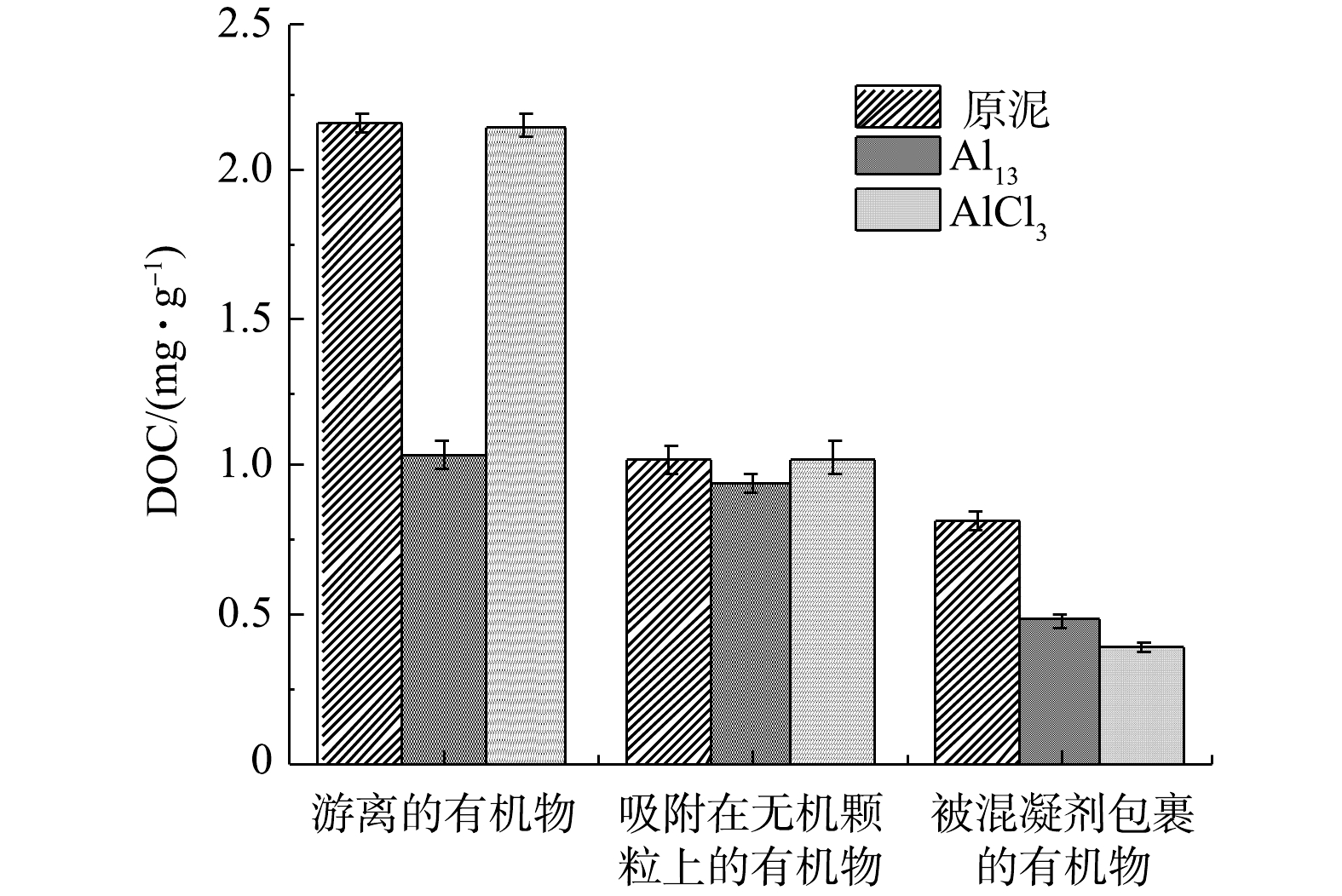 图 3 原泥和Al13、AlCl3调理后的污泥不同上清液的DOC值Figure 3. DOC content of raw sludge and conditioned sludge using Al13、AlCl3
图 3 原泥和Al13、AlCl3调理后的污泥不同上清液的DOC值Figure 3. DOC content of raw sludge and conditioned sludge using Al13、AlCl32.3 AlCl3和Al13调理对污泥絮体性质的影响
图4是在AlCl3和Al13最佳投加量时,污泥随时间变化的絮体粒径(d(0.5))和分形维数。原泥粒径为52.42 μm,快搅30 s后加入Al13和AlCl3,絮体粒径均有所增长,在快搅阶段投加Al13的污泥形成的絮体粒径平均为57.29 μm,大于加入AlCl3的54.42 μm;慢搅阶段加入AlCl3的絮体粒径增长较快,最大能够增长到126.83 μm,稳定后平均粒径为110.47 μm,而加入Al13的污泥絮体大小则较稳定,一直保持在82.00 μm左右,稳定后平均粒径为81.70 μm;加入AlCl3和Al13的污泥絮体破碎阶段的粒径分别为88.83和69.96 μm;AlCl3在破碎后再生成的絮体稳定后平均粒径为137.39 μm,大于破碎前的粒径,Al13为78.36 μm。AlCl3和Al13的强度因子分别为80.41%和85.63%,恢复因子分别为224.40%和71.79%。强度因子越高,絮体抗剪切能力越强,恢复因子越高,絮体破坏后越容易重新团聚成絮体,因此Al13形成的污泥絮体抗剪切能力更强,AlCl3形成的絮体受到破坏后更容易恢复[22]。
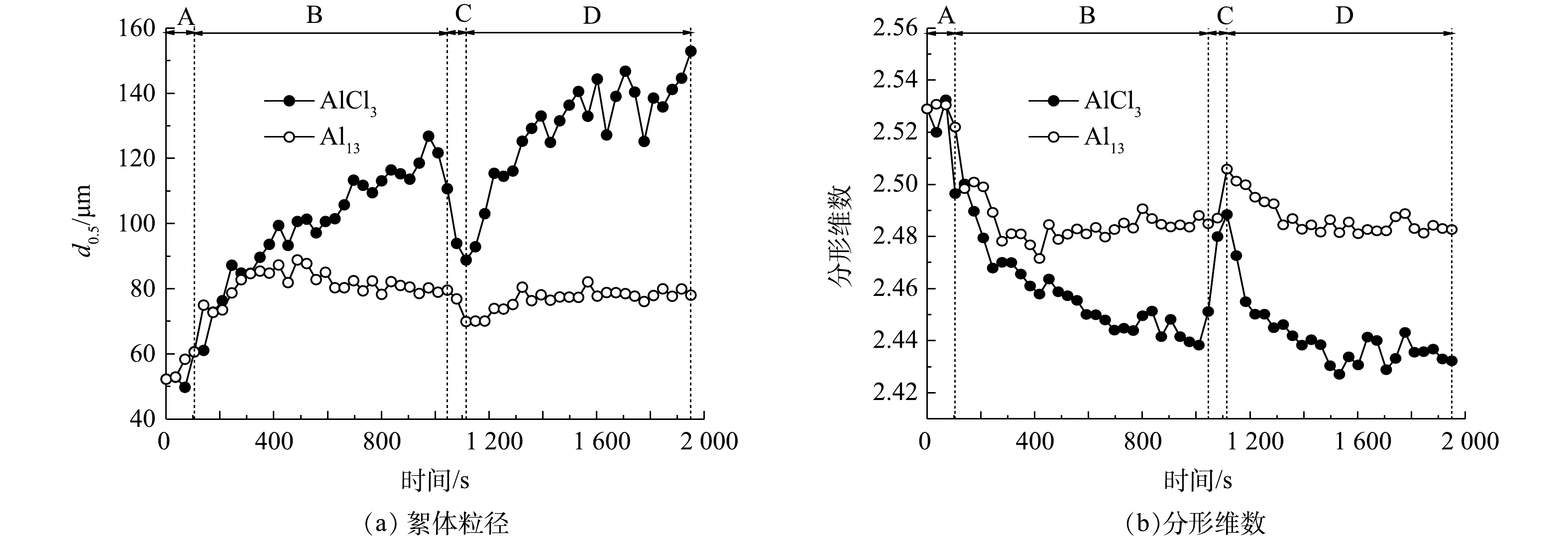 图 4 Al13、AlCl3最佳投加量时污泥的絮体粒径和分形维数变化Figure 4. Particle size and fractal dimension of flocs under the optimal dosage of Al13 and AlCl3
图 4 Al13、AlCl3最佳投加量时污泥的絮体粒径和分形维数变化Figure 4. Particle size and fractal dimension of flocs under the optimal dosage of Al13 and AlCl3分形维数是表示分形体不规则的量度[23],分形维数越高,絮体越紧实[24]。从图4中可看出,Al13和AlCl3的分形维数稳定时分别在2.48和2.44左右,相差不大,2者絮体密实程度无明显差别。从图5扫描电镜图中可以看出,原泥表面光滑,絮体规则而紧密,经AlCl3和Al13调理过后的污泥絮体松散。
 图 5 原泥和AlCl3、Al13最佳投加量时污泥絮体的扫描电镜Figure 5. SEM of raw sludge and conditioned sludge under the optimal dosage of Al13 and AlCl3
图 5 原泥和AlCl3、Al13最佳投加量时污泥絮体的扫描电镜Figure 5. SEM of raw sludge and conditioned sludge under the optimal dosage of Al13 and AlCl32.4 AlCl3和Al13调理对污泥上清液性质的影响
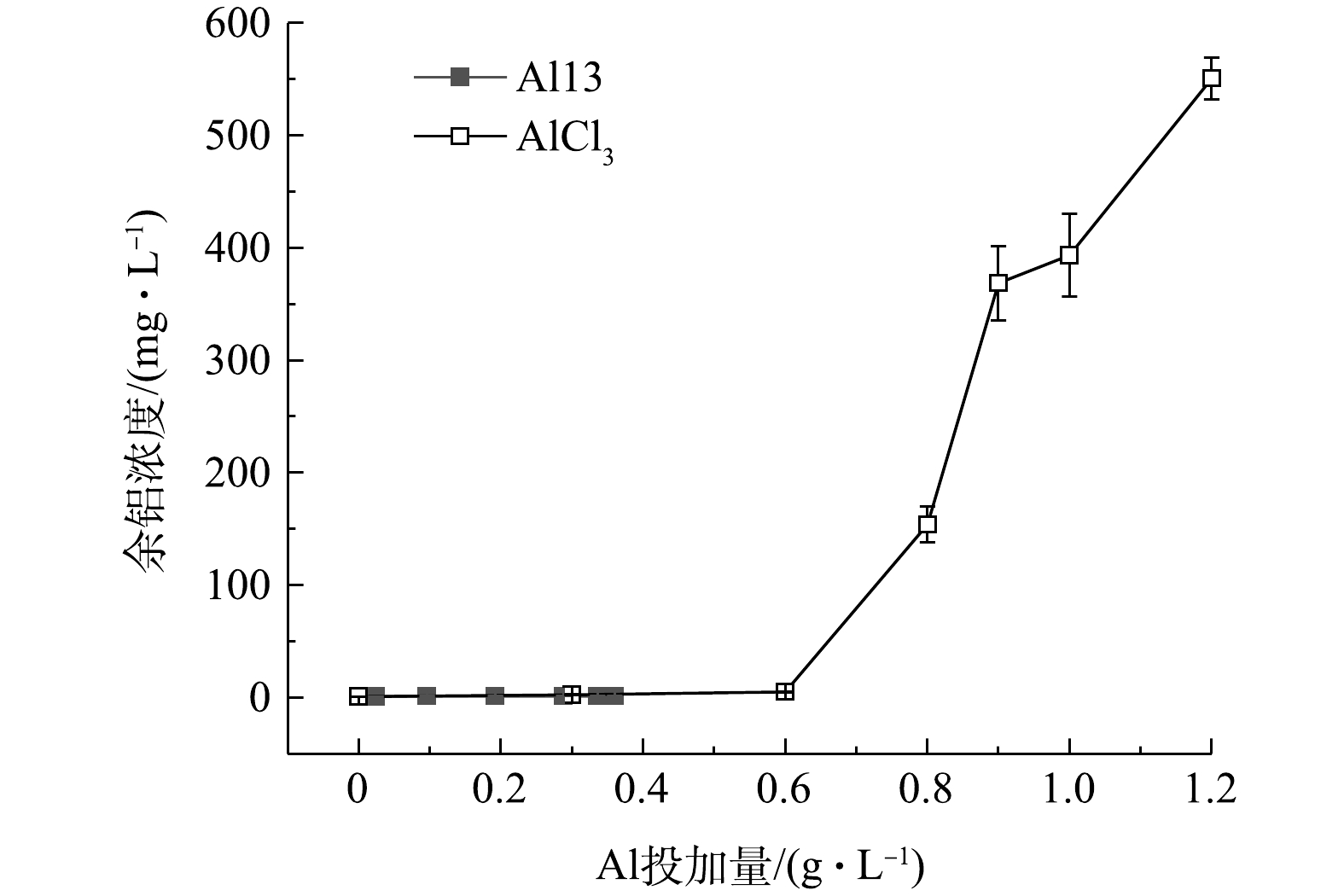
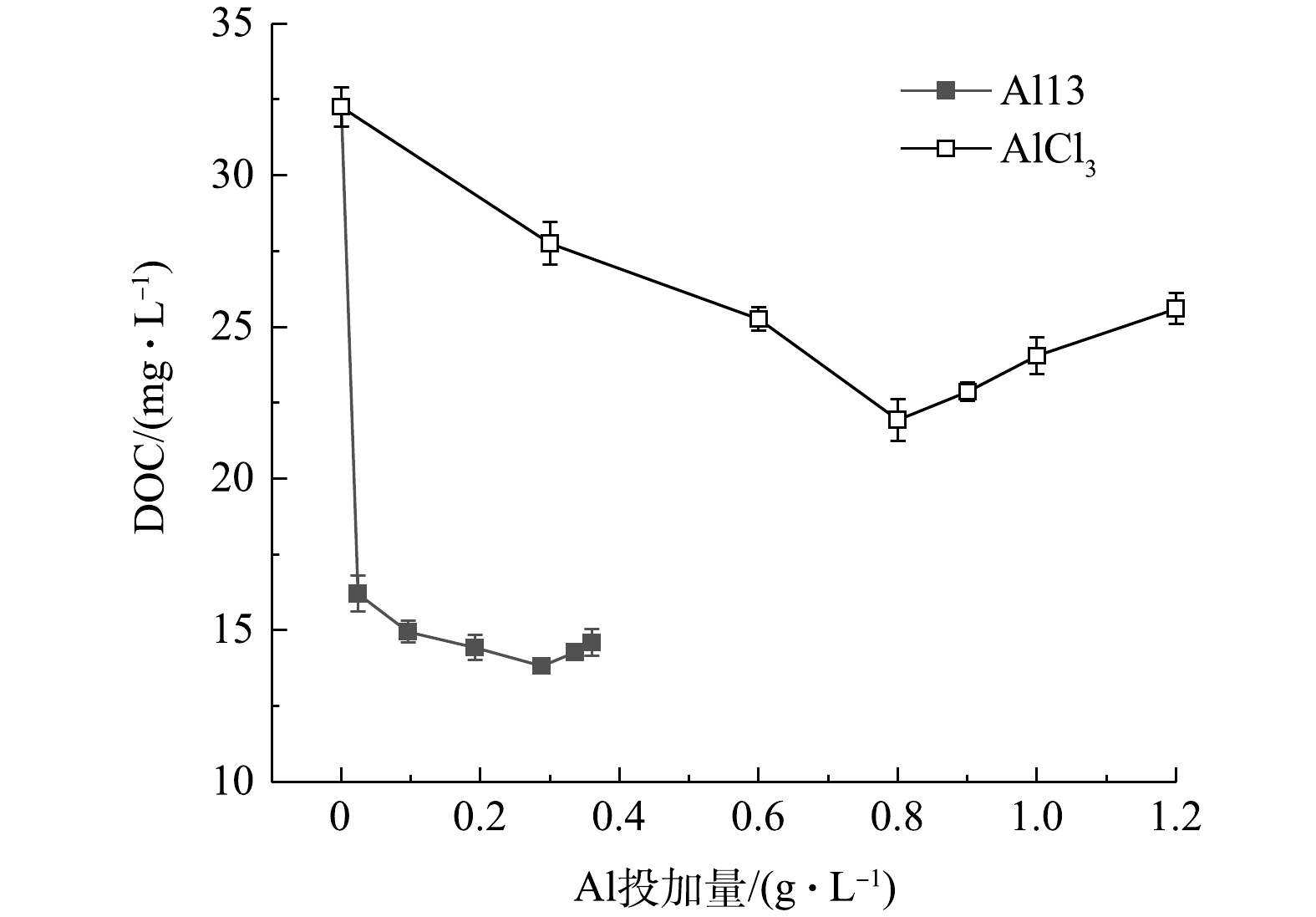
图6是Al13和AlCl3不同投加量下的污泥上清液余铝浓度,Al13调理后的污泥上清液余铝浓度最高为1.35 mg·L−1,远远低于AlCl3调理后的污泥上清液余铝浓度。未投加铝盐混凝剂的原泥上清液中余铝浓度为0.83 mg·L−1。这是因为,制水工艺中投加的铝盐混凝剂沉降下来,富集了这部分混凝剂的沉后水污泥排入污泥池中,使得污泥中铝离子含量偏高。因此,如何在保证调理效果的基础上降低余铝浓度,是下一步要解决的重要问题。图7为投加2种调理剂后溶液DOC含量的变化,原泥DOC浓度为32.25 mg·L−1,Al13的加入使得DOC含量迅速下降到16.21 mg·L−1,继续投加,DOC变化幅度较小,最佳投加量时为14.27 mg·L−1;随着投加量的增加,AlCl3调理后的污泥DOC变化较为平缓,最佳投加量时的浓度为21.93 mg·L−1,再继续增加投加量,DOC浓度变高,这是因为投加量过多,体系pH下降,引起了颗粒的复稳。考虑到污泥脱水后脱出水的回用,如果投加Al13的污泥上清液余铝和溶解性有机物含量更低,则回用后续处理将会更简单。
2.5 AlCl3和Al13污泥调理机制
综合前4节的分析可知,Al13和AlCl3加入给水厂污泥后,与污泥中的有机质相互作用,降低了污泥体系中有机物的含量,使得污泥颗粒聚集成尺寸更大、结构更疏松的絮体,最终造成水分更容易脱出。同时发现,有机物的含量与污泥脱水性能联系密切,在加入Al13的污泥中,有机物去除量更高,因此该调理剂的脱水性能优于AlCl3。图8为Al13、AlCl3与有机物的相互作用模型,AlCl3(Ala)可通过络合作用与给水污泥中的有机物结合,并形成Ala-有机物络合物,由于给水污泥中的有机物来源于天然水体,其中的大部分有机物分子量较小,不足以给Ala形成的絮凝体提供凝核,因此部分Ala-有机物络合物未能沉淀;相比之下,带有更多正电荷的Al13(Alb)不仅能够有效中和有机物的负电荷,还可作为核团与可溶性有机物形成絮体[25]。因此,Al13调理后的污泥相对AlCl3来讲,上清液中的有机物浓度更低。
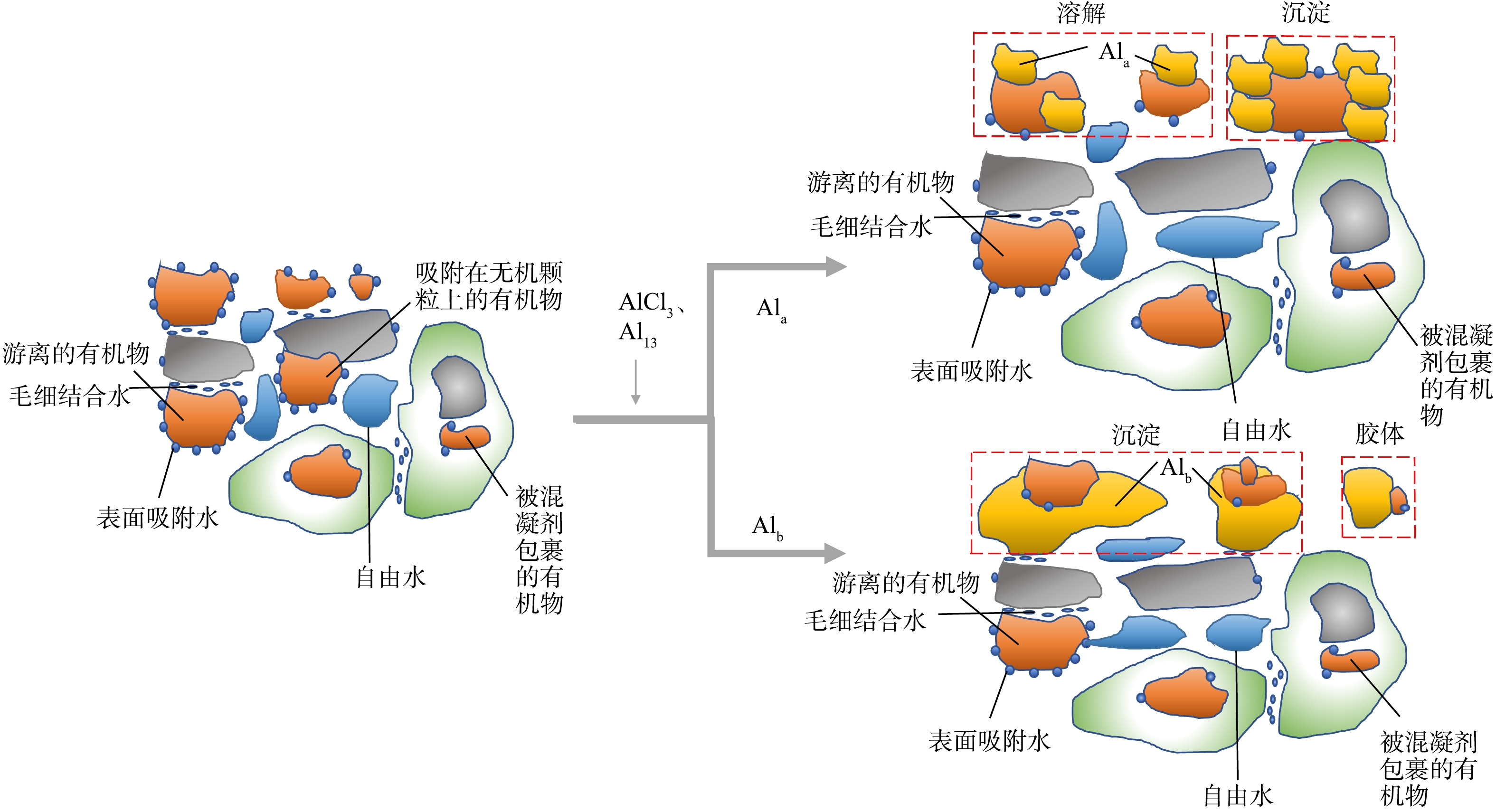 图 8 Al13、AlCl3与有机物相互作用模型Figure 8. Model of interaction of Al13、AlCl3 and organic matters
图 8 Al13、AlCl3与有机物相互作用模型Figure 8. Model of interaction of Al13、AlCl3 and organic matters3. 结论
1)以Al含量计,Al13和AlCl3的最佳投加量分别为0.336和0.800 g·L−1,此时Al13的CST和SRF均小于AlCl3,Al13在投加量较少的情况下便可取得良好的脱水效果。调理后的污泥絮体相对原泥更松散,有利于水分的脱出。
2)相比于AlCl3,Al13更能有效地与有机物形成沉淀,调理后的污泥有机物含量更低,在蛋白质含量降低的体现上尤为突出。Al13调理后的污泥上清液余铝浓度也远远低于AlCl3,更利于给水厂的回用。
-
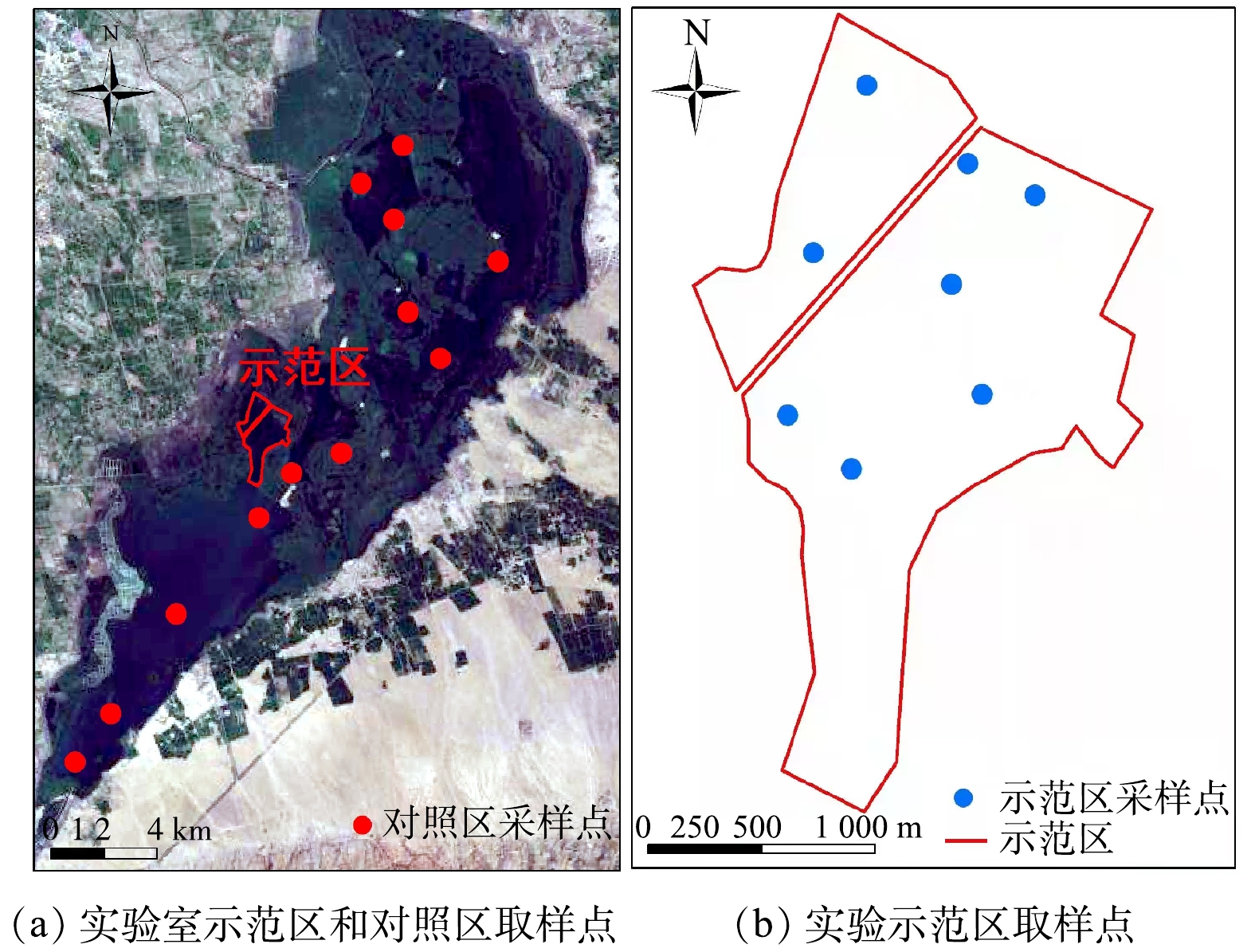
图 1 实验区地理位置及采样点示意图
Figure 1. Geographic location and sketch map of sampling points in the experimental area

图 2 实验示范研究的总体思路、野外实验现场、太阳能曝气设施
Figure 2. General idea frame diagram of the experimental demonstration research, field experiment scene and schematic diagram of solar aeration facility
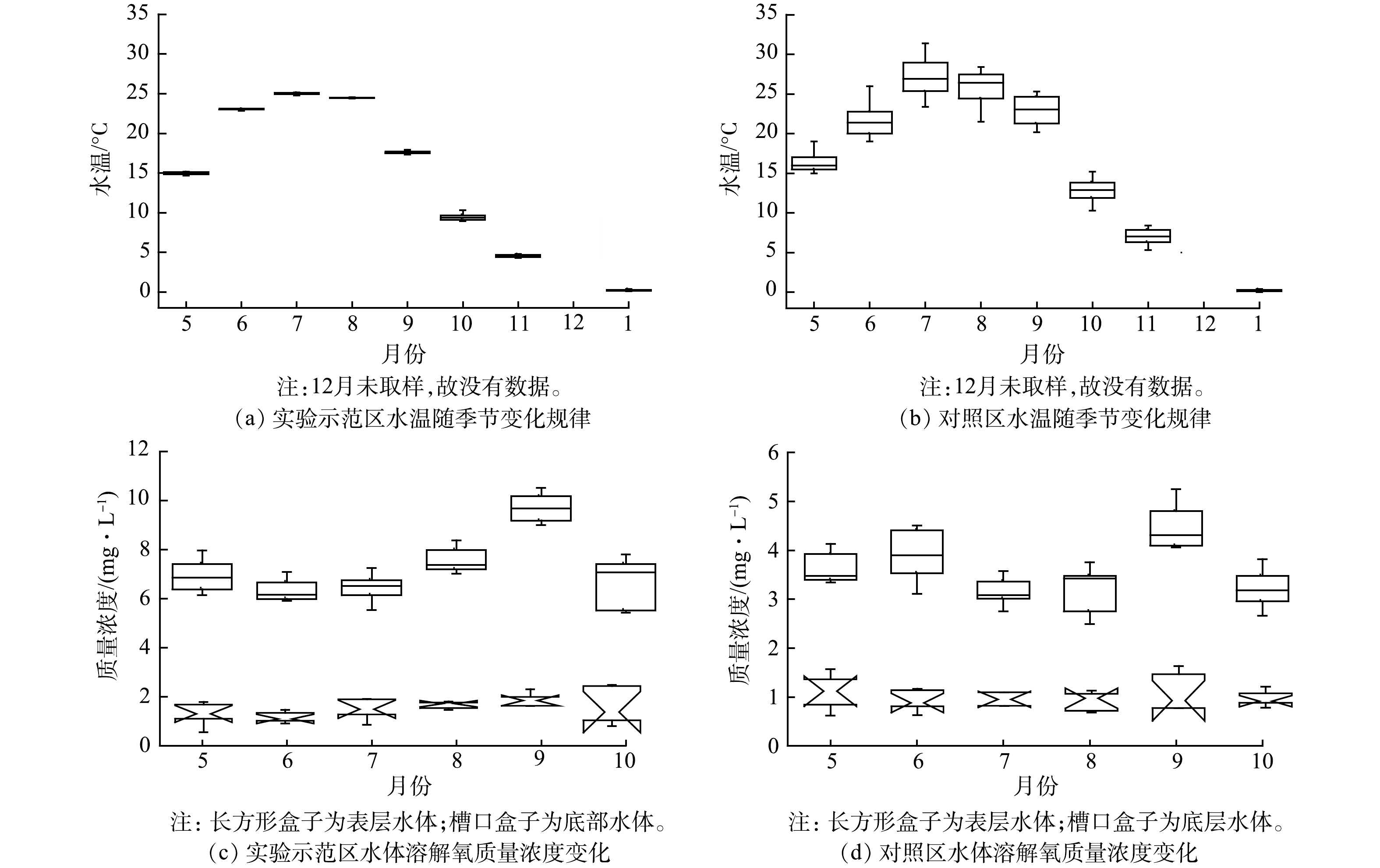
图 3 乌梁素海实验示范区和对照区水温随季节变化的规律及水体表层和底部溶解氧质量浓度变化
Figure 3. Seasonal variation of temperature, dissolved oxygen mass concentration at surface and bottom of water body at experimental demonstration area and the control area of Wuliangsu Lake

图 4 乌梁素海实验示范区和对照区水体电导率和pH值随季节变化的规律
Figure 4. Seasonal variation of water conductivity and pH value in experimental demonstration area and the control area of Wuliangsu Lake

图 5 乌梁素海实验示范区和对照区水体总氮、总磷质量浓度随季节变化规律
Figure 5. Seasonal variation of total nitrogen and total phosphorus mass concentrations in water at experimental demonstration area and the control area of Wuliangsu Lake
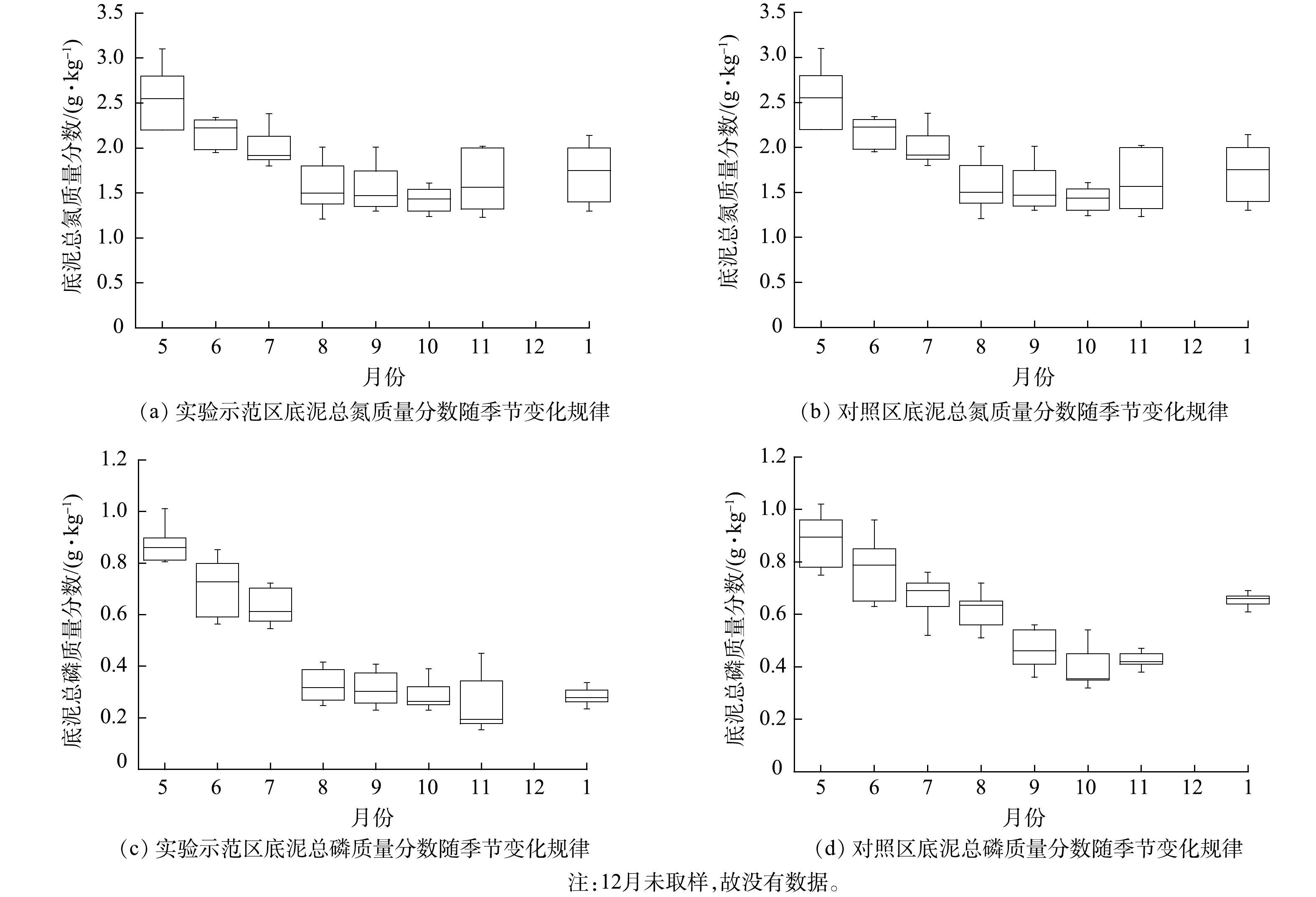
图 6 乌梁素海实验示范区和对照湖区底泥(0~10 cm)总氮、总磷质量分数随季节变化规律
Figure 6. Seasonal variation of total nitrogen and phosphorus mass fraction in sediment (0~10 cm) of the experimental demonstration area and the control area of Wuliangsu Lake
-
[1] 李干杰. 中国环境质量状况公报[M]. 北京: 中华人民共和国生态环境部, 2018. [2] CONLEY D J, PAERL H W, HOWARTH R W, et al. Controlling eutrophication: nitrogen and phosphorus[J]. Science, 2009, 323: 1014-1015. doi: 10.1126/science.1167755 [3] MARTINS G, PEIXOTO L, BRITO A G, et al. Phosphorus-iron interaction in sediments: can an electrode minimize phosphorus release from sediments? [J] Reviews in Environment Science and Bio/Technology, 2014, 13: 265-275. [4] 延霜. 水体-沉积物界面氮迁移转化的生物化学过程[D]. 西安: 西安建筑科技大学, 2010. [5] YIN H B, HAN M X, KONG M, et al. Influence of sediment resuspension on the efficacy of geoengineering materials in the control of internal phosphorus loading from shallow eutrophic lakes[J]. Environmental Pollution, 2016, 219: 568-579. doi: 10.1016/j.envpol.2016.06.011 [6] MCGOLDRICK D J, MURPHY E W. Concentration and distribution of contaminants in lake trout and walleye from Laurentian Great Lakes (2008-2012)[J]. Environmental Pollution, 2016, 217: 85-96. doi: 10.1016/j.envpol.2015.12.019 [7] WU Y H, WEN Y J, ZHOU J X, et al. Phosphorus release from lake sediments: Effects of pH, temperature and dissolved oxygen[J]. KSCE Journal of Civil Engineering, 2014, 18(1): 323-332. doi: 10.1007/s12205-014-0192-0 [8] SCHINDLER D W. Evolution of phosphorus limitation in lakes[J]. Science, 1977, 195: 260-262. doi: 10.1126/science.195.4275.260 [9] SCHELSKE C L. Eutrophication: Focus on phosphorus[J]. Science, 2009, 324: 722-722. doi: 10.1126/science.324_722 [10] ABELL J M, ÖZKUNDAKCI D, HAMILTON D P. Nitrogen and phosphorus limitation of phytoplankton growth in new Zealand Lakes: Implications for eutrophication control[J]. Ecosystems, 2010, 13: 966-977. doi: 10.1007/s10021-010-9367-9 [11] 李卫平, 李畅游, 史小红, 等. 内蒙古乌梁素海氮、磷营养元素分布特征及地球化学环境分析[J]. 资源调查与环境, 2008, 29(2): 131-138. doi: 10.3969/j.issn.1671-4814.2008.02.008 [12] 乌兰, 王俊. 乌梁素海2014—2015水环境质量评价分析[J]. 内蒙古师范大学学报, 2017, 46(4): 885-890. [13] 肖博文, 成文连, 姚荣, 等. 内蒙古乌梁素海 N、P 的变化趋势研究[J]. 水资源与水工程学报, 2015, 26(1): 43-51. doi: 10.11705/j.issn.1672-643X.2015.01.008 [14] 李晓霞, 白洋. 浅谈河套灌区农田氮磷流失量及对乌梁素海输入量的估算[J]. 内蒙古环境科学, 2009, 21(3): 44-49. [15] 冯湘云, 李娇, 白妙馨. 乌梁素海流域污染来源分析[J]. 科技创新与应用, 2015(14): 116-117. [16] 王一舒, 吴仁人, 荣楠. 乌梁素海氮磷污染通量输入及海区水质空间异质性研究[C]//中国环境科学学会. 2020中国环境科学学会科学技术年会论文集: 第二卷, 2020: 1540-1546. [17] 乌梁素海[EB/OL]. (2006-04-26)[2022-01-31]. https://baike.baidu.com/item/%E4%B9%8C%E6%A2%%E7%B4%A0%E6%B5%B7/1102650?fr=aladdin. [18] CHIAAPMAN D V. Water Quality Assessments: A Guide to the Use of Biota, Sediments and Water in Environmental Monitoring [M]. Melbourne Madras: Chapman & Hall, 1992. [19] SCHUBLE J R. Report of workshop “The second phase of an assessment of alternatives to biological nutrient removal at sewage treatment plants for alleviating hypoxia in western Long Island Sound”[R]. NYSB: Marine Science Research Center, 1991. [20] 国家环保总局. 地表水环境质量标准: GB 3838-2002[S]. 北京: 中国环境科学出版社, 2002. [21] 赵锁志, 孔凡吉, 赵军, 等. 内蒙古乌梁素海底泥总氮空间分布特征[J]. 现代地质, 2009, 23(1): 82-85. doi: 10.3969/j.issn.1000-8527.2009.01.012 [22] WANG Q P, LIAO Z Y, YAO D X, et al. Phosphorus immobilization in water and sediment using iron-based materials: A review[J]. Science of the Total Environment, 2021, 767: 144246. doi: 10.1016/j.scitotenv.2020.144246 [23] 谢玉斌, 权全, 邹昊, 等. 人工湿地挺水植物净水能力的 Meta 分析[J]. 中国水土保持, 2021, 8(4): 50-53. doi: 10.3969/j.issn.1000-0941.2021.04.017 [24] 段晓男, 王效科, 欧阳志云, 等. 乌梁素海野生芦苇群落生物量及影响因子分析[J]. 植物生态学报, 2004, 28(2): 246-251. doi: 10.3321/j.issn:1005-264X.2004.02.016 [25] 李兴, 李畅游, 勾芒芒, 等. 挺水植物对湖泊水质数值模拟过程的影响[J]. 环境科学, 2010, 21(12): 2890-2895. [26] 商传莲. 物理材料与沉水植物组合对富营养化水体的水质改善效果研究[D]. 上海: 华东理工大学, 2012. [27] 常宝亮, 上官凌飞, 沈志国, 等. 六种沉水植物对三种不同氮磷浓度水体的净化效果[J]. 给水排水, 2021, 47(S1): 230-236. [28] 林春风, 曹国军, 武鹏, 等. 四种沉水植物对富营养化水体的净化效果研究[J]. 安徽农业科学, 2012, 40(10): 6083-6085. doi: 10.3969/j.issn.0517-6611.2012.10.131 [29] 钟继承, 刘国锋, 范成新, 等. 湖泊底泥疏浚环境效应: I. 内源磷释放控制作用[J]. 湖泊科学, 2009, 21(1): 84-93. doi: 10.3321/j.issn:1003-5427.2009.01.011 [30] LIU X, SHENG H, JIANG S Y, et al. Intensification of phosphorus cycling in China since the 1600s[J]. Proceedings of the National Academy of Sciences, 2016, 113: 2609-2614. doi: 10.1073/pnas.1519554113 [31] TONG Y D, ZHANG W, WANG X J, et al. Decline in Chinese lake phosphorus concentration accompanied by shift in sources since 2006[J]. Nature Geoscience, 2017, 10: 507-511. doi: 10.1038/ngeo2967 [32] 徐熊鲲, 谢翼飞, 陈政阳, 等. 曝气强化微生物功能菌修复黑臭水体[J]. 环境工程学报, 2017, 11(8): 4559-4565. doi: 10.12030/j.cjee.201606021 [33] 李畅游, 史小红. 乌梁素海沉积物环境地球化学特征研究[M]. 北京: 科学出版社, 2014. [34] 王立志. 两种沉水植物对间隙水磷浓度的影响[J]. 生态学报, 2015, 35(4): 1051-1058. [35] 张苗, 李明梦, 刘律, 等. 超富营养城市湖泊原位生态修复效果分析: 以武汉南湖为例[J]. 海洋与湖沼, 2021, 52(6): 1399-1407. doi: 10.11693/hyhz20210500116 [36] WU Z, LIU Y, LIANG Z, et al. Internal cycling not external loading, decides the nutrient limitation in eutrophic lake: A dynanmic model with temporal Bayesian hierarchical inference[J]. Water Research, 2017, 116: 231-240. doi: 10.1016/j.watres.2017.03.039 [37] DING S M, WANG Y, WANG D, et al. In situ, high-resolution evidence for iron-coupled mobilization of phosphorus in sediments[J]. Scientific Report, 2016, 6: 2434. [38] ARONSON J, CLEWELL A F, BLIGNAUT J N, et al. Ecological restoration: A new frontier for nature conservation andeconomics[J]. Journal for Nature Conservation, 2006, 14(3/4): 135-139. [39] 卢俊平, 马太玲, 刘廷玺, 等. 大河口水库底泥释磷强度环境影响机理研究[J]. 环境科学与技术, 2017, 40(7): 72-78. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 4883
- HTML全文浏览数: 4883
- PDF下载数: 103
- 施引文献: 0
