

 下载:
下载:
</td><td class="table_top_border" align="center" valign="middle">COD/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle">SS/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle">TN/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle"><inline-formula>$ {\rm{NH}}_4^ + $<img class="inline-formula" src="data:image/svg+xml,<svg xmlns='http://www.w3.org/2000/svg' width='350' height='600'><foreignObject width='2000' height='100%'><div xmlns='http://www.w3.org/1999/xhtml' style='font-size:16px;'><table>
<thead><tr><td class="table_top_border" align="center" valign="middle">水样及标准</td><td class="table_top_border" align="center" valign="middle">水量/(m<sup>3</sup>·d<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle">COD/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle">SS/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle">TN/(mg·L<sup>−1</sup>)</td><td class="table_top_border" align="center" valign="middle"><inline-formula>$ {\rm{NH}}_4^ + $<alternatives><img class="graphic" src="202003047_M2.jpg"><img class="graphic" src="202003047_M2.png"></alternatives></inline-formula>-N/(mg·L<sup>−1</sup>)</td></tr></thead>
<tbody><tr><td class="table_top_border2" align="center" valign="middle">清液</td><td class="table_top_border2" align="center" valign="middle">1 800~2 100</td><td class="table_top_border2" align="center" valign="middle">45 000~65 000</td><td class="table_top_border2" align="center" valign="middle">5 000~8 000</td><td class="table_top_border2" align="center" valign="middle">13 00~16 00</td><td class="table_top_border2" align="center" valign="middle">10~23</td></tr><tr><td align="center" valign="middle">低浓度水</td><td align="center" valign="middle">360~400</td><td align="center" valign="middle">100~200</td><td align="center" valign="middle">20~80</td><td align="center" valign="middle">30~50</td><td align="center" valign="middle">5~10</td></tr><tr><td class="table_bottom_border" align="center" valign="middle">出水标准</td><td class="table_bottom_border" align="center" valign="middle"><b>—</b></td><td class="table_bottom_border" align="center" valign="middle">100</td><td class="table_bottom_border" align="center" valign="middle">60</td><td class="table_bottom_border" align="center" valign="middle">50</td><td class="table_bottom_border" align="center" valign="middle">10</td></tr></tbody>
</table></div></foreignObject></svg>"></inline-formula>-N/(mg·L<sup>−1</sup>)</td></tr></thead>
<tbody><tr><td class="table_top_border2" align="center" valign="middle">清液</td><td class="table_top_border2" align="center" valign="middle">1 800~2 100</td><td class="table_top_border2" align="center" valign="middle">45 000~65 000</td><td class="table_top_border2" align="center" valign="middle">5 000~8 000</td><td class="table_top_border2" align="center" valign="middle">13 00~16 00</td><td class="table_top_border2" align="center" valign="middle">10~23</td></tr><tr><td align="center" valign="middle">低浓度水</td><td align="center" valign="middle">360~400</td><td align="center" valign="middle">100~200</td><td align="center" valign="middle">20~80</td><td align="center" valign="middle">30~50</td><td align="center" valign="middle">5~10</td></tr><tr><td class="table_bottom_border" align="center" valign="middle">出水标准</td><td class="table_bottom_border" align="center" valign="middle"><b>—</b></td><td class="table_bottom_border" align="center" valign="middle">100</td><td class="table_bottom_border" align="center" valign="middle">60</td><td class="table_bottom_border" align="center" valign="middle">50</td><td class="table_bottom_border" align="center" valign="middle">10</td></tr></tbody>
</table></div></foreignObject></svg>)




-
燃料乙醇是我国重点发展的新能源之一,其生产废水的处理和资源化利用很受关注[1]。燃料乙醇的生产以稻谷为原料,经过发酵、精馏后的酒糟通过简单的挤压处理后,所得滤渣经干燥后进入干酒糟工艺(distillers dried grains,DDG)处理流程[2]。产生废水主要成分为糖类、蛋白质、纤维素等,其中粗蛋白含量约占27%[3]。该废水经厌氧生物处理后,TN主要以
NH+4 -N形式存在,且浓度较高,一般为300~600 mg·L−1;即使经过好氧硝化反应也仅发生氮形态的变化,而出水TN未降低,这主要是由于好氧进水碳氮比失调导致反硝化反应效果差。为实现TN达标排放,需要在好氧系统段补充进水碳源。影响反硝化效率的因素较多,活性污泥中微生物组成种类也会影响碳源的选择。在调整脱氮系统工艺前,通过在现场开展指导性实验,研究投加不同碳源时的反硝化脱氮效率,以期获得适宜的碳源补充方案,提高A/O反硝化系统脱氮效率。本研究从脱氮反应速率和经济性综合分析以选择适宜的碳源,并优化运行条件,应用于工程实践中,指导DDG废水处理的A/O脱氮系统高效运行。
全文HTML
-
研究采用快速消解法测定水样中的COD;重量法测定活性污泥浓度;玻璃电极测定水样中的pH;碱性过硫酸钾消解-紫外分光光度法测定TN;半自动凯氏定氮仪测定TNK。
-
某污水站主要有高浓度废水和低浓度废水两股来水。高浓度废水即为DDG废水(俗称清液)。低浓度水中包含实验室废水、净水器排泥水、锅炉除盐水、设备冲洗水、循环水排水和生活区废水等。经过处理后的外排水水质满足《广东省地方标准-水污染物排放限值》(DB 44/26-2001)第二时段一级标准,TN满足《发酵酒精和白酒工业水污染排放标准》(GB 27631-2001)(即50 mg·L−1)排放标准,排至市政污水管网。水质指标如表1所示。
-
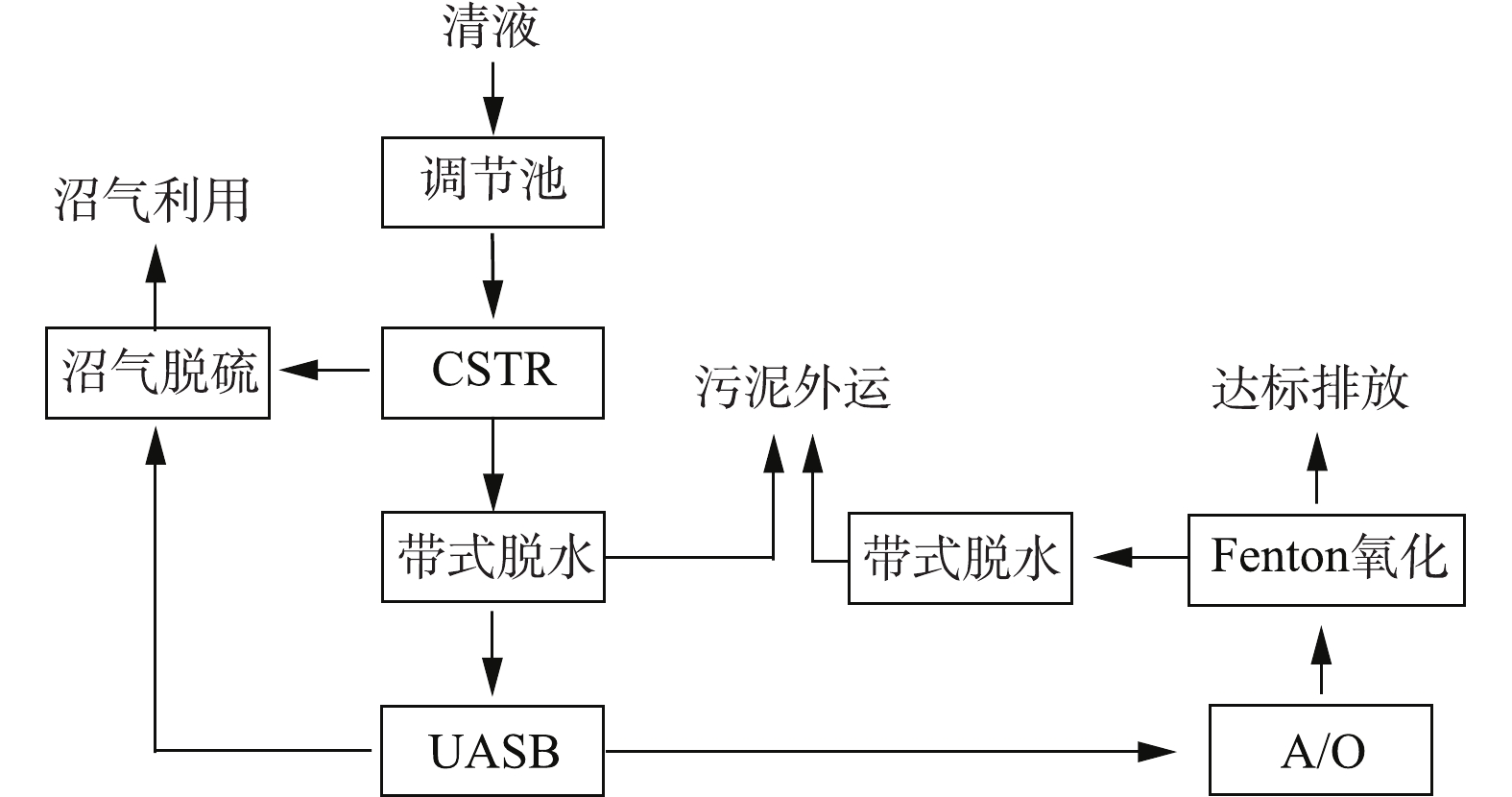
该污水站采取的主要工艺为CSTR+UASB+A/O+芬顿,废水处理量约2 800~3 400 m3·d−1。工艺流程见图1。
-
该污水站运行稳定,除TN外,其他指标均达到排放要求。外排水TN一直为300~360 mg·L−1。为解决这个问题,对该污水站A/O系统的进、出水水质进行了分析,结果如表2所示。
由表2可知,该污水站A/O系统进水的COD/TN约3.1~4.0。TNK反映了水中
NH+4 -N和有机氮的情况,说明系统进水中的总氮主要以NH4+-N和有机氮为主,故可认为A/O工艺适合该废水的处理。杨健等[4]的研究表明,实验室酒精废水经过两级厌氧消化后系统BOD/COD约为0.5。据此计算,该污水站A/O系统进水BOD/TN约为1.6~2.0,明显低于反硝化过程BOD/TN>3的需求[5-6]。因此,可以判断该污水站TN去除率低归因于A/O生化系统进水的BOD/TN失调。影响TN去除率因素较多,针对上述问题结合工程实践,通过对比研究寻找合适的碳源、确定优化运行条件。根据生产实际,把葡萄糖、乙醇和清液作为主要考察对象,主要原因有:1)葡萄糖作为典型碳源添加剂,广泛应用于市政污水处理厂,操作方便且安全稳定;2)该工厂主要产品为燃料乙醇,有丰富的乙醇可作碳源;3)该污水站来水清液含有大量氨基酸、残糖和纤维素等有机物,而发酵酒精清液的BOD/COD一般为0.4~0.5,可生化性较好,因此,清液可作为碳源加以利用,也可实现污水处理的可持续发展[7-8]。
-
1)取400 mL A/O系统生化污泥,用去离子水进行清洗以除去原污水中TN对实验的干扰。按照实际进水流量比例,取A/O出水250 mL与上述污泥混合,混合后的污泥浓度约8 500 mg·L−1。
2)取上述比例混合后的污泥300 mL,分别添加葡萄糖、乙醇和清液,每组实验取3组平行样。在500 mL烧杯内搅拌,搅拌强度以水面无漩涡为标准,防止搅拌剧烈充氧。通过前期摸索性实验得知,当反应起始的COD/TN不低于20时,反硝化过程的最大反应速率不受碳源浓度的影响,因此,实验中确保COD/TN不低于20。为避免因添加碳源带来的pH干扰会影响反应初期的反硝化速率,在添加碳源后统一调整pH为6.9。
3)在不同碳源和不同COD/TN的情况下,通过监测反应系统上清液pH和TN的变化来研究反硝化过程。
1.1. 分析方法
1.2. 水质概况
1.3. 工艺流程
1.4. 污水站水质问题分析及碳源种类选择
1.5. 实验方法
-
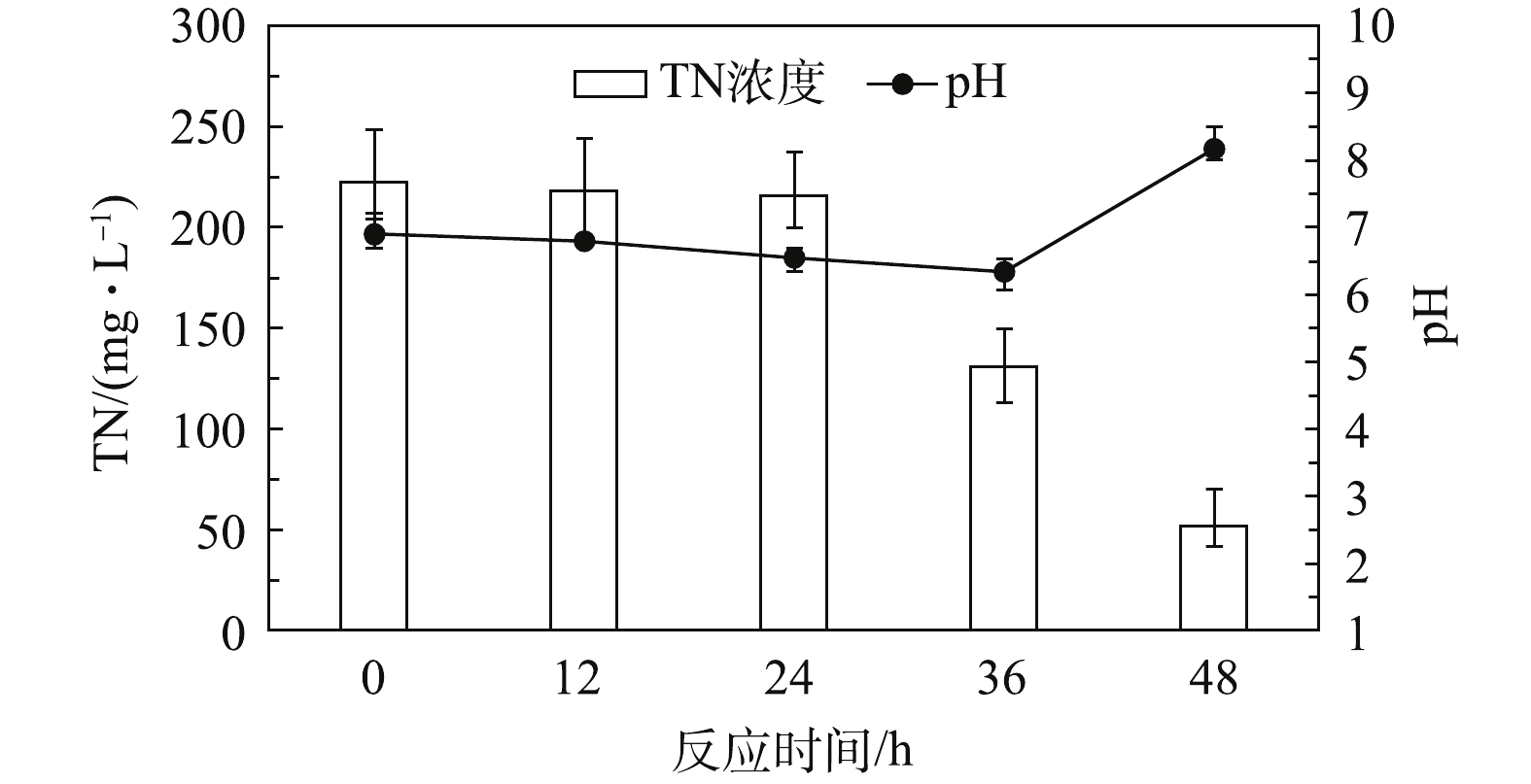
以葡萄糖为碳源的研究结果如图2所示。由图2可知,在反应初始阶段(0~24 h),TN浓度从222.5 mg·L−1下降至215.6 mg·L−1,而反应初始pH由6.90降至6.55。此阶段pH的下降可能是葡萄糖水解产生有机酸导致的[9]。该阶段TN去除量很少,可能是污泥系统中的微生物种群对于基质的变化需要有适应过程,原污水中的葡萄塘较少。加入葡萄糖后,参与反硝化过程的大多数微生物需要时间去适应。反应第2阶段(24~36 h),TN浓度从215.6 mg·L−1下降至130.9 mg·L−1,而pH从6.55下降至6.34。这主要是因为在该系统内主要存在两大类反应:水解酸化和反硝化。在此阶段,葡萄糖水解产酸的速率明显大于反硝化的产碱速率,所以虽然TN已经有了明显下降,但pH仍然下降。反应第3阶段(36~48 h),TN浓度从130.9 mg·L−1下降至52.0 mg·L−1,而pH从6.34上升至8.17,这是典型的反硝化过程,表明在此阶段系统的反硝化产碱速率已明显超过了有机物的水解酸化速率,故系统的pH明显上升。
-
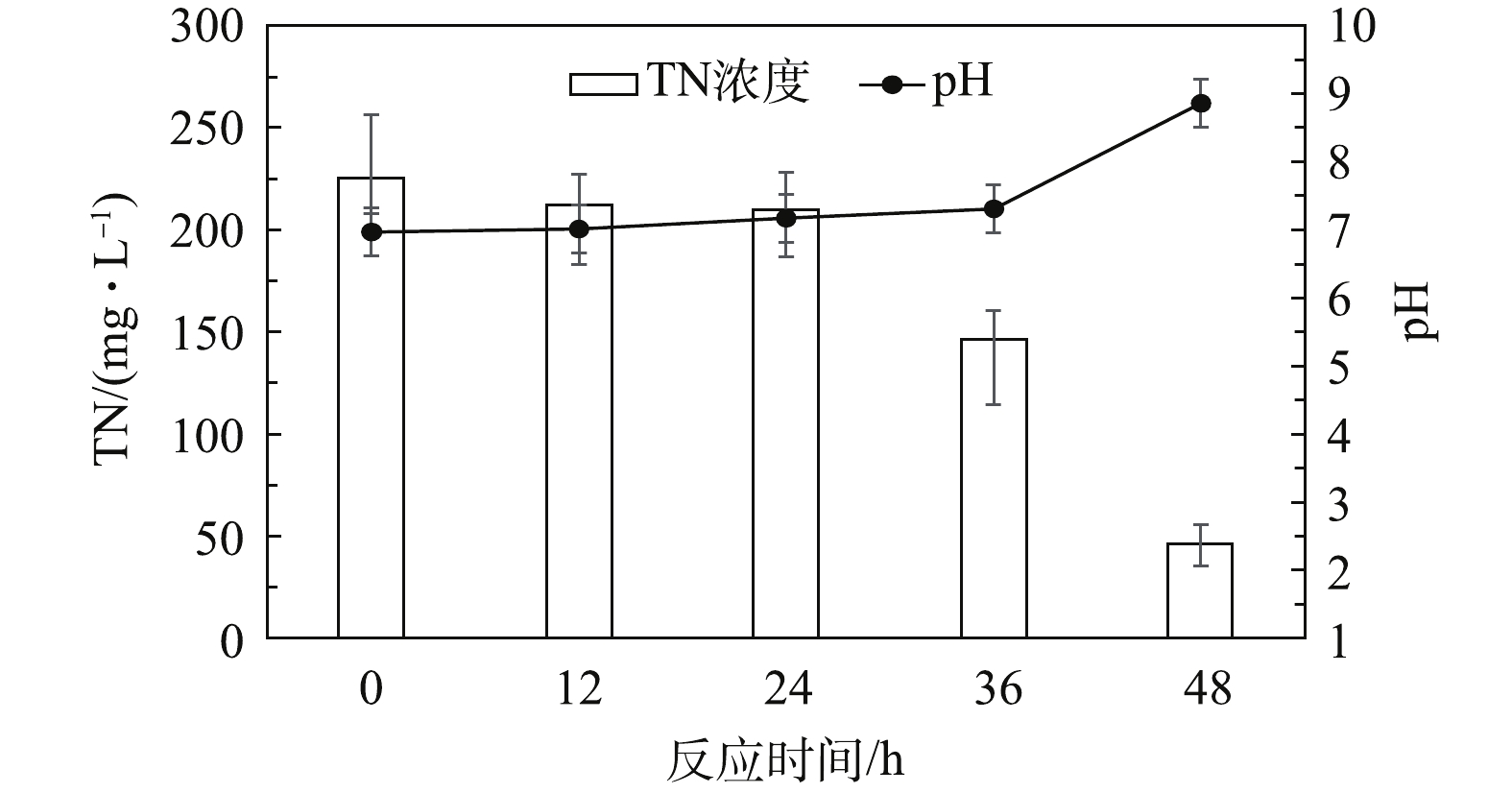
以乙醇为碳源的实验结果如图3所示。反应初始阶段(0~24 h)的TN由225.6 mg·L−1下降至210 mg·L−1,而反应初始pH由6.97上升至7.17。结果表明:反应初始阶段系统中已经有了微弱的反硝化作用;与葡萄糖作为碳源情况类似,污泥中的微生物菌群在更换反应基质后都需要一定的时间去适应[10]。
反应第2阶段(24~36 h)的TN由210.0 mg·L−1下降至146.4 mg·L−1,而pH从7.17上升至7.31。说明微生物经一定时间的适应后,反硝化反应速率明显增加。反应第3阶段(36~48 h)的TN由146.4 mg·L−1下降至46.5 mg·L−1,而pH从7.31上升至8.86,表明此阶段反硝化过程反应速率达到了较高水平。
与葡萄糖作为碳源相比,乙醇为碳源的反应体系中pH值提升速率更快,这可能是由于实际生产废水中本来残存乙醇,微生物菌群对于乙醇的适应能力要强于对葡萄糖的适应能力。
-
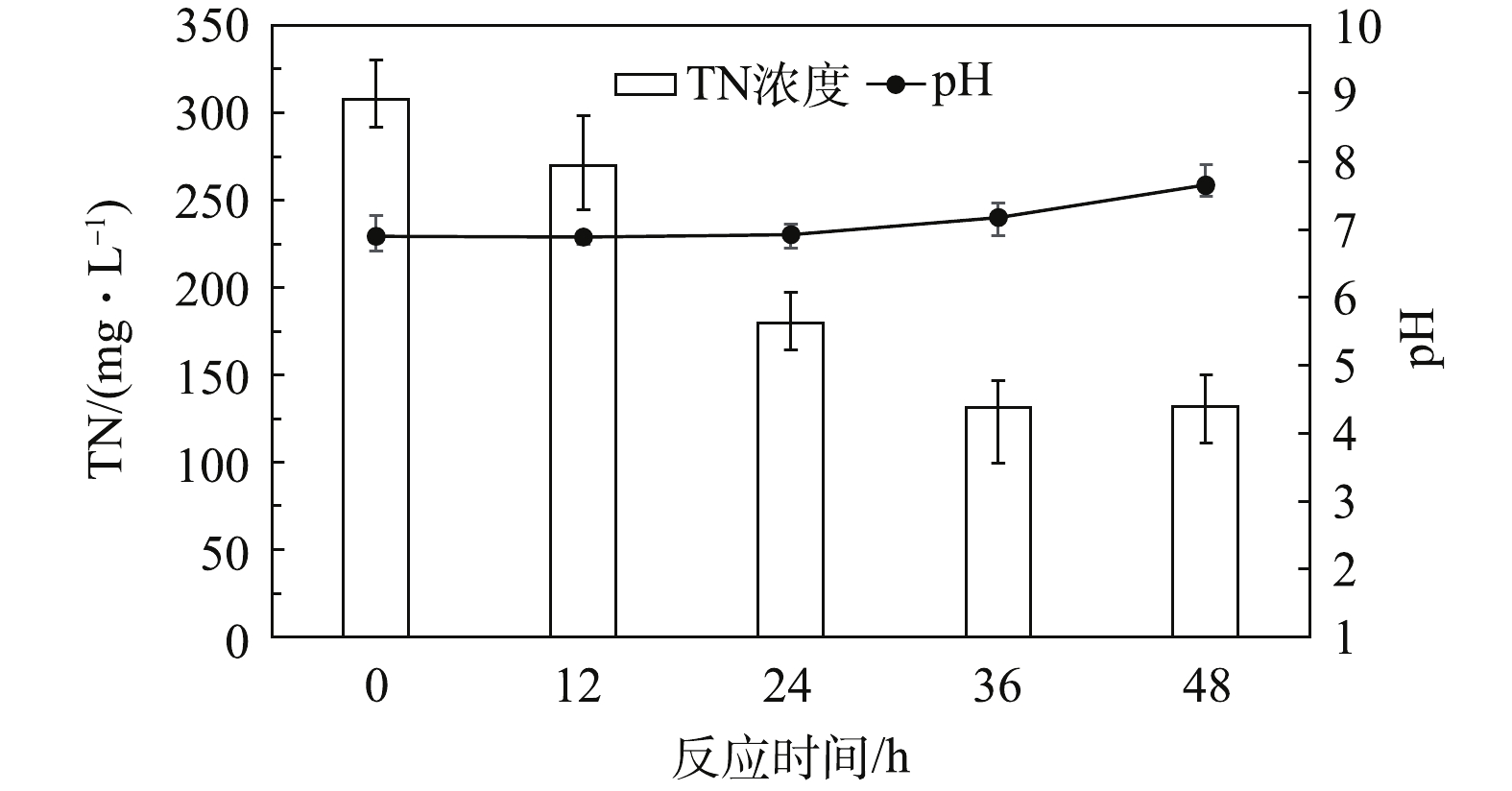
清液实际上是发酵酒精产物,其中所含营养物主要有糖分、蛋白质、纤维素、残糖、短纤维、挥发酸、有机氮、有机磷、钾等,还有大量胶质和菌丝体等[11]。以清液为补充碳源加入后,体系中各参数变化如图4所示。反应初始阶段(0~15 h)的TN由308.1 mg·L−1下降至270.3 mg·L−1,而反应pH基本维持不变。这表明在反应初期,虽然有明显的反硝化作用,但清液中存在的有机酸、氨基酸等物质会消耗反硝化过程产生的碱性物质,故反应体系的pH无明显变化。反应第2阶段(15~24 h)的TN由270.3 mg·L−1下降至179.6 mg·L−1,pH由6.89上升至6.93。在此阶段,TN下降明显,而pH上升速率较慢。这是由于清液中存在的酸性物质继续缓冲了反硝化产生的碱,导致系统的pH变化缓慢。反应第3阶段(24~36 h)的TN由179.6 mg·L−1下降至131.9 mg·L−1,pH由6.93上升至7.18。该阶段TN的反应速率开始下降,但pH上升幅度比第2阶段要快。这可能是由于清液中的酸性物质已消耗殆尽,缓冲作用已明显削弱,故系统pH变化明显。反应第4阶段(36~48 h)的TN在132 mg·L−1左右不再继续下降,但pH从7.15上升至7.65。这可能是由于经过一段时间的反应,清液中的含氮有机物开始了氨化反应,氨化反应产生的游离氨使系统的pH升高[12-13]。
相比葡萄糖和乙醇,清液作为碳源时,系统的最终pH最低。这可能是由于清液中含有大量酸性物质所致。这部分酸性物质主要由乳酸、琥珀酸、乙酸和丁酸等发酵副产物组成。而葡萄糖比乙醇更容易发生水解酸化,使得葡萄糖作碳源时反硝化反应的最终pH更低。
-
根据上述实验结果,对比3种碳源在反硝化反应中的TN去除速率,计算公式如式(1)所示,结果如表3所示。
式中:ΔN为TN反应速率即在单位时间内TN下降浓度,mg·(L·h)−1;
Ns 为在一定反应时间后的TN浓度,mg·L−1;N0为在反应初始期的TN浓度,mg·L−1;ΔT为反应时间,h。由表3可知,在较长反应时间内,以上3种碳源条件下的平均TN去除速率接近,分别为3.55、3.73和3.67 mg·(L·h)−1。这表明在碳源充足的情况下,当反应时间足够长时,碳源种类对反硝化过程的影响较小。但对于某一时段内的反应速率,3种碳源还是有明显区别的。分时段内3种碳源下的最大TN去除速率分别为葡萄糖7.06 mg·(L·h)−1、乙醇8.33 mg·(L·h)−1、清液7.56 mg·(L·h)−1。乙醇去除速率最快主要是由于反硝化细菌优先利用小分子醇类[14],而葡萄糖作为相对复杂的有机物,需要水解酸化后才被吸收。而清液由于能被反硝化细菌快速利用的小分子酸类物质占比较小,该条件下反应速率低于乙醇做碳源的条件。吴代顺等[15]研究表明,当利用乙醇作为碳源进行反硝化反应时,最大反硝化速率超过葡萄糖等有机物。
-
假定A/O系统的进水参数为:流量200 m3·h−1;COD 1 800 mg·L−1、TN 450 mg·L−1;投加碳源后的COD/TN=20。以此计算3种碳源投加条件下,生物脱氮过程增加的运行成本,结果见表4,其中葡萄糖和乙醇的单价采用市场采购价格,而清液单价以清液未进行厌氧反应而损失的沼气发电收益来计。
上述计算过程未考虑添加碳源后系统运行成本的增加。这是由于即使添加的碳源全部以曝气方式去除,在该污水站的曝气系统效率及电价基础上,每吨污水的处理成本增加量也不到0.5元。与表4中的成本增加相比,影响较小,所以未作过多考虑。表4计算结果显示,清液成本为仅为乙醇成本的9%,葡萄糖成本的37%。从经济角度出发,应该充分利用废水中的有机物来进行生物脱氮;而充分利用原废水中的碳源也可实现污水资源利用,符合污水处理可持续发展的需要。
2.1. 葡萄糖为碳源对处理效果的影响
2.2. 乙醇为碳源对处理效果的影响
2.3. 清液为碳源对处理效果的影响
2.4. 不同碳源反应条件下TN去除速率对比
2.5. 经济性比较
-
投加不同碳源时,清液的最大反硝化速率是乙醇的90%;从经济成本方面考虑,投加清液的成本仅是乙醇的10%。通过实验研究分析,综合脱氮反应速率和运行成本经济性两方面考虑,以清液作为反硝化碳源具有较明显的优势。
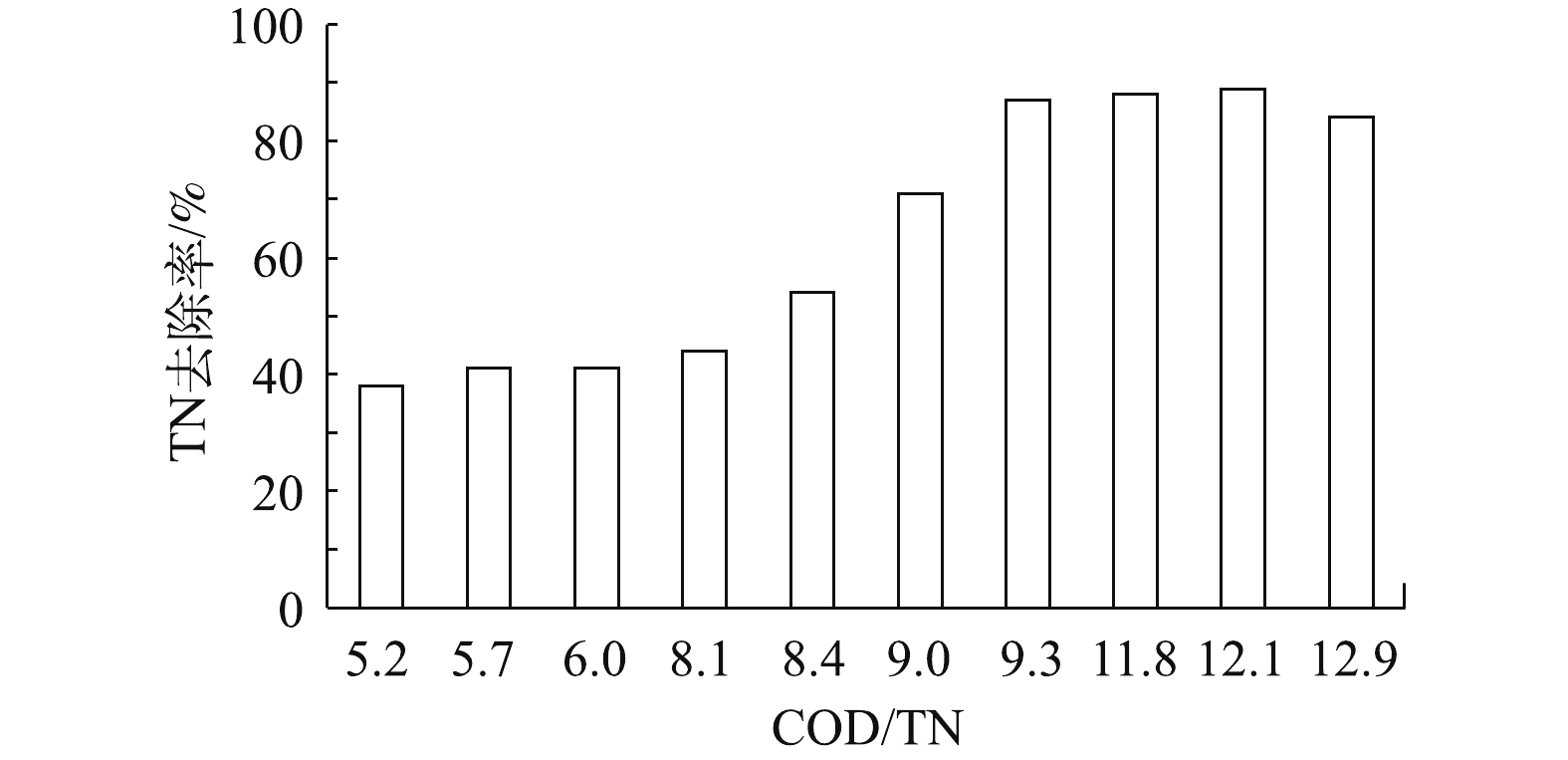
该污水站A/O系统进水流量200 m3·h−1,反硝化停留时间为50 h,内回流比为450%,外回流比100%。未添加清液时,A/O进水COD为1 792~2 023 mg·L−1。外加碳源清液的COD为50 000~62 000 mg·L−1,TN为1 100~1 500 mg·L−1,对不同清液投加量下的A/O系统TN去除率进行了比较,结果如图5所示。未添加清液前,A/O系统进水的COD/TN在5~6,TN去除率约41%;在添加清液后,COD/TN在6.0~9.3,TN去除率上升幅度较快。这主要是由于清液中的有机物被反硝化细菌快速利用,TN去除得以加速。当COD/TN在9.3~12.1时,进水COD在3 058~5 097 mg·L−1,TN在378~422 mg·L−1,出水COD在425~504 mg·L−1,TN在46~48 mg·L−1,TN去除率可以稳定在85%以上,最高达到89%,且水中TNK未检出,说明系统中氨化反应和硝化反应进行彻底,则TN主要以
NO−3 -N存在。而当COD/TN继续升高至12.9时,TN去除率反而下降。这主要是因为清液自身带有一定浓度的有机氮,这些氮的存在影响了原水中TN去除量的计算,从而影响了系统的TN去除率。根据上述实验结果对污水站的运行参数进行了相应调整,当混合液回流比和污泥回流比分别为450%和100%时,采用COD/TN=12.1,即每kg TN投加清液约307 L,该污水站的外排TN可稳定在50 mg·L−1以下。综上所述,DDG项目废水脱氮工艺中,A/O系统进水TN约300~600 mg·L−1时,可通过投加清液作为碳源,以确保A/O系统废水COD/TN在12.1以上,使A/O系统保持较高的TN去除率的同时,对运行成本的影响有限。
-
1)对于反硝化脱氮碳源不足废水处理,通过投加葡萄糖、乙醇、清液3种不同碳源进行反硝化实验,结果表明:采用葡萄糖、乙醇和清液分别做反硝化碳源时,脱氮效率分别为76.6%、79.4%和57.2%;同时,以葡萄糖作碳源时的TN去除速率最小,清液次之,乙醇作碳源的TN去除速率最大为8.33 mg·(L·h)−1,反硝化细菌优先利用小分子醇和酸类有机物作为反硝化碳源。
2)对3种碳源条件的经济性比较发现,清液为乙醇成本的9%、为葡萄糖的37%。利用原废水中的有机物经济性最佳,也符合污水处理可持续发展的需要。
3)在DDG项目废水处理脱氮工艺中,A/O系统进水TN约300~600 mg·L−1时,可通过投加清液作为碳源确保A/O系统废水的COD/TN在12.1以上,从而使A/O具有较高TN去除率的同时,对运行成本的影响有限。
4)实际工程应用研究为DDG废水的处理提供了经济合理的碳源补充方案,能为可生化性较好的发酵行业废水处理提供参考。为获得更好的脱氮效率,下一步还需探讨好氧系统内回流比及反硝化停留时间对反硝化效率的影响。
