


 百度学术
百度学术


全文HTML
多氯联苯(polychlorinated biphenyls, PCBs)因良好的化学稳定性、抗热性和绝缘性,曾广泛用于电容和变压器的冷却剂和润滑油,以及电子设备中的导热油和液压油[1]。PCBs结构稳定,具有高毒性和持久性,一旦进入环境就会长期存在,而水体沉积物被认为是环境中PCBs的主要归宿之一[2]。由于前期管理的无序化,我国电子垃圾拆解地区各种环境介质中PCBs被频繁检出。路风辉等[3]在研究清远龙塘镇水体沉积物中PCBs的分布特征时,发现其中有29种PCBs浓度高出正常水平(8.67 ng·g-1)的2~3个数量级。
目前,国内外对于受PCBs污染的土壤及沉积物修复技术主要包括生物修复和非生物(物理和化学)修复2种[4-5]。相对于土壤淋洗、工业性焚烧、改性纳米零价铁还原、Fenton氧化等物理及化学处理方法,原位生物修复PCBs污染场地具有环境可持续性、基本无二次污染等优点[6]。许多研究证实生物电化学技术可以应用于水体沉积物和地下水的原位修复,通过外部电压刺激加速特定污染物的去除,如石油烃[7]、多环芳烃[8]、氯代有机物[9]等。近期有研究[10-13]表明,利用电化学技术与微生物相耦合的方式,借助微弱外电场刺激土著脱氯微生物的生长和繁殖,可促进PCBs的厌氧还原脱氯。SUN等[10]将碳毡埋覆在底泥中通过施加恒定电压来控制底泥环境的氧化还原电位,从而促进PCBs的氧化还原反应,证实了电化学促进微生物在底泥环境中还原PCBs的可行性。CHUN等[11]通过施加恒定电场(1.5~3.0 V)促进水电解,阴极表面析H2以作为电子供体还原高氯代PCBs;阳极表面产O2形成好氧环境,氧化低氯代PCBs直至其矿化。与电解产氢介导PCBs还原脱氯不同,YU等[12]和LIU等[13]以PCB 61(2, 3, 4, 5-CB)为研究对象,采用微电压(如+0.2 V, SCE)刺激还原脱氯,在过电位不至于析H2情况下,证实PCB 61还原速率和程度得到了极大提高。尽管利用微电场耦合原位修复PCBs在概念上具有先进性和新颖性,但目前相关研究仍处于起步阶段,很多原理性问题尚待解决。其中,施加电场的大小是决定脱氯微生物结构、类型和丰度的关键因素,进而影响还原脱氯效果,这是因为不同微生物能够在热力学上适应不同电场环境,甚至以外加电场作为能量供给改变其代谢方式[14-15]。目前尚未见文献报道不同电压对PCBs厌氧还原脱氯的影响规律,不同电压刺激下菌群结构的变化有待进一步研究。
本研究利用生物电化学技术研究不同恒定电压(-0.1、-0.3、-0.5、-0.7 V, SCE)对沉积物中微生物厌氧还原PCBs的影响;较为系统地考察了不同电压及开路情况下PCB 61降解速率、效率及对产物PCB 23和PCB 29浓度的影响;通过对不同实验条件驯化后的微生物及起始接种底泥的微生物菌群结构进行对比分析,明确了微生物群落结构变化和优势微生物功能差异,分析了功能微生物与PCB 61还原脱氯的关系。
1 材料与方法
1.1 反应器的构建
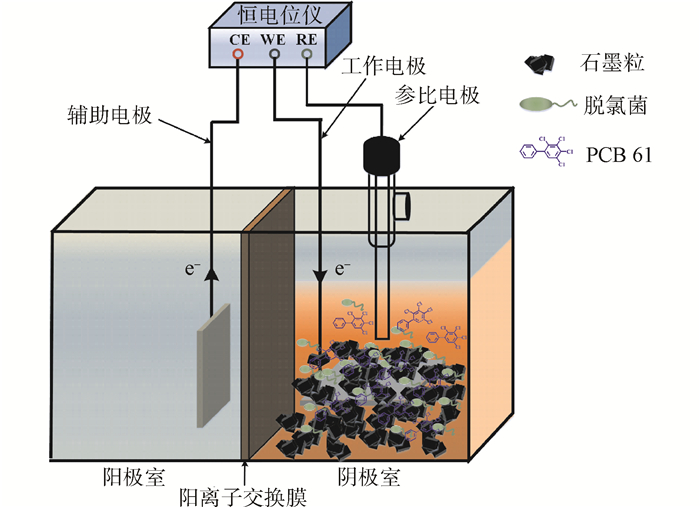
本研究所用的生物电化学反应器由阳极室和阴极室2个室构成,2室中间用阳离子交换膜隔开,双极室的容积均为100 mL,如图 1所示。其中阴、阳两极的电极材料均为10%双氧水预处理的石墨毡,尺寸均为2.5 cm×2.5 cm×0.5 cm, 反应器按照文献[13]的方法进行安装。随后将其对电极、工作电极和参比电极与CHI1000C型电化学工作站相接,其中参比电极和工作电极安置在含有底泥沉积物的阴极室,对电极安置在阳极室。
图1 生物电化学反应器
Fig. 1 Bioelectrochemical reactor
Fig. 1 Bioelectrochemical reactor
1.2 反应器的接种与运行
在进行生物电化学还原多氯联苯的实验前,双室电解池阳极室加入磷酸盐缓冲液(pH=7.0);阴极室加入体积为15 mL石墨颗粒(直径为6 mm)及与PCB 61(120 μmol·kg-1,即35.04 mg·kg-1)均匀混合的河涌底泥沉积物(取自广东省清远市龙塘镇电子垃圾拆解场地区河涌),随后加入pH=7.0的磷酸盐缓冲液培养基[16](2.70 g·L-1 NaHCO3, 0.25 g·L-1 K2HPO4,0.10 g·L-1 KCl,0.25 g·L-1 NH4Cl,0.005 0 g·L-1 Ca(OH)2,10 mL·L-1矿物质和10 mL·L-1维生素)和乙酸钠(最终控制浓度为20 mmol·L-1)。插入饱和甘汞电极,待底泥沉降稳定后,连接电化学工作站,并维持工作电极电位值恒定在-0.1、-0.3、-0.5、-0.7 V(除特别标注外,文中出现的电位值均相对于SCE电极)。在电化学反应进行前,向反应器持续通氮气30 min,驱除底泥及水中的氧气。反应器的取样口用胶塞密封,缝隙用胶水密封,保证反应器在厌氧条件下运行。同时,为了对比差异,设定了开路条件(反应器不连接外电路)对照,反应器均置于30 ℃的恒温恒湿培养箱中,每次实验设定2个平行实验组且在相同条件下运行。
1.3 PCBs的提取及分析
称取0.4 g干泥放于萃取瓶中,加入含有4.0 mg·L-1 PCB 204的正己烷(色谱纯)作为回收率指示物,待正己烷挥发完毕后,加入15.0 mL正己烷-丙酮(体积比1:1)提取溶剂,超声混溶15 min,离心,重复3次。提取液经旋转蒸发仪浓缩至3.0~4.0 mL,再进行氮气吹扫浓缩至1.0 mL左右。采用CNWBOND Florisil SPE小柱对提取液进行净化[13]。净化液用旋转蒸发仪浓缩至2.0 mL,再用氮气吹扫浓缩至1.0 mL。
经过预处理之后的样品,采用GC-MS进行定性定量分析。色谱条件[16]:BD-5MS(尺寸为30 m×0.25 mm×0.25 μm)色谱柱;升温程序:起始温度100 ℃,维持2 min后,以20 ℃·min-1的速率上升到180 ℃,然后以2 ℃·min-1的速率上升到200 ℃,最后以15 ℃·min-1的速率上升到300 ℃再维持5 min;使用氦气(He)作为载气,流速为1.0 mL·min-1,在280 ℃的注射器温度下,将1.0 μL提取物样品以不分流模式注入GC端口。
1.4 微生物群落分析
底泥中微生物采用MOBIO PowerSoil DNA Isolation Kit进行基因组DNA提取,然后对16S rDNA基因的V4~V5区域进行扩增处理,片段大小为430 bp(带有barcode),所用的正向引物为515F:5′-GTGCCAGCMGCCGCGGTAA-3′,反向引物为907R:5′-CCGTCAATTCMTTTRAGTTT-3′。PCR扩增体系的60 μL混合物包括10x ExTaq Buffer 6.0 μL,dNTP 6.0 μL,BSA 0.6 μL,Ex Taq 0.3 μL,Primer F 1.2 μL,Primer R 1.2 μL,DNA 1.0 μL,ddH2O 43.7 μL。扩增反应的升温过程为94 ℃下预变性5 min,再进行31个周期的操作:94 ℃下变性30 s,52 ℃下低温退火30 s,72 ℃延伸扩增45 s。完成以上所有循环后,再在72 ℃下延伸扩增10 min[17]。
2 结果与讨论
2.1 PCB 61还原脱氯效率及途径分析
2.1.1 电极电位对PCB 61还原脱氯的影响
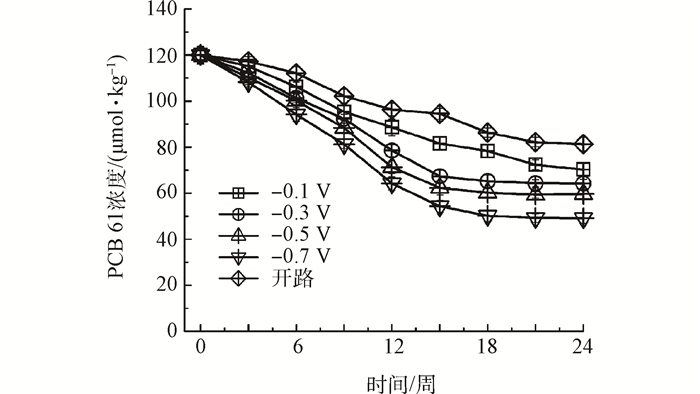
本文主要研究电极电位大小对PCB 61的还原脱氯的影响,因此,实验过程设定了一系列电极电位条件:-0.1、-0.3、-0.5、-0.7 V和开路对照。不同电极电位条件下,反应器中底泥PCB 61浓度的变化如图 2所示。结果显示:各实验组在运行到第6周时,底泥中的PCB 61出现不同程度的降解响应;而在第6~15周时,各实验组底泥中PCB 61的降解速率达到峰值;第15周后,PCB 61的降解速率趋于平缓,主要受动力学的限制[18],PCB 61的进一步降解变得更难了。相对于开路条件,不同外加电压下PCB 61的脱氯转化效果有明显的差异。开路条件下,体系相当于传统的厌氧生物还原脱氯过程,反应器运行第24周后,PCB 61的降解率达到了32.22%,因此,也表现出一定的降解效果。在反应时间达到24周时,施加-0.1、-0.3、-0.5和-0.7 V电压的反应器底泥中PCB 61的降解率分别达到了41.40%、46.49%、50.30%、59.05%,相较于开路条件下PCB 61的降解率(32.22%)有明显提高。在负电位刺激微生物还原PCB 61下,施加的电位越小,所表现出的PCB 61脱氯转化速率越高,其中最小的电位值-0.7 V高于实际情况中析氢反应所需电位值,因此,可以排除H2作为电子供体还原PCB 61的可能。
图2 不同电位条件下底泥中PCB 61浓度随时间的变化
Fig. 2 Change of PCB 61 concentration with time in sediment at various potential
Fig. 2 Change of PCB 61 concentration with time in sediment at various potential
图2 不同电位条件下底泥中PCB 61浓度随时间的变化
Fig. 2 Change of PCB 61 concentration with time in sediment at various potential
Fig. 2 Change of PCB 61 concentration with time in sediment at various potential
为进一步分析其降解速率的差异,分别对各个实验条件下PCB 61的降解进行准一级动力学模型的拟合,结果如表 1所示。可以看出,在生物电化学体系中,反应速率k表现为随着电极电位的减小而增加。在施加-0.7 V电位条件下, 还原脱氯速率最快,其准一级反应动力学常数达到0.006 0 d-1,是开路条件下的2.30倍。
表1 PCB 61还原脱氯准一级动力学拟合参数
Table 1 Pseudo-first order kinetics fitting parameter of PCB 61 dechlorination
Table 1 Pseudo-first order kinetics fitting parameter of PCB 61 dechlorination
表1 PCB 61还原脱氯准一级动力学拟合参数
Table 1 Pseudo-first order kinetics fitting parameter of PCB 61 dechlorination
Table 1 Pseudo-first order kinetics fitting parameter of PCB 61 dechlorination
| 实验条件 | k/d-1 | R2 |
| -0.1 V | 0.003 4 | 0.981 1 |
| -0.3 V | 0.004 3 | 0.941 2 |
| -0.5 V | 0.004 8 | 0.922 3 |
| -0.7 V | 0.006 0 | 0.943 2 |
| 开路 | 0.002 6 | 0.986 9 |
2.1.2 还原脱氯产物分析
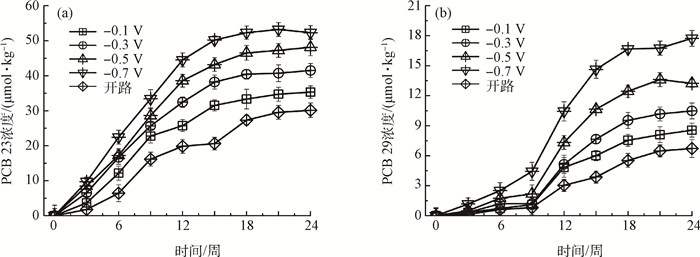
通过GC-MS分析PCB 61脱氯转化过程的产物可知,各实验条件下脱氯产物均为PCB 23(2, 3, 5-CB)和PCB 29(2, 4, 5-CB),而未检测到进一步的脱氯产物二氯联苯,由此可以推断,体系中PCB 61的脱氯途径主要为双侧对位脱氯途径和双侧间位脱氯途径[19]。图 3显示了底泥中PCB 23及PCB 29的浓度随时间的变化情况,PCB 23和PCB 29的产生量与PCB 61的脱氯转化程度呈现正相关的对应关系。BEDARD等[20]对河流沉积物中微生物对Aroclor 1260还原脱氯过程进行了研究,发现体系中的脱氯产物主要通过双侧对位及双侧间位途径得到。除此之外,GOMES等[21]利用电化学方法强化底泥中的微生物还原PCBs的研究结果同样表明:PCBs的还原脱氯途径为双侧对位及双侧间位脱氯,而双侧邻位的氯原子很难被还原脱氯。可以看出本文的实验结果与上述研究得出的PCBs脱氯途径一致,即施加恒定电位没有改变脱氯的途径,只是提高了电子传递的速率,从而强化PCB 61还原反应过程。
图3 底泥中脱氯产物PCB 23(a)及PCB 29(b)的浓度变化
Fig. 3 Change of concentration of dechlorinating products (PCB 23 (a) and PCB 29 (b)) in sediments
Fig. 3 Change of concentration of dechlorinating products (PCB 23 (a) and PCB 29 (b)) in sediments
图3 底泥中脱氯产物PCB 23(a)及PCB 29(b)的浓度变化
Fig. 3 Change of concentration of dechlorinating products (PCB 23 (a) and PCB 29 (b)) in sediments
Fig. 3 Change of concentration of dechlorinating products (PCB 23 (a) and PCB 29 (b)) in sediments
2.2 微生物群落组成分析
为探明生物电化学体系中PCBs脱氯微生物的种群结构的变化,我们通过高通量测序技术对样品中微生物16S rRNA基因上的V4~V5区域进行分子生物学分析,包括不同条件下的12个样品,每个样品中平均长度达到430 bp的高质量序列总共2万多条,根据99%的相似性聚类分成94 362个操作分类单元(OTUs)。各实验条件的样品表示如下:初始底泥(S0),-0.1 V电位(S1),-0.3 V电位(S2),-0.5 V电位(S3),-0.7 V电位(S4),开路条件(S5)。其余的S0′~S5′则分别表示相应条件下的重复样品。
2.2.1 β多样性分析
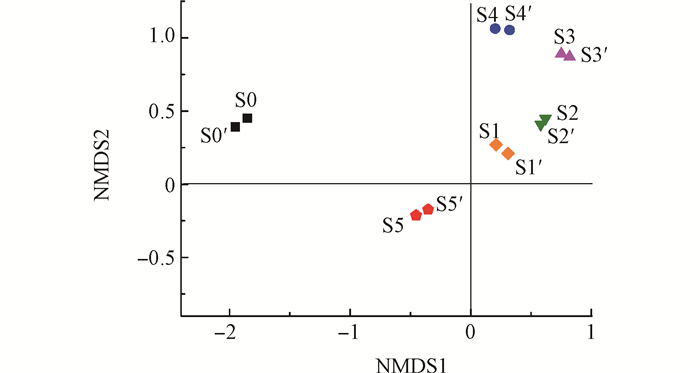
β多样性是指不同样品之间的生物多样性比较,与其他样品相关。其中的非度量多维尺度分析(non-metric multi-dimensional scaling,NMDS)是一种将多维空间的研究对象(样本或变量)简化到低维空间进行定位、分析和归类,同时又保留对象间原始关系的数据分析方法。其特点是根据样品中包含的物种信息,以点的形式反映在多维空间上;而对不同样品间的差异程度,则是通过点与点间的距离体现的,最终获得样品的空间定位点。从图 4可以看出,不同实验条件下的底泥样品与初始底泥(S0、S0′)均存在较大距离,即施加恒定电压使得微生物群落结构发生了明显变化。除此之外,各重复样品之间的距离较近,说明其微生物群落结构差异不大。从图 4还可看出,不同电极电位对于微生物群落组成的影响亦不同。例如,在接种培养了24周后开路条件(S1)与接种底泥(S0)的距离较大,而施加不同负电位的条件则均与之存在较大距离。而且,-0.5 V电位条件(S3)与-0.7 V电位条件(S4)的生物组成比较接近。
图4 基于OTUs的非度量多维尺度分析
Fig. 4 Analysis of nonmetric multi-dimensional scaling based on classified OTUs
Fig. 4 Analysis of nonmetric multi-dimensional scaling based on classified OTUs
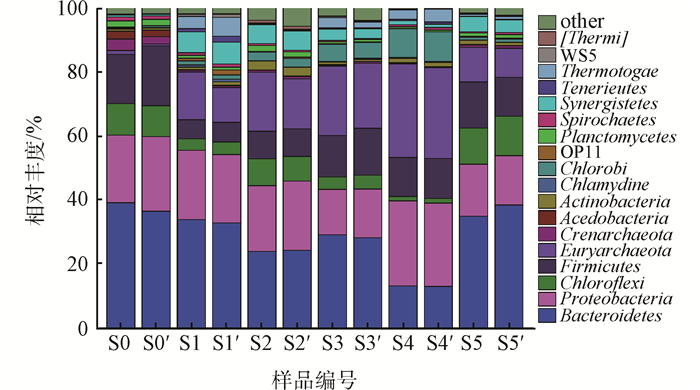
2.2.2 门水平微生物群落组成分析
门水平的微生物组成如图 5所示,反映出体系中微生物的总体差别。总共鉴定出18个门水平的微生物,剩余未分类的微生物仅占0.19%~5.66%。其中,拟杆菌门(Bacteroidetes)在各个条件下均表现出显著优势,相对丰度为13.00%~39.22%,且初始底泥(S0)、开路条件(S5)高于负电位条件。其他主要优势菌门包括:变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)和绿弯菌门(Chloroflexi),这些菌门都被报道具有还原氯代有机物(PCBs等)的功能[22-24]。值得注意的是,广古菌门(Euryarchaeota)发生了显著增长。初始底泥中,Euryarchaeota的相对丰度低于1%,而其他各条件下均发生了不同程度的增长,且表现为负电位条件明显高于开路条件。研究[25]表明,Euryarchaeota主要由产甲烷古菌组成,而部分甲烷古菌可以降解底泥中的PCBs。另外一种相对于初始底泥(S0)显著增加的菌门为养菌门(Synergistetes),Synergistetes被报道在PCBs脱氯的底泥或无底泥体系中为主要菌门[26]。
图5 门水平的微生物组成分析
Fig. 5 Analysis of microbial composition at phylum level
Fig. 5 Analysis of microbial composition at phylum level
2.2.3 优势菌属分析
为更好地分析不同电极电位对于生物电化学体系中微生物群落结构的影响,阐明生物电化学系统中优势菌属与脱氯还原性能的关系,对微生物群落结构的属水平进行了分类分析,结果见表 2。表 2中所列的微生物均至少在一个平行实验组中相对丰度超过1%。在接种底泥中微生物(S0)相对丰度超过1%的有8种。其中微生物主要有2类:一类为脱氯或脱卤菌,如Acinetobacter[27]和Flavobacterium[28];另一类为发酵、产氢菌,包括Fusibacter[29]、Anaerovorax[30]和Citrobacter[31],这类微生物的存在说明底泥中的有机质含量较高。
表2 优势菌属相对丰度分析(>1%)
Table 2 Analysis of dominant genera abundance (>1%)
Table 2 Analysis of dominant genera abundance (>1%)
表2 优势菌属相对丰度分析(>1%)
Table 2 Analysis of dominant genera abundance (>1%)
Table 2 Analysis of dominant genera abundance (>1%)
| 序号 | 门 | 属 | 种 | S0 | S1 | S2 | S3 | S4 | S5 |
| 1 | Proteobacteria | Enterobacteriaceae | Citrobacter | 1.04 | 0.82 | 0.80 | 0.39 | 0.59 | 0.76 |
| 2 | Proteobacteria | Moraxellaceae | Acinetobacter | 1.12 | 0.90 | 0.72 | 0.40 | 0.65 | 0.69 |
| 3 | Chloroflexi | Anaerolinaceae | WCHB1-05 | 2.42 | 0.55 | 0.79 | 0.38 | 0.01 | 0.64 |
| 4 | Firmicutes | Tissierellaceae | Tissierella | 1.01 | 0.44 | 0.24 | 0.26 | 0.10 | 0.25 |
| 5 | Proteobacteria | Gallionellaceae | Gallionella | 1.13 | 0.02 | 0.12 | 0.00 | 0.00 | 0.09 |
| 6 | Firmicutes | Mogibacteriaceae | Anaerovorax | 2.36 | 0.54 | 0.02 | 0.32 | 0.04 | 0.02 |
| 7 | Firmicutes | Acidaminobacteraceae | Fusibacter | 9.42 | 0.03 | 0.04 | 0.21 | 0.14 | 0.04 |
| 8 | Bacteroidetes | Flavobacteriaceae | Flavobacterium | 2.18 | 0.71 | 0.02 | 0.54 | 0.05 | 0.02 |
| 9 | Proteobacteria | Syntrophaceae | Syntrophus | 0.18 | 2.24 | 2.20 | 0.36 | 0.20 | 5.54 |
| 10 | Thermotogae | Thermotogaceae | Kosmotoga | 0.01 | 4.77 | 0.17 | 2.46 | 3.40 | 6.71 |
| 11 | Chloroflexi | Anaerolinaceae | SHD-231 | 0.46 | 1.06 | 1.48 | 1.42 | 0.14 | 9.19 |
| 12 | Euryarchaeota | Methanosarcinaceae | Methanosarcina | 0.09 | 3.24 | 9.80 | 15.91 | 16.38 | 1.53 |
| 13 | Proteobacteria | Comamonadaceae | Hydrogenophaga | 0.38 | 0.85 | 1.82 | 1.14 | 2.76 | 1.59 |
| 14 | Bacteroidetes | Rikenellaceae | Blvii28 | 0.10 | 1.65 | 2.08 | 1.86 | 3.10 | 1.60 |
| 15 | Euryarchaeota | Methanosaetaceae | Methanosaeta | 0.16 | 7.67 | 10.50 | 16.52 | 22.45 | 3.10 |
| 16 | Euryarchaeota | Methanobacteriaceae | Methanobrevibacter | 0.01 | 0.95 | 2.13 | 4.04 | 5.17 | 0.47 |
| 17 | Chlorobi | Ignavibacteriaceae | Ignavibacterium | 0.02 | 1.03 | 2.61 | 3.49 | 4.52 | 0.78 |
| 18 | Proteobacteria | Geobacteraceae | Geobacter | 0.05 | 1.96 | 2.41 | 5.47 | 7.54 | 0.19 |
| 19 | Proteobacteria | Oceanospirillaceae | Marinobacterium | 0.12 | 2.26 | 4.71 | 8.26 | 16.43 | 0.70 |
| 20 | Firmicutes | Erysipelotrichaceae | PSB-M-3 | 0.52 | 2.04 | 0.04 | 0.34 | 1.04 | 3.25 |
| 21 | Proteobacteria | Syntrophobacteraceae | Syntrophobacter | 0.11 | 0.35 | 0.80 | 0.67 | 0.18 | 5.01 |
| 22 | Thermi | Trueperaceae | B-42 | 0.01 | 0.17 | 0.96 | 0.14 | 0.05 | 2.18 |
反应器运行到24周时,闭路组(S1、S2、S3、S4)和接种底泥(S0)在微生物群落组成上差异较大,这一规律在之前的非度量多维尺度分析图(图 4)上也有体现。而丰度差异明显的第1种微生物是电化学活性菌,在闭路组中具有电化学活性的微生物Geobacter、Ignavibacterium相对丰度较高,其中S4中Geobacter、Ignavibacterium相对丰度最高,达到了4.52%和7.54%,而Geobacter本身还具备还原PCBs的能力[23]。第2种是产甲烷微生物,这类菌与PCBs的还原脱氯有密切关系。YE等[32]的研究报道直接证明了产甲烷菌对于PCBs对位脱氯的功能,拓宽了对于PCBs脱氯微生物的认识。大量研究报道表明,产甲烷条件或产甲烷纯菌可以实现氯代有机物的还原脱氯[33-34],并且有研究推测这是由于产甲烷菌可以在厌氧环境中还原过渡金属辅酶从而催化还原脱氯反应的发生[35]。Methanosaeta的主要特点是乙酸发酵型产甲烷,并被报道可以与Geobacter发生直接的种间电子传递[36]。Methanosarcina和Methanosaeta隶属于广古菌门(Euryarchaeota),在接种底泥中这2种菌的相对丰度较小。施加电压刺激并经过24周培养后,这2种菌在底泥中成了显著优势菌,因此,底泥中PCB 61的还原脱氯可能受Methanosarcina和Methanosaeta这2种菌支配。第3种是发酵、产氢相关微生物:Blvii28[37],Kosmotoga[38]、Hydrogenophaga[39]和Syntrophus[40],这类菌可以将底泥中的有机质发酵为小分子有机酸或氢气,可作为在PCB 61脱氯过程中被脱氯菌利用的优质碳源和电子供体。不同微生物驱动PCBs生物还原的效能本身存在差别,不同微生物在相对丰度上的差异与不同电压下PCB 61的还原程度存在对应关系。
在开路条件中,主要优势菌为Syntrophus、Kosmotoga和Syntrophobacter[41],其种类较少,且主要功能与发酵产氢相关,对应的PCBs脱氯速率也较低。在施加不同电压的条件下,PCBs脱氯速率较快,分析其中的优势菌发现:1)电化学活性菌Geobacter在闭路条件下的丰度显著高于初始底泥和开路条件,Ignavibacterium相对于开路的丰度差异更显著,且在初始底泥中相对丰度较低。在电极充当电子供体的生物阴极体系中,这2种电化学活性菌所具备的加速电子传递的性能可强化脱氯微生物驱动的得电子还原过程,实现脱氯性能的提升。2)2种典型的产甲烷脱氯菌Methanosarcina和Methanosaeta占主要优势,其特点是对PCBs具有对位脱氯功能,与本实验的主要脱氯途径一致。
3 结论
1) 在施加负电位的条件下,河涌底泥中PCB 61的微生物还原脱氯效率变大,且电极电位越小越有利于PCB 61的还原脱氯。24周后,施加-0.7 V电位的条件下,PCB 61的还原率达到59.05%,是开路条件的1.83倍。在生物电化学底泥体系下,PCB 61的脱氯途径为对位和间位脱氯,即脱氯产物为PCB 23和PCB 29。
2) 电极在恒电位装置的控制下保持稳定电位(-0.1、-0.3、-0.5和-0.7 V),电极电位所提供的电子可以被电化学活性菌利用,促进电化学活性菌生长代谢,从而强化了微生物对PCB 61的还原脱氯过程。
3) 高通量测序结果表明,恒定电位刺激对底泥中微生物的代谢生长有促进效果,对PCB 61的还原脱氯有促进作用的Geobacter、Ignavibacterium电化学活性菌和Methanosarcina、Methanosaeta产甲烷菌的丰度得到显著提升,而电化学活性菌与产甲烷菌可发生直接的种间电子传递,强化了PCB 61的还原脱氯。
4) 对位脱氯的途径是由体系中Methanosarcina、Methanosaeta产甲烷菌的存在并占主导作用决定的,而体系中还原脱氯过程增强可能是通过电化学活性菌和产甲烷菌的共同作用实现的。
 下载:
下载:
